

Зоологический журнал, 2019, T. 98, № 7, стр. 792-803
Склериты и мускулатура гениталий самцов семейства Platypezidae (Diptera)
Т. В. Галинская 1, 2, *, А. И. Шаталкин 3, **, О. Г. Овчинникова 4, ***
1 Биологический факультет Московского государственного университета
им. М.В. Ломоносова
119234 Москва, Россия
2 Научно-методический отдел энтомологии Всероссийского центра карантина растений
140150 Московская область, Раменский район, п. Быково, Россия
3 Научно-исследовательский Зоологический музей Московского государственного университета
им. М.В. Ломоносова
125009 Москва, Россия
4 Зоологический институт РАН
199034 Санкт-Петербург, Россия
* E-mail: nuha1313@gmail.com
** E-mail: shatalkin@zmmu.msu.ru
*** E-mail: brach@zin.ru
Поступила в редакцию 15.09.2018
После доработки 22.11.2018
Принята к публикации 22.11.2018
Аннотация
Впервые приведено описание мускулатуры гениталий самцов Platypezidae (Diptera). Показано, что гипандрий представителей подсемейств Platypezinae и Callomyiinae принадлежит к двум разным структурным типам. У Platypezinae гипандрий принадлежит к исходному структурному типу, показывающему большое сходство с гениталиями прямошовных мух (Brachycera Orthorrhapha), у Callomyiinae гипандрий принадлежит к производному структурному типу, характерному для большинства круглошовных мух (Cyclorrhapha). Первый тип гипандрия представляет по форме желоб и является переходным состоянием к развитию трубчатого гипандрия. Образование трубчатого гипандрия привело к резкому укорочению фаллуса. Ориентация его полового отверстия меняется с дорсальной в первом подсемействе Platypezinae на вентральную во втором подсемействе Callomyiinae. Прегенитальные склериты, а также мускулатура гипандриального комплекса Platypezinae демонстрируют более продвинутое состояние признаков. Приведены дополнительные свидетельства в пользу гипотезы о вхождении гонопод в структуру гипандрия у круглошовных мух. Наличие собственных мышц у прегонитов дает основание утверждать, что эти склериты в структуре гениталий грибных мух не являются выростами гипандрия, но представляют собой производные гонопод.
Статья продолжает серию публикаций, посвященных строению склеритов и мускулатуры абдоминальных сегментов и терминалий самцов двукрылых (Diptera). Ранее изучение мускулатуры позволило уточнить функции и гомологии прегенитальных и генитальных склеритов (Овчинникова, 1989, 1993; Ovtshinnikova, Yeates, 1998; Ovtshinnikova, Galinskaya, 2017; Galinskaya et al., 2018).
Platypezidae Fallén 1815 (грибные мухи) – мелкие, реже среднего размера мухи (1.4–10.0 мм в длину), встречающиеся на грибах, валяющихся на земле ветках, на листьях кустарников и деревьев. Окраска тела черная или серая, часто с добавлением желтых, красных, оранжевых и серебристых участков. Усики с концевой трехчлениковой аристой. Семейство включает около 250 видов из примерно 20 родов в зависимости от принятой классификации (Chandler 2001). Распространено всесветно, кроме Антарктиды. Для представителей подсемейств Callomyiinae и Platypezinae характерен ярко выраженный половой диморфизм в окраске тела, хетотаксии и других признаках.
Брюшко самца состоит из 6 сегментов, 7–8-й сегменты являются прегенитальными и сильно видоизменены в связи с поворотом гениталий самца на 360° по часовой стрелке. Сегменты 9–11 являются генитальными. Их основу составляют видоизмененные склериты девятого сегмента брюшка – гипандрий и эпандрий, отвечающие стерниту и тергиту соответственно (Crampton, 1923). Гоноподы в качестве независимых морфологических элементов генитального аппарата у прямошовных короткоусых (Brachycera Orthorrhapha) отсутствуют у самцов круглошовных мух, но гипандрий и эпандрий несут лопасти, которые в разное время соотносились с гоноподами (McAlpine, 1981). В этом случае предполагались два основных варианта: гоноподы слились либо с гипандрием, либо с эпандрием. Возможен третий сценарий, связанный с полной редукцией гонопод. Кроме того, Чилкотт (Chillcott, 1958) предположил, что стернит 9 полностью редуцирован и замещен слившимися своими вентральными краями гоноподами. С этим связана путаница и несогласованность в использовании терминов, обозначающих гипандриальные и эпандриальные лопасти у Platypezidae. В частности, лопасти гипандрия могут трактоваться как производные гонопод (гонококситов и гоностилей), а дорсальные по расположению склериты – как производные склеротизованных обкладок фаллуса (парамеры авторов – см. McAlpine J.F., 1981); либо обе или одна пара гипандриальных лопастей могут быть новообразованиями, т.е. могут не иметь никакого отношения ни к гоноподам, ни к парамерам. Для практической систематики такая разноголосица создает большие трудности. Поэтому мы следуем предложению Эмдена и Хеннига (Emden, Hennig, 1956, p. 120): “Пока интерпретация остается неопределенной, нейтральные имена подобные терминам “прегонит” и “постгонит”, введенные Крэмптоном, будут предпочтительными для тех, кто считает, что… [их использование] может воспрепятствовать постоянно возникающей путанице”. Крэмптон (Crampton, 1942, 1944) предложил использовать эти термины взамен широко принятых в его время терминов “передние и задние гонапофизы”. В определителях и при описании новых видов термины прегониты и потсгониты для вентрально и дорсально расположенных гипандриальных лопастей используются достаточно часто. Они также приняты и в настоящей работе. Однако мы предполагаем, что эти лопасти возникали параллельно в разных группах круглошовных (Galinskaya, Ovtshinnikova, 2015, 2015а; Ovtshinnikova, Galinskaya, 2016, 2016a), и могут быть не гомологичны, например, лопастям калиптратных двукрылых (Diptera, Calyptratae).
