

Зоологический журнал, 2019, T. 98, № 6, стр. 634-642
Распространение и морфологическая изменчивость в популяциях Coenonympha leander (Lepidoptera, Satyridae) Южного урала и Приуралья
Е. Ю. Захарова *
Институт экологии растений и животных Уральского отделения РАН
620144 Екатеринбург, Россия
* E-mail: zakharova@ipae.uran.ru
Поступила в редакцию 03.04.2018
После доработки 27.07.2018
Принята к публикации 07.09.2018
Аннотация
Изучено современное распространение в Уральском регионе западноевразиатского суббореального вида Coenonympha leander и уточнены границы распространения на северо-восточном крае ареала: 56° с.ш. на севере, 65° в.д. на востоке. В условиях лесостепи C. leander обитает в виде отдельных изолированных локальных популяций, в то время как в степной зоне распространен достаточно равномерно. В степях Южного Урала и Приуралья вид толерантен к умеренной антропогенной трансформации сообществ, не является редким и не может служить видом-индикатором, как в других частях ареала. Проанализирована географическая изменчивость длины крыла, диаметров и частоты встречаемости глазчатых пятен крылового рисунка. Наибольшим морфологическим своеобразием отличаются краевые форпостные популяции, особенно самая северная популяция из окрестностей дер. Нижнее Тукбаево, обитающая в уникальных условиях азональной Месягутовской лесостепи. По направлению с севера на юг существует тенденция к формированию клины: размеры крыльев уменьшаются, диаметры глазчатых пятен и частота их встречаемости увеличиваются.
Coenonympha leander (Esper [1784]) (Lepidoptera, Satyridae) – широко распространенный элемент энтомоценозов открытых луговых и степных пространств. Предковая форма, вероятно, возникла в Западной Палеарктике, затем вид распространился на восток до Зауралья и юга Западной Сибири (Kodandaramaiah, Wahlberg, 2009). Известно, что в европейской части ареала к концу 20–началу 21 века С. leander регистрировали на территории Албании, Болгарии, Македонии, Греции, Румынии, Турции и Югославии (van Swaay, Warren, 1999; van Swaay et al., 2006), а также в ряде областей Кавказского и Волго-Уральского регионов (Каталог чешуекрылых…, 2008; Anikin et al., 2017). Практически повсеместно на европейской части ареала численность популяций сокращается. Так, например, в Саратовской обл. вид является редким, локально встречающимся из-за возрастающей нагрузки в виде выпаса скота, весенних и осенних палов травы (Красная книга Саратовской…, 2006). Кроме того, согласно результатам, полученным на основе моделирования размера потенциального ареала вида по совокупности нескольких климатических факторов, таких как сумма активных температур, содержание воды в верхних слоях почвы, суммарная годовая температура, количество осадков и др., С. leander является видом категории HHHR (extremely high climate change risk) (Settele et al., 2008).
Наиболее восточная часть ареала С. leander соответствует лесостепной и степной зонам Южного Урала и Приуралья. Граница распространения вида на востоке проходит по территории Курганской обл., на севере – по территории Челябинской обл. и республики Башкортостан. Естественные сообщества Южного Урала в значительной степени подвержены антропогенному воздействию, включающему сплошные рубки коренных лесов, лесные и степные пожары, выпас скота, сенокошение, раскорчевку и распашку наиболее плодородных участков под пашни и огороды. В результате хозяйственной деятельности человека во многих районах лесостепной и степной зон Южного Урала естественный ландшафт преобразован в культурно-аграрный (Колесников, 1964; Куликов, 2005). На настоящий момент по данным официальной статистики сельскохозяйственные угодья в республике Башкортостан занимают около 51%, в Челябинской обл. – 58%, в Курганской – 63%, в Оренбургской – 88% (Природа России, 2018).
Изменения климата и трансформация естественных сообществ в агроценозы приводят к смещению границ ареалов. Особенно чувствительными к воздействию антропогенной и климатической нагрузок оказываются популяции на краю распространения, существующие в субоптимальных условиях. Такие периферические популяции, обитающие на границе видовых ареалов, принято рассматривать в качестве естественных форпостных группировок (Васильев, 2012).
Цель данной работы – оценить современное распространение и проанализировать морфологическую изменчивость в популяциях Coenonympha leander на северо-восточном крае ареала.
МАТЕРИАЛ И МЕТОДЫ
Материалом для статьи послужили данные, собранные в различных районах Южного Урала и Приуралья с 2003 г. по настоящее время. Географическое расположение 13 изученных локалитетов приведено на карте-схеме (рис. 1). Объем выборок, географические координаты и зональная приуроченность 13 изученных локалитетов C. leander приведены в табл. 1. Всего проанализировано 879 экз. При статистическом анализе использовали только репрезентативные выборки, поэтому из-за небольшого объема материала не включали самцов C. leander из окр. пос. Сухтелинский. Выборки самок (объемом не менее 8 экз.) анализировали только по частотам встречаемости глазчатых пятен крылового рисунка.
