

Зоологический журнал, 2019, T. 98, № 9, стр. 965-977
Центрохелидные солнечники (Centroplasthelida Febvre-Chevalier et Febvre 1984) Вьетнама
К. И. Прокина a, b, *, Л. В. Радайкина a, А. П. Мыльников a
a Институт биологии внутренних вод им. И.Д. Папанина РАН
152742 Ярославская обл., Некоузский р-н,
пос. Борок, Россия
b Зоологический институт РАН
199034 Санкт-Петербург, Россия
* E-mail: kristin892@mail.ru
Поступила в редакцию 11.12.2018
После доработки 04.03.2019
Принята к публикации 04.03.2019
Аннотация
Исследована морфология покровных неорганических чешуек 10 видов центрохелидных солнечников из разнотипных местообитаний Вьетнама. Все обнаруженные виды – новые для Вьетнама. Радиальные чешуйки Acanthocystis turfacea отличаются характерными конически заостренными концами фурок и большой длиной фурок длинных радиальных чешуек относительно длины ствола. Исследованные пластинчатые чешуйки клеток Pterocystis tropica соответствуют подвиду P. tropica paucistriata, а радиальные чешуйки – подвиду P. tropica tropica. Исследованные пластинчатые чешуйки вида Raineriophrys erinaceoides undulata не имеют радиальных прорезей в отличие от первоописания этого подвида. Описаны два новых вида центрохелидных солнечников. Pterocystis vietnamica sp. n. характеризуется листовидными радиальными чешуйками c поперечными прорезями на латеральных крыльях и крупным сферическим субапикальным выпячиванием. Pseudoraphidocystis dentata sp. n. характеризуется наличием маргинальных зубцов на краях воронковидных радиальных чешуек.
Центрохелидные солнечники – группа амебоидных протистов, занимающая положение внутри Haptista Cavalier-Smith, 2003 вместе с гаптофитовыми водорослями (Adl et al., 2019). Центрохелидные солнечники являются важным компонентом микробентоса морских и пресноводных экосистем, осуществляя роль пассивных хищников, поедающих простейших, водорослей, и даже микроскопических многоклеточных (Микрюков, 2002).
Практически все центрохелидные солнечники (за исключением рода Oxnerella) имеют наружные элементы клетки – тангенциальные и/или радиальные кремниевые (реже органические) чешуйки. Изучение морфологии таких покровных структур является исключительно важным для видовой идентификации центрохелид и необходимо при фаунистических исследованиях. Однако проведение таких работ возможно лишь с применением особых методов – сканирующей (СЭМ) и трансмиссионной (ТЭМ) электронной микроскопии. На сегодняшний день известно немногим более 100 видов центрохелидных солнечников, имеющих уникальные морфологические особенности чешуек, однако представляется возможным, что большая часть видов центрохелид еще не изучена, т.к. постоянно описываются новые виды (Леонов, 2010; Cavalier-Smith, Heyden, 2007; Prokina et al., 2019; Tikhonenkov, Mylnikov, 2011; Zlatogursky, 2015; Zlatogursky et al., 2017).
Разнообразие центрохелид и их распределение по всему миру изучены крайне неравномерно, при этом данные по видовому составу этой группы протистов полностью отсутствуют для многих стран, в том числе и для Вьетнама. Также отмечается, что морские и соленые континентальные местообитания изучены значительно хуже пресноводных. При этом накопление подобных данных может позволить в дальнейшем ответить на вопросы о космополитизме и эвригалинности данной группы микроорганизмов, пока предполагаемые, но не подтвержденные в настоящее время (Plotnikov, Gerasimova, 2017; Mikrjukov, 1999; Leonov, 2010). Накопление данных о морфологии чешуек солнечников из различных регионов мира позволит, совместно с применением методов молекулярно-филогенетического анализа, лучше понять границы внутривидовой изменчивости для этих организмов.
Цель нашей работы заключается в изучении морфологии покровных структур (чешуек) клеток и выявлении видового состава центрохелидных солнечников в некоторых пресноводных и морских местообитаниях Вьетнама методами трансмиссионной и сканирующей электронной микроскопии.
МАТЕРИАЛ И МЕТОДЫ
Исследованы пресноводные и морские местообитания Вьетнама в январе и феврале 2018. Отбор проб производился Н.Р. Крупицкой (МГУ) из трех пресноводных объектов в пределах заповедника Донгнай [Dong Nai] в провинции Донгнай: р. Донгнай [Dong Nai River], 11°25′57.0″ с.ш., 107°25′51.0″ в.д.; озеро Бау Сау [Bau Sau], или крокодилово озеро, 11°27′42.7″ с.ш., 107°20′16.4″ в.д.; небольшой безымянный пруд, 11°25′75″ с.ш., 107°25′74″ в.д.; а также морские местообитания в окрестностях г. Нячанг [Nha Trang], провинция Кханьхоа [Khanh Hoa] 12°14′25.3″ с.ш., 109°12′13.9″ в.д.
Пробы воды с частью придонного грунта помещали в пластиковые пробирки емкостью 15 мл каждая и транспортировали в лабораторию при температуре 4°С. В лабораторных условиях пробы помещали в чашки Петри, обогащали суспензией бактерий Pseudomonas fluorescens Migula 1895, культурой жгутиконосцев Bodo saltans Ehrenberg 1838 (для пресной среды) и Procryptobia sorokini (Zhukov 1975) Frolov, Karpov et Mylnikov 2001 (для морской среды), выдерживали в термостате при температуре 22°С в полной темноте в течение нескольких дней для выявления скрытого видового разнообразия. С целью обнаружения солнечников и приготовления препаратов для электронной микроскопии использовали световые микроскопы AxioScope A1 (Carl Zeiss, Германия) и Olympus CKX 41 (Olympus, Япония). Полученные с помощью аналоговой видеокамеры AVT HORN MC1009/S изображения оцифровывали и хранили в виде файлов формата AVI. Препараты для электронной микроскопии подготавливали согласно общепринятым методикам (Микрюков, 2002; Moestrup, Thomsen, 1980) и просматривали в трансмиссионном электронном микроскопе JEM-1011 (Jeol, Япония) и сканирующем электронном микроскопе JSM–6510 LV (Joel, Япония).
РЕЗУЛЬТАТЫ
В исследованных водных объектах было выявлено 10 видов центрохелидных солнечников, два из которых – новые для науки. В работе использована современная макросистема эукариот (Adl et al., 2019), в которой не используются формальные ранги таксонов. Статус макротаксонов обозначен звездочками – чем их больше, тем ниже ранг таксона. Морфологическая характеристика изученных неорганических чешуек выявленных видов представлена ниже.
DIAPHORETICKES Adl et al. 2012
*Haptista Cavalier-Smith 2003
**Centroplasthelida Febvre-Chevalier et Febvre 1984
***Pterocystida Cavalier-Smith et von der Heyden 2007 emend. Shishkin et Zlatogursky 2018
****Raphidista Shishkin et Zlatogursky 2018
*****Choanocystidae Cavalier-Smith et von der Heyden 2007
Choanocystis perpusilla (Petersen et Hansen 1960) Siemensma 1991 [bas.: Acanthocystis perpusilla Petersen et Hansen 1960] (рис. 1A–1C)
Рис. 1.
Морфология чешуек центрохелид родов Choanocystis и Pterocystis (СЭМ): A–C – Ch. perpusilla, D–F – P. foliacea, G–J – P. tropica; а.гр – аксиальный гребень, вм – выемка основания радиальной чешуйки, л.к – латеральные крылья, о.ч – основание чешуйки, р.р – радиальные ребра, р.ч – радиальная чешуйка, стб – “стебелек”, ств – ствол радиальной чешуйки, фр – фурка. Масштаб (мкм): D – 10; A – 5; B–C, E–F, H–J – 1.

Материал. Две клетки (СЭМ) и 1 клетка (ТЕМ) из р. Донгнай, детрит и песок под камнем, 02.02.2018.
Морфология. Радиальные чешуйки состоят из полого цилиндрического ствола 3.29–8.73 мкм в длину, 0.16–0.20 мкм в диаметре, и плоского сердцевидного основания 1.19–1.69 мкм в диаметре. Основание имеет небольшую выемку с одного края, от которого отходит ствол. Дистальный конец ствола делится на 2 заостренные фурки. Овальные пластинчатые чешуйки 1.44–2.83 × 1.38–1.98 мкм, с вогнутой центральной частью и аксиальным гребнем (1.35–1.26 мкм в длину).
Замечания. Наиболее часто в литературе указывается меньшая длина радиальных чешуек, 1.4–5.6 мкм (Шатилович и др., 2010; Croome et al., 1987; Dürrschmidt, 1985), однако есть находки с длиной 8–10 мкм (Леонов, Мыльников, 2012). Размеры исследованных нами чешуек в наибольшей степени согласуются с размерами чешуек подвида Ch. cordiformis parvula, описанного Дюршмит (Dürrschmidt, 1987) и в последствии признанного младшим синонимом Ch. perpusilla.
Распространение. Пресные воды европейской части России (Леонов, Мыльников, 2012; Леонов, Плотников, 2009; Leonov, 2010), Эстонии (Mikrjukov, 1993a), Чили (Dürrschmidt, 1985; Prokina, Mylnikov, 2019), Шри-Ланки ([как Acanthocystis cordiformis parvula] Dürrschmidt, 1987). Морские воды Антарктики 35‰ (Croome et al., 1987); Балтийское море, 5–6‰ (Vørs, 1992); соленые внутренние воды европейской части России, 2–22‰ (Ермоленко, Плотников, 2013). Почвы Восточной Сибири (Шатилович и др., 2010).
****Pterista Shishkin et Zlatogursky 2018
*****Pterocystidae Cavalier-Smith et von der Heyden 2007
Pterocystis foliacea (Dürrschmidt 1985) Siemensma 1991 [bas.: Acanthocystis foliacea Dürrschmidt 1985] (рис. 1D–1F)
Материал. Две клетки (СЭМ) из небольшого безымянного пруда с опавшей листвой (детрит) близ реки Донгнай, 22.01.2018; четыре клетки (СЭМ) и три клетки (ТЭМ) из бентосных проб озера Бау Сау, 29.01.2018.
Морфология. Листовидные радиальные чешуйки состоят из полого цилиндрического ствола, базального и латеральных крыльев. Ствол 1.85–3.32 мкм в длину, латеральные крылья тянутся вдоль ствола, дистально сужаясь, в базальной части соединяются с базальным крылом, формируя субапикальное выпячивание, которое сильно сужаясь переходит в небольшой полый “стебелек”. Латеральные крылья в самой расширенной части 1.16–2.02 мкм шириной, стебелек 0.22–0.48 мкм длиной. Яйцевидные пластинчатые чешуйки 1.66–2.97 × 1.22–1.83 мкм, с центральным вдавлением, аксиальным гребнем и немного утолщенной маргинальной каймой.
Замечания. Размеры и морфология исследованных чешуек соответствуют размерам и морфологии подвида P. foliacea truncata, приводимые в литературе (Dürrschmidt, 1987a; Prokina et al., 2019).
Распространение. Пресные воды Украины (Gaponova, 2008), Нигерии (Wujek, Ogundipe, 2002), Австралии (Croome, 1987), Чили (Dürrschmidt, 1985; Prokina, Mylnikov, 2019), Новой Зеландии, Японии, Шри-Ланки (Dürrschmidt, 1987a). Соленые внутренние воды европейской части России, 16‰ (Plotnikov, Gerasimova, 2017), Черное море, 16–18‰ (Prokina et al., 2019).
Pterocystis tropica (Dürrschmidt 1987) Siemensma 1991 [bas.: Acanthocystis tropica Dürrschmidt 1987] (рис. 1G–1J)
Материал. Одна клетка (СЭМ) из реки Донгнай, детрит и песок под камнем, 02.02.2018.
Морфология. Радиальные чешуйки состоят из полого цилиндрического ствола, базального и латеральных крыльев. Ствол 3.26–7.43 мкм в длину, 0.16–0.26 мкм в диаметре, в базальной части изогнут дорсально, дистальный конец конически заострен. Латеральные крылья тянутся вдоль ствола на 2.36–4.35 мкм, в дистальной части узкие, 1.28–3.12 мкм в длину, проксимальная часть резко расширяется, почти прямоугольная в контуре, 1.19–1.78 мкм длиной, 1.07–1.55 мкм шириной, с вытянутыми дистально плечами. В базальной части латеральные крылья соединяются с перпендикулярно ориентированным базальным крылом и формируют ковшеобразное выпячивание. Крылья орнаментированы радиально отходящими от ствола ребрами. Овальные пластинчатые чешуйки 2.7–3.29 × 1.23–1.98 мкм, бесфактурные, за исключением аксиального гребня.
Замечания. Размеры исследованных чешуек в целом схожи с размерами чешуек, указываемых ранее. Дюршмит описывала 2 подвида: Acanthocystis tropica tropica и A. tropica paucistriata (Dürrschmidt, 1987a). Первый подвид отличается от второго значительно большим количеством радиальных ребер и хорошо развитыми плечами на латеральных крыльях радиальных чешуек, а также хорошо заметными радиальными прорезями пластинчатых чешуек. Исследованные нами радиальные чешуйки больше соответствуют подвиду A. tropica tropica, в то время как пластинчатые чешуйки схожи с таковыми у подвида A. tropica paucistriata.
Распространение. Пресные воды европейской части России (Prokina et al., 2017), Германии, Чили, Шри-Ланки (Dürrschmidt, 1987a).
Pterocystis vietnamica Prokina, Radaykina, Mylnikov sp. n. (рис. 2)
Рис. 2.
Морфология чешуек Pterocystis vietnamica sp. n. (СