

Зоологический журнал, 2019, T. 98, № 8, стр. 915-927
Анализ фауны и населения птиц южных отрогов Колымского нагорья
А. А. Романов a, *, Е. В. Мелихова b, **, Н. А. Миклин c, ***, В. О. Яковлев d, ****
a Географический факультет Московского государственного университета
им. М.В. Ломоносова
119991 Москва, Россия
b Всероссийский научно-исследовательский институт
охраны окружающей среды
117628 Москва, Россия
c Институт биологии и химии, Московский педагогический
государственный университет
129164 Москва, Россия
d Русское общество сохранения и изучения птиц им. М.А. Мензбира
121099 Москва, Россия
* E-mail: putorana05@mail.ru
** E-mail: max-kun@yandex.ru
*** E-mail: nikolaymik@gmail.com
**** E-mail: bird-yakovlev@yandex.ru
Поступила в редакцию 22.10.2018
После доработки 12.03.2019
Принята к публикации 18.03.2019
Аннотация
Проанализированы экологические закономерности высотно-поясной дифференциации фауны и населения птиц в южных отрогах Колымского нагорья. Исследования проведены в районах (n = 11), где четко выражены три высотных пояса: горно-таежный (до 1000 м над ур. м.), подгольцовый (1000–1700 м над ур. м.) и гольцовый (до 2600 м над ур. м.). Обследование вели методом маршрутного учета на трансектах неограниченной ширины. Фауна гнездящихся птиц была проанализирована с точки зрения принадлежности видов к фаунистическим комплексам и зонально-ландшафтным группам. Сходство орнитофаун сравниваемых районов и высотных поясов определялось по коэффициенту фаунистической общности Серенсена и Жаккара. Для выявления различий в населении птиц разных участков был использован коэффициент сходства населения. Высока общность таксономической структуры и видового состава гнездовой авифауны региона, насчитывающей 74 вида. Ряд видов (n = 16) впервые зарегистрирован нами на гнездовье в южных отрогах Колымского нагорья на удалении 20–800 км от северных границ основного ареала. Таксономическая структура гнездовой авифауны соответствует зональным и ландшафтным особенностям Северо-Восточной Азии с доминированием видов из отрядов воробьеобразных, ржанкообразных, гусеобразных, суммарно составляющих 83%. Наиболее значимы в формировании авифауны виды сибирского фаунистического комплекса (33–49%). В зоогеографическом отношении авифауна южных отрогов Колымского нагорья имеет специфичные черты за счет видов (n = 8), экологически связанных с горными ландшафтами. Авифауна южных отрогов Колымского нагорья формируется в системе общих зонально-ландшафтных и высотно-поясных закономерностей. Сокращение видового разнообразия птиц происходит с увеличением высоты – от подножий к вершинам. Изменение видового состава птиц с высотой происходит постепенно. В горно-таежном поясе южных отрогов Колымского нагорья гнездится 68 видов (92%), в подгольцовом – 33 вида (45%), гольцовом – 15 видов (20%). В широком диапазоне высот, охватывающем не менее двух высотных поясов, обитают 45% видов. В южных отрогах Колымского нагорья с высотой поступательно сокращаются плотность населения птиц и обилие абсолютного большинства видов. Основное сокращение плотности населения птиц происходит при переходе из подгольцового пояса в гольцовый. Плотность населения птиц в горно-таежном поясе южных отрогов Колымского нагорья 176–674 особей/км2, в подгольцовом – 76–230 особей/км2, гольцовом – 20–72 особей/км2.
Итоги представленных исследований лежат в сфере изучения пространственной организации фауны и населения птиц и направлены на оценку биоразнообразия гор Северной Азии на примере обширной горной области, образованной южными отрогами Колымского нагорья. Несмотря на то, что известны обобщающие орнитологические работы по горным районам Северной Азии (Кищинский, 1988; Романов, 2013), закономерности формирования фауны и населения птиц горных территорий этой обширной части суши до сих пор изучены неудовлетворительно. Монография Кищинского (1968) основана на полевых исследованиях с широкой географией в целом. Но при этом эти исследования были проведены лишь в некоторых частях региона, и не имели основную цель по изучению закономерностей пространственной дифференциации фауны и населения птиц в условиях высотной поясности. Внутренние части южных районов нагорья остались почти не исследованы. Имеющиеся в публикации сведения собраны 50 лет назад, поэтому правомерно предположить, что какая-то их часть могла устареть и требует существенной корректировки. Знания об экологических особенностях высотно-поясной дифференциации населения птиц в горных условиях Колымского нагорья до сих пор остаются фрагментарными. При этом в сфере изучения биологического разнообразия познание фаунистических комплексов обширных горных регионов признается одним из актуальных вопросов (Баранов, 2007; Ruggiero, Hawkins, 2008; McCain, 2009; Вартапетов, Гермогенов, 2011; Graham et al., 2014). Очевидным вкладом в его решение может стать выявление биогеографических закономерностей формирования авифауны Колымского нагорья.
Основная цель – выявление эколого-географических закономерностей пространственной дифференциации авифауны Колымского нагорья для использования при мониторинге и разработке мер сохранения биологического разнообразия. В соответствии с этим, результаты исследований 2016 г. позволили решить 3 основные задачи: 1 – выполнение инвентаризации видового состава авифауны; 2 – выявление или уточнение границ ареалов, особенностей распространения и обилия видов; 3 – установление высотно-поясной, широтной и меридиональной дифференциации фауны и населения птиц.
МАТЕРИАЛЫ И МЕТОДЫ
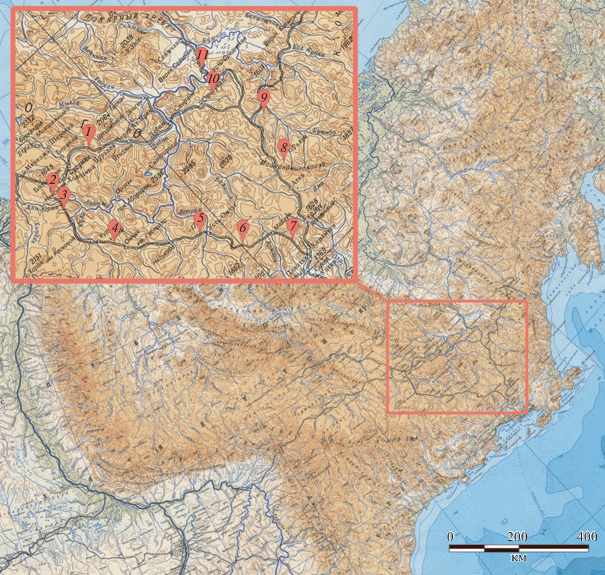
Исследования проводились с 7 по 29 июня 2016 г. в южных отрогах Колымского нагорья: хребты Черге и Майманджинский, горы Делурэкчен, перевалы Лошкалах, Гаврюшка, Кулу, горные массивы в бассейнах рек Омчуг, Салтахан, Герба, Кинжал, Колыма. Обследованы обширные территории, сопредельные с федеральной трассой “Колыма” (Якутск–Магадан) и расположенной южнее Тенькинской трассой. Протяженность кольцевого маршрута, пройденного на автомашине по этим трассам, составила около 1400 км. Обследована территория в пределах 60°–63° с.ш., 147°–153° в.д. (рис. 1).