Рис. 1.
Карта-схема района исследования на Южном Урале и в Приуралье. Треугольники – находки Coenonympha leander по данным Горбунова и др., 1992; квадраты – находки C. leander по данным Lukhtanov, Lukhtanov, 1994; кружки с арабскими цифрами – точки сбора материала. 1 – Башкортостан, Дуванский р-н, окр. д. Нижнее Тукбаево, сборы Е.Ю. Захаровой (2012, 2013 гг.). 2 – Башкортостан, Учалинский р-н, хр. Нурали, окр. с. Яльчигулово, сборы Е.Ю. Захаровой, Т.К. Туневой (2008 г.). 3 – Курганская обл., окр. с. Звериноголовское, Прорывинский государственный природный комплексный (ландшафтный) заказник, сборы П.Ю. Горбунова, А.О. Шкурихина, Е.Ю. Захаровой (2013, 2014 гг.). 4 – Курганская обл., Целинный р-н., Кислянский сельсовет, окр. д. Белозерки, сборы Е.Ю. Захаровой, Ю.М. Чибиряка (2015 г.). 5 – Челябинская обл., Троицкий р-н., окр. д. Осиповка, б. р. Уй, сборы Е.Ю. Захаровой, Ю.М. Чибиряка (2014 г.). 6 – Челябинская обл., Верхнеуральск. р-н, окр. пос. Сухтелинский, б.р. Курасан, Шелудивые и Ущельные горы, сборы Е.Ю. Захаровой, Т.С. Ослиной, Ю.М. Чибиряка (2012 г.). 7 – Челябинская обл., Верхнеуральск р-н, окр. пос. Вятское Леоновские горы, сборы Е.Ю. Захаровой, Т.С. Ослиной (2009 г.). 8 – Челябинская обл., Брединский р-н, окр. д. Варшавка, берег р. Карагайлы-Аят, сборы Е.Ю. Захаровой, Т.С. Ослиной (2012 г.). 9 – Челябинская обл., Кизильский р-н, окр. пос. Ждановский, г. Чека, сборы Е.Ю. Захаровой, П.В. Рудоискателя (2010 г.). 10 – Оренбургская обл., Кувандыкский р-н, окр. г. Кувандык, сборы Е.Ю. Захаровой, А.О. Шкурихина, П.В. Рудоискателя (2005, 2007, 2010 гг.). 11 – Оренбургская обл., Кувандыкский р-н, окр. г. Медногорск, сборы Е.Ю. Захаровой, А.О. Шкурихина, (2007 г.). 12 – Оренбургская обл., окр. с. Губерля, сборы П.Ю. Горбунова, П.В. Рудоискателя (2003, 2004 гг.). 13 – Оренбургская обл., Оренбургский государственный заповедник, участок Айтуарская степь, сборы А.О. Шкурихина, Т.С. Ослиной (2013 г.)
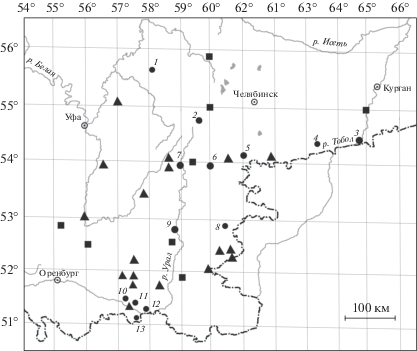
Таблица 1.
Характеристика изученных локалитетов и объем выборок Coenonympha leander с территории Южного Урала и Приуралья
| № | Локалитет | Географические координаты |
Природная зона | Подзона | Антропогенная трансформация | Количество экземпляров C. leander |
||
|---|---|---|---|---|---|---|---|---|
| с.ш. | в.д. | самцы | самки | |||||
| 1 | Нижнее Тукбаево | 55°40′ | 58°09′ | Лесостепь | Северная лесостепь | Умеренная | 33 | 4 |
| 2 | хребет Нурали | 54°44' | 59°40' | Лесостепь | Северная лесостепь | Умеренная | 50 | 50 |
| 3 | Звериноголовское | 54°25′ | 64°45′ | Степь | Разнотравно-дерновинно-злаковые степи | Умеренная | 13 | 15 |
| 4 | Белозерки | 54°21′ | 63°24′ | Степь | Разнотравно-дерновинно-злаковые степи | Умеренная | 18 | 4 |
| 5 | Осиповка | 54°07′ | 61°05′ | Степь | Ковыльно-разнотравные северные степи | Умеренная | 99 | 36 |
| 6 | Сухтелинский | 53°55′ | 60°00′ | Степь | Ковыльно-разнотравные степи | Сильная | 3 | 4 |
| 7 | Леоновские горы | 53°55′ | 59°01′ | Лесостепь | Южная лесостепь | Слабая | 6 | 9 |
| 8 | Варшавка | 52°49′ | 60°28′ | Степь | Ковыльно-разнотравные степи | Умеренная | 14 | 26 |
| 9 | г. Чека | 52°34' | 59°06' | Степь | Ковыльно-разнотравные степи | Умеренная | 60 | 129 |
| 10 | Кувандык | 51°28′ | 57°17′ | Степь | Разнотравно-дерновинно-злаковые степи | Умеренная | 91 | 8 |
| 11 | Медногорск | 51°23′ | 57°35′ | Степь | Разнотравно-дерновинно-злаковые степи | Сильная | 123 | 27 |
| 12 | Губерля | 51°08′ | 57°57′ | Степь | Типчаково-ковыльные степи | Умеренная | 19 | 3 |
| 13 | Айтуарская степь | 51°06′ | 57°41′ | Степь | Типчаково-ковыльные степи | Слабая | 35 | 0 |
При изучении изменчивости крылового рисунка C. leander применяли подход, использованный нами ранее. Глазчатые пятна рассматривали одновременно в качестве как метрических, так и неметрических (фенетических) признаков (Захарова, 2010, 2017; Захарова и др., 2017).
