

Зоологический журнал, 2019, T. 98, № 7, стр. 836-842
Гаплотипическое разнообразие домашней овцы (Ovies aries) (по материалам археологических памятников ранней бронзы юга Западной Сибири)
М. А. Дымова 1, 2, *, А. А. Тишкин 2, О. В. Мишукова 1, Е. А. Храпов 1, С. П. Грушин 2, М. Л. Филипенко 1, 2
1 Институт химической биологии и фундаментальной медицины СО РАН
630090 Новосибирск, Россия
2 Алтайский государственный университет
656049 Барнаул, Россия
* E-mail: maya.a.rot@gmail.com
Поступила в редакцию 10.11.2017
После доработки 24.12.2018
Принята к публикации 04.01.2019
Аннотация
Сравнительный анализ генетического разнообразия древних и современных овец может внести ясность в вопросы происхождения этих животных и их дальнейшего распространения, а также помочь определить роль человека на каждом этапе формирования пород. Для оценки популяционной истории овец по материнской и отцовской линиям, костные остатки которых обнаружены в археологических комплексах периода ранней бронзы на юге Западной Сибири, мы анализировали локус SRY (sex-determining region Y) и микросателлитный участок SRYM18 в 19 образцах. Для шести образцов было показано, что животные были самцами. В частности, две особи имели гаплотип H6, еще две – гаплотип H8 и по одной особи – гаплотипы H4 и H10, соответственно. Относительно высокое гаплотипическое разнообразие, а также показанное ранее авторским коллективом наличие двух базальных гаплотипов контрольного региона мтДНК свидетельствуют, что юг Западной Сибири мог быть своеобразным “транспортным узлом” при миграции людей в период ранней бронзы. Необходимы дальнейшие исследования с привлечением существенных выборок пригодных костных материалов из археологических раскопок и с использованием нуклеотидных последовательностей большего числа генов.
Одним из первых одомашненных животных, сыгравших важную роль в системе жизнеобеспечения древнейших социумов, является овца (Ovis aries (Linnaeus 1758)). Различные породы овец в ходе миграций и взаимодействий человеческих сообществ распространились по Евразии и другим континентам. Предполагается, что овцы были одомашнены около 9–11 тыс. лет назад на территории “плодородного полумесяца” (территория от северного Загроса до юго-восточной Анатолии) (Tapio et al., 2006). Последующее разведение этих животных, наряду с естественной и интенсивной искусственной селекцией, привело к появлению не менее 1400 пород. В течение указанного времени деятельность людей играла важную роль в формировании необходимых хозяйственно-полезных признаков у овец. Сравнительный анализ генетического разнообразия древних и современных овец способствует лучшему пониманию происхождения этих животных и их распространения, а также позволяет оценить роль человека на разных этапах формирования современного разнообразия пород овец (Lv et al., 2015).
Регион Монгольского плато играл роль “транспортной развязки” миграционных потоков, имевших место 5–7 тыс. лет назад (Lv et al., 2015). Поэтому археологические памятники, находящиеся на этой и сопредельных территориях, могут содержать важную информацию для исследования происхождения разных популяций домашних овец. Они также позволяют проследить возможные пути распространения животных, установить наличие у древнего населения опыта и методов искусственного отбора наиболее продуктивных мясных, мясомолочных и шерстяных пород домашней овцы.
Одним из актуальных вопросов современной науки является определение периода становления производящего хозяйства на территории Сибири (к северу от Монгольского плато). Фактическим маркером этого важного исторического этапа является наличие костных остатков овец в археологических памятниках Западной и Южной Сибири. Наиболее древними считаются такие находки из объектов афанасьевской культуры Алтая (конец IV–1-я половина III тысячелетия до н.э.). Однако их палеогенетическое изучение и AMS-датирование пока не осуществлялись. В течение длительного времени сотрудники Алтайского государственного университета исследовали поселенческие и погребально-поминальные комплексы, относящиеся к елунинской археологической культуре, которая в настоящее время определяется такими хронологическими рамками: последняя треть III – первая четверть II тысячелетия до н.э. Среди этих изученных памятников важное значение имеет древнее поселение Березовая Лука, которое характеризуется как культурно-хозяйственный центр и на основании радиоуглеродного анализа датируется XXII–XIX веками до н.э. (Кирюшин и др., 2011). Раскопки указанного памятника позволили получить колоссальный объем археологических находок (сотни тысяч экземпляров). Среди найденных костей преобладают остатки домашних животных (около 99%), при этом установлено, что доминирующее место занимают остатки мелкого рогатого скота (свыше 55%). Эти и другие свидетельства из древних и средневековых памятников Верхнего Приобья, Алтая, Восточного Казахстана и Северо-Западной Монголии позволяют приступить к молекулярно-генетическим исследованиям изменчивости древней домашней овцы на юге Западной Сибири. Данному процессу будут способствовать радиоуглеродные датировки и археологический контекст.
