

Зоологический журнал, 2019, T. 98, № 8, стр. 933-948
Ареалы млекопитающих на Урале в конце позднего плейстоцена
О. П. Бачура a, *, П. А. Косинцев a, **
a Институт экологии растений и животных УрО РАН
620144 Екатеринбург, Россия
* E-mail: olga@ipae.uran.ru
** E-mail: kpa@ipae.uran.ru
Поступила в редакцию 16.08.2018
После доработки 22.11.2018
Принята к публикации 24.01.2019
Аннотация
На основании данных из многочисленных местонахождений ископаемых фаун описаны динамика ареалов и период обитания млекопитающих на Урале в конце позднего плейстоцена. Выделено 4 типа ареалов: постоянный, расширявшийся, сокращавшийся и флуктуирующий. Постоянный ареал имели эвритопные виды, ряд степных, тундровые виды, а также представители мамонтового комплекса. Расширение ареалов в период постепенного потепления климата в течение позднеледниковья и перехода к голоцену происходило преимущественно у видов, связанных с древесно-кустарниковой растительностью или водными биотопами. Сокращение ареалов при переходе от интерстадиала к максимально холодной стадии позднего плейстоцена к югу или в результате вымирания произошло у лесостепных, степных и полупустынных видов. Группа млекопитающих с флуктуирующими ареалами включает виды, связанные с древесно-кустарниковой растительностью, один околоводный вид и один степной. Северная граница их ареалов в максимально холодную стадию позднего плейстоцена смещалась к югу, а в период позднеледниковая их ареалы восстановились в прежних границах или продвигались севернее.
Современное распространение млекопитающих – это итог непрерывного процесса эволюции фаун. Конкретное современное географическое распределение видов животных определяется сложившимися параметрами окружающего их пространства, географической изменчивостью, а также во многом историей формирования их ареалов, спецификой образования фаунистических комплексов. Следовательно, для полноценного объяснения современных ареалов млекопитающих необходимы результаты историко-фаунистического анализа.
Основной этап формирования современных ареалов млекопитающих происходил в течение последнего межледникового периода – голоцена. Но начался этот процесс еще в позднем плейстоцене, для которого характерны значительные изменения природной среды. Этот период в истории распространения млекопитающих интересен еще и тем, что в позднем плейстоцене в Северном полушарии господствовали особые природные условия, аналогов которым в настоящее время нет (Маркова и др., 2008).В этих особых условиях существовали виды млекопитающих, обитающих в настоящее время в разных природных зонах.
Территория Урала является одним из удобных регионов для исследований, посвященных изучению ареалов животных в прошлом. Это обусловлено большой протяженностью Уральских гор с севера на юг, что создает хорошо выраженный широтный градиент природно-климатических условий (Урал и Приуралье, 1968). На всем протяжении Уральских гор распространены карстующиеся породы с многочисленными карстовыми полостями (Лавров, Андрейчук, 1992), вмещающие местонахождения ископаемых остатков млекопитающих. Кроме того, для отдельных видов млекопитающих получены серии радиоуглеродных дат, которые позволяют установить точное время их распространения в позднем плейстоцене на Урале. Все это обеспечивает большое количество источников фактического материала для изучения истории ареалов млекопитающих в прошлом.
Изменение ареалов ряда видов для отдельных регионов Урала в позднем плейстоцене уже рассматривалось ранее (Кузьмина, 1971; Смирнов, 1993; 1996). Накопленные в настоящее время новые материалы из многочисленных местонахождений позволяют рассмотреть динамику ареалов большинства видов млекопитающих в конце позднего плейстоцена для всей территории Урала (Кузьмина, 1971;Смирнов и др., 1990; Смирнов, 1993; 1995; 1996; Кузьмина и др., 1999; Пономарев, 2001; Bachura, Kosintsev, 2007; Фадеева, Смирнов, 2008; Косинцев, Пластеева, 2009; Kuzmina, 2009; Teterina, 2009; Ponomarev et al., 2012; Kosintsev, Bachura, 2013).
Цель работы: изучить характер изменения ареалов и, по возможности, границы распространения для разных видов млекопитающих на Урале в конце позднего плейстоцена.
МАТЕРИАЛ И МЕТОДЫ
Традиционно Урал делят на четыре области: Полярный (от 67°30′ с.ш. до 64°00′ с.ш.), Северный (от 64°00′ с.ш. до 59°15′ с.ш.), Средний (от 59°15′ с.ш. до 55°54′ с.ш.) и Южный (от 55°54′ с.ш. до 51°00′ с.ш.) (Урал и Приуралье, 1968). Хотя это деление основано на орографическом строении Уральского хребта, вместе с тем каждая из этих областей имеет свои природно-климатические особенности. В этой работе мы рассматриваем Полярный, Северный, Средний и Южный Урал в широком плане, включая горную часть и районы Предуралья и Зауралья.
В настоящей статье рассматриваются представители отрядов Lagomorpha, Rodentia, Carnivora, Perissodactyla и Artiodactyla. Данные о представителях отрядов Insectivora и Chiroptera не включены в работу, т.к. в публикациях чаще всего не приведены видовые определения костных остатков представителей этих отрядов.
Остатки мелких и крупных млекопитающих во всех ископаемых фаунах происходят из зоогенных отложений карстовых полостей или древних присад хищных птиц (Кузьмина, 1971; Смирнов, 1993; 1995; 1996; Косинцев, 1996; Кузьмина и др., 1999; Пономарев, 2001; Косинцев, Орлова, 2002; Тетерина, 2002; Кузьмина, 2003; Ражев и др., 2005; Изварин, 2006; Струкова и др., 2006; Косинцев, Крахмальная, 2008; Фадеева, Смирнов, 2008; Yakovlev et al., 2006; Danukalova et al., 2008; 2011; Golovachev, Smirnov, 2009; Kuzmina, 2009; Ponomarev et al., 2012; 2013). Наряду с опубликованными данными в работе использованы и новые данные. В анализ включены ископаемые фауны, временные рамки существования которых подтверждены радиоуглеродными датами (более 200 дат). Работа выполнена на основании данных из 56 локальных фаун мелких млекопитающих (Смирнов и др., 1990; Смирнов и др., 1992; Смирнов, 1993; 1995; 1996; Яковлев, 1997; Бородин др., 2000; Струкова, 2000; Кузьмина и др., 2001; Бачура, Струкова, 2002; Струкова, 2002; Тетерина, 2002; Кузьмина, 2003; Яковлев и др., 2005; Фадеева, Смирнов, 2008; Изварин, 2004; 2009; Kuzmina, 2009; Golovachev, Smirnov, 2009; Danukalova et al., 2008; 2011; Кряжева и др., 2012; Ponomarev et al., 2013; Кряжева, Пономарев, 2014) и 61 локальных фаун крупных млекопитающих (Смирнов и др., 1990; Кузьмина, 1971; 1975; Кузьмина и др., 1999; Щербакова, 1994; Косинцев, 1995; 1996; Подопригора, 1999; Бородин и др., 2000; Бородин и др., 2000а; Косинцев, Воробьев, 2000; Кузьмина, 2000; Бачура, Струкова, 2002; Косинцев, Орлова, 2002; Ражев и др., 2005; Косинцев, Крахмальная, 2008;). Так же использованы данные по отдельным находкам костей млекопитающих, датированных радиоуглеродным методом.
Коллекции ископаемых костных остатков из местонахождений, которые были использованы в работе, хранятся в Музее ИЭРиЖ УрО РАН, г. Екатеринбург.
В рамках данной статьи рассматривается период конца позднего плейстоцена – 50–10.2 тыс. лет назад. Анализ данных проводился по нескольким хронологическим периодам:
1. Невьянский интерстадиал (средневалдайский интерстадиал, MIS 3) – 58–25 тыс. лет назад;
2. Максимально холодная стадия позднего плейстоцена (LGM, early MIS2) – 25–16.5 тыс. лет назад;
3. Начало позднеледниковья (LGT; early MIS2) – 16.5–12.4 тыс. лет назад;
4. Конец позднеледниковья (late MIS2) – 12.4 – 10.2 тыс. лет назад.