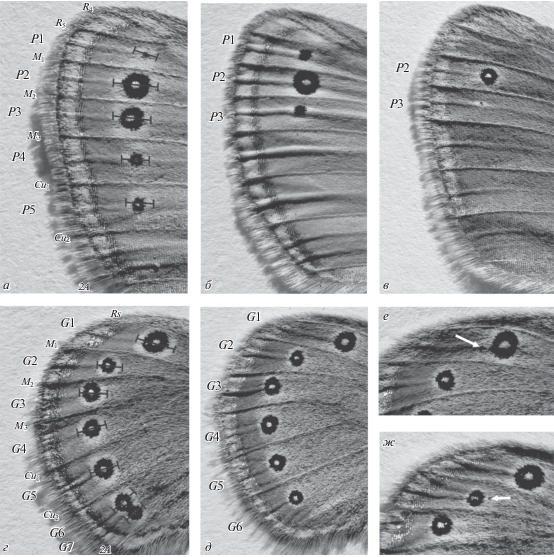
Камеральная обработка материала включала измерения длины крыльев (переднего – от основания жилки Sc до вершины крыла, заднего – от основания жилки Rs до вершины жилки Cu1) и диаметров глазчатых пятен крылового рисунка с испода. Типичное глазчатое пятно представляет собой черный круг с фокусом в центре и окаймляющее его желтое кольцо. Диаметр глазчатого пятна измеряли как максимальный диаметр наружного желтого кольца вдоль срединной линии той ячейки, в которой оно расположено, как показано на рис. 2а, 2г. Пятна рисунка переднего крыла обозначены буквой Р, заднего крыла – буквой G. Все измерения сделаны по левой стороне особи на бинокулярном микроскопе МБС-10 с использованием окулярного микрометра при увеличении 8×0.6.
Рис. 2.
Глазчатые пятна в рисунке переднего (а–в) и заднего (г–ж) крыльев C. leander. Буквами Р обозначены пятна на переднем крыле, буквами G – на заднем. Отрезки иллюстрируют способ измерения диаметров пятен (а, г). Белые стрелки указывают на мелкие дополнительные пятна (е, ж).
Анализ изменчивости комплекса метрических признаков (размеров всех глазчатых пятен и длин крыльев) проводили с использованием дискриминантного анализа. Для наглядности и при сравнении выборок по размерам пятен крылового рисунка использовали параметр “суммарный диаметр пятен на крыле”. Для этого значения диаметров всех присутствующих на крыле пятен у особи суммировали, после чего рассчитывали внутривыборочное среднее и его ошибку. Наличие значимых различий между выборками по суммарным диаметрам пятен оценивали с помощью однофакторного дисперсионного анализа (ANOVA). Данные расчеты выполнены в пакете программ Statistica 6.0.
Максимально у C. leander на переднем крыле может присутствовать 5 пятен (рис. 2а), на заднем – 7 пятен (рис. 2г). Возможны различные варианты редукции числа пятен в крыловом рисунке, например случай отсутствия пятен Р4 и Р5 (рис. 2б) или пятен Р1, Р4 и Р5 (рис. 2в). На заднем крыле наиболее типично расположение ряда из шести пятен G1–G6 (рис. 2д). Помимо основных, в крыловом рисунке иногда присутствуют мелкие дополнительные пятна. Это явление было описано Шванвичем (Schwanwitsch, 1935) и названо “budding” — “почкование”. Некоторые из обнаруженных вариантов проиллюстрированы на рис. 2е, 2ж. В данной статье мы не рассматриваем изменчивость этих элементов крылового рисунка, поскольку у C. leander они крайне редки, и их суммарная частота встречаемости не превышает 0.1%. Статистическую значимость различий между выборками по частотам встречаемости основных глазчатых пятен оценивали с помощью критерия χ2 в программе Past 2.17c (Hammer et al., 2001).