Молекулярно-генетические исследования древних овец базируются как на изучении митохондриальной ДНК, так и на анализе ядерных генов. Митохондриальная ДНК является полезным маркером для изучения эволюции по материнской линии, что обусловлено высокой скоростью мутирования и высокой копийностью митохондриального генома. Для оценки вклада отцовской линии в развитие породы необходимо получить информацию по последовательностям нескольких генов Y-хромосомы. На сегодняшний день в качестве филогенетических маркеров на Y-хромосоме рассматривают 5'-промоторной район локуса SRY, в частности маркеры OY1 (AY604734.2:g.67A>G), OY5 (AY604734:g.157C>T), и микросателлитный участок SRYM18, в частности ([TTTTG]m, Indel(G/-), [TG]n, Allele (bp)) (Meadows, Kijas, 2009). Исследование данных локусов, вероятно, имеет особое значение для одомашненных видов, где контролируемые спаривания приводят к тому, что небольшое количество самцов вносит непропорционально большой генетический вклад в последующие поколения. Для филогении современных представителей Ovis aries локусы, находящиеся на Y-хромосоме, впервые использованы в работе Мэдоус с соавторами (Meadows et al., 2006), где для 14 особей мужского пола определили последовательность девяти фрагментов генов MSY (the male-specific region). Исследования генетического разнообразия 5'-промоторного района SRY локуса Y-хромосомы древних образцов овцы уже проведены в Эстонии (Rannamäe et al., 2016) и Финляндии (Niemi et al., 2013). Однако археологические образцы относились к более позднему времени, начиная с периода поздней бронзы и заканчивая поздним средневековьем, а локус SRYM18 в этих работах изучен не был. При этом в обоих исследованиях был найден аллель G-OY1 SRY локуса, характерный для современных североевропейских пород овец. Ранее нами обнаружено, что в период ранней бронзы на юге Западной Сибири существовали не только гаплогруппы A и B, но и новые, ранее не описанные гаплогруппы (Dymova et al., 2017).
Целью данной работы стало дальнейшее исследование генетического разнообразия древних овец периода ранней бронзы с использованием маркеров Y-хромосомы. В частности, не только 5'-промоторного района локуса SRY, но и, впервые, микросателлитного участка SRYM 18, что позволяет получить более ценную информацию для понимания происхождения домашних овец в Сибири.
МАТЕРИАЛ И МЕТОДЫ
Археологические памятники
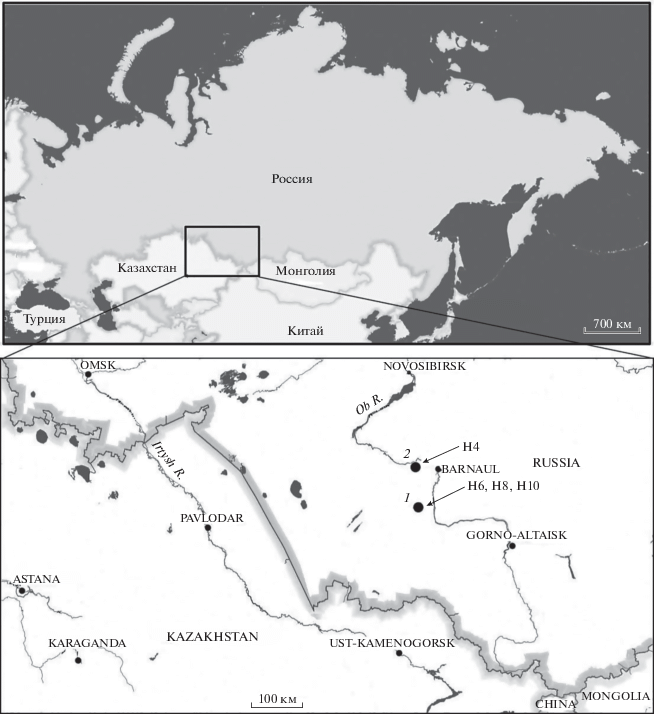
В данной работе использованы костные остатки овец из следующих археологических памятников: поселение Березовая Лука (Россия, Алтайский край, елунинская археологическая культура, конец III–начало II тыс. до н.э., раскопки А.А. Тишкина и С.П. Грушина), поселение Колыванское-I (Россия, Алтайский край, елунинская археологическая культура, конец III–начало II тыс. до н.э., раскопки С.П. Грушина); курганный могильник Яломан-II (Россия, Республика Алтай, булан-кобинская археологическая культура, 2-я половина IV – 1-я половина V вв. н.э., раскопки А.