Семейство Platypezidae относится к надсемейству Platypezoidea, которое считается базальным среди Brachycera Cyclorrhapha (McAlpine, 1989; Cumming et al., 1995; Sinclair et al., 2013). Мускулатура гениталий самцов не была изучена ни у одного представителя надсемейства Platypezoidea. Этот факт делает семейство Platypezidae крайне важным для исследования.
В данной работе представлены результаты изучения склеритов и мускулатуры абдоминальных сегментов и терминалий самцов двух представителей семейства Platypezidae: Platypeza consobrina Zetterstedt 1844 из подсемейства Platypezinae и Callomyia amoena Meigen 1824 из подсемейства Callomyiinae.
МАТЕРИАЛ И МЕТОДИКА
Склериты гениталий самцов были изучены следующим образом. Сухой материал был размочен, прегенитальная и генитальная часть брюшка была отделена, выдержана в 10%-ном растворе KOH, затем отпрепарирована. Для обозначения склеритов принята терминология по Каммингу и Вуду (Cumming, Wood, 2009).
Абдоминальные мышцы у изученных нами экземпляров частично мацерированы. Свежий спиртовой материал Platypezidae очень редок, и мы были ограничены доступным нам материалом. Поэтому в дальнейшем тексте фраза “мышцы не обнаружены” может означать как отсутствие этих мышц, так и их утрату при фиксации и хранении.
Мускулатура гениталий самцов была изучена с помощью ручной препаровки насекомых, хранящихся в 70%-ном этаноле, микроножами под стереомикроскопом Leica MZ95. Иллюстрации подготовлены в программе Photoshop CS6 после обработки фотографий, сделанных камерой Canon 5D Mark II на тринокуляре Leica MZ95. Мышцы гениталий самцов были разделены на следующие группы: абдоминальные мышцы, прегенитальные мышцы, тергостернальные мышцы, мышцы гипандриального комплекса, мышцы эпандриального комплекса. Мышцы сгруппированы по месту прикрепления их передней части и описаны под номерами согласно классификации Овчинниковой (1989, 2000).
Буквенные обозначения на рисунках: аэ – аподема эякулятора, гип – гипандрий, гонк – гонококсит, гонс – гоностиль, гипдгон – гоноподальная часть гипандрия, гипдст – стернальная часть гипандрия, дм – дорсальный мост, оцм – околоцерочная мембрана, пост – постгониты, пре – прегониты, ст – стернит, суб – субэпандриальный склерит, сур– сурстили, тг – тергит, ф – фаллус, фп – фаллаподема, ц – церки, эп – эпандрий, ISM1–ISM7 – брюшные межсегментные стернальные мышцы, М1–М43 – прегенитальные и генитальные мышцы, ITM1–ITM7 – брюшные межсегментные тергальные мышцы, TSM1–TSM7 – брюшные тергостернальные мышцы.
РЕЗУЛЬТАТЫ
Подсемейство Callomyiinae
Callomyia amoena Meigen 1824
Сухой материал. 1 ♂, Моск. обл., Голицыно, А. Шаталкин, 20.VIII.1978.
Спиртовой материал. 1 ♂, РОССИЯ, Московская обл., г. Наро-Фоминск, (55.39453° с.ш., 36.7152° в.д.), 01.IX.2009, Д. Гаврюшин (ЗММУ).
Склериты гениталий самцов
Брюшко (рис. 1). Тергиты и стерниты коричневые, слегка блестящие. Тергит 1 шире своей длины. Тергиты 2–5 округлые. Шестой тергит крупный, широкий. Латеральные мембраны желто-коричневые. Стернит 1 шире своей длины. Стерниты 2–6 узкие, коричневые.
Рис. 1.
Детали строения самца Callomyia amoena: A – брюшные стерниты и мускулатура, вид изнутри и B – брюшные тергиты и мускулатура, вид изнутри.
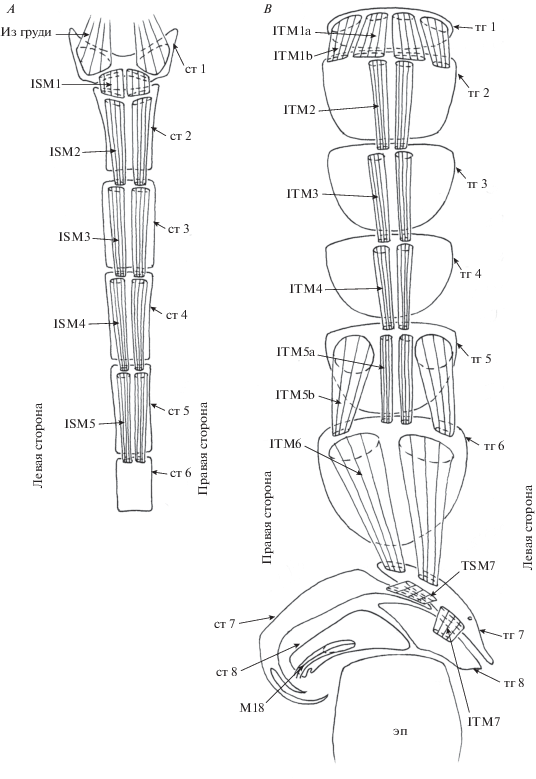
Прегенитальные сегменты 7 и 8 несимметричные. Тергит 7 узкий, длинный, расположен на дорсальной стороне брюшка, слева. Стернит 7 узкий, длинный, расположен на дорсальной стороне брюшка справа, имеет два длинных выроста, заходящих на вентральную сторону тела. Тергит 8 узкий, короткий, расположен на дорсальной стороне брюшка, слева. Стернит 8 узкий, короткий, расположен на дорсальной стороне брюшка справа.
