

Зоологический журнал, 2019, T. 98, № 7, стр. 819-824
К организации пения длиннохвостого сорокопута (Lanius schach erythronotus)
А. С. Опаев *
Институт проблем экологии и эволюции им. А.Н. Северцова РАН
119071 Москва, Россия
* E-mail: aleksei.opaev@gmail.com
Поступила в редакцию 31.08.2018
После доработки 25.10.2018
Принята к публикации 12.12.2018
Аннотация
На примере одного самца впервые описано песенное поведение длиннохвостого сорокопута. Проанализировано 44.2 мин записей этой особи, полученных на юге Казахстана 24 мая–16 июня 2007 г. Рекламная вокализацию включала два чередующихся типа акустических структур – серии позывок (длительностью 3–282 с каждая, медиана 52 с, n = 18) и эпизоды пения (6–202 с каждый, медиана 67 с, n = 22). Репертуар позывок самца включал 13 их типов. Каждая серия позывок включала один или несколько типов позывок. Эпизоды пения состояли из песенных слогов – всего 143 типа в репертуаре самца. Каждый песенный слог относился к одной из одиннадцати композиций, каждая композиция содержала от 3 до 33 типов песенных слогов (медиана 8 слогов). Во время одного эпизода пения, самец сначала исполнял слоги одной композиции, затем переходил к следующей композиции, и т.д.
Песня воробьиных птиц – один из самых сложных сигналов в мире животных. К настоящему времени накопилось много примеров сложной организации пения. Особенно впечатляют способности некоторых видов запоминать и воспроизводить в неизменном виде довольно протяженные акустические конструкции. Так, самцы пеночек голосистой (Phylloscopus schwarzi) и сероголовой очковой (Ph. (Seicercus) tephrocephalus) способны к многократному повторению последовательностей из нескольких десятков стереотипных типов песен, длительностью около секунды каждая (Ivanitskii et al., 2012; Opaev, 2016). Это – яркие примеры так называемого линейного синтаксиса, при котором каждый данный тип песни определяет следующий за ним (Иваницкий, 2015). Не менее характерен для воробьиных птиц комбинаторный синтаксис. В этом случае выделяются кластеры взаимно-ассоциированных вокальных элементов (например, типов песен), а порядок их исполнения внутри кластера варьирует (Иваницкий, 2015; Sasahara et al., 2012; Weiss et al., 2014; Cody et al., 2016). Обычны и варианты организации пения, переходные между линейным и комбинаторным синтаксисами.
Структура и организация пения изучена далеко не у всех видов, вокализацию которых по первому впечатлению можно отнести к “сложной”. Одна из таких групп – сорокопуты (семейство Laniidae). По характеру сигнального поведения, Панов (2008) делит все виды рода Lanius (29 видов), центрального в семействе, на (1) группу серых сорокопутов и (2) всех прочих. Рекламная вокализация первых довольно проста. Так, индивидуальные репертуары серого L. excubitor и клинохвостого L. spenocercus сорокопутов включают всего несколько (два–три) типов коротких песен длительностью не более 0.5 с каждая. При пении разные типы песен чередуются по принципу периодической вариативности: сорокопут много раз (обычно – более 10) повторяет один и тот же тип песни, а затем переходит к следующему типу (Опаев, 2012, 2013). По-иному обстоит дело у прочих Lanius (вторая группа) – по крайней мере, у видов фауны бывшего СССР, по которым имеются детальные наблюдения Панова (2008). У них функциональным аналогом песен являются, во-первых, короткие широкополосные позывки, до нескольких типов в репертуаре каждой особи. Во-вторых, в моменты наибольшего возбуждения серию позывок сменяет “импровизационная” песня (Панов, 2008). Такая песня может быть более характерна для холостых самцов (например, по нашим наблюдениям, туркестанского жулана (L. phoenicuroides) и длиннохвостого сорокопута), либо может быть использована при взаимодействиях с самкой. Так, у самцов чернолобых сорокопутов (L. minor) пение, сопровождаемое серией поклонов, – характерный элемент ухаживания (Панов, 2008; наши наблюдения). “Импровизационное” пение сорокопутов на первый взгляд выглядит разнообразным и сложным. Однако детальный анализ его структуры и организации до сих пор не был проведен ни для одного из видов. Одной из причин этого может быть необходимость анализа очень длинных фонограмм. Между тем, для сорокопутов собрать такой материал не просто. Ведь в отличие, например, от соловья (Luscinia luscinia), сорокопуты никогда не поют долго. Эпизоды пения редко превышают несколько минут и разделены сериями позывок либо продолжительными периодами молчания.
