

Зоологический журнал, 2019, T. 98, № 7, стр. 773-777
Акустические сигналы кузнечиков трибы Gampsocleidini (Orthoptera, Tettigoniidae) России
Р. Д. Жантиев 1, *, О. С. Корсуновская 1
1 Биологический факультет Московского государственного университета
им. М.В. Ломоносова
117234 Москва, Россия
* E-mail: zhantiev@mail.ru
Поступила в редакцию 13.12.2018
После доработки 25.12.2018
Принята к публикации 26.12.2018
Аннотация
Описаны временны́е и частотные параметры акустических призывных сигналов самцов Uvarovites inflatus и 4 видов рода Gampsocleis, распространенных на территории России. Показано, что кузнечики трибы Gampsocleidini способны издавать территориальные звуки. Приводятся данные о строении звуковых аппаратов 4 видов Gampsocleis.
Триба Gampsocleidini (Orthoptera, Tettigoniidae) включает монотипический род Uvarovites и род Gampsocleis (Cigliano et al., 2018). На территории России встречаются 7 видов этой трибы. Ареал Gampsocleis glabra (Herbst 1786) простирается от Франции до Алтая, Красноярского края и Западной Монголии. G. schelkovnikovae Adelung 1916 обитает на юго-востоке Европы. G. sedakovii (Fischer von Waldheim 1846) с двумя подвидами: G. s. sedakovii и G. s. obscura (Walker 1869), встречается на обширной территории от востока европейской части России до Сахалина и Приморского края. Восточная граница ареала G. s. sedakovii доходит до Забайкалья и Якутии; G. s. obscura обитает на юге Амурской обл., Хабаровского края, в Приморье и на Сахалине. G. beybienkoi Cejchan 1968 и G. gratiosa Brunner von Wattenwyl 1862 на территории России известны с юга Читинской обл., G. ussuriensis Adelung 1910 – из Забайкалья, Uvarovites inflatus (Uvarov 1924) распространен на юге Амурской обл. и Хабаровского края, а также в Приморье (Стороженко, 2004, 2014).
Сигналы широко распространенного G. glabra хорошо изучены (Latimer, 1980; Heller, 1988). Недавно появились данные о поведении и сигнализации G. sedakovii и U. inflatus из китайских популяций (Li et al., 2010; Zhang et al., 2015; Wang et al., 2016). Однако авторы либо не приводят осциллограммы и частотные спектры, либо не указывают температуру, при которой регистрировали существенно различающиеся по ритмическому рисунку сигналы G. sedakovii. В связи с этим мы считаем необходимым обобщить данные о призывных сигналах и звуковых аппаратах европейских и сибирских представителей трибы Gampsocleidini.
МАТЕРИАЛ И МЕТОДЫ
Кузнечики были собраны в следующих географических точках: Gampsocleis glabra – в окрестностях Курска, в Центрально-черноземном заповеднике (1 самец, 24.07.1970, И. Крицкая) и Северной Осетии, Северо-Осетинском заповеднике (окрестности села Верхний Цей) (1 самец, 08.1985, Д. Тишечкин); G. schelkovnikovae – на Северном Кавказе, в окрестностях Грозного (2 самца, 05.06.1986, В. Чуканов) и Астраханской обл., в окрестностях оз. Баскунчак (1 самец, 02.08.1996, В. Савицкий); G. sedakovii sedakovii – в южной Туве, в окрестностях п. Эрзин, в пойме р. Тес-Хем (4 самца 11.09.1986 и 05.08.1988, О. Корсуновская); G. gratiosa – в КНР, автономном районе Внутренняя Монголия (1 самец, 11.2006, точное местонахождение неизвестно); Uvarovites inflatus – в южном Приморье, Хасанский р-н, окрестностях д. Рязановка (1 самец, 22.09.2001, А. Полилов).
Запись звуковых сигналов осуществляли при помощи студийного магнитофона МЗ-23 (нелинейность частотной характеристики ±2 дБ в диапазоне 2–50 кГц) либо усовершенствованного в лаборатории магнитофона “Юпитер-202 Стерео” с линейной частотной характеристикой в диапазоне 0.063–70 кГц. Некоторые записи производили непосредственно на компьютер с использованием АЦП L-305 (фирма L-Card, Москва) и программы L-Graph (L-Card, Москва). Для регистрации звуков использовали микрофон 4135 и измерительный усилитель 2606 фирмы Вruel and Kjaer (Дания). Обработку временны́х и частотных параметров акустических сигналов проводили в программах TurboLab v. 4.2 и Cool Edit Pro 2.1. Микрофотографии стридуляционного аппарата (pars stridens) получали с помощью сканирующего электронного микроскопа JSM-6380 в лаборатории электронной микроскопии биологического факультета МГУ им. М.В. Ломоносова.