К сожалению, материалы по всем хронологическим периодам на настоящий момент имеются не для всей территории Урала. Наиболее полно данные хронопериоды представлены на территории Среднего Урала. Для Полярного Урала на настоящий момент для мелких млекопитающих имеются датированные материалы только конца позднеледниковья, а для крупных млекопитающих немногочисленные материалы невьянского интерстадиала. На Северном Урале отсутствуют данные о фауне конца позднеледниковья. На Южном Урале для крупных млекопитающих имеются данные по всем хронопериодам, а для мелких млекопитающих – от максимально холодной стадии позднего плейстоцена до конца позднеледниковья. Таким образом, в ряде случаев могут быть восстановлены лишь предположительные границы распространения млекопитающих в те или иные периоды конца позднего плейстоцена.
Список видов изменился по сравнению со списком, который был опубликован ранее, в связи с полученными за последнее время новыми данными. В результате изучения палео-ДНК установлено, что донской заяц (Lepus tanaiticus Gureev 1964), который считался типичным видом позднего плейстоцена Восточной Европы и Сибири, является экоморфой зайца-беляка (L. timidus L. 1758) (Prost et al., 2010). Ранее предполагалось существование на Южном Урале в позднем плейстоцене двух видов лошадей – Equus (Equus) latipes Gromova 1949 и E. (E.) uralensis Kuzmina 1975 (Смирнов и др., 1990). Проведенный анализ показал, что это подвиды одного вида E. ferus Boddaert 1785. (Косинцев, Пластеева, 2009).
Вид Microtus arvalis Pallas 1779 рассматривается как обыкновенная полевка в широком смысле (sensu lato) (Markova et al., 2012). Копытный лемминг (Dicrostonyx sp.) и лесные мыши рода Apodemus рассматриваются нами на уровне родов, т.к. во многих источниках, которые были использованы для написания данной статьи, не указана их видовая принадлежность.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
В результате сравнительного анализа региональных таксономических списков млекопитающих было выделено несколько групп видов, которые различались по характеру динамики ареалов в течение позднего плейстоцена.
Группа I – современные виды млекопитающих, границы ареалов которых были постоянными в течение всего позднего плейстоцена (табл. 1). Ареалы зайца-беляка, степной пищухи (Ochotona pusilla Pallas 1768), копытного лемминга, сибирского лемминга (Lemmus sibiricus Kerr 1792), узкочерепной полевки (Lasiopodomys gregalis Pallas 1779), волка (Canis lupus L. 1758), песца (Vulpes lagopus L. 1758), северного оленя (Rangifer tarandus L. 1758) охватывали всю территорию Урала. Ареалы лисицы (Vulpes vulpes L. 1758), росомахи (Gulo gulo L. 1758), соболя (Martes zibellina L. 1758), горностая (Mustela erminea L. 1758), ласки (M. nivalis L. 1766), светлого хоря (M. eversmanni Lesson 1827) и сайги (Saiga tatarica L. 1766) на юге занимали всю территорию Южного Урала, а на севере – всю территорию Северного Урала. Сайга в Восточной Сибири в позднем плейстоцене была распространена на север вплоть до 73° с.ш. (Орлова, 1995). Вероятно, на Урале этот вид мог доходить до тех же широт. Северную границу распространения остальных видов на настоящий момент установить невозможно, т.к. находок позднего плейстоцена этих видов на Полярном Урале неизвестно.
Таблица 1.
Ареалы млекопитающих на Урале в конце позднего плейстоцена
| Вид | Предпочитаемые местообитания | Ареал | Полярный | Северный | Средний | Южный | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| периоды позднего плейстоцена | ||||||||||||||||||
| 1 | 2 | 3 | 4 | 1 | 2 | 3 | 4 | 1 | 2 | 3 | 4 | 1 | 2 | 3 | 4 | |||
| Dicrostonyx sp. | Тундровые | Постоянный | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + |
| Lemmus sibiricus | Постоянный | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | |
| Vulpes lagopus | Постоянный | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | |
| Lasiopodomys gregalis | Тундровые и степные | Постоянный | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + |
| Rangifer tarandus | Тундровые и таежные | Постоянный | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + |
| Alexandromys middendorffi | Постоянный | + | + | + | + | + | + | + | + | + | + | + | + | – | – | – | – | |
| Martes zibellina | Таежные | Постоянный | – | – | – | – | + | + | + | + | + | + | + | + | + | + | + | + |
| Equus ferus | Лесостепные и степные | Постоянный | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + |
| Mustela eversmanni | Постоянный | – | – | – | – | + | + | + | + | + | + | + | + | + | + | + | + | |
| Spermophilus major | Степные | Постоянный | – | – | – | – | + | + | + | + | + | + | + | + | + | + | + | + |
| Marmota bobak | Постоянный | – | – | – | – | + | + | + | + | + | + | + | + | + | + | + | + | |
| Ochotona pusilla | Постоянный | ? | ? | ? | + | + | + | + | + | + | + | + | + | + | + | + | + | |
| Cricetulus migratorius | Постоянный | – | – | – | – | + | + | + | + | + | + | + | + | + | + | + | + | |
| Lagurus lagurus | Постоянный | – | – | – | – | + | + | + | + | + | + | + | + | + | + | + | + | |
| Saiga tatarica | Степные и полупустынные | Постоянный | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + |
| Eolagurus luteus | Полупустынные и пустынные | Постоянный | – | – | – | – | – | – | – | – | + | + | + | + | + | + | + | + |
| Panthera spelaea | Перигляциальные степи позднего плейстоцена | Постоянный | + | + | + | + | + | + | + | + | + | + | + | + | ||||
| Mammuthus primigenius | Постоянный | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | |
| Coelodonta antiquitatis | Постоянный | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | |
| Bison priscus | Постоянный | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | |
| Ovibos moschatus | Перигляциальные степи позднего плейстоцена, современные тундры | Постоянный | + | + | + | + | + | + | + | + | + | + | + | + | – | – | – | – |
| Lepus timidus | Широкий спектр | Постоянный | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + |
| Canis lupus | Постоянный | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | |
| Vulpes vulpes | Постоянный | ? | ? | ? | ? | + | + | + | + | + | + | + | + | + | + | + | + | |
| Gulo gulo | Постоянный | ? | ? | ? | ? | + | + | + | + | + | + | + | + | + | + | + | + | |
| Mustela erminea | Постоянный | ? | ? | ? | ? | + | + | + | + | + | + | + | + | + | + | + | + | |
| M. nivalis | Постоянный | ? | ? | ? | ? | + | + | + | + | + | + | + | + | + | + | + | + | |
| M. agrestis | Таежные | Расширялся | – | – | – | + | – | – | – | + | + | + | + | + | + | + | + | + |
| Sicista betulina | Лесные | Расширялся | – | – | – | – | – | – | – | – | – | – | + | + | – | – | + | + |
| Apodemus sp. | Лесные | Расширялся | – | – | – | – | – | – | – | – | – | – | – | + | – | – | – | + |
| M. arvalis | Лесостепные | Расширялся | – | – | – | – | – | – | – | – | + | + | + | + | + | + | + | + |
| Meles meles | Расширялся | – | – | – | – | – | – | – | – | – | – | – | + | – | – | – | ? | |
| Alexandromys oeconomus | Влажные и околоводные биотопы в лесной зоне | Расширялся | – | – | – | + | + | + | + | + | + | + | + | + | + | + | + | + |
| Arvicola amphibius | Околоводные | Расширялся | – | – | – | + | – | – | – | + | + | + | + | + | + | + | + | + |
| Myopus shisticolor | Таежно-болотные | Расширялся | – | – | – | + | – | + | + | + | + | + | + | + | – | – | – | – |
| Allactaga jaculus | Степные | Расширялся | – | – | – | – | – | – | – | – | – | – | – | – | – | + | + | + |
| Ellobius talpinus | Степные и полупустынные | Расширялся | – | – | – | – | – | – | – | – | – | – | – | – | – | + | + | + |
| Ursus spelaeus | Перигляциальные степи позднего плейстоцена | Сократился | – | – | – | – | + | – | – | – | + | – | – | – | + | – | – | – |
| Ursus savini | Сократился | – | – | – | – | – | – | – | – | + | – | – | – | + | – | – | – | |
| Crocuta c. spelaea | Сократился | – | – | – | – | – | – | – | – | + | – | – | – | + | – | – | – | |
| Cervus elaphus | Лесостепные | Сместился | – | – | – | – | + | – | – | – | – | – | – | – | + | + | + | – |
| Ovis ammon | Степные | Сократился | – | – | – | – | – | – | – | – | – | – | – | – | + | – | – | – |
| Allocricetulus eversmanni | Степные и полупустынные | Сократился | – | – | – | – | – | – | – | – | ? | + | + | – | + | + | + | – |
| Pygeretmus pumilio | Пустынные и полупустынные | Сместился | – | – | – | – | – | – | – | – | – | – | – | – | ? | + | + | – |
| Camelus ferus | Полупустынные и пустынные | Сократился | – | – | – | – | – | – | – | – | – | – | – | – | + | – | – | – |
| Craseomys rufocanus | Лесные | Флуктуирующий | – | – | – | + | + | – | + | + | + | + | + | + | + | + | + | + |
| Myodes rutilus ex. gr. glareolus | Флуктуирующий | – | – | – | + | + | – | + | + | + | + | + | + | + | + | + | + | |
| Alces alces | Флуктуирующий | – | – | – | – | – | – | – | + | + | – | + | + | + | + | + | + | |
| Ursus arctos | Таежные | Флуктуирующий | – | – | – | – | + | – | + | + | + | + | + | + | + | + | + | + |
| Lynx lynx | Флуктуирующий | – | – | – | – | – | – | – | – | – | – | + | + | – | – | – | ? | |
| Megaloceros giganteus | Лесостепные | Флуктуирующий | – | – | – | – | – | – | – | – | – | – | + | + | – | – | + | + |
| Cricetus cricetus | Луговые и лесостепные | Флуктуирующий | – | – | – | – | – | – | – | – | – | – | – | + | + | – | + | + |
| Castor fiber | Околоводные | Флуктуирующий | – | – | – | – | – | – | – | – | – | – | – | + | + | – | – | + |
| Vulpes corsac | Степные и полупустынные | Флуктуирующий | – | – | – | – | – | – | – | – | + | – | – | – | + | + | + | + |
Периоды позднего плейстоцена: 1– невьянский интерстадиал, 2 – максимально холодная стадия позднего плейстоцена, 3 – начало позднеледниковья, 4 – конец позднеледниковья. + – вид обитал в данный период; прочерк – вид не обитал в данный период; ? – вид мог обитать в этот период, но его остатки не найдены.
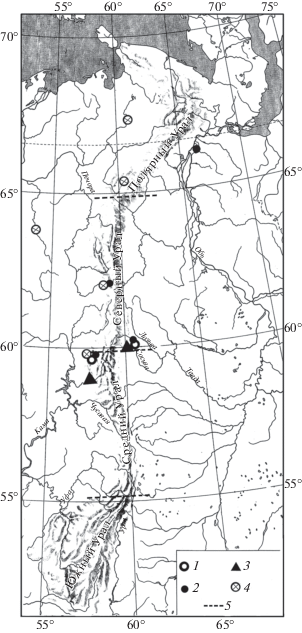
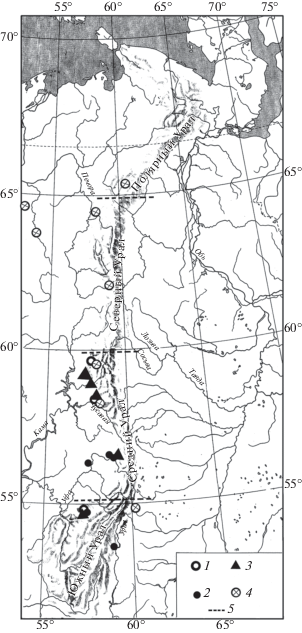
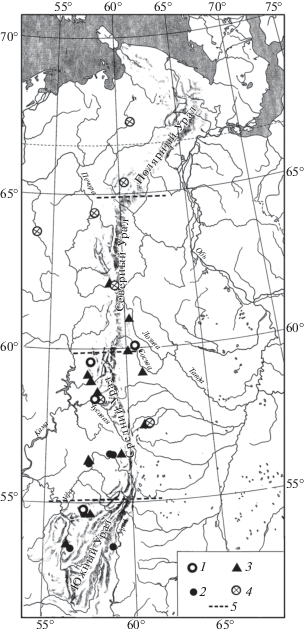
Находки серого хомячка (Cricetulus migratorius Pallas 1773), большого суслика (Spermophilus major Pallas 1779) (рис. 1), сурка (Marmota bobak Müller 1776) (рис. 2), степной пеструшки (Lagurus lagurus Pallas 1773) (рис. 3) известны в локальных фаунах, которые расположены к югу от 61° с.ш., а находки желтой пеструшки (Eolagurus luteus Eversmann 1840) к югу от 57° с.ш. (рис. 4). Вероятно, эти широты являются северными границами ареалов этих видов. Все находки полевки Миддендорфа (Alexandromys middendorffi Poljakov 1881) на Урале в конце позднего плейстоцена известны в локальных фаунах к северу от 58° с.ш. до 67° с.ш. (рис. 5).
Рис. 1.
Распространение Cricetulus migratorius и Spermophilus major на Урале в конце позднего плейстоцена: 1 – местонахождения невьянского интерстадиала, 2 – местонахождения максимально холодной стадии, 3 – местонахождения начала позднеледниковья, 4 – местонахождения конца позднеледниковья, 5 – граница областей Урала.
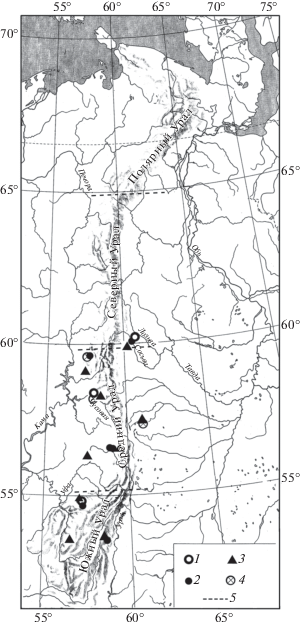
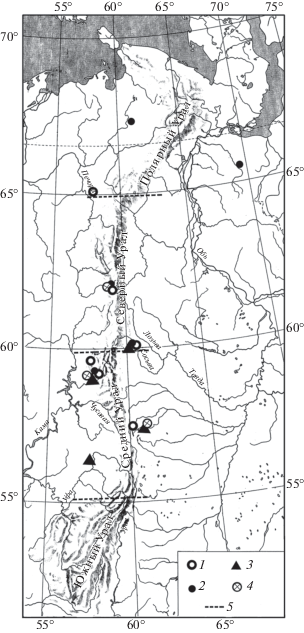
Рис. 2.
Распространение Marmota bobak на Урале в конце позднего плейстоцена. Обозначения как на рис. 1.
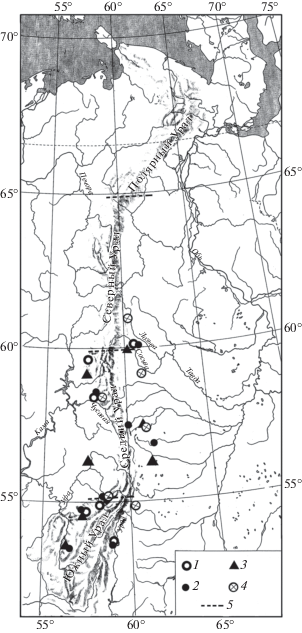
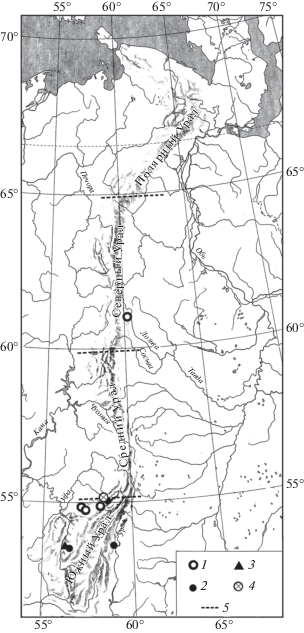
Рис. 3.
Распространение Lagurus lagurus на Урале в конце позднего плейстоцена. Обозначения как на рис. 1.
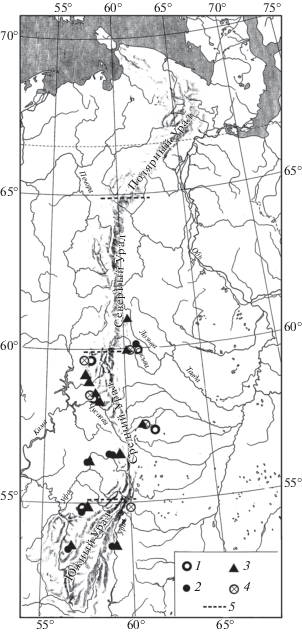
Рис. 4.
Распространение Eolagurus luteus на Урале в конце позднего плейстоцена. Обозначения как на рис. 1.
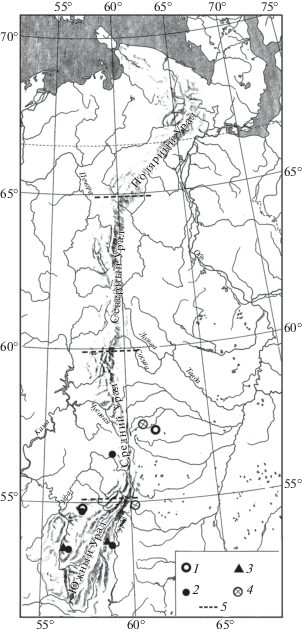
Рис. 5.
Распространение Alexandromys middendorffi на Урале в конце позднего плейстоцена. Обозначения как на рис. 1.
Группа II объединяет вымершие виды, ареалы которых были постоянными на территории Урала в течение всего конца позднего плейстоцена.
Ареалы пещерного льва (Panthera spelaea Goldfuss 1810), мамонта (Mammuthus primigenius Blumenbach 1799), шерстистого носорога (Coelodonta antiquitatis Blumenbach 1799), плейстоценовой лошади (Equus ferus Boddaert 1785), первобытного бизона (Bison priscus Bojanus 1827) занимали всю территории Урала в течение всего рассматриваемого периода. Овцебык (Ovibos moschatus Zimmermann 1780) обитал на Полярном, Северном и Среднем Урале. Южная граница ареала овцебыка в позднем плейстоцене проходила, вероятно, между 56–57° с.ш. Южнее находок этого вида неизвестно (рис. 6). Вымирание этих видов на территории Урала происходило в разное время и с разной скоростью, но в рамках данной статьи мы не будем касаться этого вопроса.
Рис. 6.
Распространение Ovibos moschatus на Урале в конце позднего плейстоцена. Обозначения как на рис. 1.
Группа III объединяет виды, у которых происходило сокращение ареала на Урале в течение плейстоцена.
Ареал тарбаганчика (Pygeretmus pumilio Kerr 1792) занимал территорию к югу от 55° с.ш. в период максимально холодной стадии позднего плейстоцена до середины позднеледниковья. В конце позднеледниковья этот вид сократил свой ареал и больше не входил в состав фауны Урала.
Хомячок Эверсманна (Allocricetulus eversmanni Brandt 1859) обитал к югу от 57° с.ш., вероятно, до середины позднеледниковья. К концу этого периода ареал хомячка Эверсманна сокращается, и он исчезает из фауны Урала.
Плейстоценовых находок костей благородного оленя (Cervus elaphus L. 1758) на Урале очень мало. Часть из них имеет радиоуглеродные даты. На Южном Урале благородный олень обитал на протяжении всего рассматриваемого периода позднего плейстоцена. В более ранние периоды позднего плейстоцена 43–47 тыс. лет назад ареал благородного оленя доходил до Северного Урала, вероятно, до широты 61° с.ш. К началу невьянского интерстадиала ареал благородного оленя смещается на Южный Урал, где северная граница ареала проходила между 55–56° с.ш. (рис. 7). Расширение ареала благородного оленя вновь к северу началось в начале раннего голоцена.
Рис. 7.
Распространение Cervus elaphus на Урале в конце позднего плейстоцена. Обозначения как на рис. 1.
Малый пещерный медведь (Ursus savini Andrews 1922) постоянно обитал на Среднем и Южном Урале вплоть до 59° с.ш. в течение первой половины позднего плейстоцена до середины невьянского интерстадиала. По остаткам с Урала получены наиболее поздние радиоуглеродные даты (Paсher, Stuart, 2009), которые указывают, что этот вид вымирает здесь в конце невьянского интерстадиала (рис. 8).
Рис. 8.
Находки остатков млекопитающих на Урале в конце позднего плейстоцена: 1 – Ursus spelaeus, 2 – U. savini, 3 – Crocuta c. spelaea, 4 – Camelus ferus и 5 – Ovis ammon, 6 – граница областей Урала.
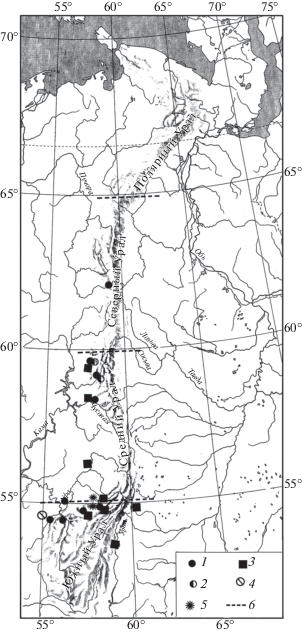
Большой пещерный медведь (U. spelaeus Rosenmüller 1794) постоянно обитал на западном склоне Северного, Среднего и Южного Урала в течение первой половины позднего плейстоцена. Около 40 тыс. лет назад ареал большого пещерного медведя постепенно сокращается и к концу невьянского интерстадиала вид исчезает из фауны Урала (рис. 8).
В эту же группу были отнесены несколько видов крупных млекопитающих, для которых известны лишь единичные находки для определенных периодов позднего плейстоцена. Строго говоря, отнесение этих видов в данную группу условное, т.к. остается неясным характер динамики их ареалов в начале позднего плейстоцена. Это такие виды как пещерная гиена (Crocuta crocuta spelaea Goldfuss 1823), архар (Ovis ammon L. 1758), верблюд (Camelus ferus Przewalski 1883). Ареал гиены до 40 тыс. лет назад охватывал территорию Южного Урала и западный склон Среднего Урала до 58° с.ш. Около 38 тыс. лет назад ареал пещерной гиены смещается на Южный Урал, и северная граница уже проходила примерно по 54° с.ш. (рис. 8). Около 35 тыс. лет назад восточная граница ареала гиены проходила уже по территории современной Западной Европы и юга Восточной Европы, где этот вид дожил до 25–26 тыс. лет назад (Stuart, Lister, 2014).
Остатки архара известны из отложений пещеры Усть-Катавская, которые датируются по кости пещерной гиены 38 тыс. лет назад (Kosintsev, Bachura, 2013). Так же остатки этого вида найдены в двух других местонахождениях Сикаяз-Тамак-1 и Сикаяз-Тамак-7 (Данукалова и др., 2018). Отложения из местонахождения Сикияз-Тамак-7, где была найдена кость архара, датируются первой половиной позднего плейстоцена (первая половина МИС 3 – МИС 5 a–d). (Данукалова и др., 2018). Для отложений из местонахождения Сикияз-Тамак-1 радиоуглеродных дат нет. Предположительно, эти отложения датируются невьянским интерстадиалом. Вероятно, северная граница ареала архара на Урале в период невьянского интерстадиала проходила по 55° с.ш. (рис. 8). Позднее этот вид не обитал на Урале.
Период обитания верблюда в предгорной части Южного Урала на настоящий момент не совсем понятен. Остатки этого вида найдены в местонахождении Горново в Башкирии (Danukalova et al., 2016) в отложениях невьянского интерстадиала (рис. 8). Вероятно, верблюд обитал в предгорной части Южного Урала до 54° с.ш. в период невьянского интерстадиала. К началу максимально холодной стадии позднего плейстоцена этот вид исчезает с данной территории.
Группа IV объединяет виды млекопитающих, ареал которых расширялся в течение позднего плейстоцена.
Обыкновенная полевка в период максимально холодной стадии и, вероятно, невьянского интерстадиала позднего плейстоцена обитала на территории Южного Урала и юге Среднего Урала. Северная граница ареала обыкновенной полевки в эти периоды проходила, вероятно, между 56° с.ш. и 57° с.ш. В середине позднеледниковья (не ранее 13 тыс. лет назад) ареал обыкновенной полевки начинает расширяться к северу, и к концу этого периода она расселяется до 58° с.ш. Далее на север обыкновенная полевка в позднем плейстоцене не расселялась.
Полевка-экономка (Alexandromys oeconomus Pallas 1776) обитала на территории Северного, Среднего и Южного Урала в течение всего рассматриваемого периода. Расселение ее на Полярный Урал произошло, скорее всего, только в конце позднеледниковья.
Водяная (Arvicola amphibius L. 1758) и темная (M. agrestis L. 1761) полевки весь период в конце позднего плейстоцена обитали на Среднем и Южном Урале на север до 58°с.ш.. В конце позднеледниковья около 12 тыс. лет назад эти виды постепенно расселяются на Северный Урал до 62° с.ш., а затем, позднее, около 11 тыс. лет назад – на Полярный Урал до 67° с.ш. (рис. 9).
Рис. 9.
Распространение Arvicola amphibius на Урале в конце позднего плейстоцена. Обозначения как на рис. 1.
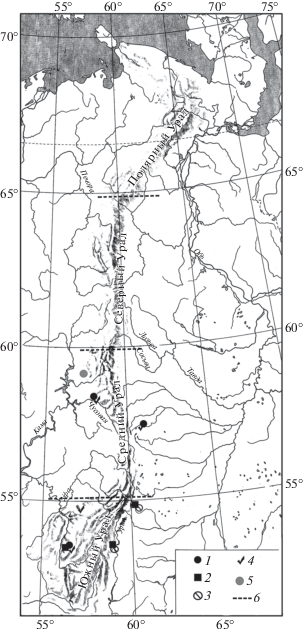
В конце позднеледниковья в южную часть Среднего Урала и на Южный Урал расселяются лесные мыши рода Apodemus (рис. 10). К концу позднего плейстоцена лесные мыши расселяются по всему Среднему Уралу. В конце максимально холодной стадии позднего плейстоцена на Южный Урал расселяется обыкновенная слепушонка (Ellobius talpinus Pallas 1770). Северная граница ареала лесных мышей проходила в позднеледниковье по 52° с.ш., а у слепушонки – по 55° с.ш. (рис. 10).
Рис. 10.
Находки остатков млекопитающих на Урале в конце позднего плейстоцена: 1 – Apodemus sp., 2 – Ellobius talpinus, 3 – Allactaga jaculus, 4 – Sicista betulina, 5 – Meles meles, 6 – граница областей Урала.
Находок лесного лемминга (Myopus schisticolor Lilljeborg 1844)в локальных фаунах позднего плейстоцена на Урале очень мало (рис. 12), но они охватывают весь период конца позднего плейстоцена на Среднем Урале. Вероятно, на Среднем Урале лесной лемминг обитал на протяжении всего конца позднего плейстоцена между 56° и 59° с.ш. В конце максимально холодной стадии позднего плейстоцена лесной лемминг расселяется к северу сначала на Северный Урал, а позднее в конце позднеледниковья и на Полярный Урал (рис. 11). Имеющиеся данные указывают на то, что расселение проходило по западному склону, т.к. на восточном склоне его находок нет (рис. 11). Расселение лесного лемминга на Южный Урал происходило уже в голоцене.
Рис. 11.
Распространение Myopus schisticolor на Урале в конце позднего плейстоцена. Обозначения как на рис. 1.
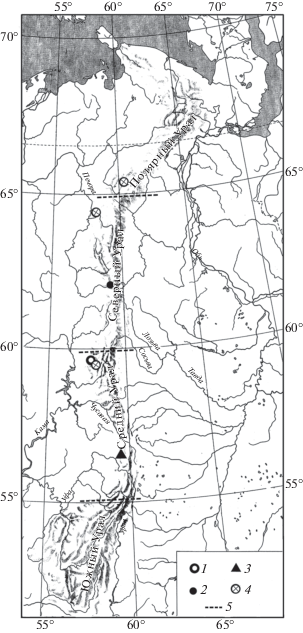
Рис. 12.
Распространение лесных полевок на Урале в конце позднего плейстоцена. Обозначения как на рис. 1.
В период максимально холодной стадии позднего плейстоцена в Южное Зауралье, а позднее, в начале позднеледниковья и в горную часть Южного Урала, расселяется большой тушканчик (Allactaga jaculus Pallas 1778). Северная граница его ареала не выходила за пределы 55° с.ш. (рис. 10).
Лесная мышовка (Sicista betulina Pallas 1778) появляется в горной части Южного Урала в начале позднеледниковья. Северная граница ее ареала в этот период проходила по 55° с.ш. В течение этого периода лесная мышовка расселяется дальше на север, и к концу позднеледниковья северная граница ее ареала проходила уже по 58° с.ш. (рис. 10). Дальнейшее расселение этого вида к северу по Уралу происходило уже в голоцене.
Европейский барсук (Meles meles L. 1758) в конце позднеледниковья, вероятно, не ранее 12 тыс. лет назад, расселяется на западный склон Среднего Урала до широты 58–59° с.ш. (рис. 10). Дальнейшее его расселение к северу и к югу происходило уже в голоцене.
Группа V объединяет виды с флуктуирующим ареалом. Лесные полевки: красно-серая (Craseomys rufocanus Sundervall 1846–1847), рыжая (Myodes glareolus Schreber 1780) и красная (M. rutilus Pallas 1779) обитали на территории Среднего и Южного и южной части Северного Урала в период невьянского интерстадиала, когда северная граница ареала этих видов проходила, вероятно, по 62° с.ш. К началу максимально холодной стадии позднего плейстоцена ареал этих видов смещается к югу, в южную часть Среднего Урала, и северная граница ареала проходила, вероятно, между 56–57° с.ш. Позднее, в начале позднеледниковья эти виды вновь постепенно расселяются к северу до 62° с.ш., а к концу позднеледниковья до 67° с.ш. (рис. 12).
Ареал бурого медведя (Ursus arctos L. 1758) весь рассматриваемый период занимал территорию Среднего и Южного Урала. На Северном Урале известны находки бурого медведя невьянского периода, а так же позднеледниковья. Вероятно, в период максимально холодной стадии позднего плейстоцена бурый медведь на Северном Урале не обитал.
Корсак (Vulpes corsac L. 1768) на протяжении всего позднего плейстоцена обитал на территории Южного Урала. В период невьянского интерстадиала ареал корсака расширяется к северу на Средний Урал до 57° с.ш. Позднее, в период максимально холодной стадии позднего плейстоцена ареал корсака восстанавливается в своих прежних границах.
На основании единичных находок хомяка (Cricetus cricetus L. 1758), бобра (Castor fiber L. 1758), рыси (Lynx lynx L. 1758), лося (Alces alces L. 1758) и гигантского оленя (Megaloceros giganteus Blumenbach 1803) могут быть восстановлены только предположительные ареалы.
Рысь обитала на территории Южного Урала до начала невьянского интерстадиала. К началу максимально холодной стадии ареал рыси сокращается, она исчезает с территории Урала. Вновь этот вид заселяет Южный и Средний Урал в начале позднеледниковья около 14–15 тыс. лет назад до 59° с.ш.
Хомяк в невьянский период обитал на территории Южного Зауралья, позднее его ареал сокращается, и вновь этот вид расселяется на Южный Урал в начале позднеледниковья, а к концу позднеледниковья расселяется на Средний Урал до 59° с.ш.