В этой связи представляет интерес анализ 14 фонограмм рекламной вокализации одного самца длиннохвостого сорокопута общей длительностью более 44 мин, записанных на протяжении 24 дней. Это – первое аналитическое описание “импровизационной” песни сорокопутов, хотя и выполненное на примере одного самца.
МАТЕРИАЛ И МЕТОДЫ
Объект исследования, сбор и объем материала
Длиннохвостый сорокопут, или шах имеет обширную область распространения, обитая на большей части Средней, Южной и Юго-восточной Азии, а также на Филиппинах и Зондских островах. На этом ареале выделяют не менее 9 подвидов (Панов, 2008; del Hoyo, Collar, 2016), окрасочные и морфометрические различия между которыми могут быть достаточно велики. В настоящей работе речь идет о среднеазиатской форме L. s. erythronotus. На юге Казахстана и в Средней Азии эти птицы – типичные обитатели культурного ландшафта, реже встречаются на удалении от человеческих жилищ. В антропогенный ландшафт этого сорокопута притягивает древесная растительность, с которой он здесь тесно связан. Птицы живут в садах, парках, кишлаках и аулах, а также в зеленых кварталов городов (Панов, 2008; наши данные).
В 2007 г. один самец этого вида на протяжении почти месяца держался среди десятка деревьев, растущих у одиночно стоящей кошары на южном берегу озера Бийликоль (южный Казахстан, Жамбылская обл.). Наблюдения и запись акустических сигналов проведены в период с 24 мая по 16 июня (24 дня). Почти все это время самец был холостым и токовал в одиночестве. Лишь 30 мая мы наблюдали на его участке предположительно самку, с которой самец в тот день активно взаимодействовал. Однако уже на следующий день эта вторая птица исчезла.
Для записи пения самца использована видеокамера Panasonic NV-GS11, оснащенная встроенным микрофоном с полосой пропускания до 10 кГц. Извлечение звуков с видео осуществляли при помощи программы Adobe Audition 1.5. Для анализа выбрано 14 непрерывных фонограмм, содержащих либо только пение (4 записи), либо пение и позывки (10 записей). Длительность каждой фонограммы 70–509 с, медиана 165.5 с. Суммарная длительность всех записей 2651 с (44.2 мин).
Описание и анализ фонограмм
Визуализацию и обработку записей проводили в программе Syrinx. При построении спектрограмм использовали окно Блэкмана и длину быстрого преобразования Фурье равную 256 точек.
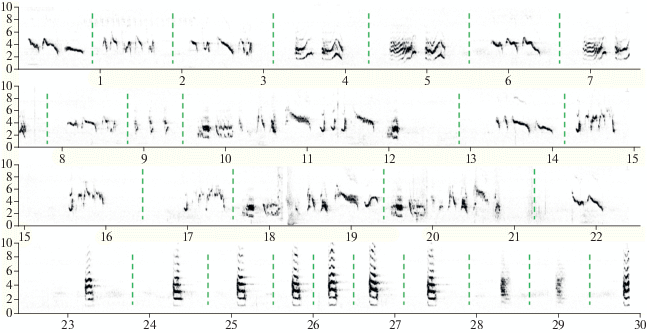
В качестве единицы анализа мы использовали слоги – стереотипные сочетания из нескольких звуков. Одиночные звуки, не образующие устойчивых сочетаний с другими, мы, по примеру других исследователей, также считали слогами (Catchpole, Slater, 2008). Все слоги делили на две группы: позывки и песенные слоги. Позывки состоят из одного звука (реже двух звуков) каждая и исполняются обычно в составе гомотипических серий (т.е. серий из одного типа слога). Песенные слоги чаще включают несколько звуков и встречаются в виде акустической последовательности из разных типов слогов (рис. 1). В изученных фонограммах мы выделили и проанализировали 1029 позывок и 1328 песенных слогов (т.е. всего 2357 слогов).
Рис. 1.