Гениталии самца (рис. 2) симметричные. Эпандрий небольшой, полушаровидный. Короткие сильно склеротизованные двухлопастные сурстили соединяются с эпандрием дорсально и с субэпандриальным склеритом вентрально. Субэпандриальный склерит широкий, хорошо развитый. Церки хорошо развиты, округлые, покрыты короткими волосками. Околоцерочная мембрана, называемая обычно у платипезид гипопроктом (Chandler, Shatalkin, 1998; Cumming, Wood, 2009), расположена между сурстилями, вентральнее церков, и несет две склеротизированные пластинки у края субэпандриального склерита.
Рис. 2.
Детали строения самца C. amoena: A – генитальные и прегенитальные сегменты, вид справа; B – гипандрий и эпандрий, вид изнутри; C – гипандрий, дорсальный вид; D – гипандрий, вид справа; E – гипандрий, вид справа с мышцами; F – эпандрий, вентральный вид и G – эпандрий, дорсальный вид.
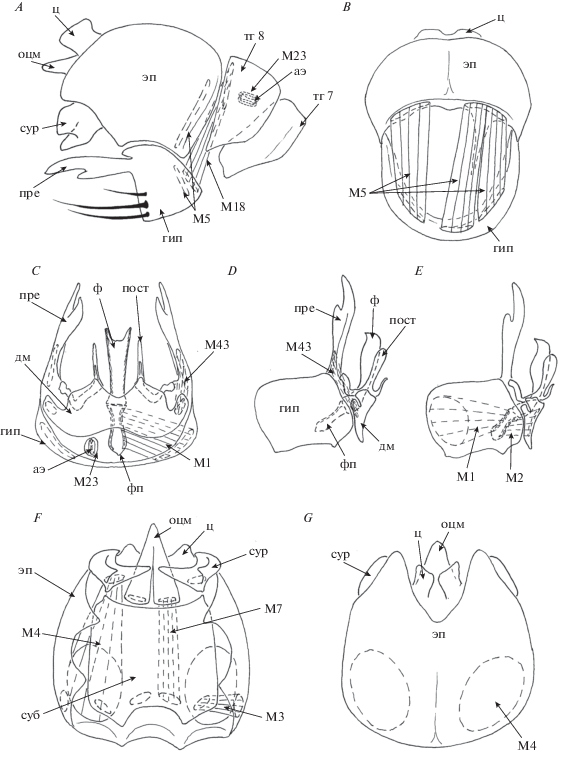
Гипандрий широкий, хорошо развитый, вентрально по заднему краю несет ряд толстых длинных черных щетинок. Гипандрий латерокаудально соединен с прегонитами. Прегониты апикально раздвоены, дорсально соединены с постгонитами. Постгониты дорсально соединены с дорсальным мостом. Фаллус короткий, имеет два продольных склеротизированных тяжа, отличается вентральной ориентацией генитального отверстия (фаллотремы). Фаллаподема хорошо развита. Аподема эякулятора хорошо развита и расположена внутри сегмента 8.
Мускулатура гениталий самцов
Грудные мышцы. К латеральному краю 1-го стернита прикрепляются парные мышцы, берущие начало в груди.
Мускулатура 1–7 сегментов. От заднего края 1-го стернита к переднему краю 2-го стернита идет пара широких коротких плоских мышц ISM1. От переднего края 1-го тергита к переднему краю 2-го тергита идут две пары широких коротких плоских мышц ITM1.
От переднего края 2-го стернита к переднему краю 3-го стернита идет пара длинных мышц ISM2. Набор и места прикреплений стернальных мышц сегментов 3–5 сходны с таковыми сегмента 2. Мышцы, идущие от 6-го стернита, не обнаружены.
От центральной части переднего края 2-го тергита к центральной части переднего края 3-го тергита идет пара длинных мышц ITM2. Набор и места прикреплений тергельных мышц сегментов 3–4 сходны с таковыми сегмента 2. От центральной части переднего края 5-го тергита к центральной части переднего края 6-го тергита идет пара длинных мышц ITM5a. От латеральной части 5-го тергита к латеральной части переднего края 6-го тергита идет пара длинных мошщных мышц ITM5b. От переднего края 6-го тергита к правой части переднего края 7-го тергита идет пара длинных мышц ITM6.
Тергостернальные мышцы TSM в сегментах 1–6 не были обнаружены.
От правой части заднего края 7-го тергита к левой части 7-го стернита идет непарная мышца TSM7. От левой части 7-го тергита к 8-му тергиту идет непарная мышца ITM7.
Прегенитальные мышцы. Длинная спирально закрученная непарная мышца М18 идет от правой части 8-го стернита к гипандрию. Мышца M19 не обнаружена.
Длинные парные асимметричные тергостернальные мышцы M5 идут от латеро-базального края эпандрия к латеро-базальному краю гипандрия; левая мышца шире правой.
Мышцы гипандриального комплекса симметричны. Короткие широкие парные мышцы M1 соединяют латеральные части гипандрия и латеровентральную часть фаллаподемы. Короткие парные мышцы M2 соединяют латеральные части дорсального моста и латеродорсальную часть фаллаподемы. Короткие мощные парные мышцы М43 соединяют базальную часть прегонитов и латеральные выросты дорсального моста. Мышцы М23 окружают аподему эякулятора.
Мышцы эпандриального комплекса симметричны. Длинные плоские парные мышцы M7 идут от переднего края субэпандриального склерита к базальным лопастям околоцерочной мембраны. Парные короткие тонкие мышцы M3 идут от передних латеральных углов эпандрия к латеральным частям переднего края субэпандриального склерита. Длинные мощные парные мышцы M4 идут от переднего края эпандрия к сурстилям. Мышцы М25 не обнаружены.