Степень антропогенной нагрузки оценивали по наличию на конкретной территории выпаса, наличию сельскохозяйственных угодий и рекреационной нагрузки. Отнесение географических локалитетов к той или иной природной зоне осуществляли на основе районирования, принятого в следующих источниках: Географический атлас Оренбургской…, 1999; Куликов, 2005; Науменко, 2008; Определитель высших растений…, 1988.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Распространение на Южном Урале и в Приуралье. Самая северная находка вида – окрестности д. Нижнее Тукбаево (55°40′ с.ш., 58°09′ в.д.) в азональной Месягутовской лесостепи. Согласно ботанико-географическому районированию данный локалитет относится к лесостепному району Айско-Юрезанского водораздела Башкирского Предуралья. По характеру поверхности район представляет сложное чередование резко очерченных увалов и мелких хребтов с крутыми в верхней части, внизу пологими растянутыми склонами, переходящими в речные долины (Крашенинников, Кучеровская-Роженец, 1941). Бабочек отлавливали в верховьях р. Кушкаяк, где глубокая узкая долина пересыхающего левого притока разрезает увалы. Здесь основу растительности составляют луговые степи и остепненные луга. В целом данная территория характеризуется высокой комплексностью растительности, представляя фрагмент типичной горной лесостепи. Следует отметить, что специальный поиск C. leander на соседних территориях Месягутовской лесостепи не увенчался успехом. Например, в окрестностях д. Юсупово, удаленной от Нижнего Тукбаево на 15 км, C. leander не найден. На настоящий момент популяцию C. leander из окрестностей д. Нижнее Тукбаево можно считать наиболее северной островной локальной популяцией в Предуралье. В европейской части ареала C. leander также не распространен севернее 56° с.ш. Например, самая северная популяция вида, обнаруженная на территории Нижегородской обл., обитает на участке степного разнотравья в окрестностях г. Княгинино (около 55°50′ с.ш., 45°02′ в.д.) (Корб, 2006).
Помимо Нижнего Тукбаево в лесостепной зоне обнаружены только два локалитета C. leander: хребет Нурали (северная лесостепь) и Леоновские горы (южная лесостепь). Данные популяции также представляют значительный интерес при анализе изменчивости и могут быть рассмотрены в качестве краевых северных популяций на данной части ареала. Подробное геоботаническое описание растительных сообществ этих территорий приведено нами ранее (Захарова и др., 2014).
Самое восточное местообитание C. leander – окрестности с. Звериноголовское (54°25′ с.ш., 64°45′ в.д.) Курганской обл. Вид обнаружен в Прорывинском заказнике, территория которого расположена в пределах современного хорошо развитого пойменного комплекса и первой надпойменной террасы р. Тобол и характеризуется равнинным плоским рельефом. Популяция C. leander обитает на примыкающих к пойме Тобола по краю борового массива открытых участках песчаных степей и остепненных пойменных лугов, где в травостое в массе присутствует ковыль перистый.
На территории Южного Урала и Зауралья, ограниченной с севера 54° с.ш., с юга 51° с.ш., где преобладают ковыльно-разнотравные, разнотравно-дерновинно-злаковые и типчаково-ковыльные степи, C. leander обнаруживали практически во всех обследованных локалитетах. Это свидетельствует о том, что в степной зоне на настоящий момент вид распространен достаточно равномерно и проявляет высокую степень устойчивости к антропогенной нагрузке и трансформации естественных сообществ по сравнению, например, с Coenonympha amaryllis (Stoll 1782). Оценка распространения C. leander в степной зоне Урала и Приуралья позволяет сделать вывод о том, что здесь вид не является редким и не может служить индикатором неповрежденных степных сообществ, как в других частях ареала (Красная книга Саратовской…, 2006; Полтавский, 2017; Poltavsky, 2005).
Изменчивость длины крыла и глазчатых пятен крылового рисунка. Результаты измерений длины крыла и значения суммарных диаметров глазчатых пятен крылового рисунка приведены в табл. 2. В среднем по размеру самцы значимо мельче самок (t = –22.99, df = 1772, p < 0.001). Для уральского региона длина переднего крыла самцов составляет 18.5 ± 0.1 мм, самок – 19.4 ± 0.1 мм, длина заднего крыла самцов 14.4 ± 0.1 мм, самок – 15.3 ± 0.1 мм. Как и у других видов Coenonympha, глазчатые пятна крупнее у самок, чем у самцов (табл. 2). Различия между выборками самцов по признаку “суммарный диаметр пятен рисунка” согласно результатам ANOVA статистически достоверны как по передним крыльям (F = 6.81, df 1 = 13, df 2 = 1113, p < < 0.001), так и по задним (F = 12.83, df 1 = 13, df 2 = = 1101, p < 0.001).
Таблица 2.