Непрерывный фрагмент рекламной вокализации длиннохвостого сорокопута. Сначала самец пел в течение около 22 с (первые три строки иллюстрируют этот эпизод пения), а затем перешел к исполнению позывок (в данном случае двух разных типов: четверная строка). Вертикальные линии разделяют разные типы слогов при пении и разные позывки.
На первом этапе мы составили каталог типов слогов. Каждый тип был обозначен цифрой, далее все фонограммы были представлены в виде последовательности цифр. По этой последовательности или ее частям в программе Past3 строили матрицы переходных вероятностей. В данном случае последовательность слогов рассматривалась как Марковская цепь первого порядка – т.е. такая, где событие n + 1 зависит только от события n. По матрицам переходных вероятностей рассчитывали относительную энтропию первого порядка RE1 = E1/E0 (Briefer et al., 2010). Для расчета E1 (энтропия первого порядка) и E0 (энтропия нулевого порядка) использовали формулу Шеннона:
где К – размер репертуара (количество типов слогов); где Pi – наблюдаемая вероятность каждого типа переходов между разными типами слогов.RE1 варьирует от 0 до 1. Этот параметр можно использовать как показатель “свободы выбора” самцом каждого следующего слога. Например, если RE1 = 0.70, можно считать, что самец имеет 70% такой “свободы” (Briefer et al., 2010). Или, по-другому, – последовательность слогов на 70% случайна.
Дальнейшая обработка проведена в среде программирования R 3.3.2 (R Core Team, 2016). Для визуализации наблюдаемых переходов между разными типами слогов использовали пакет ‘markovchain’ (Spedicato et al., 2017). Для выявления кластеров взаимно-ассоциированных слогов использовали методы теории графов в пакете ‘igraph’ (Csardi, Nepusz, 2006).
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Рекламная вокализация изученного самца длиннохвостого сорокопута включала два последовательно сменяющих друг друга (чередующихся) типа вокализаций: позывки и пение (рис. 1). Например, самец сначала издавал серию позывок, затем переходил к серии песенных слогов (пению), далее вновь возвращался к позывкам и т.д. Непрерывную последовательность песенных слогов мы далее будет называть эпизодом пения (рис. 1). Длительности каждой серии позывок составила 3–282 с (медиана 52 с, n = 18), а длительность каждого эпизода пения – 6–202 с (медиана 67 с, n = 21).
Репертуар позывок включал 13 типов (на рис. 1, в качестве примера, приведены только два типа). В составе одной непрерывной серии позывок все типы не использовались – максимально до 5 типов. Большинство типов позывок включали только один звук (как на рис. 1), но 4 типа из 13 включали по два звука каждый.
Всего изучено 25 эпизодов пения. Суммарно в них было 1328 песенных слогов, относящихся к 143 разным типам. Количество типов звуков в данном типе слога варьировало от 1 до 12, медиана 2 (n = 143). Известно, что длиннохвостый сорокопут охотно включает в свое пение имитации голосов других птиц (Rabor, 1936; Панов, 2008). Мы специально не занимались этим вопросом, но, по первому впечатлению, некоторые из выделенных 143 типов слогов птица могла позаимствовать у своих соседей. Так, некоторые слоги фокального самца, по нашему впечатлению, напоминали фрагменты вокализаций чернолобого сорокопута, индийской иволги (Oriolus kundoo), майны (Acridotheres tristis), воробьев Passer sp., деревенской ласточки (Hirundo rustica) и южной бормотушки (Iduna rama).
При пении песенные слоги объединялись в композиции (кластеры слогов). Мы выделили 11 типов композиций: сначала “вручную”, а затем подтвердив это методом теории графов. Каждая композиция объединяла 3–33 (медиана 8, n = 11) типов слогов. Слоги одной композиции взаимно-ассоциированы во времени. Приступая к очередной композиции, самец исполняет относящиеся к ней слоги, все или большинство. А затем переходит либо к следующей композиции, либо к секвенции позывок, либо замолкает. Разные типы песенных слогов в составе данной композиции чередуются не случайно, хотя порядок их следования определен не жестко. Так, значения энтропии RE1, описывающие чередование слогов, варьировали в разных композициях от 0.22 до 0.54 (медиана 0.30, n = 11). Кроме того, внутри каждой композиции можно было выделить более мелкие кластеры взаимно-ассоциированных слогов. Конкретные примеры можно найти на рис. 2.