Подсемейство Platypezinae
Platypeza consobrina Zetterstedt 1844
Сухой материал. 1 ♂, РОССИЯ, Краснодарский край, окрестности г. Адлер, 43.4766° N, 39.9067° E, 28.X.2009, Д. Гаврюшин.
Спиртовой материал. 3 ♂♂, РОССИЯ, Московская обл., г. Наро-Фоминск, (55.39453° с.ш., 36.7152° в.д.), 03.IX.2009, Д. Гаврюшин (ЗММУ).
Склериты гениталий самцов
Брюшко (рис. 3). Тергиты и стерниты коричневые, слегка блестящие. Тергит 1 шире своей длины. Тергиты 2–5 округлые. Шестой тергит крупный, широкий. Латеральные мембраны желто-коричневые. Стернит 1 шире своей длины. Стерниты 2–5 округлые, коричневые. Стернит 6 шире своей длины.
Рис. 3.
Детали строения самца Platypeza consobrina : A – брюшные стерниты и мускулатура, вид изнутри; и B – брюшные тергиты и мускулатура, вид изнутри.
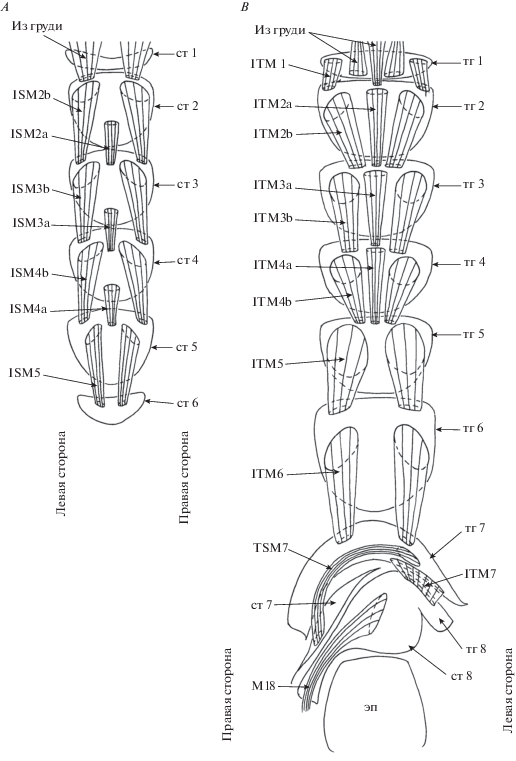
Сегменты 7 и 8 несимметричные. Тергит 7 узкий, длинный, расположен на дорсальной стороне брюшка. Стернит 7 узкий, расположен на дорсальной стороне брюшка справа. Тергит 8 узкий, короткий, расположен на дорсальной стороне брюшка, слева, частично слит с 8 стернитом. Синтергостернит 8: стернит 8 длинный, расположен на дорсальной стороне брюшка справа; стернит 8 продолжен с правой стороны в длинный отросток, заходящий на вентральную сторону и причленяющийся к гипандрию.
Гениталии самца (рис. 4) симметричные. Эпандрий небольшой, полушаровидный. Короткие сильно склеротизованные сурстили соединяются с эпандрием дорсально и с субэпандриальным склеритом вентрально. Субэпандриальный склерит широкий, хорошо развитый, имеет выросты, заходящие в сурстили. Церки хорошо развиты, округлые, покрыты короткими волосками. Околоцерочная мембрана расположена между сурстилями, вентральнее церков.
Рис. 4.
Детали строения самца P. consobrina : A – генитальные и прегенитальные сегменты, дорсальный вид; B – гипандрий и эпандрий, вид изнутри; C – гипандрий, латеральный вид, в разрезе; D – гипандрий, дорсальный вид; E – эпандрий, вентральный вид и F – эпандрий, дорсальный вид.
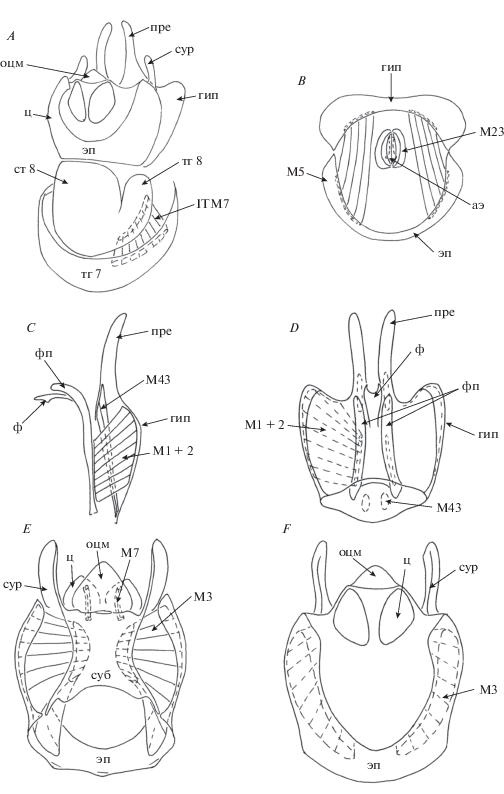
Гипандрий широкий, хорошо развитый, не несет толстых длинных щетинок. Гипандрий каудально с одной парой лопастей, которые мы определяем как прегониты, исходя из следующих соображений. Гипандрий C. amoena, как было рассмотрено выше, несет две пары лопастей, из которых одна пара (прегониты) имеет собственные мышцы, тогда как другая (постгониты) их не имеет. Лопасти гипандрия P. consorbina, как будет показано при обсуждении мускулатуры, имеют собственные мышцы. Поэтому предположительно эти лопасти можно соотнести с прегонитами C. amoena. Фаллус относительно короткий, сильно склеротизованный, саблевидный и имеет дорсальную ориентацию генитального отверстия (фаллотремы). Фаллаподема хорошо развита. Аподема эякулятора небольшая, расположена внутри сегмента 8.