Средние значения длины крыльев, суммарных диаметров пятен и частота их встречаемости в крыловом рисунке C. leander
| № | Выборка | Пол | Длина, мм | Суммарный диаметр глазчатых пятен, мм | Частота встречаемости, % | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| переднее крыло | заднее крыло | переднее крыло | заднее крыло | P1 | P2 | P3 | P4 | P5 | G2 | G4 | G7 | |||
| 1 | Нижнее Тукбаево | Самцы | 19.3 ± 0.1 | 15.1 ± 0.1 | 0.8 ± 0.1 | 5.9 ± 0.2 | 0.0 | 100.0 | 31.8 | 0.0 | 0.0 | 100.0 | 100.0 | 9.1 |
| 2 | хребет Нурали | Самцы | 18.1 ± 0.1 | 14.0 ± 0.1 | 0.9 ± 0.1 | 5.6 ± 0.1 | 4.0 | 99.0 | 66.7 | 1.0 | 0.0 | 100.0 | 100.0 | 27.0 |
| Самки | 19.3 ± 0.1 | 15.0 ± 0.1 | 1.3 ± 0.1 | 6.3 ± 0.1 | 8.0 | 98.0 | 85.0 | 24.5 | 0.0 | 100.0 | 100.0 | 53.1 | ||
| 3 | Леоновские горы | Самцы | 18.7 ± 0.3 | 14.5 ± 0.3 | 0.7 ± 0.2 | 5.6 ± 0.4 | 0.0 | 100.0 | 16.7 | 0.0 | 0.0 | 100.0 | 100.0 | 0.0 |
| Самки | 19.2 ± 0.2 | 15.1 ± 0.2 | 1.4 ± 0.2 | 6.3 ± 0.3 | 0.0 | 100.0 | 88.9 | 16.7 | 0.0 | 100.0 | 100.0 | 5.6 | ||
| 4 | Звериноголовское | Самцы | 18.9 ± 0.1 | 14.7 ± 0.1 | 1.0 ± 0.1 | 6.1 ± 0.2 | 7.0 | 100.0 | 56.3 | 0.0 | 0.0 | 98.1 | 100.0 | 28.7 |
| Самки | 19.9 ± 0.1 | 15.7 ± 0.1 | 1.5 ± 0.1 | 6.4 ± 0.2 | 18.7 | 100.0 | 86.4 | 17.0 | 3.5 | 100.0 | 100.0 | 30.0 | ||
| 5 | Белозерки | Самцы | 18.0 ± 0.2 | 13.9 ± 0.2 | 0.9 ± 0.1 | 5.6 ± 0.1 | 5.6 | 100.0 | 47.2 | 0.0 | 0.0 | 100.0 | 100.0 | 22.2 |
| 6 | Осиповка | Самцы | 18.8 ± 0.1 | 14.6 ± 0.1 | 1.1 ± 0.1 | 6.4 ± 0.1 | 5.6 | 100.0 | 69.7 | 5.6 | 1.5 | 100.0 | 100.0 | 50.8 |
| Самки | 19.5 ± 0.1 | 15.4 ± 0.1 | 1.6 ± 0.1 | 6.7 ± 0.2 | 8.3 | 100.0 | 97.2 | 30.6 | 5.6 | 100.0 | 98.6 | 45.8 | ||
| 7 | Варшавка | Самцы | 18.4 ± 0.2 | 14.0 ± 0.2 | 1.0 ± 0.1 | 5.6 ± 0.2 | 2.1 | 100.0 | 84.7 | 4.2 | 0.0 | 100.0 | 100.0 | 40.2 |
| Самки | 19.3 ± 0.2 | 15.1 ± 0.1 | 1.4 ± 0.1 | 6.3 ± 0.2 | 15.7 | 100.0 | 90.2 | 24.9 | 2.4 | 100.0 | 100.0 | 40.7 | ||
| 8 | г. Чека | Самцы | 18.2 ± 0.1 | 14.0 ± 0.1 | 1.0 ± 0.1 | 5.9 ± 0.1 | 4.4 | 99.1 | 59.6 | 4.7 | 0.0 | 100.0 | 99.2 | 44.9 |
| Самки | 19.2 ± 0.1 | 15.1 ± 0.1 | 1.3 ± 0.1 | 6.2 ± 0.1 | 15.8 | 100.0 | 87.2 | 22.0 | 3.6 | 100.0 | 100.0 | 48.9 | ||
| 9 | Кувандык | Самцы | 18.2 ± 0.1 | 14.1 ± 0.1 | 1.3 ± 0.1 | 6.5 ± 0.1 | 1.7 | 98.9 | 52.0 | 5.8 | 0.6 | 98.9 | 100.0 | 12.1 |
| Самки | 19.4 ± 0.3 | 15.2 ± 0.2 | 1.6 ± 0.2 | 6.2 ± 0.4 | 12.5 | 100.0 | 100.0 | 12.5 | 6.3 | 100.0 | 100.0 | 13.4 | ||
| 10 | Медногорск | Самцы | 18.5 ± 0.1 | 14.3 ± 0.1 | 1.0 ± 0.1 | 5.7 ± 0.1 | 5.2 | 98.7 | 58.5 | 4.6 | 0.9 | 99.6 | 100.0 | 14.5 |
| Самки | 19.5 ± 0.1 | 15.4 ± 0.1 | 2.3 ± 0.1 | 8.5 ± 0.2 | 35.2 | 100.0 | 92.5 | 41.5 | 7.5 | 100.0 | 100.0 | 26.0 | ||
| 11 | Губерля | Самцы | 18.1 ± 0.2 | 13.9 ± 0.1 | 1.2 ± 0.1 | 6.1 ± 0.2 | 21.1 | 100.0 | 60.5 | 7.9 | 0.0 | 100.0 | 100.0 | 44.7 |
| 12 | Айтуарская степь | Самцы | 18.6 ± 0.1 | 14.3 ± 0.1 | 1.3 ± 0.1 | 6.5 ± 0.2 | 11.4 | 100.0 | 81.4 | 8.6 | 1.4 | 100.0 | 100.0 | 59.4 |
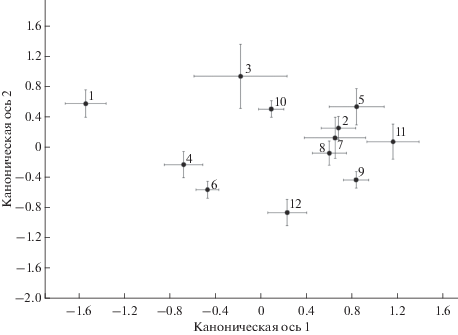