Лось в период невьянского интерстадиала был распространен на территории Среднего и Южного Урала до 59° с.ш. Позднее в период максимально холодной стадии позднего плейстоцена ареал лося отступает к югу на Южный Урал. Вероятно, в этот период северная граница ареала проходила по 55° с.ш. К началу позднеледниковья ареал вида вновь расширяется на Средний Урал до 59° с.ш., а в конце этого периода на Северный Урал до 62° с.ш. Согласно находкам расселение лося к северу в позднеледниковье происходило по западному склону.
Бобр в период невьянского интерстадиала обитал на территории Южного Урала до 54° с.ш. К началу максимально холодной стадии позднего плейстоцена бобр исчезает из состава фауны Урала. В конце позднеледниковья этот вид вновь расселяется на территорию Южного и Среднего Урала до 57–58° с.ш. Дальнейшее его расселение к северу происходило уже в голоцене. Гигантский олень обитал на Южном Урале в горной части и в Среднем и Южном Зауралье до начала невьянского интерстадиала. Северная граница его ареала проходила на широте 54°. Позднее ареал гигантского оленя сокращается, и олень исчезает из состава фауны региона. Вновь этот вид появляется на Среднем и Южном Урале в середине позднеледниковья, когда северная граница его ареала доходила до 57° с.ш.
Полученные данные показывают, что у 27 видов из 54 описанных ареал оставался постоянным в течение всего рассматриваемого периода. У 8 видов произошло сокращение ареала, еще 10 видов расширили свой ареал, а у 9 видов ареал колебался в течение позднего плейстоцена. По ландшафтно-биотопическим характеристикам среди видов с постоянным ареалом присутствуют эврибионтные, степные, тундровые и вымершие виды млекопитающих, обитавшие в перигляциальных ландшафтах позднего плейстоцена (табл. 1). Также в эту группу входит один вид – соболь, обитающий в настоящее время исключительно в таежной зоне (табл. 1). Очевидно, этот вид мог обитать в поймах рек, где на протяжении всего позднего плейстоцена сохранялась древесная растительность (Струкова и др., 2006; Маркова и др., 2008; Лаптева, 2009; Широков и др., 2011).
Виды, ареал которых в позднем плейстоцене сократился, имеют различные экологические характеристики (табл. 1). У большей части видов этой группы сокращение ареала произошло в конце невьянского интерстадиала, после которого последовал период с экстремально холодными климатическими условиями. Сокращение ареалов у трех видов (гиены, большого и малого пещерных медведей) на Урале связано с постепенным их вымиранием на территории Евразии, которое в свою очередь обусловлено ухудшением климатических условий в середине позднего плейстоцена (Pacher, Stuart, 2009; Stuart, Lister, 2014). Вероятно, смещение северной границы ареала благородного оленя на юг от 55° с.ш в середине позднего плейстоцена можно объяснить этими же причинами. В Западной Европе в период максимально холодной стадии позднего плейстоцена ареал благородного оленя так же сократился и занимал территорию к югу от 45° с.ш. (Sommer et al., 2008). Расселение благородного оленя из южных частей Европы к северу начинается вновь в теплый период Беллинга 12 500 лет назад, (Sommer et al., 2008). В современных условиях сурового климата Якутии отмечается активное расселение благородного оленя по территории. Исследователи объясняют это явление несколькими факторами, и в том числе потеплением климата (Смирнов, 2006; Степанова, 2009). Дальнейшему расширению ареала благородного оленя в северном направлении в Якутии, по мнению ученых, кроме прочих причин (естественные географические преграды, высокая плотность людского населения и другие) могут препятствовать продолжительные зимы и крайне низкие температуры (Боескоров, 1999).
Единичные находки архара и верблюда не позволяют реконструировать характер динамки ареалов этих видов на Урале. Скорее всего, сокращение ареалов этих видов в конце невьянского интерстадиала, также обусловлено похолоданием климата, т.к. в современных условиях они обитают в теплом или умеренно-теплом климате.
Сокращение ареала тарбаганчика произошло позднее, чем у предыдущих видов, в конце позднеледниковья. Вероятнее всего, сокращение ареала этого вида также обусловлено постепенным потеплением и увлажнением климата. В современных условиях представители этого вида предпочитают пустынные местообитания (Громов, Ербаева, 1995). В конце позднеледниковья последовало несколько быстро сменяющих друг друга потеплений и похолоданий. В какой из этих периодов произошло сокращение ареала тарбаганчиков – возможности установить нет, а, следовательно, нет возможности выявить, какие именно климатические изменения повлияли на сокращение ареала у этого вида.
Расширение ареалов у млекопитающих в позднем плейстоцене на Урале обусловлено как вселением одних видов, так и расширением ареалов к северу у других. Эта группа объединяет виды с различными экологическими характеристиками (табл. 1). У видов, приуроченных к лесным местообитаниям, расширение ареала началось в период постепенного потепления климата в течение позднеледниковья и перехода к голоцену. Большой тушканчик – обитатель пустынных местообитаний, расселяется на Южный Урал в период максимально холодной стадии позднего плейстоцена, когда господствовали очень сухие и холодные климатические условия. Другой представитель пустынных местообитаний – обыкновенная слепушонка расселяется на Урал в конце позднеледниковья, вероятно, в один из периодов потепления. Для этого зверька, ведущего норный образ жизни, решающим фактором для распространения, вероятно, была деградация вечной мерзлоты.
И, наконец, последняя группа млекопитающих с флуктуирующим ареалом включает виды, связанные с древесно-кустарниковой растительностью, один околоводный вид и один степной вид (табл. 1). Северная граница распространения большинства видов этой группы в максимально холодную стадию позднего плейстоцена смещалась к югу от территории, на которой они обитали ранее. Позднее, в период позднеледниковья, их ареалы восстановились в прежних границах, а у ряда видов к началу голоцена продвинулись севернее. Обратную картину динамики ареала имел единственный представитель степных местообитаний – корсак. Его ареал продвинулся к северу к началу максимально холодной стадии, а затем восстановился в прежних границах. Малое количество сведений о находках этого вида не позволяет выявить причины такой динамики.
Отдельно следует отметить границы обитания некоторых видов млекопитающих на Урале в позднем плейстоцене. По западному склону Урала в позднем плейстоцене проходила восточная граница ареала большого пещерного медведя. К северу от 57° с.ш. обитали полевка Миддендорфа и овцебык. К югу от 61° с.ш. обитали степная пеструшка и серый хомячок, к югу от 58° с.ш. – лесная мышовка, обыкновенная полевка, хомячок Эверсманна, желтая пеструшка, пещерная гиена, корсак и гигантский олень. Ареалы таких видов как бобр, хомяк, рысь, барсук и малый пещерный медведь в позднем плейстоцене не выходили за границы Среднего Урала (Bachura, Kosintsev, 2007). Все эти виды имеют различные экологические характеристики. Овцебык – обитатель арктических тундр; полевка Миддендорфа – обитатель северо-таежных лесов и кустарниковой тундры; рысь – облигатный обитатель таежных лесов; лесная мышовка, бобр, барсук и благородный олень – виды, предпочитающие местообитания умеренного климата; обыкновенная полевка – лесостепной вид; серый хомячок и хомячок Эверсманна, желтая и степная пеструшка, корсак, малый пещерный медведь – степные и пустынно-степные виды. Границы их распространения маркируют, вероятно, границы зональных вариантов позднеплейстоценового комплекса мамонтовой фауны (Смирнов и др., 2014).
Таким образом, изменение ареалов млекопитающих в позднем плейстоцене на Урале тесным образом связано с динамикой климата в течение этого периода. По характеру динамики ареалов в позднем плейстоцене можно выделить четыре основных типа: постоянный, расширявшийся, сокращавшийся и флуктуирующий. В пределах этих типов ареалов входят виды с различными экологическими требованиями к среде. Причем виды со сходными экологическими требованиями зачастую оказываются в разных группах. Стабильный (постоянный) ареал на Урале в позднем плейстоцене имели виды с очень широким спектром экологических характеристик, ряд степных видов, тундровые виды, а также представители мамонтового комплекса. Эти виды млекопитающих составляли основу териофауны позднего плейстоцена на Урале.