А. Тишкина); могильник Мышиный Лог-I (Россия, Алтайский край, сросткинская археологическая культура, IX–X вв. н.э., раскопки С.П. Грушина); могильник Телеутский Взвоз-I (Россия, Алтайский край, елунинская археологическая культура, конец III – начало II тыс. до н.э.; кармацкая археологическая культура, XIII–XIV вв. н.э., раскопки А.А. Тишкина и С.П. Грушина). Три указанных памятника (Березовая Лука, Колыванское-I и Телеутский Взвоз-I) являются базовыми для характеристики елунинской культуры периода ранней бронзы (согласно локальной периодизации), демонстрирующей производящий тип животноводческого хозяйства. Особое внимание привлекает образец № 77 из могилы-24 памятника Телеутский Взвоз-I. Он происходит из объекта, который характеризуется относительно большими размерами, особой формой и наличием деревянной погребальной камеры. Такими параметрами обладали 13 из 51 исследованного погребения на указанном могильнике. Остальные перечисленные археологические памятники имеют более позднее происхождение (Яломан-II – предтюркское время, Мышиный Лог-I – раннее Средневековье). Материалы из них привлекались для сравнительного анализа. Ниже приведена географическая карта-схема (рисунок) с нанесенными на ней археологическими комплексами, которые находятся на юге Западной Сибири (в южном Обь-Иртышском междуречье). Именно на них найдены костные остатки особей мужского пола. Там же указаны их гаплотипы.
Коллекция костных остатков
В работе использованы 19 костных остатков овец, обнаруженных в выше обозначенных археологических памятниках. Для шести из них удалось получить ампликоны ДНК гена SRY и локуса SRYM18: их месторасположение и название, культурная принадлежность и датировка, а также тип костных остатков и другая информация представлена в табл. 1.
Таблица 1.
Название, структура и размер фланкирующего праймерами региона
| Название | Структура | Размер, п. н. |
|---|---|---|
| mm-Probe | 5'-FAM-CTGGGCTCCCTACAACATCGTCC-BHQ-3' | 79 |
| mm-R2 | 5'-CAAAGATTTCCTGGAAGGTGC-3' | |
| mm-U3 | 5'-GTGATCATGATTGTCTACTTTCTCT-3' | |
| SRYM18F | 5'-GGCATCACAAACAGGATCAGCAAT-3' | 139 |
| SRYM18R | 5'-GTGATGGCAGTTCTCACAATCTCCT-3' | |
| SRY_U1 | 5'-AGCTCCAGAATATTTCACTGACCT-3' | 130 |
| SRY_D1 | 5'-GAAGGCAAATGCAGAGACAA-3' |
Выделение аДНК
Все манипуляции осуществлялись по правилам работы с археологической ДНК. Выделение аДНК проводили по описанному ранее методу с некоторыми модификациями, параллельно ставили отрицательный контроль выделения (Pääbo, Gifford, Wilson, 1988; Dymova et al., 2017).
Амплификация аДНК
Для проверки эффективности выделения ДНК ставили ПЦР в “режиме реального времени” с праймерами mm-U3/mm-R2 и TaqMan зондом mm-Probe. Для амплификации локуса SRY (sex-determining region Y) размером 130 н.п. и микросателлитного участка SRYM18 размером 139 п.н. были использованы праймеры SRY U1/D1 и SRYM18F/R, соответственно (Meadows et al., 2006; Meadows, Kijas, 2009). Структура олигонуклеотидных праймеров и зонда приведена в табл. 2.
Таблица 2.