На основе комплекса всех метрических признаков (длины переднего и заднего крыльев, диаметры пятен крылового рисунка) были оценены различия между 12 выборками самцов C. leander с помощью канонического дискриминантного анализа: λ Уилкса = 0.39, F = 6.49, df 1 = 154, df 2 = 8685, p < 0.001. На первую каноническую переменную приходится 46.7% дисперсии, на вторую – 24.3%. Результаты анализа проиллюстрированы на рис. 3. Вдоль первой канонической оси проявились географические различия. Наибольшим морфологическим своеобразием отличаются самцы C. leander из самого северного местообитания (Нижнее Тукбаево): они имеют самые крупные крылья и при этом самые мелкие пятна крылового рисунка переднего крыла (табл. 2). Несколько обособлены в пространстве канонических осей выборки из подзоны разнотравно-дерновинно-злаковых степей (Звериноголовское и Осиповка). Довольно тесную группу образуют центроиды выборок из соседних степных местообитаний, что говорит об их морфологическом сходстве.
Рис. 3.
Результаты канонического дискриминантного анализа географической изменчивости длины крыльев и диаметров глазчатых пятен крылового рисунка самцов C. leander с территории Урала и Приуралья Нумерация выборок соответствует номерам локалитетов в табл. 2 и 3. Приведены центроиды выборок с учетом величин стандартных ошибок.
Следующим этапом работы был анализ изменчивости глазчатых пятен крылового рисунка C. leander в качестве неметрических (фенетических) признаков. Почти всегда (не менее чем в 98% случаев) во всех проанализированных выборках все особи на исподе переднего крыла имеют пятно в ячейке М1–М2, обозначенное Р2. Пятно Р3 было обнаружено у более чем половины всех самцов и самок за исключением двух северных выборок (Нижнее Тукбаево и Леоновские горы) (табл. 2). Следует заметить, что крыловой рисунок с двумя пятнами Р2 и Р3 на переднем крыле (рис. 2в) является самым часто встречающимся у уральских C. leander. Пятна Р1, Р4, Р5 в целом чаще проявляются в выборках из степных локалитетов, чем в выборках из лесостепных и северных разнотравно-дерновинно-злаковых степей (Звериноголовское и Белозерки).
Для C. leander характерна достаточно высокая степень стабильности рисунка заднего крыла. Наиболее часто встречающимися вариантами является крыловой рисунок с 6 пятнами (рис. 2д) либо с 7 пятнами (рис. 2г). Частота встречаемости пятен G2, G4 и G7 приведена в табл. 2. Остальные пятна рисунка заднего крыла были обнаружены у всех исследованных самцов и самок. Направление географической изменчивости пятен рисунка на заднем крыле имеет тот же характер, что и на переднем. Частота встречаемости фена G7 несколько больше в южных степных локалитетах, чем в северных лесостепных.
Результаты проверки статистической значимости наблюдаемых различий по частотам встречаемости всех пятен рисунка переднего и заднего крыльев методом χ2 приведены в табл. 3. Самцы C. leander из Нижнего Тукбаево статистически значимо отличаются от самцов из всех остальных локалитетов. То же характерно и для выборки самцов с Леоновских гор. Остальные выборки оказываются в той или иной степени сходны друг с другом. Например, между выборками из соседних локалитетов ковыльно-разнотравных степей (г. Чека и Варшавка) не обнаружены различия ни по комплексу метрических признаков (рис. 3), ни по частотам встречаемости фенов (табл. 3).