Виды млекопитающих с нестабильным ареалом имели невысокую численность в течение позднего плейстоцена, а ряд из них были очень редкими. Реакция млекопитающих, связанных с древесно-кустарниковой растительностью, была неодинаковой у различных видов. Одни виды смогли расселиться на Урале только с началом потепления климата, у других происходило смещение северной границы ареала к северу в периоды потепления и к югу в периоды похолодания, единичные виды имели стабильный ареал. Представители степных и пустынных видов млекопитающих имели все указанные типы ареалов.
Описанная нами картина показывает, что динамики ареалов млекопитающих в позднем плейстоцене сопряжена с динамикой климата. Реакция видов со сходными экологическими требованиями на климатические изменения у каждого вида была индивидуальная.
Список литературы
Бачура О.П., Струкова Т.В., 2002. Остатки млекопитающих из местонахождения Черемухово-1: (раскоп 4) // Фауна Урала в плейстоцене и голоцене. Сб. науч. трудов. Екатеринбург: Изд-во “Университет”. С. 37–55.
Боескоров Г.Г., 1999. К систематическому положению и истории благородных оленей Якутии // Редкие виды млекопитающих России и сопредельных территорий. М.: Наука. С. 40–53.
Бородин А.В., Косинцев П.А., Струкова Т.В., Некрасов А.Е., 2000. Млекопитающие, птицы и рыбы из местонахождения Черемухово1 (раскоп 2) // Плейстоценовые и голоценовые фауны Урала. Челябинск: Изд-во “Рифей”. С. 59–80.
Бородин А.В., Струкова Т.В., Косинцев П.А., Некрасов А.Е., Панова Н.К., 2000а. Новые данные о природной среде Среднего Урала в позднечетвертичное время (местонахождение Шайтаноозерский Каменный остров) // Плейстоценовые и голоценовые фауны Урала. Челябинск: Изд-во “Рифей”. С. 17–35.
Громов И.М., Ербаева М.А., 1995. Млекопитающие фауны России и сопредельных территорий. СПб.: ЗИН РАН. 522 с.
Данукалова Г.А., Юрин В.И., Косинцев П.А., Осипова Е.М., Курманов Р.Г., 2018. Биостратиграфические исследования отложений верхнего плейстоцена и голоцена пещеры Сикияз-Тамак 7 (Южный Урал, Россия) // Геологический вестник. № 1. С. 144–161.
Изварин Е.П., 2004.Новые материалы по фауне мелких млекопитающих плейстоцен-голоценового перехода на западном склоне Среднего Урала // Экологические механизмы динамики и устойчивости биоты. Екатеринбург: Изд-во “Академкнига”. С. 91–92.
Изварин Е.П., 2006. Новые материалы по фауне мелких млекопитающих позднего плейстоцена и раннего голоцена на Западном склоне Среднего Урала // Экология в меняющемся мире. Екатеринбург: Изд-во “Академкнига”. С. 82–83.
Изварин Е.П., 2009. Ископаемые фауны мелких млекопитающих из местонахождения навес Светлый на реке Серге (Средний Урал) // Эволюционная и популяционная экология (назад в будущее). Екатеринбург: Изд-во “Гощицкий”. С. 75–79.
Косинцев П.А., 1995. Остатки крупных млекопитающих из Лобвинской пещеры // Материалы по истории современной биоты Среднего Урала. Екатеринбург: Изд-во “Екатеринбург”. С. 58–102.
Косинцев П.А., 1996. Фауна крупных млекопитающих Северного Урала в позднем плейстоцене и голоцене // Материалы и исследования по истории современной фауны Урала. Екатеринбург: Изд-во “Екатеринбург”. С. 84–110.
Косинцев П.А., Воробьев А.А., 2000. Остатки крупных млекопитающих из местонахождения. Виашер на Среднем Урале // Плейстоценовые и голоценовые фауны Урала. Челябинск: Изд-во “Рифей”. С. 105–122
Косинцев П.А., Крахмальная Т.В., 2008. Фауна млекопитающих из пещеры Георгиевская (Средний Урал) // Фауна и флора Северной Евразии в позднем кайнозое. Екатеринбург– елябинск: ООО “ЦИКР “Рифей”. С. 124–136.
Косинцев П.А., Орлова М.В., 2002. Крупные млекопитающие из местонахождений “Лобвинская пещера” и “Лобва I” // Плейстоценовые и голоценовые фауны Урала. Екатеринбург: Изд-во “Университет”. С. 136–145.
Косинцев П.А., Пластеева Н.А., 2009. Лошади подрода (подрод Equus) Южного Урала в позднем плейстоцене // Вестник Оренбургского государственного университета. № 6. С. 161–163.
Кряжева И.В., Пономарев Д.В., 2014. Позднечетвертичная микротериофауна бассейна р. Кожим (Приполярный Урал) // Вестник. № 5. С. 3–8.
Кряжева И.В., Пономарёв Д.В., ван Кольфсхотен Т., ван дер Плихт Х., 2012. История формирования современных сообществ микромаммалий Приполярного Урала // Экология. № 6. С. 434–441.
Кузьмина Е.А., 2003 Позднеплейстоценовые и голоценовые сообщества млекопитающих из пещерных местонахождений Южного Зауралья // Четвертичная палеозоология на Урале. Екатеринбург: Изд-во Уральского университета. С. 193–210.
Кузьмина Е.А., Смирнов Н.Г., Коурова Т.П., 2001. Фауны грызунов Южного Зауралья в позднем плейстоцене и голоцене // Современные проблемы популяционной, исторической и прикладной экологии. Екатеринбург: Изд-во “Екатеринбург”. С. 121–127.
Кузьмина И.Е., 1971. Формирование териофауны Северного Урала в позднем антропогене // Труды Зоол. ин-та АН СССР. Т. 49. Л.: Наука. С. 44–122.
Кузьмина И.Е., 1975. Некоторые данные о млекопитающих Среднего Урала в позднем плейстоцене // Бюллетень комиссии по изучению четвертичного периода. № 43. М.: Наука. С. 63–78.
Кузьмина И.Е., Саблин М.В., Цыганова С.А., 1999. Видовой состав и морфологические особенности млекопитающих из грота Большой Глухой на Среднем Урале // Охранные археологические исследования на Среднем Урале. Сб. ст. Вып. 3. Екатеринбург: Банк культурной информации. С. 4–14.
Кузьмина С.А., 2000. Фаунистические данные по позднепалеолитической стоянке Смеловская II на Южном Урале // Плейстоценовые и голоценовые фауны Урала. Челябинск: Изд-во “Рифей”. С. 137–153.
Лавров И.А., Андрейчук В.Н., 1992. Пещеры Урала и Приуралья. Пермь: Изд-во Пермского университета. 77 с.
Лаптева Е.Г., 2009. Ландшафтно-климатические изменения на восточном склоне Северного Урала за последние 50 тыс. лет назад // Экология. № 4. С. 284–290.
Маркова А.К., Кольфсхотен Т. ван, Бохнкке Ш., Косинцев П.А., Мол И. и др., 2008. Эволюция экосистем Европы при переходе от плейстоцена к голоцену (24–8 тыс. л.н.). М.: Товарищество научных изданий КМК. 556 с.
Орлова Л.А., 1995. Радиоуглеродное датирование археологических памятников Сибири и Дальнего Востока // Методы естественных наук в археологических реконструкциях. Новосибирск: Институт археологии и этнографии СО РАН С. 208–232.
Подопригора И.Н., 1999. Фауна крупных млекопитающих из пещер Пермского Приуралья // Развитие идей академика С.С. Шварца в современной экологии. Екатеринбург: Изд-во “Екатеринбург”. С. 143–146.
Пономарев Д.В., 2001. Крупные млекопитающие Европейского Северо-Востока в позднем плейстоцене и голоцене. Сыктывкар: Изд-во Коми научного центра УрО РАН. 48 с.
Ражев Д.И., Косинцев П.А., Улитко А.И., 2005. Фауна крупных млекопитающих позднего плейстоцена и голоцена из грота Бобылек (Средний Урал) // Фауны Урала и Сибири в плейстоцене и голоцене. Челябинск: Изд-во “Рифей”. С. 190–211.