Мускулатура гениталий самцов
Грудные мышцы. К антеролатеральному краю 2-го стернита прикрепляются парные мышцы, берущие начало в груди. К центральной части заднего края 1 тергита прикрепляются парные мышцы, берущие начало в груди. К центральной части переднего края 2 тергита прикрепляется непарная мышца, берущая начало в груди.
Мускулатура 1–7 сегментов. Мышц 1 стернита не обнаружено. От центральной части 2-го стернита к переднему краю 3-го стернита идет длинная непарная мышца ISM2a. От переднего антеролатерального края 2-го стернита к переднему краю 3-го стернита идет пара длинных мышц ISM2b. Набор и места прикреплений стернальных мышц сегментов 3–5 сходны с таковыми сегмента 2. Мышцы, идущие от 6-го стернита, не обнаружены.
От латеральных частей переднего края 1-го тергита к латеральным частям переднего края 2-го тергита идет пара широких коротких плоских мышц ITM1. От центральной части переднего края 2-го тергита к центральной части переднего края 3-го тергита идет длинная мышца ITM2a. От латеральной части 2-го тергита к латеральной части переднего края 3-го тергита идет пара длинных мощных мышц ITM2b. Набор и места прикреплений тергальных мышц сегментов 3–4 сходны с таковыми сегмента 2. От латеральных частей переднего края 5-го тергита к латеральным частям переднего края 6-го тергита идет пара широких мышц ITM5. Подобные мышцы (ITM6) соединятю 6 и 7 тергиты.
Тергостернальные мышцы TSM в сегментах 1–6 не были обнаружены.
Широкая непарная мышца ITM7 идет от левого заднего края 7-го тергита к левому переднему краю 8-го синтергостернита. Длинная непарная мышца TSM7 идет от правой части 7-го тергита к левой части 7-го стернита.
Прегенитальные мышцы. Длинная спирально закрученная непарная мышца М18 идет от правой части 8-го синтергостернита к гипандрию. Мышца M19 не обнаружена.
Длинные парные симметричные тергостернальные мышцы M5 идут от латеро-базального края эпандрия к латеро-базальному краю гипандрия; левая мышца шире правой.
Мышцы гипандриального комплекса симметричны. Короткие широкие парные мышцы M1 + 2 соединяют всю латеральную поверхность гипандрия и латеральную часть фаллаподемы. Короткие парные мышцы M43 соединяют вентральную медиальную часть гипандрия и его лопасти (прегониты). Топографически эти мышцы соответствуют флексорам гипандрия M33, которые во многих группах прямошовных короткоусых (Brachycera Orthorrhapha), например, у Choerades amurensis Hermann (Asilidae) связывают середину гипандрия с базальным краем гонококситов (базистилей) (Овчинникова, 1989, с. 53). Эти данные подкрепляют ранее выдвинутое на обсуждение предположение о соответствии гипандриальных мышц M33 мышцам, связывающим прегониты с гипандрием, изученным в других группах круглошовных мух (Овчинникова, 1993; Ovtshinnikova, 1994). С учетом сказанного можно предположить, что прегониты P. consorbina являются производными гонопод, вошедших в структуру гипандрия в результате их слияния со стернитом 9. Соответственно прегониты C. amoena также следует рассматривать в качестве производных гонопод. Мышцы М23 окружают аподему эякулятора.
Мышцы эпандриального комплекса симметричны. Узкие парные мышцы M7 идут от заднего края субэпандриального склерита к базальным частям церков. Этот факт доказывает, что у вида Callomyia amoena мембранозная лопасть у церков не является гипопроктом (10 стернитом), а основанием церок; у прямошовных двукрылых (Diptera Orthorrhapha) мышцы идут именно так: от 9 тергита к 10 стерниту (субэпандриальному склериту), от 10 стернита к церкам. Длинные мощные парные мышцы M3 широко начинаются на передних латеральных частях эпандрия и идут к субэпандриальному склериту, занимая практически всю внутреннюю поверхность эпандрия. Мышцы М25 не обнаружены.
ОБСУЖДЕНИЕ
Изученные нами два вида Platypezidae сильно различаются между собой по строению как склеритов, так и мышц.
В частности, у C. amoena 8-й сегмент представлен отдельно лежащими тергитом и стернитом, а у P. consorbina имеется 8-й синтергостернит с заметной границей слияния тергита и стернита. При этом набор мышц прегенитальных сегментов у изученных видов идентичен и подтверждает природу склеритов 7-го и 8-го сегментов. У C. amoena имеется асимметрия мышц М5, а у P. consorbina мышцы М5 полностью симметричные.
Строение склеритов и мышц гипандриального комплекса у C. amoena сложнее, чем у P. consorbina. Прегониты у C. amoena расположены латерально и имеют место сочленения, в то время как у P. consorbina они расположены медиально и не имеют мест сочленения. У C. amoena имеются отдельно лежащие склериты, постгониты, не имеющие собственной мускулатуры, в то время как у P. consorbina таких склеритов нет. У C. amoena имеется отдельный склерит – дорсальный мост, служащий местом прикрепления мышц М43 и М2; у P. consorbina дорсальный мост отсутствует, а гипандрий изнутри закрыт мембраной, к которой не подходят мышцы; в то же время мышцы М43 связывают лопасти гипандрия и его медиальную часть. У C. amoena мышцы М1 и М2 разделены, в то время как у P. consorbina имеется одна пара мышц М1 + 2. У C. amoena фаллус имеет явное сочленения с фаллаподемой, в то время как у P. consorbina между ними нет четкой границы. Строение склеритов и мышц эпандриального комплекса у C. amoena заметно сложнее, чем у P. consorbina. У C. amoena сурстили обособлены от субэпандриального склерита и имеют собственную мускулатуру (М4), в то время как у P. consorbina сурстили частично образованы выростами субэпандриального склерита и не имеют собственной мускулатуры.