Таблица 3.
Значения χ2 для выборок самцов (верхняя треугольная матрица) и самок (нижняя треугольная матрица) C. leander по частотам встречаемости фенов глазчатых пятен крылового рисунка
| № | Выборка | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | Н. Тукбаево | – | 22.0 | 12.9 | 20.8 | 12.5 | 45.5 | 39.9 | 34.8 | 12.7 | 17.6 | 71.0 | 63.2 |
| 2 | хр. Нурали | – | 50.5 | 2.7 | 4.2 | 10.4 | 5.5 | 6.9 | 11.0 | 7.3 | 19.6 | 18.5 | |
| 3 | Леоновские горы | 41.8 | – | 47.1 | 35.5 | 74.6 | 71.3 | 62.7 | 33.1 | 40.6 | 76.9 | 92.7 | |
| 4 | Звериноголовское | 15.3 | 35.1 | – | 1.3 | 12.1 | 12.3 | 8.3 | 15.7 | 10.3 | 15.8 | 19.0 | |
| 5 | Белозерки | 8.6 | 28.5 | 26.1 | – | 17.2 | 16.6 | 12.8 | 11.7 | 8.3 | 20.5 | 26.6 | |
| 6 | Осиповка | 7.0 | 39.1 | 10.5 | 13.7 | – | 6.0 | 2.2 | 22.3 | 17.3 | 11.9 | 3.0 | |
| 7 | Варшавка | 6.7 | 38.9 | 3.0 | 17.6 | 4.4 | – | 5.0 | 17.6 | 15.3 | 21.0 | 11.7 | |
| 8 | г. Чека | 6.4 | 46.4 | 4.5 | 18.7 | 4.6 | 1.2 | – | 18.1 | 15.0 | 11.1 | 7.4 | |
| 9 | Кувандык | 35.1 | 20.9 | 9.8 | 41.1 | 23.0 | 18.9 | 22.9 | – | 2.3 | 30.1 | 32.8 | |
| 10 | Медногорск | 36.7 | 51.9 | 13.4 | 46.0 | 23.8 | 16.1 | 20.9 | 24.9 | – | 22.6 | 25.1 | |
| 11 | Губерля | – | 8.4 | ||||||||||
| 12 | Айтуарская степь | – |
Таким образом, наибольшим морфологическим своеобразием обладают самые крайние популяции, которые могут рассматриваться в качестве форпостных: наиболее северные (Нижнее Тукбаево и Леоновские горы) и самая восточная (Звериноголовское).
ЗАКЛЮЧЕНИЕ
Уточнены современные границы распространения C. leander на северо-восточном крае ареала: 56° с.ш. на севере, 65° в.д. на востоке. В условиях лесостепи C. leander обитает в виде отдельных изолированных друг от друга расстоянием локальных популяций, в то время как в степной зоне вид распространен достаточно равномерно. Анализ морфологической изменчивости по комплексу метрических (длина крыла и диаметры пятен крылового рисунка) и фенетических (стабильность присутствия пятен в крыловом рисунке) признаков позволил следующим образом охарактеризовать географическую изменчивость на данной части ареала. Наибольшим морфологическим своеобразием отличаются краевые форпостные популяции. Самая северная популяция из окрестностей Нижнее Тукбаево, обитающая в уникальных условиях азональной Месягутовской лесостепи, обладает особым набором признаков: крупные крылья, редукция рисунка на переднем крыле (мелкие пятна Р2, Р3, отсутствие пятен Р1, Р4, Р5), мелкие пятна в рисунке заднего крыла. По направлению с севера на юг существует тенденция к формированию клины: размеры крыльев уменьшаются, диаметры глазчатых пятен и частота их встречаемости увеличиваются.
На настоящий момент состояние популяций C. leander в степной зоне Уральского региона не вызывает опасений, и вид не является редким в отличие от основной части ареала. Однако особого внимания заслуживают самые северные лесостепные и восточные степные популяции, которые можно рассматривать в качестве форпостных, поскольку каждая из них представляет собой эволюционно-экологический феномен – элементарный уровень эволюционно-экологической иерархии вида, на котором происходят микроэволюционные события (Васильев, 2012; Шварц, 1980).
Список литературы
Васильев А.Г., 2012. Проблема устойчивости форпостных популяций и сообществ: от теории к методам оценки // Биологические системы: устойчивость, принципы и механизмы функционирования. Ч. 1. Материалы IV Всерос. науч.-практ. конф. с международн. участием. Нижний Тагил. С. 76–80.
Географический атлас Оренбургской области, 1999. Ред. Чибилев А.А. М.: Изд-во ДИК. 96 с.
Горбунов П.Ю., Ольшванг В.Н., Лагунов А.В., Мигранов М.Г., Габидуллин А.Ш., 1992. Дневные бабочки Южного Урала. Екатеринбург. 132 с.