Рис. 1.
Пункты исследования в южных отрогах Колымского нагорья: 1 – хребет Черге, 2 – перевал Лошкалах, 3 – перевал Гаврюшка, 4 – перевал Кулу, 5 – р. Омчуг, 6 – р. Салтахан, 7 – горы Делурэкчэн, 8 – Майманджинский хребет, 9 – р. Герба, 10 – р. Кинжал, 11 – р. Колыма.
Объект исследований – авифауна южных отрогов Колымского нагорья. Проанализированы данные о гнездовой авифауне горных регионов, в пределах которых выражены гольцовый, подгольцовый и горно-таежный высотно-ландшафтные пояса. Для анализа привлечены результаты маршрутных учетов птиц по методике Равкина (1967), проведенных на высотах 240–1450 м над ур. м. Суммарная протяженность пеших учетных маршрутов составила 356 км, из которых 255 км – в горно-таежном поясе, 74 км – в подгольцовом поясе, 27 км – в гольцовом поясе. Достоверность гнездования определяли в соответствии с критериями, рекомендованными Европейским комитетом по учету птиц (The EBCC Atlas, 1997). Сходство авифаун сравниваемых горных систем определяли по коэффициенту фаунистической общности – КФО, который рассчитывали по формулам Серенсена и Жаккара (Песенко, 1982; Чернов, 2008). Для выявления различий в населении птиц разных участков был использован коэффициент сходства населения – КСН (Наумов, 1964). Доминантными считали виды, доля которых в общей плотности населения составила более 10%, субдоминантными – 1–10%. Фауна гнездящихся птиц охарактеризована по типам фаун (Штегман, 1938) и в свете представлений о зонально-ландшафтных (географо-генетических) группах птиц (Чернов, 1975, 1978; Кищинский, 1988). При этом в категории фаунистических комплексов за широкораспространенные приняты виды с обширным ареалом и неясным центром происхождения. В категории зонально-ландшафтных групп широкораспространенными считали виды с обширным ареалом, современное распространение которых совпадает с несколькими природными зонами. В номенклатуре и при составлении списков птиц мы следовали конспекту орнитологической фауны России и сопредельных территорий Степаняна (2003). Названия некоторых видов приняты по Списку птиц Российской Федерации (Коблик и др., 2006).
ФИЗИКО-ГЕОГРАФИЧЕСКАЯ ХАРАКТЕРИСТИКА РАЙОНА ИССЛЕДОВАНИЙ
Описание физико-географических условий Колымского нагорья содержится в ряде монографий (Гвоздецкий, Михайлов, 1987; Голубчиков, 1996; Куваев, 2006). Поэтому укажем лишь, что для обследованной территории, где по абсолютным высотам (1000–3000 м над ур. м.) господствуют среднегорные участки, свойственно мозаичное сочетание горных хребтов с типично альпийскими формами рельефа и горных массивов с плоскими выровненными вершинами. Для большей территории характерны континентальный или резко континентальный типы умеренного климата. Обследованные районы расположены в подзоне северной тайги. В связи с четко выраженной вертикальной поясностью растительность принято подразделять на три высотно-ландшафтных пояса: горно-таежный (лесной), подгольцовый (горные редколесья и кустарники) и гольцовый (горнотундровый) (Юрцев, 1968; Сочава, 1980; Куваев, 2006). Горно-таежный пояс, где повсеместно господствует лиственница Каяндера (Larix kajanderii), охватывает пространство от подножий склонов до 700–1000 м над ур. м. С высотой горная лиственничная тайга постепенно переходит в лиственничные редколесья и редины. Подгольцовый пояс расположен в пределах высот 1000–1700 м над ур. м. Здесь господствует кедровый стланик (Pinus pumila (Pall.) Regel) с участием березы тощей (Betula exilis Sukaczev), березы Миддендорфа (Betula middendorffii Trautv. & C.A. Mey.), рододендрона мелколистного (Rhododendron parvifolium Adams), можжевельника (Juniperus sibirica Burgsd.). Пояс горных тундр и гольцовых пустынь (гольцовый пояс) распространен до высот 2000–2600 м над ур. м. В гольцовом поясе преобладают ерниковые тундры с карликовой березой (Betula nana L.), мохово-травяные тундры, дриадовые тундры с дриадой точечной (Dryas punctata Juz.), мертвые и накипно-лишайниковые пустыни. По глубоким речным долинам распространены галерейные лиственные леса из благовонного тополя (Рopulus suaveolens Fisch.) и чозении (Chosenia arbutifolia (Pall.) A.K. Skvortsov).
РЕЗУЛЬТАТЫ
Таксономическая структура авифауны
Опубликованный Кищинским (1968) список птиц Колымского нагорья включает 133 вида, из которых 114 – гнездящихся. В обследованных нами пунктах (n = 11) южных отрогов Колымского нагорья зарегистрировано 74 вида птиц с подтвержденным, вероятным или возможным гнездованием (в дальнейшем все они условно именуются гнездящимися), что составляет 65% от всей гнездовой авифауны региона.
Таксономическая структура гнездовой авифауны южных отрогов Колымского нагорья соответствует зональным и ландшафтным особенностям рассматриваемой части Северо-Восточной Азии, в целом сохраняется на всем протяжении от бассейна реки Колымы на северо-востоке до перевала Кулу на юго-западе и включает в себя 7 отрядов с доминированием трех из них, наиболее характерных для бореального и гипоарктического поясов Палеарктики: воробьеобразных (Passeriformes) (42 вида, 57%), ржанкообразных (Charadriiformes) (11 видов, 15%) и гусеобразных (Anseriformes) (8 видов, 11%). Суммарно доля этих отрядов в южных отрогах Колымского нагорья составляет 83% отмеченных видов. В обследованных нами пунктах южных отрогов Колымского нагорья отмечены представители 4–7 отрядов. По числу представленных видов везде преобладают воробьеобразные (59–83%), в авифауне половины пунктов также значимы ржанкообразные (11–18%).