Общая информация об археологических образцах, исследуемых в работе
| № | Год раскопок |
Номер GenBank | Регион | Археологический комплекс/ культура | Могила | Датировка | Объект | SRYM18 | SRY | Y-гаплотип ([TTTTG]m, Indel G/-, [TG]n, Allele (bp), OY1, OY5) | Гаплотип | Гаплотип мт ДНК |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 44 | 1998 | KU714622 | Алейский р-н Алтайского края, около с. Безголосово, правый берег р. Алей | Поселение Березовая Лука, елунинская археологическая культура | Раскоп 1, квадрат PС – 19–20, зольник в жилище № 1 | Конец III– 1-я четверть II тыс. до н.э. (в рамках периода ранней бронзы) | Кусок лопатки | + | + | 3,G, 15, 143, A, C | H6 | B |
| 45 | 1998 | KU714623 | Алейский р-н Алтайского края, около с. Безголосово, правый берег р. Алей | Поселение Березовая Лука, елунинская археологическая культура | Раскоп 1, квадрат Р–21, жилище № 1 | Конец III– 1-я четверть II тыс. до н.э. (в рамках периода ранней бронзы) | Кость нижней конечности (пястная или плюсневая) | + | + | 3,G, 14, 141, A, C | H8 | B |
| 54 | 1997 | KU714625 | Алейский р-н Алтайского края, около с. Безголосово, правый берег р. Алей | Поселение Березовая Лука, елунинская археологическая культура | Раскоп 1, квадрат К–1, слой 3 | Конец III– 1-я четверть II тыс. до н.э. (в рамках периода ранней бронзы) | Фаланга 2 | + | + | 3,G, 14, 141, A, C | H8 | B |
| 74 | 2000 | KU714631 | Алтайский край, Павловский р-н, 5 км к северо-востоку от с. Елунино | Погребально-поминальный комплекс Телеутсский Взвоз-I, елунинская археологическая культура | Могила-28 | Конец III– 1-я четверть II тыс. до н.э. (в рамках периода ранней бронзы) | Нижняя челюсть, Р/1, Р/2, Р/3, Р/4, М/1 | + | + | 3,G, 16, 145, A, C | H4 | B |
| 88 | 2006 | N/A | Алейский р-н Алтайского края, около с. Безголосово, правый берег р. Алей | Поселение Березовая Лука, елунинская археологическая культура | Раскоп 3, квадрат 36–96, яма 694 | Конец III– 1-я четверть II тыс. до н.э. (в рамках периода ранней бронзы) | Тазовая кость | – | + | N/A, N/A, N/A, N/A, A, C | H10 | N/K |
| 90 | 2004 | N/A | Алейский р-н Алтайского края, около с. Безголосово, правый берег р. Алей | Поселение Березовая Лука, елунинская археологическая культура | Раскоп 2, квадрат ЗИ – 25′–26′/ слои 3–4 | Конец III– 1-я четверть II тыс. до н.э. (в рамках периода ранней бронзы) | Лопатка | + | + | 3,G, 15, 143, A, C | H6 | N/K |
Полимеразную цепную реакцию проводили в конечном объеме 20 мкл, содержащем 65 мМ Трис-НСl (рН 8.9), 23 мМ (NH4)2SO4, 3.0 мМ MgCl2, 0.05% Tween 20, 0.2 мМ дНТФ, 0.3 мкМ растворы олигонуклеотидных праймеров, 0.1 мкМ растворы TaqMan-зондов (в случае проверки эффективности выделения ДНК), 20 нг ДНК и 1 ед. акт. Pfu-ДНК-полимеразы. ПЦР в режиме реального времени с использованием TaqMan-зондов проводили на ДНК-амплификаторе CFX96 (BioRad, США) с начальной денатурацией при температуре 96°С – 3 мин, далее в течение 50 циклов: 96°С – 10 с, 63°С – 40 с и съемом флуоресценции. Каждую реакцию ПЦР дублировали. ПЦР для наработки фрагмента мтДНК, и локусов SRY и SRYM18 проводили с начальной денатурацией при температуре 96°С – 3 мин, далее в течение 40 циклов: 96°С – 5 с, 60°С – 5 с, 72°С – 15 с, завершая конечной элонгацией при температуре 72°С – 3 мин. Наличие продуктов амплификации проверяли электрофоретически в 6% полиакриламидном геле с последующей визуализацией бромистым этидием.
РЕЗУЛЬТАТЫ
В данном исследовании с помощью секвенирования локусов SRY и SRYM18 среди 19 древних овец обнаружены шесть самцов. Для типирования мы использовали ранее описанную номенклатуру гаплотипов, основанную на полиморфизме 5'-промоторного района локуса SRY, в частности маркеры OY1, OY5, и микросателлитного участка SRYM18, в частности ([TTTTG]m, Indel (G/-), [TG]n, Allele (bp)). В результате типирования выявлено, что две особи (образцы № 44, № 90) имели гаплотип H6, две (№45, №54) – гаплотип H8, по одной – гаплотипы H4 (№ 74) и H10 (№ 88). Результаты типирования приведены в табл. 2.