Терминология, используемая при описании амплитудно-временнóй структуры сигналов, принята по Жантиеву (1981): пульс – элемент, возникающий при закрывании надкрылий, интерпульс – при их открывании. Обычно амплитуда последнего меньше, чем у пульса. Однако у ряда видов Gampsocleis движения надкрылий во время стридуляции очень сложны, каждый цикл открывания–закрывания, по-видимому, осуществляется в несколько этапов (Latimer, 1980; Heller, 1988), и с уверенностью отнести каждую звуковую посылку к категории пульсов или интерпульсов невозможно. Поэтому в описаниях таких сигналов звуковые посылки большей амплитуды названы макропульсами, а чередующиеся с ними низкоамплитудные элементы – микропульсами.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Призывные сигналы Gampsocleis glabra (рис. 1А–1В) и G. sedakovii sedakovii (рис. 1F–1G) представляют собой трели, в которых макропульсы чередуются с микропульсами. Структура призывного сигнала G. schelkovnikovae сходна с таковой двух предыдущих видов, однако это не трель, а ритмически повторяющиеся серии длительностью около 3 с, состоящие из многочисленных макропульсов нарастающей амплитуды (рис. 1С). Паузы между макропульсами также заполнены микропульсами (рис. 1D). У всех трех видов в начале сигнала, как правило, присутствует высокоамплитудный пульс c предшествующим интерпульсом, отделенный от последующей трели/длительной серии небольшой паузой. Такой пульс может продуцироваться независимо от последующей длительной фазы сигнала, часто – в ответ на посторонние звуковые раздражители. В последнем случае этот элемент, по-видимому, выполняет функцию территориального сигнала (рис. 1Е). Временнóй паттерн призывных сигналов корейских и японских представителей Gampsocleis (G. buergeri, G. ussuriensis) не отличается от паттерна сигналов изученных нами видов: они издают длительные серии, состоящие из макро- и микропульсов; одиночные громкие пульсы продуцируются независимо от продолжительного сигнала, предшествуют ему или завершают его (Kim, 2010; Hashimoto, 2018).
Рис. 1.
Осциллограммы призывных (A–D, F–K) и территориального (E) сигналов самцов Gampsocleis spp. (A–I) и Uvarovites inflatus (J–K) при разных скоростях развертки. Внизу – отметки времени. Цифры справа – температура при записи сигналов.
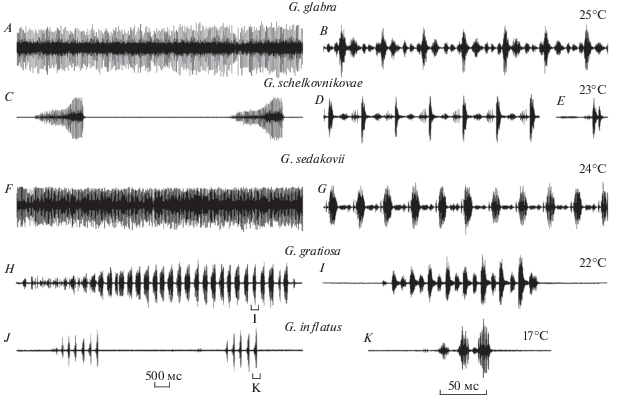
Существенные отличия от описанных звуков наблюдаются в паттерне сигнала у G. gratiosa и U. inflatus. У этих видов призывный сигнал самцов представляет собой ритмически повторяющиеся серии из четырех пульсов (U. inflatus, рис. 1J–1K) или фразы (рис. 1H), состоящие из серий с нарастающей амплитудой, включающие по 7–8 пульсов (G. gratiosa, рис. 1I). Каждому пульсу предшествует интерпульс. Микропульсов и отдельных коротких одиночных звуковых посылок у этих видов не зарегистрировано.
Таким образом, призывные сигналы кузнечиков трибы Gampsocleidini по временнóй структуре можно разделить на две группы. В первую входят сигналы, представляющие собой трели или длительные (от 0.5 до нескольких секунд) серии из многочисленных пульсов, вторая образована последовательностью коротких серий или фраз, состоящих из коротких серий.