Смирнов М.Н., 2006. Благородный олень в Южной Сибири: монография. Ч. 1. Красноярск: РИО КрасГУ. 250 с.
Смирнов Н.Г., 1993. Мелкие млекопитающие Среднего Урала в позднем плейстоцене и голоцене. Екатеринбург: УИФ Наука. 64 с.
Смирнов Н.Г., 1995. Материалы к изучению исторической динамики разнообразия грызунов таежных районов Среднего Урала // Материалы по истории современной биоты Среднего Урала. Екатеринбург: Изд-во “Екатеринбург”. С. 24–57.
Смирнов Н.Г., 1996. Разнообразие мелких млекопитающих Северного Урала в позднем плейстоцене и голоцене // Материалы и исследования по истории современной фауны Урала. Екатеринбург: Изд-во “Екатеринбург”. С. 39–83.
Смирнов Н.Г., Большаков В.Н., Косинцев П.А., Панова Н.К., Коробейников Ю.И., Ольшванг В.Н. и др., 1990. Историческая экология животных Южного Урала. Свердловск: УрО РАН. 245 с.
Смирнов Н.Г., Ерохин Н.Г., Быкова Г.В., Лобанова А.В., Корона О.М., Широков В.Н. и др., 1992. Грот Сухореченский – памятник истории природы и культуры в Красноуфимской лесостепи // История современной фауны Южного Урала. Свердловск: УрО РАН СССР. С. 20–43.
Смирнов Н.Г., Косинцев П.А., Кузьмина Е.А., Изварин Е.П., Кропачева Ю.Э., 2014. Экология четвертичных млекопитающих на Урале // Экология. № 6. С. 403–409.
Степанова В.В., 2009. Расширение ареала благородного оленя в Якутии // Российский журнал биологических инвазий. № 2. С. 49–59.
Струкова Т.В., 2000. Типология фаун мелких млекопитающих позднего плейстоцена и голоцена Среднего Зауралья // Биосфера и человечество. Екатеринбург: Изд-во “Екатеринбург”. С. 251–260.
Струкова Т.В., 2002. Степная пеструшка и узкочерепная полевка в ископаемых фаунах Среднего и Южного Зауралья // Биота горных территорий: история и современное состояние. Екатеринбург: Изд-во “Академкнига”. С. 207–215.
Струкова Т.В., Бачура О.П., Бородин А.В., Стефановский В.В., 2006. Первые находки млекопитающих в аллювиально-спелеогенных образованиях позднего плейстоцена и голоцена Северного Урала (Черемухово-1) // Стратиграфия. Геологическая корреляция. Т. 14. № 1. С. 98–108.
Тетерина А.А., 2002. Ископаемые фауны мелких млекопитающих из местонахождений Северного Урала // Фауны Урала в плейстоцене и голоцене. Екатеринбург: Изд-во “Университет”. С. 111–135.
Урал и Приуралье, 1968. М.: Наука. 461 с.
Фадеева Т.В., Смирнов Н.Г., 2008. Мелкие млекопитающие Пермского Предуралья в позднем плейстоцене и голоцене. Екатеринбург: Изд-во “Гощицкий”. 171 с.
Щербакова Т.И., 1994. Материалы Верхнепалеолитической стоянки Талицкого (Островской). Екатеринбург: Ин-т истории и археологии. УрО РАН. 64 с.
Широков В.Н., Волков Р.Б., Косинцев П.А., Лаптева Е.Г., 2011. Палеолитическая стоянка Богдановка (Южный Урал) // Российская археология. № 1. С. 125–139.
Яковлев А.Г., 1997. Мелкие млекопитающие из позднеплейстоценовых и голоценовых пещерных местонахождений западного склона Южного Урала // Пещерный палеолит Урала. Уфа: Принт. Вып. 6. С. 174–181.
Яковлев А.Г., Данукалова Г.А., Алимбекова Л.И., Яковлева Т.И., Еремеев А.А., Морозова Е.М., 2005. Биостратиграфическая характеристика отложений позднего неоплейстоцена – голоцена в районе памятника природы “Водопад Атыш” (Южный Урал) // Фауны Урала и Сибири в плейстоцене и голоцене. Челябинск: Изд-во “Рифей”. С. 260–303.
Bachura O.P., Kosintsev P.A., 2007. Late Pleistocene and Holocene small- and large-mammal faunas from the Northern Urals // Quaternary International. V. 160. P. 121–128.
Danukalova G., Kurmanov R., Yakovlev A., Osipova E., Zinovyev E., Arslanov K., 2016. Palaeoenvironment of the Middle and Upper Neopleistocene at the Gornovo Upper Palaeolithic site (Southern Ural foreland, Russia) // Quaternary International. V. 420. P. 24–46.
Danukalova G., Yakovlev A., Alimbekova L., Yakovleva T., Morozova E., et al., 2008. Biostratigraphy of the Upper Pleistocene (Upper Neopleistocene) – Holocene deposits of the Lemeza River valley of the Southern Urals region (Russia) // Quaternary International. V. 190. Issue 1. P. 38–57
Danukalova D., Yakovlev A., Osipova E., Alimbekova L., Yakovleva T., Kosintcev P., 2011. Biostratigraphy of the Late Upper Pleistocene (Upper Neopleistocene) to Holocene Deposits of the Belaya River Valley (Southern Urals, Russia) // Quaternary International. V. 231. Issue 1–2. P. 28–43.
Golovachev I.B., Smirnov N.G., 2009. The Late Pleistocene and Holocene rodents of the Pre-Urals subarctic // Quaternary International. V. l (201). P. 37–42.
Kosintsev P.A., Bachura O.P., 2013. Late Pleistocene and Holocene mammal fauna of the Southern Urals // Quaternary International. V. 284. P. 161–170.
Kuzmina E.A., 2009. Late Pleistocene and Holocene small mammal faunas from the South Trans-Urals. Quaternary International. V. l (201). P. 25–30.
Markova E., Beeren Z., van Kolfschoten T., Strukova T., Vrieling K., 2012. Differentiating sibling species in the Quaternary fossil record: a comparison of morphological and molecular methods to identify Microtus arvalis and M. rossiaemeridionalis (Arvicolinae, Rodenia) // Journal of Systematic Palaeontology. V. 10. Issue 3. P. 585–597
Pacher M., Stuart A., 2009. Extinction chronology and palaeobiology of the cave bear (Ursus spelaeus) // Boreas. V. 38. P. 189–206.
Ponomarev D.V., van Kolfschoten T., van der Plicht J., 2012. Late Glacial and Holocene micromammals of northeastern Europe // Russian J. Theriol. V. 11(2). P. 121–130.
Ponomarev D., van Kolfschoten T., van der Plicht J., 2013. Late Glacial and Holocene small mammals of the Timan Ridge (Komi Republic, Russia) // Quaternary International. V. 284. P. 177–183.
Prost S., Knapp M., Flemmig J., Hufthammer A.K., Kosintsev P., et al., 2010. Phantom extinction? New insights into extinction dynamics of the Don-hare Lepus tanaiticus // Journal of Evolutionary Biology. V. 23. P. 2022–2029.
Sommer R.S., Zachos F.E., Street M., Jöris O., Skog A., Benecke N., 2008. Late Quaternary distribution dynamics and phylogeography of the red deer (Cervus elaphus) in Europe // Quaternary Science Reviews. V. 27. 714–733.
Stuart A., Lister A., 2014. New radiocarbon evidence on the extirpation of the spotted hyaena (Crocuta crocuta (Erxl.)) in northern Eurasia // Quaternary Science Reviews. V. 96. P. 106–116.
Teterina A., 2009. Rodents of the North Urals in the Late Pleistocene and Holocene // Quaternary International. V. 201. P. 31–36.
Yakovlev A., Danukalova G., Kosintcev P., Alimbekova L., Morozova E., 2006. Biostratigraphy of the Late Palaeolithic site of “Bajslan-Tash cave” (the Southern Urals) // Quaternary International. V. 149. Issue 1. P. 115–121.
Дополнительные материалы отсутствуют.
Инструменты
Зоологический журнал