Прослежены изменения соотношения таксономических групп с высотой (табл. 1).
Таблица 1.
Изменение с высотой соотношения таксономических групп в гнездовой авифауне южных отрогов Колымского нагорья, число видов
| Отряд | Число видов | |||||
|---|---|---|---|---|---|---|
| горно-таежный пояс | подгольцовый пояс | гольцовый пояс | ||||
| абс. | % | абс. | % | абс. | % | |
| Гусеобразные | 8 | 12 | – | – | – | – |
| Соколообразные | 4 | 6 | 2 | 6 | 2 | 13 |
| Курообразные | 4 | 6 | 1 | 3 | 1 | 7 |
| Ржанкообразные | 10 | 15 | 4 | 12 | 1 | 7 |
| Кукушкообразные | 2 | 3 | 2 | 6 | 1 | 7 |
| Дятлообразные | 1 | 1 | – | – | – | – |
| Воробьеобразные | 39 | 57 | 24 | 73 | 10 | 66 |
Широтная и меридиональная дифференциация авифауны
Установлено, что в обследованных пунктах (n = 11) южных отрогов Колымского нагорья локальные гнездовые авифауны насчитывают от 23 до 44 видов. Максимальное видовое разнообразие птиц зарегистрировано в долине р. Колыма, что объясняется высоким разнообразием экологических условий, и как следствие – весьма широким спектром местообитаний для самых разных видов птиц, особенно – водно-околоводных.
Показатель представленности общей гнездовой авифауны всего обследованного региона в отдельных пунктах – 31–59%. Одновременно во всех или почти всех районах (n = 10–11) гнездится 14 видов птиц, формирующих общее фаунистическое ядро: обыкновенная кукушка (Cuculus canorus (Linnaeus 1758)), пятнистый конек (Anthus Hodgsoni (Richmond 1907)), горная трясогузка (Motacilla cinerea (Tunstall 1771)), кедровка (Nucifraga caryocatactes macrorhynchos (Linnaeus 1758)), ворон (Corvus corax (Linnaeus 1758)), таловка (Phylloscopus borealis (Blasius 1858)), зарничка (Phylloscopus inornatus (Blyth 1842)), корольковая пеночка (Phylloscopus (proregulus) proregulus (Pallas 1811)), бурая пеночка (Phylloscopus fuscatus (Blyth 1842)), соловей-красношейка (Luscinia calliope (Pallas 1776)), синехвостка (Tarsiger cyanurus (Pallas 1773)), вьюрок (Fringilla montifringilla (Linnaeus 1758)), обыкновенная чечевица (Caprodacus erythrinus (Pallas 1770)), овсянка-крошка (Ocyris pusillus (Pallas 1776)). В подавляющем большинстве обследованных пунктов (n = 7–9) встречены также чеглок (Falco subbuteo (Linnaeus 1758)), тундряная куропатка (Lagopus muta (Montin 1781)), перевозчик (Actitis hypoleucos (Linnaeus 1758)), глухая кукушка (Cuculus (saturatus) optatus (Gould 1845)), белая трясогузка (Motacilla alba (Linnaeus 1758)), зелёная пеночка (Phylloscopus trochiloides (Sundevall 1837)). Большинство всех перечисленных видов широко распространены в северной тайге, лесотундре и, частично, в южной тундре. Коэффициенты взаимной общности локальных гнездовых авифаун обследованных районов южных отрогов Колымского нагорья, полученные по формулам Серенсена и Жаккара, составляют 45(29)–75(60)%. Это свидетельствует о формировании однородной авифауны в пределах Колымского нагорья, что, вероятно, предопределено относительно стабильным развитием местных экосистем в постледниковую эпоху (последние 12–14 тысяч лет) (Кищинский, 1988; Голубчиков, 1996). В это время отдельные элементы авифауны Северо-Восточной Сибири имели возможность почти беспрепятственно расселяться по региону, что поддерживалось и поддерживается сходством современных экологических условий. В целом однородная бореально-гипоарктическая авифауна в современном ее виде сформировалась в пределах единой области Северо-Восточной Азии с повсеместным господством северо-таежных лесов, гипоарктических редколесий и южных кустарниковых тундр (Кищинский, 1988). В условиях мозаичного сочетания этих ландшафтов и повсеместной сопряжeнности их горных и равнинных аналогов подавляющее большинство видов птиц освоило их повсеместно, так как не имело непреодолимых преград, препятствующих расселению.
Границы гнездовых ареалов
В 2016 г. нами зарегистрирован ряд видов (n = 16), статус пребывания и характер географического распространения которых в южных отрогах Колымского нагорья до сих пор были не известны, не точны, противоречивы или подтверждены единичными наблюдениями (Кищинский, 1968; Cтепанян, 2003; Андреев и др., 2006; Рябицев, 2014; Андреев и др., 2015, 2015а). По нашим данным, гнездование этих видов в обследованном регионе достоверно или вероятно. Встречи обсуждаемых видов зафиксированы в 1–11 пунктах на расстоянии от 20 до 800 км от основной области своего распространения. Не исключено, что некоторые виды, впервые зарегистрированные нами в горах Северо-Восточной Сибири, появились здесь в результате гнездования за пределами своего ареала или, возможно, даже его расширения.
Весьма мозаичный ареал большого песочника (Calidris tenuirostris (Horsfield 1821)) (Tomkovich, 1997) дополнен новыми фрагментами, впервые обнаруженными нами в горах бассейнов рек Салтахан (60°45′ с.ш., 149°56′ в.д.) и Кинжал (62°17′ с.ш., 151°57′ в.д.), удаленными на 500 км от известных районов обитания. Территориальные гнездовые пары альпийской завирушки (Prunella collaris (Scopoli 1769)) зафиксированы нами в горах Делурэкчен (60°26′ с.ш., 150°58′ в.д.) и горных массивах бассейна р. Салтахан (60°45′ с.ш., 149°56′ в.д.), где ее гнездование ранее считалось неподтвержденным (Рябицев, 2014).
Актуальна регистрация в пределах Колымского нагорья таких видов, как зимняк (Buteo lagopus (Pontoppidan 1763)), обыкновенный канюк (Buteo buteo (Linnaeus 1758), халей (Larus heuglini (Bree 1876)), зеленая пеночка, синий соловей (Luscinia cyane (Pallas 1776)), соловей-свистун (Luscinia sibilans (Swinhoe 1863)), оливковый дрозд (Turdus obscurus (Gmelin 1789)), рябинник (Turdus pilaris (Linnaeus 1758)), чиж (Spinus spinus (Linnaeus 1758)). Они впервые зарегистрированы в пределах Колымского нагорья на удалении 300–800 км от известных ранее границ основного ареала этих видов (табл. 2). Большинство из них (n = 7) обнаружено к северу от основной области своего распространения, и только зимняк и халей – к югу. Почти все указанные виды встречены в лесных массивах речных долин горно-таежного пояса, и лишь зимняк – в гольцовом поясе.
Таблица 2.
География встреч видов птиц, зарегистрированных за пределами известных границ гнездовых ареалов
| Виды | Географические координаты 11 обследованных пунктов южных отрогов Колымского нагорья | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| хребет Черге | перевал Лошкалах | перевал Гаврюшка | перевал Кулу | р. Омчуг | р. Салтахан | горы Делурэкчэн | Майман- джинский хребет | р. Герба | р. Кинжал | р. Колыма | |
| 62°42′ с.ш. 148°50′ в.д. |
62°38′ с.ш. 147°23′ в.д. |
62°27′ с.ш. 147°18′ в.д. |
61°47′ с.ш. 147°45′ в.д. |
61°09′ с.ш. 149°30′ в.д. |
60°45′ с.ш. 149°56′ в.д. |
60°26′ с.ш. 150°58′ в.д. |
61°09′ с.ш. 152°07′ в.д. |
61°43′ с.ш. 152°26′ в.д. |
62°17′ с.ш. 151°57′ в.д. |
62°28′ с.ш. 152°13′ в.д. |
|
| Зимняк | + | ||||||||||
| Обыкновенный канюк | + | ||||||||||
| Халей | + | + | |||||||||
| Зеленая пеночка | + | + | + | + | + | + | + | + | |||
| Корольковая пеночка | + | + | + | + | + | + | + | + | + | + | + |
| Синий соловей | + | + | + | + | |||||||
| Соловей-свистун | + | + | |||||||||
| Оливковый дрозд | + | + | + | ||||||||
| Рябинник | + | ||||||||||
| Чиж | + | + | + | + | |||||||
Наши наблюдения подтверждают данные Кищинского (1968) о пребывании зеленой и корольковой пеночек на Колымском нагорье. Кроме этого, нами установлено, что корольковая пеночка не только повсеместно распространена по южным отрогам Колымского нагорья, но и, как показали наши исследования 2017 г., проникает значительно северо-восточнее Колымского нагорья – вплоть до северных окраин Корякского нагорья (Романов и др., 2018). В обследованных нами в 2016 г. южных частях Колымского нагорья для зеленой и корольковой пеночек обнаружены значительные по площади очаги устойчивого гнездования с высоким (или относительно высоким) обилием, что позволяет считать их частями основной области гнездования. Для других видов выявлены лишь локальные, вероятно, изолированные территориальные группировки, площадь обитания которых не превышает 50–100 км2 и которые образуют самую окраину ареала вида в Северо-Восточной Азии. Высока вероятность того, что это области спорадического гнездования или области ареала, где граница имеет явно выраженный пульсирующий характер.
Обилие видов в пунктах, где они встречены впервые, различно. Чирок-трескунок (Anas querquedula (Linnaeus 1758)), большой улит (Tringa nebularia (Gunnerus 1767)), сибирский дрозд (Zoothera sibirica (Pallas 1776)) были редки. Корольковая пеночка повсеместно многочисленна в обследованных частях юга Колымского нагорья. Чиж единичен или редок во всех пунктах встреч (n = 5). Численность синего соловья в каждом пункте встречи (n = 4) составила от 3 до 5 территориальных пар.
В процессе изучения трендов пространственного изменения обилия птиц установлено также, например, что обилие восточного подвида щура (Pinicola enucleator kamtschatkensis (Dybowski 1883)) последовательно увеличивается с запада на восток от хребта Черского (0.04 особи/км2) через Колымское нагорье (2 особи/км2) к Корякскому нагорью (71.9 особи/км2) (Романов и др., 2018). Максимально высокие показатели обилия этого вида на северо-восточной окраине ареала обусловлены тем, что Корякское нагорье охватывает оптимум ареала кедрового стланика, заросли которого являются излюбленным гнездовым местообитанием, а его орешки – основным кормом щура.
Вероятно, существующая динамика границ ареалов некоторых видов птиц указывает на продолжение расселения видов и формирования авифауны гор Северо-Восточной Азии. Недостаточный объем наблюдений не позволяет сделать репрезентативные выводы о положительных трендах динамики северных границ ареалов. Однако в пользу этого свидетельствует тот факт, что благодаря видам иммигрантам, расселяющимся из более южных областей, за несколько прошедших десятилетий видовое разнообразие плато Путорана и субарктических гор Якутии увеличилось на 6% (Романов, 2013).
Формирование вертикальной неоднородности авифауны
В соответствии с господствующим типом высотной поясности, в южных отрогах Колымского нагорья дифференцированы сменяющие друг друга с высотой авифауны горно-таежного, подгольцового и гольцового поясов. С высотой видовое разнообразие сокращается. Максимально разнообразна авифауна горно-таежного пояса, занимающего горные долины и нижние части горных склонов. С увеличением высоты, в условиях снижения теплообеспеченности и сокращения продолжительности бесснежного периода в подгольцовом поясе, видовое богатство птиц понижается. В экстремальных условиях гольцового пояса формируется наиболее бедная гольцовая авифауна. Условия среды на практически безжизненных горных вершинах соответствуют условиям зональных арктических тундр и полярных пустынь.
В южных отрогах Колымского нагорья гнездовая авифауна горно-таежного пояса насчитывает 68 (92%), подгольцового пояса – 33 (45%), гольцового пояса – 15 (20%) видов. Как и в других горных системах Северной Азии (Романов, 2013), видовой состав авифауны обсуждаемого региона с высотой меняется постепенно. При наличии в авифауне каждого вертикального пояса своих специфичных видов, авифауны двух соседних поясов имеют в своем составе много общих видов. Из 68 видов птиц, гнездящихся в горно-таежном поясе, и 33 видов – в подгольцовом, 31 вид является общим для авифаун обоих поясов. Из 33 видов птиц, гнездящихся в подгольцовом поясе, и 15 видов – в гольцовом поясе, 10 видов являются общими для авифаун этих поясов. Столько же общих видов отмечено для горно-таежного и гольцового поясов. Коэффициент фаунистического сходства горно-таежного и подгольцового поясов южных отрогов Колымского нагорья составляет 61(44)%, подгольцового и гольцового – 42(26)%, горно-таежного и гольцового – 24(14)%.