ОБСУЖДЕНИЕ
В данной работе было исследовано генетическое разнообразие древних овец периода ранней бронзы с вовлечением маркеров Y-хромосомы. Ранее было показано, что OY1 -наиболее информативный локус среди OY1-OY8 для одомашненных овец, однако для дифференциации диких овец он не подходит. Для этого можно использовать маркер OY5 (AY604734:g.157C>T), который идентичен для трех видов, далеких от Ovis aries: гривистого барана (Ammotragus lervia (Pallas 1777)), толсторога (Ovis canadensis (Shaw 1804)), барана Далла (Ovis dalli (Nelson 1884)) (Meadows et al., 2006). Важно, что найденный во всех шести случаях аллель A-OY1 в 5’-промоторном районе локуса SRY является самым распространенным среди одомашненных современных овец (71.4%) и предковым аллелем. Мутационное событие, приведшее к образованию аллеля G-OY1, вероятно, произошло уже после завершения одомашнивания O. aries. Также стоит отметить, что гаплотип H6 является самым распространенным (как по обширности географии, так и по частоте встречаемости), остальные гаплотипы встречаются реже (Meadows, Kijas, 2009). Гаплотип Н6, наряду с H8 и H10, обнаружен у овец из поселения Березовая Лука, датируемого в пределах периода ранней бронзы (конец III–начало II тыс. до н.э.), а гаплотип H4 найден в археологическом памятнике Телеутский Взвоз-I (конец III–начало II тыс. до н.э.). Таким образом, можно предположить, что уже в период ранней бронзы на территории юга Западной Сибири существовали гаплотипы одомашненных овец, встречающиеся и по сей день.
Ранее нами уже показано разнообразие контрольного района Д-петли мтДНК древних овец периода ранней бронзы и сравнение их с современными овцами (Dymova et al., 2017). Была оценена эволюционная дивергенция между исследуемыми последовательностями древней мтДНК методом максимального правдоподобия с использованием программы MEGA6, при этом обнаружены два образца, имеющие базальное положение по отношению к другим исследуемым костным остаткам. Приблизительное время расхождения гаплотипов внутри гаплогруппы составило 1.7 млн лет назад для образца № 77 и 0.96 млн лет назад для № 56. На основании этих данных мы предположили, что древние овцы могли иметь большее разнообразие, нежели современные, и возможно, существовали еще две вымершие линии: G (№ 56) и F (№ 77). Стоит отметить, что образец (№ 77) имел гомологию с последовательностью, полученной от домашней овцы c Иберийского полуострова. Современные иберийские породы домашнего скота являются результатом влияния нескольких миграций и вторжений на протяжении истории человечества. Овцы, изначально происходившие из Азии, распространились в Европу и достигли Пиренейского полуострова в начале V тыс. до н.э. (Pedrosa et al., 2007). Вторая миграция овец из Азии произошла в результате вторжения кельтов примерно в 700 г. до н.э., а все последующие связаны с нашествием финикийцев, греков, римлян и позднее мавров.
ЗАКЛЮЧЕНИЕ
В данной работе по исследованию гаплотипического разнообразия по отцовской линии мы обнаружили уже известные гаплотипы одомашненных овец (H6, H8, H4, H10). Генетический анализ мтДНК древних овец указывает на то, что, по крайней мере, две линии (A и B) существовали на юге Западной Сибири в период ранней бронзы. Это коррелирует с литературными данными, описывающими процесс расселения ближневосточной овцы на восток Евразии (включая Монголию, Китай, Индию) через Кавказ и Центральную Азию, примерно 4.5–6.8 тыс. лет назад (Lv et al., 2015). Ранее показано, что гаплогруппа А характерна для ранней бронзы в Китае (Cai et al., 2011). В настоящее время гаплогруппа А остается доминирующей (58.73%), следом за ней идет гаплогруппа В с частотой встречаемости 24,68%, и гаплогруппа C – 16.59%. (Luo et al., 2005). Ранее нами обнаружено, что в период ранней бронзы на юге Западной Сибири существовали не только гаплогруппы A и B, но и новые, ранее не описанные гаплогруппы (Dymova et al., 2017). Существование двух базальных гаплотипов, большое гаплотипическое и нуклеотидное разнообразие указывают на высокую гетерогенность домашних овец в прошлом, а также тот факт, что одомашнивание не привело к резкому снижению генетического разнообразия из-за эффекта “бутылочного горлышка”, что характерно для других домашних животных (Kijas et al., 2012). Можно предположить, что юг Западной Сибири в период ранней бронзы являлся миграционным коридором и/или регионом, который находился в выигрышном положении от активного обмена между различными культурами, что отразилось на высоком гаплотипическом разнообразии овец.