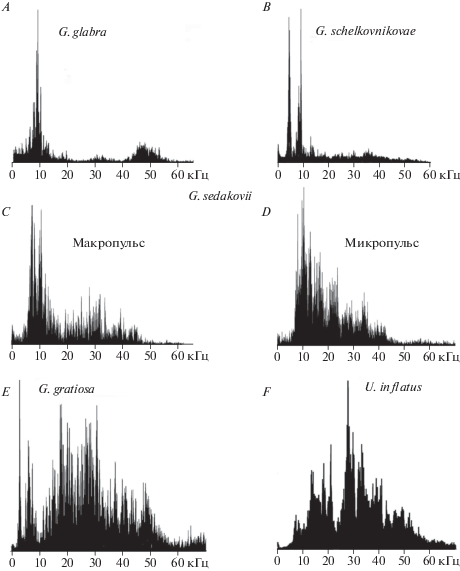
Доминирующие частоты в амплитудно-частотных спектрах призывных сигналов у кузнечиков рода Gampsocleis составляют около 9–10 кГц (G. glabra) (рис. 2А), 5 и 9 (G. schelkovnikovae) (рис. 2B), 7 кГц (G. sedakovii) (рис. 2C–2D) либо занимают широкий диапазон с несколькими максимумами приблизительно равной амплитуды в области 7, 20 и 30 кГц (G. gratiosa) (рис. 2E). Иногда в звуках G. gratiosa хорошо представлены также низкочастотные составляющие (3–4 кГц). У G. glabra в спектре в области 45–50 кГц располагается дополнительный максимум. Доминирующие частоты в спектре U. inflatus (рис. 2F) составляют 28–30 кГц, кроме того, в диапазоне 5–25 кГц можно выделить несколько дополнительных максимумов меньшей амплитуды в области 7, 12, 18 и 22 кГц. Составляющие максимальной амплитуды у G. glabra и G. schelkovnikovae занимают весьма узкий частотный диапазон, за счет чего спектры призывных звуков этих кузнечиков напоминают спектры тональных сигналов сверчков, в то время как спектры G. gratiosa и U. inflatus являются широкополосными шумовыми. Частотный спектр G. sedakovii занимает промежуточное положение. Частотный состав макро- и микропульсов у исследованных видов кузнечиков обычно различен: в спектре микропульсов и интерпульсов может отсутствовать низкочастотный максимум, могут быть представлены в большей степени высокочастотные составляющие, которые (например, у G. sedakovii) иногда образуют выраженные максимумы в области 15–40 кГц (рис. 2С и 2D).
Рис. 2.
Частотные спектры призывных сигналов самцов Gampsocleis spp. и Uvarovites inflatus. По оси абсцисс – частота звука, кГц; по оси ординат – амплитуда сигнала в линейном масштабе.
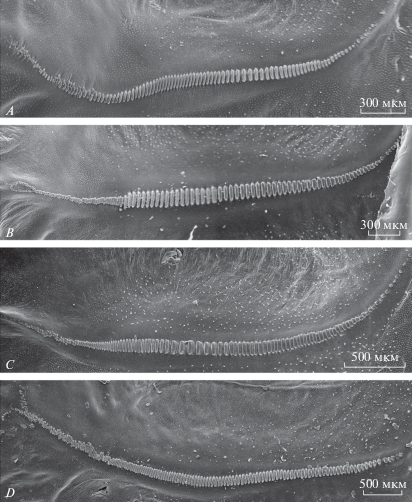
Исследование звукового аппарата кузнечиков рода Gampsocleis показало, что их стридуляционные жилки (pars stridens) обладают сходным строением (рис. 3). Для всех изученных видов характерно наличие на медиальном (противоположном крыловому суставу) участке жилки нескольких десятков мелких, часто округлых и довольно хаотично расположенных зубчиков. К ним примыкает следующий отрезок pars stridens, образованный более крупными, продолговатыми, равномерно распределенными на жилке зубчиками. Возле сустава они вновь уменьшаются. У G. glabra (рис. 3А) и G. sedakovii (рис. 3С) 5–7 серединных зубчиков разделены бóльшими интервалами, чем остальные. Специфическая медиальная зона pars stridens, возможно, используется при эмиссии микропульсов.
Рис. 3.
Стридуляционные жилки самцов: A – Gampsocleis glabra, B – G. schelkovnikovae, C – G. sedakovii, D – G. gratiosa.
У видов Gampsocleis наблюдается расширение акустического репертуара: помимо призывных сигналов они продуцируют территориальные звуки, которые в некоторых поведенческих ситуациях могут выполнять функцию сигналов протеста. Подобное явление наблюдается и у кузнечиков других родов – некоторых Platycleis s. str. (P. affinis Fieber 1853, P. escalerai Bolivar 1899), Сalopterusa pastuchovi (Uvarov 1917), Sphagniana ussuriana (Uvarov 1926) (Korsunovskaya et al., 2002; наши неопубликованные данные). Как правило, эти звуки представлены одиночным пульсом или короткой серией, идентичной по структуре пульсам (сериям) призывного сигнала. Согласно наблюдениям над U. inflatus, G. sedakovii и G. gratiosa, самцы способны продуцировать также прекопуляционные звуки (Jia, Jiang, 1999; Li et al., 2010). Не исключено, что кузнечики рода Gampsocleis, подобно Sphagniana sphagnorum (Walker 1869), могут использовать в брачном поведении автономно издаваемые низкоамплитудные компоненты призывного сигнала (Morris et al., 1975), меняя алгоритм и механику движения надкрылий (Morris, Pipher, 1972). Дальнейшие этологические исследования с применением метода записи звуков самцов синхронно с регистрацией движений их надкрылий должны дать ответ на этот вопрос.