В южных отрогах Колымского нагорья 41 вид (55% местной гнездовой авифауны) обитает только в одном высотном поясе, не заходя в соседние. Из числа таких видов горно-таежному поясу свойственны 36 (перепелятник (Accipiter nisus (Linnaeus 1758)), белая куропатка (Lagopus lagopus (Linnaeus 1758)), желна (Dryocopus martius (Linnaeus 1758)), теньковка (Phylloscopus collybita (Vieillot 1817)), синий соловей, чиж, овсянка-ремез (Ocyris rusticus (Pallas 1776)) и др.), подгольцовому поясу – 1 (обыкновенный канюк), гольцовому поясу – 4 вида (зимняк, большой песочник, гольцовый конек (Anthus rubescens (Tunstall 1771)), альпийская завирушка). Одновременно два высотных пояса в южных отрогах Колымского нагорья осваивают 24 вида (32% местной гнездовой авифауны). Два нижних пояса обследованного региона населяют 21 вид птиц, в том числе, сибирский пепельный улит (Heteroscelus brevipes (Vieillot 1816)), глухая кукушка, сибирский жулан (Lanius cristatus (Linnaeus 1758)), кедровка, зеленая пеночка, зарничка, синехвостка, бурый дрозд (Turdus eunomus (Temminck 1831)), щур, овсянка-крошка и др. Зарегистрирован только один вид, одновременно населяющий два верхних пояса – обыкновенная каменка (Oenanthe oenanthe (Linnaeus 1758)). Во всех трех высотных поясах распространены 9 видов (12% местной гнездовой авифауны): тундряная куропатка, обыкновенная кукушка, пятнистый конек, горная трясогузка, ворон, таловка, корольковая пеночка, соловей-красношейка и обыкновенная чечевица. Ареалы видов, обитающих в широком диапазоне высот (не менее двух высотных поясов), имеют явно выраженный трехмерный характер.
В составе гнездовых авифаун горно-таежного пояса всех (или почти всех) обследованных пунктов южных отрогов Колымского нагорья 14 общих видов, подгольцового – 5, гольцового – 2 вида. Среди видов, повсеместно гнездящихся в горно-таежном поясе – горная трясогузка, кедровка, таловка, зарничка, корольковая и бурая пеночки, соловей-красношейка, синехвостка и др., в подгольцовом поясе – обыкновенная кукушка, пятнистый конек, таловка, корольковая пеночка и обыкновенная чечевица, в гольцовом поясе – тундряная куропатка и обыкновенная кукушка.
Анализ степени общности гольцовых, подгольцовых и горно-таежных авифаун всех обследованных частей юга Колымского нагорья (n = 11), проведенный с использованием коэффициентов фаунистической общности (КФО) Серенсена и Жаккара, выявил следующие закономерности. Максимальное сходство авифаун обследованных пунктов Колымского нагорья проявляется на уровне горно-таежного пояса – 63(46)–81(68)%), несколько меньшее на уровне гольцов – 44(29)–52(38)%) и минимальное в средней части высотного профиля на уровне подгольцов – 26(17)–33(24)%). В целом, однородная горно-таежная авифауна в современном ее виде сформировалась в пределах единой области Северо-Восточной Азии с повсеместным господством северо-таежных лесов и гипоарктических редколесий. В условиях непрерывности лесных ландшафтов и повсеместной сопряженности горных и равнинных типов лесов, подавляющее большинство видов осваивает их повсеместно, так как не имеет непреодолимых преград, препятствующих расселению. Раздробленность, взаимная удаленность и изоляция участков подгольцовых и гольцовых поясов, в совокупности с более разнообразными и нестабильными экологическими условиями, обусловила меньшее взаимное сходство региональных авифаун на уровне каждого из этих поясов. Выявленные различия уровня общности соответствующих авифаун указывают на определенную автономность формирования авифауны на уровне каждого высотно-ландшафтного пояса.
Фаунистическая и зонально-ландшафтная структура авифауны
Авифауна южных отрогов Колымского нагорья гетерогенна по происхождению. Она формируется видами 7 типов фауны (Штегман, 1938), наиболее значимы из которых во всех высотных поясах, элементы сибирского фаунистического комплекса (33–49%) и широкораспространенные виды (27–37%). Представители китайского типа фауны играют второстепенную роль (12–20%) в местных сообществах птиц. При этом они привносят определeнную специфику в формирование качественного разнообразия видового состава авифауны каждого высотно-ландшафтного пояса (табл. 3).
Таблица 3.
Соотношение фаунистических комплексов в гнездовой авифауне южных отрогов Колымского нагорья (%), число видов
| Фаунистический комплекс | Число видов | |||||
|---|---|---|---|---|---|---|
| горно-таежный пояс | подгольцовый пояс | гольцовый пояс | ||||
| абс. | % | абс. | % | абс. | % | |
| Арктический | 2 | 3 | 1 | 3 | 2 | 13 |
| Сибирский | 30 | 44 | 16 | 49 | 5 | 33 |
| Сибирско-американский | 1 | 1 | – | – | – | – |
| Европейский | 2 | 3 | – | – | – | – |
| Тибетский | – | – | – | – | 1 | 7 |
| Китайский | 8 | 12 | 5 | 15 | 3 | 20 |
| Широкораспространенные | 25 | 37 | 11 | 33 | 4 | 27 |
Южные отроги Колымского нагорья лежат в пределах подзоны северной тайги, и региональная авифауна закономерно испытывает влияние элементов авифауны южной и средней тайги, а также лесотундры и южной тундры. Это обусловливает неоднородность авифауны региона по сочетанию формирующих ее представителей 7 зонально-ландшафтных групп, из которых наиболее значимы на всех высотах широкораспространенные (33–41%), бореальные (20–31%) и бореально-гипоарктические (7–24%) виды. Доля арктоальпийских (тундряная куропатка, обыкновенная каменка) и альпийских (большой песочник, гольцовый конек, альпийская завирушка) видов суммарно существенна (33%) в формировании сообществ птиц гольцового пояса. Именно они, наряду с еще некоторыми видами, экологически тесно связанными с горными потоками (каменушка (Histrionicus histrionicus (Linnaeus 1758)), сибирский пепельный улит, горная трясогузка), определяют в процессе формирования качественного разнообразия авифауны южных отрогов Колымского нагорья ее горную специфику (табл. 4).
Таблица 4.