БЛАГОДАРНОСТИ
Авторы благодарны В.А. Трифонову за критический обзор рукописи и ценные замечания.
Работа выполнена при финансовой поддержке РНФ, проект № 16-18-10 033 “Формирование и эволюция систем жизнеобеспечения у кочевых социумов Алтая и сопредельных территорий в поздней древности и средневековье: комплексная реконструкция”.
Список литературы
Кирюшин Ю.Ф., Грушин С.П., Тишкин А.А., 2011. Березовая Лука – поселение эпохи бронзы в Алейской степи.Т. II. Барнаул: Издательство Алтайского университета. 171 с.
Cai D., Tang Z., Yu H., Han L., Ren X., Zhao X., Zhou H., 2011. Early history of Chinese domestic sheep indicated by ancient DNA analysis of Bronze Age individuals // Journal of Archaeological Science. V. 38. I. 4. P. 896–902.
Dymova M.A., Zadorozhny A.V., Mishukova O.V., Khrapov E.A., Druzhkova A.S., Trifonov V.A., Filipenko M.L., 2017. Mitochondrial DNA analysis of ancient sheep from Altai // Animal Genetics. V. 48. I. 5. P. 615–618.
Kijas J.W., Lenstra J.A., Hayes B., Boitard S., Porto Neto L.R., San Cristobal M., 2012. Genome-wide analysis of the world’s sheep breeds reveals high levels of historic mixture and strong recent selection // PLoS Biology. V. 10. I. 2. e1001258.
Luo Y.-Z., Cheng S.-R., Batsuuri L., Badamdorj D., Olivier H., Han J.-L., 2005. [Origin and genetic diversity of Mongolian and Chinese sheep using mitochondrial DNA D-loop sequences] // Yi Chuan Xue Bao. V. 32. I. 12. P. 1256–1265.
Lv F.-H., Peng W.-F., Yang J., Zhao Y.-X., Li W.-R., et al., 2015. Mitogenomic Meta-Analysis Identifies Two Phases of Migration in the History of Eastern Eurasian Sheep // Molecular Biology and Evolution. V. 32. I. 10. P. 2515–2533.
Meadows J.R.S., Hanotte O., Drögemüller C., Calvo J., Godfrey R., et al., 2006. Globally dispersed Y chromosomal haplotypes in wild and domestic sheep // Animal Genetics. V. 37. I. 5. P. 444–453.
Meadows J.R.S., Kijas J.W., 2009. Re-sequencing regions of the ovine Y chromosome in domestic and wild sheep reveals novel paternal haplotypes // Animal Genetics. V. 40. I. 1. P. 119–123.
Niemi M., Bläuer A., Iso-Touru T., Nyström V., Harjula J., et al., 2013. Mitochondrial DNA and Y-chromosomal diversity in ancient populations of domestic sheep (Ovis aries) in Finland: comparison with contemporary sheep breeds // Genetics, Selection, Evolution: GSE. V. 45. P. 2–14.
Pääbo S., Gifford J.A., Wilson A.C., 1988. Mitochondrial DNA sequences from a 7000-year old brain // Nucleic Acids Research. V. 16. I. 20. P. 9775–9787.
Pedrosa S., Arranz J.-J., Brito N., Molina A., San Primitivo F., Bayón Y., 2007. Mitochondrial diversity and the origin of Iberian sheep // Genetics, Selection, Evolution: GSE. V. 39. P. 91–103.
Rannamäe E., Lõugas L., Niemi M., Kantanen J., Maldre L., et al., 2016. Maternal and paternal genetic diversity of ancient sheep in Estonia from the Late Bronze Age to the post-medieval period and comparison with other regions in Eurasia // Animal Genetics. V. 47. I. 2. P. 208–218.
Tapio M., Marzanov N., Ozerov M., Cinkulov M., Gonzarenko G., et al., 2006. Sheep mitochondrial DNA variation in European, Caucasian, and Central Asian areas // Molecular Biology and Evolution. V. 23. I. 9. P. 1776–1783.
Дополнительные материалы отсутствуют.
Инструменты
Зоологический журнал