Наличие собственных мышц у прегонитов грибных мух дает основание утверждать, что эти склериты не выросты гипандрия, как предполагал ряд авторов (Griffiths, 1972; Шаталкин, 1985; Sinclair, 2000, p. 63; Cumming, Wood, 2009, p. 49, Fig. 2.79 –80, p. 47), а производные гонопод–точка зрения, защищавшаяся Макалпайном (McAlpine, 1981) и поддержанная сравнительным изучением мускулатуры (Овчинникова, 1989, 1993; Ovtshinnikova, 1994). Камминг с соавт. (Cumming et al., 1995, p. 127) пришли к мнению, “что прегониты вторично дифференцировались из гонопод (т.е. у Schizophora это новообразования, которые срослись с гипандрием)”. Из этой статьи не вполне ясно, имеются ли прегониты у Aschiza, включая и представителей Platypezidae. В более поздней работе (Cumming, Wood, 2009) этот вопрос проясняется. По мнению авторов, прегониты (их определение дано на странице 49) в качестве структурных элементов гипандрия присущи лишь щеленосным мухам (Schizophora). Сходные структуры у самцов Callomyia ими обозначены (Fig. 2.79 –80, p. 47) как лопасти гипандрия (hypandrial lobe). В целом, однако, наша позиция мало чем отличается от позиции упомянутых авторов. Даже если прегониты являются новообразованием, то онтогенетически они могут возникнуть либо на базе стернита 9, либо на базе слившихся с ним гонопод. Т.е. вполне оправданы предположения и о гоноподальном, и о стернальном происхождении лопастей гипандрия. Примером последних, возможно, может служить непарная лингула у многих Syrphini (см. примеры в: Vockeroth, 1969), которую Затварницкий (Zatwarnicki, 1996, p. 112, Fig. 2) соотнес, на наш взгляд ошибочно, с прегонитом.
Ввиду существенных различий в строении гениталий у C. amoena и P. consorbina, возникает вопрос, какой из вариантов ближе к исходному в семействе Platypezidae. В качестве такового, вероятно, следует принять вариант, который обладает большим сходством с генитальными структурами прямошовных мух (Brachycera Orthorrhapha). Вигман с соавторами (Wiegmann et al., 1993, p. 48, 78; see also Chvala, 1983) предположили, что структура гениталий самцов рода Atelestus Walker, 1837 (Atelestidae) из продвинутых прямошовных мух надсемейства Empidoidea может быть исходной для Eremoneura (Empidoidea + Cyclorrhapha) в целом. Среди грибных мух сходная структура гениталий обнаружена у родов Melanderomyia Kessel 1960 (Melanderomyiinae), Microsania Zetterstedt 1837 (Microsaniinae) и Callomyia (Callomyiinae). Еще ранее предположение об исходном для грибных мух строении гениталий у представителей подсемейств Callomyiinae и Microsaniinae (включая Melanderomyia) высказывал Кессел (Kessel, 1960; см. также Kessel, Maggioncalda, 1968). По существу к тому же мнению пришел Затварницкий (Zatwarnicki, 1996, p. 112, Fig. 2), который рассматривал в качестве основного плана строения гениталий круглошовных мух вариант, характерный для рода Syrphus Fabricius, 1775 (Syrphidae). Отметим, что в структурном плане гениталии Callomyia и Syrphus близки, что хорошо видно из представленных им рисунков (Zatwarnicki, 1996, p. 112, 124). Кроме того, по нашим данным и Callomyia и Syrphidae обладают причлененными придатками гипандрия с мышцами М43, не слитым 8 тергитом и стернитом, а также и не слитыми мышцами фаллаподемы М1 и М2 и эпандриальными мышцами М3 и М4. Следовательно, можно предполагать, что изученный нами род Callomyia – представитель наиболее примитивной группы грибных мух, исходной в эволюции подсем. Platypezinae.
Однако другая часть полученных нами результатов не согласуется с этой гипотезой. Саблевидный фаллус представителей подсемейства Platypezinae, имеющий примитивную дорсальную ориентацию (Griffiths, 1972, p. 37; см. также Chandler, Shatalkin, 1998) должен рассматриваться в качестве анцестрального состояния. Соответственно, подсемейство Callomyiinae в отношении строения гениталий является производной группой. Равным образом семейство Atelestidae внутри Empidoidea является специализированной группой, а исходный статус имеет семейство Empididae, для обоих этих таксонов (Platypezinae и Empididae) характерен саблевидный фаллус с дорсальной ориентацией полового отверстия.
Учитывая вышеописанные различия, мы предполагаем, что общий предок подсемейств Callomyiinae и Platypezinae обладал набором эволюционно исходных признаков (причлененные придатки гипандрия с мышцами М43, не слитые 8 тергит и стернит, не слитые мышцы фаллаподемы М1 и М2, эпандриальные мышцы М3 и М4, саблевидный фаллус с дорсальной ориентацией). В дальнейшем линии, соответствующие подсемействам, эволюционировали независимо и сохранили разный набор анцестральных черт. Соответсвенно, в данной статье корректнее говорить об эволюционно исходных признаках, а не эволюционно исходной группе.
Таким образом, мы предлагаем двухэтапный сценарий морфофункциональных преобразований при переходе от исходного типа гениталий с хорошо развитыми гоноподами к циклорафному типу, в котором гоноподы слились со стернитом 9 (рис. 5). В исходном типе строения гениталий, который отличает прямошовных мух от круглошовных, гоноподы располагаются по бокам от чехла эдеагуса, сочленяясь с его основанием (рис. 5А). Эта трехчленная структура образует интромитантный орган: во время копуляции (данные по азилидам) во влагалище самки помимо фаллуса вводятся вторые членики гонопод.