Соотношение зонально-ландшафтных групп в гнездовой авифауне южных отрогов Колымского нагорья (%), число видов
| Зонально-ландшафтная группа | Число видов | |||||
|---|---|---|---|---|---|---|
| горно-таежный пояс | подгольцовый пояс | гольцовый пояс | ||||
| абс. | % | абс. | % | абс. | % | |
| Альпийские | – | – | – | – | 3 | 20 |
| Арктоальпийские | 1 | 1 | 2 | 6 | 2 | 13 |
| Гемиаркты | – | – | – | – | 1 | 7 |
| Гипоаркты | 2 | 3 | 1 | 3 | – | – |
| Бореально-гипоарктические | 16 | 24 | 8 | 24 | 1 | 7 |
| Бореальные | 21 | 31 | 10 | 30 | 3 | 20 |
| Широкораспространенные | 28 | 41 | 12 | 37 | 5 | 33 |
Пространственная структура населения птиц
В южных отрогах Колымского нагорья с высотой поступательно сокращаются видовое богатство, плотность населения птиц, обилие абсолютного большинства видов. Плотность населения птиц в горно-таежном поясе различных частей южных отрогов Колымского нагорья лежит в интервале 176–674 особей/км2, в подгольцовом – 76–230 особей/км2, гольцовом – 20–72 особи/км2, составляя в среднем, соответственно – 312, 133, 40 особей/км2 (табл. 5). Как и в большинстве других горных регионов Северной Азии (Романов, 2013), в пределах высотного профиля южных отрогов Колымского нагорья основное сокращение плотности населения птиц происходит при переходе из подгольцового пояса в гольцовый (в 3–7 раз). Сокращение плотности населения при переходе из горно-таежного пояса в подгольцовый менее существенно (в 2–3 раза). Сокращение плотности населения птиц с высотой в южных отрогах Колымского нагорья соответствует общему понижению продуктивности сообществ животных от подножий к вершинам гор и демонстрирует относительную самостоятельность населения птиц каждого высотного пояса. Оценка населения птиц высотных поясов с помощью коэффициента сходства населения не только подтверждает эту самостоятельность, но и указывает на выраженную автономность формирования населения птиц разных высотных поясов. Это подтверждает то, что уровень сходства населения горно-таежного и подгольцового поясов составляет 33%, подгольцового и гольцового – 20%, горно-таежного и гольцового – 6%.
Таблица 5.
Население птиц южных отрогов Колымского нагорья по данным учетов 2016 г.
| Вид | Обилие, особей/км2 | Доля участия, % | ||||
|---|---|---|---|---|---|---|
| горно- таежный пояс | подгольцовый пояс | гольцовый пояс | горно- таежный пояс |
подгольцовый пояс | гольцовый пояс | |
| Кряква | 0.7 | – | – | 0.2 | – | – |
| Чирок-свистунок | 2 | – | – | 0.7 | – | – |
| Свиязь | 0.9 | – | – | 0.3 | – | – |
| Чирок-трескунок | 0.2 | – | – | 0.1 | – | – |
| Хохлатая чернеть | 0.2 | – | – | 0.1 | – | – |
| Каменушка | 0.8 | – | – | 0.3 | – | – |
| Обыкновенный гоголь | 0.2 | – | – | 0.1 | – | – |
| Длинноносый крохаль | 0.3 | – | – | 0.1 | – | – |
| Тетеревятник | 0.1 | – | – | 0.02 | – | – |
| Перепелятник | 0.4 | – | – | 0.1 | – | – |
| Зимняк | – | – | 0.8 | – | – | 1.9 |
| Обыкновенный канюк | – | 0.1 | – | – | 0.1 | – |
| Чеглок | 0.7 | 0.1 | – | 0.2 | 0.1 | – |
| Обыкновенная пустельга | 0.01 | – | 0.1 | 0.004 | – | 0.2 |
| Белая куропатка | 0.5 | – | – | 0.2 | – | – |
| Тундряная куропатка | 0.1 | 4.7 | 3.7 | 0.03 | 3.5 | 9.3 |
| Рябчик | 2.1 | – | – | 0.7 | – | – |
| Черныш | 0.6 | – | – | 0.2 | – | – |
| Фифи | 0.1 | – | – | 0.03 | – | – |
| Большой улит | 0.9 | 0.1 | – | 0.3 | 0.1 | – |
| Сибирский пепельный улит | 0.7 | 0.5 | – | 0.2 | 0.4 | – |
| Перевозчик | 3.5 | 0.3 | – | 1.1 | 0.2 | – |
| Большой песочник | – | – | 3.7 | – | – | 9.3 |
| Бекас | 0.2 | – | – | 0.1 | – | – |
| Озерная чайка | 0.04 | – | – | 0.01 | – | – |
| Халей | 0.1 | – | – | 0.02 | – | – |
| Сизая чайка | 0.6 | 0.1 | – | 0.2 | 0.1 | – |
| Речная крачка | 0.5 | – | – | 0.2 | – | – |
| Обыкновенная кукушка | 2 | 2.4 | 1.2 | 0.7 | 1.8 | 3 |
| Глухая кукушка | 0.8 | 0.8 | – | 0.3 | 0.6 | – |
| Желна | 0.03 | – | – | 0.01 | – | – |
| Полевой жаворонок | 0.01 | – | – | 0.004 | – | – |
| Пятнистый конек | 4.2 | 10.4 | 0.8 | 1.3 | 7.9 | 1.9 |
| Гольцовый конек | – | – | 1.7 | – | – | 4.3 |
| Берингийская желтая трясогузка | 0.4 | – | – | 0.1 | – | – |
| Горная трясогузка | 7.6 | 12.7 | 10.5 | 2.4 | 9.6 | 26.2 |
| Белая трясогузка | 2.7 | 2.4 | – | 0.9 | 1.8 | – |
| Сибирский жулан | 0.2 | 0.1 | – | 0.1 | 0.1 | – |
| Кедровка | 6.6 | 3.8 | – | 2.1 | 2.9 | – |
| Черная ворона | 1 | 0.3 | – | 0.3 | 0.2 | – |
| Ворон | 0.6 | 0.5 | 0.3 | 0.2 | 0.4 | 0.7 |
| Свиристель | 0.5 | – | – | 0.2 | – | – |
| Альпийская завирушка | – | – | 3 | – | – | 7.5 |
| Сибирская завирушка | 1.2 | 2.9 | – | 0.4 | 2.2 | – |
| Пятнистый сверчок | 0.8 | – | – | 0.3 | – | – |
| Пеночка-теньковка | 2.6 | – | – | 0.8 | – | – |
| Пеночка-таловка | 20.2 | 18.9 | 2.2 | 6.5 | 14.2 | 5.6 |
| Зеленая пеночка | 3.3 | 1.1 | – | 1.1 | 0.8 | – |
| Пеночка-зарничка | 63.8 | 4.4 | – | 20.5 | 3.3 | – |
| Корольковая пеночка | 89.4 | 29.3 | 2.2 | 28.7 | 22 | 5.6 |
| Бурая пеночка | 21.8 | 12.7 | – | 7 | 9.5 | – |
| Малая мухоловка | 0.8 | – | – | 0.3 | – | – |
| Черноголовый чекан | 3 | 2 | – | 1 | 1.5 | – |
| Обыкновенная каменка | – | 3.2 | 4.7 | – | 2.4 | 11.8 |
| Соловей-красношейка | 6 | 4.2 | 1.5 | 1.9 | 3.2 | 3.7 |
| Синий соловей | 1.2 | – | – | 0.4 | – | – |
| Соловей-свистун | 0.2 | – | – | 0.1 | – | – |
| Синехвостка | 7.6 | 1.3 | – | 2.4 | 1 | – |