Рис. 2.
Чередование разных типов песенных слогов в двух композициях длиннохвостого сорокопута, показанное в виде Марковской цепи первого порядка (вверху, приведены наблюдаемые переходные вероятности) и выявленное методом теории графов (внизу). Цифры соответствуют номерам типов слогов. Для каждой композиции приведены значения относительной энтропии RE1.
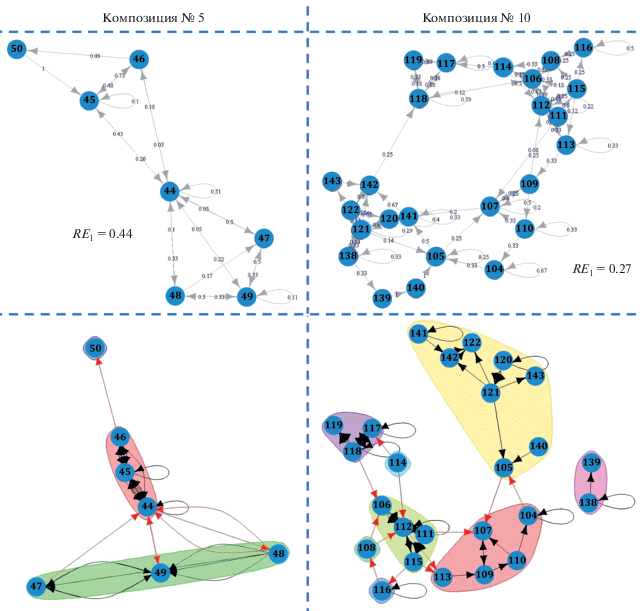
В каждом эпизоде пения было 1–6 разных композиций (медиана 2, n = 25). Удалось выявить некоторые закономерности в чередовании композиций разных типов (рис. 3), но таких правил было немного. Именно, после некоторых композиций с высокой вероятностью следовала другая, определенного типа. Переходы между такими связанными композициями показаны красными стрелками на рис. 3. Свобода выбора каждой следующей композиции была больше, нежели свобода при подборе последовательных песенных слогов в составе композиции. Действительно, значение энтропии RE1, характеризующее чередование разных композиций в эпизоде пения, оказалось равным 0.41. Это несколько больше, чем RE1 для чередования разных типов песенных слогов внутри данной композиции (медиана RE1 составила в этом случае 0.30, см. выше).
Рис. 3.
Чередование разных типов композиций (обозначены номерами), а также их взаимосвязи с секвенциями позывок (П) и началом/концом фонограммы (К) у длиннохвостого сорокопута. Красными стрелками выделены переходы, наблюдавшиеся более чем в 50% случаев.
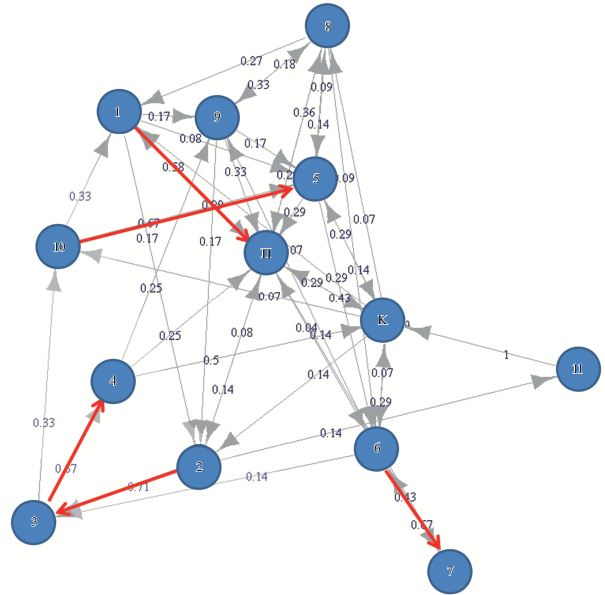
Таким образом, пение длиннохвостого сорокопута может быть устроено довольно сложно. Эта сложность выражается, во-первых, в обширном репертуаре слогов. А во-вторых, в некоторой упорядоченности следования разных акустических элементов (песенных слогов и композиций) друг за другом. Стоит отметить способность фокального самца к запоминанию и многократному исполнению протяженных акустических конструкций. Самая длинная из них включала последовательно исполняемые композиции № 2, № 3 и № 4 (рис. 3). Такую супер-композицию, состоящую из трех композиций, самец повторил 4 раза в разных эпизодах пения. Длительность супер-композиции составила 41–63 с (медиана 59 с, n = 4), она включала 69–108 слогов (медиана 89.5 слогов, n = 4). А суммарное количество типов слогов в композициях № 2, № 3 и № 4 – 36.