Захарова Е.Ю., 2010. Сезонная изменчивость длины крыла и глазчатых пятен в природных популяциях Erebia ligea L. (Lepidoptera, Satyridae) на Среднем Урале // Энтомологическое обозрение. Т. 89. № 2. С. 320–332.
Захарова Е.Ю., 2017. Фенотипическая изменчивость сенницы Coenonympha hero (L.) (Lepidoptera, Satyridae) в центральной части ареала // Энтомологическое обозрение. Т. 96. № 2. С. 272–285.
Захарова Е.Ю., Шкурихин А.О., Ослина Т.С., 2017. Морфологическая изменчивость Melanargia russiae (Esper, 1783) (Lepidoptera, Satyridae) из основной части ареала и при его расширении на север в условиях меняющегося климата // Сибирский экологический журнал. № 5. С. 567–582.
Захарова Е.Ю., Золотарева Н.В., Чибиряк М.В., 2014. Распространение и ландшафтно-биотопическая приуроченность локальных популяций сенницы Coenonympha amaryllis (Stoll, 1782) (Lepidoptera: Satyridae) на северо-западной границе ареала // Бюллетень Московского общества испытателей природы. Отд. Биол. Т. 119. № 5. С. 20–27.
Каталог чешуекрылых (Lepidoptera) России, 2008. Ред. Синев С.Ю. СПб., М.: Товарищество научных изданий КМК. 424 с.
Колесников Б.П., 1964. Растительность Челябинской области // Природа Челябинской области. Челябинск: Юж.-Урал. кн. изд-во. С. 135–158.
Корб С.К., 2006. Дневные бабочки (Lepidoptera: Rhopalocera) Нижегородской области // Бюллетень Московского общества испытателей природы. Отд. Биол. Т. 111. №. 4. С. 8–14.
Красная книга Саратовской области: Грибы. Лишайники. Растения. Животные, 2006. Саратов: Изд-во Торгово-промышленной палаты. Саратов. обл. 528 с.
Крашенинников И.М., Кучеровская-Роженец С.Е., 1941. Природные ресурсы Башкирской АССР. Т. 1. Растительность Башкирской АССР. М., Л.: Изд-во Академии наук СССР. 162 с.
Куликов П. В., 2005. Конспект флоры Челябинской области (сосудистые растения). Екатеринбург; Миасс: Геотур. 537 с.
Науменко Н.И., 2008. Флора и растительность Южного Зауралья. Курган: Изд-во Курган. гос. ун-та. 512 с.
Определитель высших растений Башкирской АССР, 1988. Алексеев Ю.Е., Алексеев Е.Б., Габбасов К.К. и др. М.: Наука. 316 с.
Полтавский А.Н., 2017. Редкие чешуекрылые (Lepidoptera) Ростовской области // Труды Русского энтомологического общества. С.-Петербург. Т. 88. № 1. С. 94–102.
Природа России. Национальный портал. Уральский федеральный округ [Электронный ресурс]. 2018. Режим доступа: http://www.priroda.ru/regions/earth/index.php?SECTION_ID=&FO_
ID=558. Дата обновления 15.03.2018.
Шварц С.С., 1980. Экологические закономерности эволюции. М.: Наука. 277 с.
Anikin V.V., Sachkov S.A., Zolotuhin V.V., 2017. “Fauna lepidopterologica Volgo-Uralensis”: from P. Pallas to present days // Proceedings of the Museum Witt Munich. 7. Munich, Vilnius. 696 p.
Hammer O., Harper D.A.T., Ryan P.D., 2001. PAST: Paleontological Statistics software package for education and data analysis // Palaeontologia Electronica. V. 4. № 1. 9 p.
Kodandaramaiah U., Wahlberg N., 2009. Phylogeny and biogeography of Coenonympha butterflies (Nymphalidae: Satyrinae) – patterns of colonization in the Holarctic // Systematic Entomology. № 34. P. 315–323.
Lukhtanov V., Lukhtanov A., 1994. Die Tagfalter Nordwestasiens. Herbipolina. Markleuthen. 440 s.
Poltavsky A.N., 2005. Concept for the preservation of the Lepidoptera biodiversity in agrolandscapes // Phegea. V. 33. № 4. P. 145–150.
Schwanwitsch B.N., 1935. Evolution of the wing-pattern in palaearctic Satyridae. III. Pararge and five other genera // Aca Zoologica. Bd. XVI. P.145–270.
Settele J., Kudrna O., Harpke A., Kühn I., van Swaay C., et al., 2008. Climatic Risk Atlas of European Butterflies. BioRisk. № 1. Pensoft. 712 p.
Van Swaay C.A.M., Warren, M.S., 1999. Red Data book of European butterflies (Rhopalocera). Nature and Environment, № 99. Council of Europe Publishing, Strasbourg. 260 p.
Van Swaay C.A.M., Warren, M.S., Loïs G., 2006. Biotope use and trends of European butterflies // Journal of Insect Conservation. № 10. P. 189–209.
Дополнительные материалы отсутствуют.
Инструменты
Зоологический журнал