| Оливковый дрозд | 0.2 | – | – | 0.1 | – | – |
| Бурый дрозд | 0.7 | 0.1 | – | 0.2 | 0.1 | – |
| Рябинник | 0.04 | – | – | 0.01 | – | – |
| Сибирский дрозд | 0.2 | – | – | 0.1 | – | – |
| Буроголовая гаичка | 3 | – | – | 1 | – | – |
| Сероголовая гаичка | 0.2 | – | – | 0.1 | – | – |
| Вьюрок | 13 | 1.2 | – | 4.2 | 0.9 | – |
| Чиж | 1.4 | – | – | 0.5 | – | – |
| Обыкновенная чечетка | 0.1 | 0.1 | – | 0.03 | 0.1 | – |
| Обыкновенная чечевица | 18.2 | 7.9 | 3.7 | 5.8 | 5.9 | 9.3 |
| Сибирская чечевица | 0.1 | 0.3 | – | 0.03 | 0.2 | – |
| Щур | 0.2 | 2.1 | – | 0.1 | 1.5 | – |
| Полярная овсянка | 0.1 | – | – | 0.03 | – | – |
| Овсянка-ремез | 2.4 | – | – | 0.8 | – | – |
| Овсянка-крошка | 7.2 | 2.2 | – | 2.3 | 1.6 | – |
| Всего | 312 | 133 | 40 | 100 | 100 | 100 |
Максимальное сходство населения птиц обследованных районов южных отрогов Колымского нагорья проявляется на уровне горно-таежного пояса – 27–33%, несколько меньшее – на уровне гольцового пояса 19–25% и минимальное сходство – в пределах подгольцового пояса – 7–12%.
В населении птиц горно-таежного пояса большинства обследованных пунктов (n = 8) численно доминируют корольковая пеночка и зарничка, в некоторых (n = 3) – таловка и бурая пеночка. В населении птиц подгольцового пояса более половины обследованных пунктов (n = 6) численно доминируют корольковая пеночка и таловка, в некоторых (n = 3) – пятнистый конек, бурая пеночка и обыкновенная чечевица. В гольцовом поясе обследованных пунктов в различном сочетании численно преобладают обыкновенная каменка, горная трясогузка, большой песочник.
Среди субдоминантов в населении птиц горно-таежного пояса 12 видов: бурая пеночка, таловка, обыкновенная чечевица, вьюрок, синехвостка, горная трясогузка, овсянка-крошка, кедровка, соловей-красношейка, пятнистый конек, перевозчик, зеленая пеночка. Среди субдоминантов горно-таежного пояса 15 видов: горная трясогузка, бурая пеночка, пятнистый конек, обыкновенная чечевица, тундряная куропатка, зарничка, соловей-красношейка, кедровка, обыкновенная каменка, сибирская завирушка, обыкновенная кукушка, белая трясогузка, овсянка-крошка, щур, черноголовый чекан (Saxicola torquata (Linnaeus 1766)). В гольцовом поясе группу видов субдоминантов в различном сочетании формируют обыкновенная чечевица, большой песочник, тундряная куропатка, альпийская завирушка, таловка, корольковая пеночка, гольцовый конек, соловей-красношейка, обыкновенная кукушка, пятнистый конек, зимняк.
ЗАКЛЮЧЕНИЕ
Получена, обобщена и проанализирована подробная оригинальная современная информация о видовом составе и характере регионального и высотно-поясного размещения птиц в южных отрогах Колымского нагорья. Представлены ранее неизвестные данные по структуре фауны и населения птиц, существенно расширяющие знания об авифауне обширной горной области Северо-Восточной Азии.
В обследованных нами пунктах (n = 11) южных отрогов Колымского нагорья зарегистрировано 23–44 вида птиц. В целом, гнездовая авифауна всего обследованного региона насчитывает 74 вида, среди которых 16 (22%) впервые обнаружены здесь на расстоянии от 20 до 800 км от известных границ гнездовых ареалов.
Показатель представленности гнездовой авифауны региона в отдельных обследованных пунктах 31–59%. Уровень фаунистической общности обследованных районов южных отрогов Колымского нагорья 45(29)–75(60)%, что свидетельствует в пользу однородности авифауны рассматриваемой части материковой суши. Структура гнездовой авифауны южных отрогов Колымского нагорья соответствует зональным и ландшафтным особенностям рассматриваемой части Северо-Восточной Азии, а ее качественная однородность во всех обследованных пунктах обусловлена повсеместным преобладанием представителей одних и тех же отрядов (воробьеобразные 59–83%, ржанкообразные 11–18%), фаунистических комплексов (сибирский – 33–49%, китайский – 12–20%) и зонально-ландшафтных групп (бореальные – 20–31%, бореально-гипоарктические – 7–24%).
Общность структуры фауны птиц поддерживается в горизонтальной плоскости видами, широко распространенными одновременно в большинстве обследованных регионов, а в вертикальной плоскости – видами, одновременно населяющими два, обычно смежных, высотно-ландшафтных пояса. Большинство видов птиц, одновременно гнездящихся во всех обследованных частях юга Колымского нагорья и формирующих общее фаунистическое ядро, широко распространено в северной тайге, лесотундре и частично в южной тундре.
Фауна и население птиц юга Колымского нагорья формируется в системе общих зональных и высотно-поясных закономерностей. С высотой сокращаются видовое богатство, плотность населения птиц, обилие абсолютного большинства видов. В силу идентичности господствующих экологических условий, авифауна всех обследованных частей Колымского нагорья имеет единый характер высотно-поясной дифференциации, в соответствии с которым выделяются сменяющие друг друга с высотой авифауны горно-таежного, подгольцового и гольцового поясов.
В южных отрогах Колымского нагорья гнездовая авифауна горно-таежного пояса насчитывает 68 (92%), подгольцового – 33 (45%), гольцового – 15 (20%) видов. Коэффициент сходства авифаун горно-таежного и подгольцового поясов обсуждаемого региона 61 (44)%, подгольцового и гольцового поясов 42 (26)%, горно-таежного и гольцового поясов 24 (14)%.
В южных отрогах Колымского нагорья 55% всех гнездящихся видов птиц ограничены в своем распространении только одним высотным поясом. В широком диапазоне высот, охватывающем не менее двух высотных поясов, обитает 45% видов, два пояса осваивает 32%, а три пояса – 12% всех гнездящихся видов. Ареалы видов, обитающих в широком диапазоне высот, охватывающем не менее двух высотных поясов, имеют явно выраженный трехмерный характер.
Плотность населения птиц в горно-таежном поясе различных частей южных отрогов Колымского нагорья 176–674 особей/км2, в подгольцовом – 76–230, гольцовом – 20–72 особей/км2, в среднем, соответственно, 312, 133, 40 особей/км2. Повсеместно наиболее существенное (в 3–7 раз) сокращение плотности населения птиц наблюдается при переходе из подгольцового пояса в гольцовый.
Горную специфику авифауны обследованных частей юга Колымского нагорья определяют виды, экологически тесно связанные с сухопутными или водно-околоводными элементами альпинотипного ландшафта на всем пространстве своего ареала (каменушка, сибирский пепельный улит, большой песочник, гольцовый конек, альпийская завирушка) или значительной его части (тундряная куропатка, горная трясогузка, обыкновенная каменка).
Список литературы
Андреев А.В., Докучаев Н.Е., Кречмар А.В., Чернявский Ф.Б., 2005. Наземные позвоночные Северо-Востока России. Магадан: СВНЦ ДВО РАН. 313 с.
Андреев А.В., Кондратьев А.В., Потапов Е.Р., 2015. Орнитофауна нижнеколымских тундр: многолетняя динамика на фоне климатических перемен. Сообщение 1. Динамика состава нижнеколымской орнитофауны в XX в. и первом десятилетии XXI в. // Вестник СВНЦ ДВО РАН. № 1. С. 49–59.
Андреев А.В., Кондратьев А.В., Потапов Е.Р., 2015а. Орнитофауна нижнеколымских тундр: многолетняя динамика на фоне климатических перемен. Сообщение 2. Статус, распространение и численность индикаторных видов // Вестник СВНЦ ДВО РАН. № 2. С. 57–68.
Баранов А.А., 2007. Пространственно-временная динамика биоразнообразия птиц Алтай-Саянского экорегиона. Автореф. дис. … докт. биол. наук. Красноярск. 49 с.
Вартапетов Л.Г., Гермогенов Н.И., 2011. Орнитофаунистическое районирование Средней и Восточной Сибири // Труды ИСиЭЖ СО РАН “Птицы Сибири: структура и динамика фауны, населения и популяций”. Отв. редактор: Вартапетов Л.Г. М.: Товарищество научных изданий КМК. С. 7 ̶ 28.
Гвоздецкий Н.А., Михайлов Н.И., 1987. Физическая география СССР. Азия. М.: Мысль. 512 с.
Голубчиков Ю.Н., 1996. География горных и полярных стран. М.: МГУ. 304 с.
Кищинский А.А., 1968. Птицы Колымского нагорья. М.: Наука. 184 с.
Кищинский А.А., 1988. Орнитофауна северо-востока Азии. М.: Наука. 288 с.
Коблик Е.А., Редькин Я.А., Архипов В.Ю., 2006. Список птиц Российской Федерации. М.: Товарищество научных изданий КМК. 256 с.
Куваев В.Б., 2006. Флора субарктических гор Евразии и высотное распределение ее видов. М.: Товарищество научных изданий КМК. 568 с.
Наумов Р.Л., 1964. Птицы в очагах клещевого энцефалита Красноярского края // Автореф. дис. … канд. биол. наук. М: Институт медицинской паразитологии и тропической медицины им. Е.И. Марциновского, 19 с.
Песенко Ю.А., 1982. Принципы и методы количественного анализа в фаунистических исследованиях. М.: Наука. 287 с.
Равкин Ю.С., 1967. К методике учета птиц лесных ландшафтов // Природа очагов клещевого энцефалита на Алтае. Отв. ред. Максимов А.А. Новосибирск: Наука. С. 66–75.
Романов А.А., 2013. Авифауна гор Азиатской Субарктики: закономерности формирования и динамики // Русское общество сохранения и изучения птиц имени М.А. Мензбира. М. 360 с.
Романов А.А., Астахова М.А., Миклин Н.А., Шемякин Е.В., 2018. Высотно-поясная дифференциация населения птиц севера Корякского нагорья // Вестник Томского гос. ун-та. Биология. № 43. С. 136–158.
Рябицев В.К., 2014. Птицы Сибири: справочник–определитель. Т. 2. Москва– Екатеринбург: Кабинетный ученый. 452 с.
Сочава В.Б., 1980. Географические аспекты сибирской тайги. Новосибирск: Наука. 256 с.
Cтепанян Л.С., 2003. Конспект орнитологической фауны России и сопредельных территорий. М.: Наука. 727 с.
Чернов Ю.И., 1975. Природная зональность и животный мир суши. М.: Мысль. 222 с.
Чернов Ю.И., 1978. Структура животного населения Субарктики. М.: Наука. 167 с.
Чернов Ю.И., 2008. Экология и биогеография. Избранные труды. М.: Товарищество научных изданий КМК. 580 с.
Штегман Б.К., 1938. Основы орнитогеографического деления Палеарктики // Фауна СССР. Птицы. Т. 1. Вып. 2. Отв. ред. Зернов С.А. М.-Л.: АН СССР. 157 с.
Юрцев Б.А., 1968. Флора Сунтар-Хаята // Проблемы истории высокогорных ландшафтов Северо-Востока Сибири. Л.: Наука. 235 с.
Graham C.H., Carnaval A.C., Cadena C.D., Zamudio K.R., Roberts T.E., et al., 2014. The origin and maintenance of montane diversity: integrating evolutionary and ecological processes // Ecography. V. 37. № 8. P. 711–719.
McCain C., 2009. Vertebrate range sizes indicate that mountains may be 'higher' in the tropics // Ecology Letters. V. 12 (6). P. 1043–1052.
Ruggiero A., Hawkins B.A., 2008. Why do mountains support so many species of birds? // Ecography. V. 31. № 3. P. 306–315.
The EBCC Atlas of European breeding birds: Their distribution and abundance, 1997. Hagemmeijer W.J.M., Blair M.J., eds. London: T&A D Poyser Publ. 903 p.
Tomkovich P.S., 1997. Breeding distribution, migrations and conservation status of the Great Knot Calidris tenuirostris in Russia // Emu. V. 97. № 4. P. 265–282.
Дополнительные материалы отсутствуют.
Инструменты
Зоологический журнал