БЛАГОДАРНОСТИ
Авторы благодарят чл.-кор. НАН Украины Л.И. Францевича за предоставленную возможность работы с Gampsocleis gratiosa, В.С. Чуканова, Д.Ю. Тишечкина, В.Ю. Савицкого и А.А. Полилова за сбор и доставку живых насекомых для биоакустических исследований.
Работа выполнена в соответствии с планом НИР кафедры энтомологии Московского государственного университета им. М.В. Ломоносова (“Решение теоретических вопросов общей и прикладной энтомологии”, № договора 32-1-16) и при поддержке РНФ (Russian Science Foundation) (14-50-00029 – “Научные основы создания национального банка-депозитария живых систем” (Scientific basis of the national biobank – depository of the living systems)).
Список литературы
Жантиев Р.Д., 1981. Биоакустика насекомых. М.: Издательство Московского университета. 256 с.
Стороженко C.Ю., 2004. Длинноусые прямокрылые насекомые (Orthoptera: Ensifera) азиатской части России. Владивосток: Дальнаука. 280 с.
Стороженко С.Ю., 2014. Ортопрероидные насекомые (Orthoptera, Mantoptera, Dermaptera) островов залива Петра Великого, Приморский край // Чтения памяти А.И. Куренцова. Вып. 25. С. 49–59.
Cigliano M.M., Braun H., Eades D.C., Otte D., 2018. Orthoptera Species File. Version 5.0/5.0. [Электронный ресурс]. Режим доступа: http://Orthoptera.SpeciesFile.org. Дата обновления: 03.12.2018.
Hashimoto K., 2018. Songs of crickets and katydids from Japan [Электронный ресурс]. Режим доступа: http://mushinone.sakura.ne.jp. Дата обновления: 03.12.2018.
Heller K.-G., 1988. Bioakustik der europaeischen Laubheuschrecken. Weikersheim: Verl. Josef Margraf. 358 S.
Jia Z., Jiang Z., 1999. Ethograms and mating behaviors of Velerifictorus micado and Gampsocleis gratiosa // Acta Zoologica Sinica. V. 45. № 1. P. 49–56.
Kim T.W., 2010. Sound guide to Korean katydids. Seoul. 136 p.
Korsunovskaya O.S., Zhantiev R.D., Savitsky V.Yu., 2002. The songs of the Palaearctic bush crickets of the tribe Drymadusini (Orthoptera, Tettigoniidae) // Russian Entomological Journal. V. 11. P. 335–350.
Latimer W., 1980. Song and spacing in Gampsocleis glabra (Orthoptera, Tettigoniidae) // Journal of Natural History. V. 14. № 2. P. 201–213.
Li T., Yang C., Lin Y., 2010. Comparative study on singing behaviors of the three bushcrickets // Journal of Shandong Agricultural University (Natural Science Edition). V. 41. P. 65–69.
Morris G.K., Kerr G.E., Gwynne D.T., 1975. Calling song function in the bog katydid, Metrioptera sphagnorum (F. Walker) (Orthoptera, Tettigoniidae): female phonotaxis to normal and altered song // Zeitschrift fuer Tierpsychologie. Bd 37. S. 502–514.
Morris G.K., Pipher R.E., 1972. The relation of song structure to tegminal movement in Metrioptera sphagnorum (Orthoptera: Tettigoniidae) // Canadian Entomologist. V. 104. P. 977–985.
Wang Y., Zhao H., Zhang X., Ren B., 2016. The complete mitochondrial genome and song evolution of the monotypic genus U. Tarbinsky, 1932 (Orthoptera: Tettigoniidae) // Environmental Entomology. V. 45. № 3. P. 737–746.
Zhang X., Wen M., Li J., Zhu H., Wang Y., Ren B., 2015. Acoustic, genetic and morphological variations within the katydid Gampsocleis sedakovii (Orthoptera, Tettigonioidea) // ZooKeys. V. 529. P. 105–121.
Дополнительные материалы отсутствуют.
Инструменты
Зоологический журнал