В заключение отметим, что полученные данные также прекрасно иллюстрируют отсутствие четкой границы между линейным и комбинаторным синтаксисами. Действительно, композиции и супер-композиции, хотя и исполняются в основном по принципу комбинаторного синтаксиса, содержат много элементов синтаксиса линейного.
Список литературы
Иваницкий В.В., 2015. Песня птиц как микрокосм современной науки: между биоакустикой и лингвистикой // XIV орнитологическая конференция Северной Евразии. II. Доклады. Алматы. С. 418–435.
Опаев А.С., 2012. Поведение и вокализация серого сорокопута Lanius excubitor на ранних стадиях гнездового цикла // Труды Окского государственного биосферного заповедника. Вып. 27. Рязань: НП “Голос губернии”. С. 17–34.
Опаев А.С., 2013. Поведение и вокализация клинохвостого сорокопута (Lanius sphenocercus Cabanis, 1873) на ранних стадиях гнездового цикла // Поволжский экологический журнал. № 3. С. 271–279.
Панов Е.Н., 2008. Сорокопуты (семейство Laniidae) мировой фауны. Экология, поведения, эволюция. М.: Товарищество научных изданий КМК. 650 с.
Briefer E., Osiejuk T., Rybak F., Aubin T., 2010. Are bird song complexity and song sharing shaped by habitat structure? An information theory and statistical approach // Journal of Theoretical Biology. V. 262. P. 151–164.
Catchpole C.K., Slater P.J.B., 2008. Bird song: biological themes and variations. Cambridge: Cambridge University Press. 335 p.
Cody M.L., Stabler E., Castellanos H.M.S., Taylor C.E., 2016. Structure, syntax and “small-world” organization in the complex song of California Thrashers (Toxostoma redivivum) // Bioacoustics. V. 25. P. 41–54.
Csardi G., Nepusz T., 2006. The igraph software package for complex network research // Complex Systems. V. 1695. P. 1–9.
del Hoyo J., Collar N.J., 2016. HBW and BirdLife International illustrated checklist of the birds of the world. V. 2. Passerines. Barcelona: Lynx Edicions. 1013 p.
Ivanitskii V.V., Marova I.M., Malykh I.M., 2012. Between order and chaos: contrasting syntax in the advertising song of dusky (Phylloscopus fuscatus) and Radde’s (Ph. schwarzi) warblers // Journal of Ornithology. V. 153. P. 337–346.
Opaev A., 2016. Relationships between repertoire size and organization of song bouts in the Grey-crowned Warbler (Seicercus tephrocephalus) // Journal of Ornithology. V. 157. P. 949–960.
R Core Team, 2016. R: A language and environment for statistical computing. Vienna: R Foundation for Statistical Computing. https://www.r-project.org
Rabor D.S., 1936. Life histories of some birds in the vicinity of Novaliches, Rizal Province, Luzon. 1. The Large-nosed Shrike, L. schach nasutus // Phillipine Journal of Science. V. 59. P. 337–355.
Sasahara K., Cody M.L., Cohen D., Taylor C.E., 2012. Structural design principles of complex bird songs: a network-based approach. PLoS ONE. V. 7. e44436
Spedicato G.A., Kang T.S., Yalamanchi S.B., Thoralf M., Yadav D., et al., 2017. Easy handling discrete time Markov chains. https://cran.r-project.org/web/packages/markovchain
Weiss M., Hultsch H., Adam I., Scharff C., Kipper S., 2014. The use of network analysis to study complex animal communication systems: a study on nightingale song // Proceedings of the Royal Society B. V. 281. e20140460
Дополнительные материалы отсутствуют.
Инструменты
Зоологический журнал