Рис. 5.
Сценарий морфофункциональных преобразований при переходе от исходного типа гениталий с хорошо развитыми гоноподами к циклорафному типу: A – схема связи фаллуса и гонопод в структуре гениталий некоторых Asilidae (вентральный вид); B – структурная схема гипандрия в подсемействе Platypezinae (вентральный вид) и C – структурная схема гипандрия в подсемействе Callomyiinae (вентральный вид).
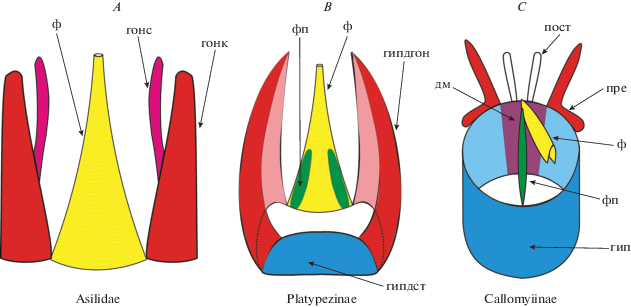
На первом этапе преобразования гениталий исходного типа (рис. 5А) имело место слияние гонопод со стернитом 9 с образованием структуры, имеющей вид желоба (U-образной в сечении). С дорсальной стороны этот желоб прикрывается массивным чехлом эдеагуса, который сохраняет исходный примитивный тип строения, отличающий основную массу продвинутых прямошовных мух (Brachycera Orthorrhapha) из надсемейств Asiloidea и Empidoidea. Образуется промежуточный тип гениталий (рис. 5B), которым, в частности, обладают представители подсемейства Platypezinae. Если связь гипандрия с основанием фаллуса обеспечивается слившимися со стернитом 9 гоноподами, то стернальная часть гипандрия может быть небольшой относительно гоноподиальной части гипандрия, как это характерно для многих Brachycera Orthorrhapha. У Platypezinae стернальная часть гипандрия расположена вентрально. Последующая эволюция, как мы предполагаем, шла в направлении увеличения стернальной части гипандрия за счет уменьшения его гоноподальной части.
На втором этапе преобразования гениталий (рис. 5С) желоб гипандрия замыкается в трубчатую структуру за счет латерального разрастания стернальной части гипандрия и разрастания дорсального моста – склерита, прикрывающего внутреннее пространство гипандрия с дорсальной стороны (он уже имеется у прямошовных двукрылых). Соответственно, гоноподальная часть гипандрия, защищавшая фаллус с боков, уменьшается, образуя пару хорошо различимых лопастей, прегонитов. Фаллус существенно укорачивается; дорсальная пластинка чехла эдеагуса в его базальной части участвует в образовании дорсального моста (Wood, 1991), а вентральная пластинка чехла – в образовании аподемы фаллуса (Овчинникова, 1993, 2000).
БЛАГОДАРНОСТИ
Авторы выражают благодарность Д.И. Гаврюшину и А.Л. Озерову (Зоологический музей МГУ им. М.В. Ломоносова) за предоставленный материал и советы при написании статьи.
Работа Т.В. Галинской (обработка сухого и спиртового материала, подготовка иллюстраций) поддержана грантом РНФ (14-14-00208), подготовка текста рукописи выполнена на базе кафедры энтомологии биологического факультета МГУ им. М.В. Ломоносова (гостема № AAAA-A16-116021660095-7). Работа А.И. Шаталкина поддержана грантом РФФИ (16-04-01358-a) и выполнена на базе Зоологического музея МГУ (классические подходы – гостема № АААА-А16-116021660077-3). Работа О.Г. Овчинниковой выполнена на базе Зоологического института РАН (гостема АААА-А19-119020690082-8) и при финансовой поддержке РФФИ (18-04-00354-а).
Список литературы
Овчинникова О.Г., 1989. Мускулатура гениталий самцов двукрылых Brachycera–Orthorrhapha (Diptera) // Труды Зоол. ин-та АН СССР. Т. 190. Л. С. 1–167.
Овчинникова О.Г., 1993. Гомологизация склеритов гениталий самцов двукрылых (Diptera, Brachycera) на основе изучения мускулатуры // Энтомологическое обозрение. Т. 72. Вып. 4. С. 737–746.
Овчинникова О.Г., 2000. Мускулатура гениталий самцов двукрылых семейства Syrphidae (Diptera) // Чтения памяти Н.А. Холодковского. Вып. 52(2). С. 1–69.
Шаталкин А.И., 1985. Обзор грибных мух (Diptera, Platypezidae) фауны СССР // Сборник трудов Зоологического музея МГУ. Т. 23. С. 69–136.
Chandler P.J., 2001. The flat-footed flies (Diptera: Opetiidae, Platypezidae) of Europe. Fauna Entomologica Scandinavica. V. 36. Leiden-Boston-Köln: E.J. Brill. 276 p.
Chandler P.J., Shatalkin A.I. 1998. Family Platypezidae // Contribution to a Manual of Palaearctic Diptera. Papp L., Darvas B. (Eds). V. 3. Budapest: Science Herald. P. 27–49.
Chillcott J.G.T., 1958. The comparative morphology of the male genitalia of muscoid Diptera // Proceedings tenth International Congress of Entomology. 1956, Montreal. V. 1. P. 387–392.
Chvála M., 1983. The Empidoidea (Diptera) of Fennoscandia and Denmark. II. General Part. The families Hybotidae, Atelestidae and Microphoridae // Fauna Entomologica Scandinavica. V. 12. P. 1–279.
Crampton G.C., 1923. The genitalia of male Diptera and Mecoptera compared with those of related insects from the standpoint of phylogeny // Transaction of the American Entomological Society. V. 48. P. 207–225.
Crampton G.C., 1942. The external morphology of the Diptera // Guide to the Insects of Connecticut. Part VI. The Diptera or true flies of Connecticut. 1st fascicle. Connecticut State Geological and Natural History Survey. Bulletin. V. 64. P. 10–165.
Crampton G.C., 1944. A comparative morphological study of the terminalia of male calyptrate cyclorrhaphous Diptera and their acalyptrate relatives // Bulletin of the Brooklyn Entomological Society. V. 39. P. 1–31.
Cumming J.M., Wood D.M., 2009. Adult Morphology and Terminology // Manual of Central American Diptera. Brown B.V., Borkent A., Cumming J.M., Wood D.M., Woodley N.E., Zumbado M. (Eds). V. 1. Ottawa, Canada: NRC Research Press. P. 9–50.
Cumming J.M., Sinclair B.J., Wood D.M., 1995. Homology and phylogenetic implications of male genitalia in Diptera – Eremoneura // Entomologica Scandinavica. V. 26. P. 121–152.
Emden F. van, Hennig W., 1956. Diptera // Taxonomist’s glossary of genitalia in Insects (Tuxen S.L. ed.). Kopenhagen: Munksgaard. P. 111–122.
Galinskaya T.V., Ovtshinnikova O.G., 2015. Musculature of the male genitalia in the tribe Ulidiini (Diptera, Ulidiidae) // Entomological Review. V. 95(1). P. 31–37.
Galinskaya T.V., Ovtshinnikova O.G., 2015a. Musculature of the male genitalia in Rivellia (Diptera: Platystomatidae) // ZooKeys. V. 545. P. 149–158.
Galinskaya T.V., Gafurova D., Ovtshinnikova O.G., 2018. X‑ray microtomography (microCT) of male genitalia of Nothybus kuznetsovorum (Nothybidae) and Cothornobata sp. (Micropezidae) // ZooKeys. V. 744. P. 139–147.
Griffiths G.C.D., 1972. The phylogenetic classification of Diptera Cyclorrhapha, with special reference to the structure of the male postabdomen // Series Entomologica. V. 8. The Hague: Dr. W. Junk N.V. Publishers. P. 340.
Kessel E.L., 1960. The systematic positions of Platycnema Zetterstedt and Melanderomyia, new genus, together with the description of the genotype of the latter (Diptera: Platypezidae) // Wasmann Journal of Biology. V. 18. P. 87–101.
Kessel E.L., Maggioncalda E.A., 1968. A revision of the genera of Platypezidae, with the description of five new genera, and consideration of phylogeny, circumversion, and hypopygia (Diptera) // Wasmann Journal of Biology. V. 26. P. 33–106.
McAlpine J.F., 1981. Morphology and terminology. Adults // Manual of Nearctic Diptera. McAlpine J.F. (Ed.). V. 2. Canada, Ottawa: Research Branch Agric. V. 1. № 27. P. 9–64.
McAlpine J.F., 1989. Phylogeny and classification of the Muscomorpha // Manual of Nearctic Diptera. McAlpine J.F. (Ed.). Canada, Ottawa: Research Branch Agric. V. 3. P. 1397–1578.
Ovtshinnikova O.G., 1994. On the homology of male genital sclerites of Brachycera Orthorrhapha and Cyclorrhapha (Diptera) based on musculature // Dipterological Research. V. 5. № 4. P. 263–269.
Ovtshinnikova O.G., Galinskaya T.V. 2016. Musculature of the abdomen and male genitalia of a member of Celyphidae (Diptera, Cyclorrhapha) // Oriental Insects. V. 50(4). P. 178–186.
Ovtshinnikova O.G., Galinskaya T.V. 2016a. Musculature of the male genitalia of a member of the genus Teleopsis Rondani, 1875 (Diopsidae, Diptera) // Oriental Insects. V. 50(2). P. 61–68.
Ovtshinnikova O.G., Galinskaya T.V. 2017. The male abdominal, genital and pregenital sclerites and musculature in Neria commutata (Czerny, 1930) (Diptera, Micropezidae) // Entomological Review. V. 97(3). P. 282–287.
Ovtshinnikova O.G., Yeates D.K. 1998. Male genital musculature of Therevidae and Scenopinidae (Diptera: Asiloidea): structure, homology and phylogenetic implications // Australian Journal of Entomology. V. 37 (1). P. 27–33.
Sinclair B.J., 2000. 1.2. Morphology and terminology of Diptera male terminalia // Contributions to a Manual of Palaearctic Diptera. General and Applied Dipterology (Papp L., Darvas B. eds). V. 1. Budapest: Science Herald. P. 53–74.
Sinclair B.J., Cumming J.M., Brooks S.E. 2013. Male terminalia of Diptera (Insecta): a review of evolutionary trends, homology and phylogenetic implications // Insect Systematics & Evolution. V. 44. P. 373–415.
Vockeroth J.R., 1969. A revision of the genera of the Syrphini (Diptera: Syrphidae) // Memoirs of the Entomological Society of Canada. V. 62. P. 1–176.
Wiegmann B.M., Mitter C., Thompson F.C., 1993. Evolutionary origin of the Cyclorrhapha (Diptera): tests of alternative morphological hypotheses // Cladistics. V. 9. P. 41–81.
Wood D.M., 1991. Homology and phylogenetic implications of male genitalia in Diptera. The ground plan. Proceedings of the Second International Congress of Dipterology held in Bratislava, Czechoslovakia, August 27 – September 1, 1990. Weismann, L., Országh, I., Pont A.C. (eds). The Hague. P. 255–284.
Zatwarnicki T., 1996. A new reconstruction of the origin of the eremoneuran hypopygium and its implications for classification (Insecta: Diptera) // Genus. V. 3. P. 103–175.
Дополнительные материалы отсутствуют.
Инструменты
Зоологический журнал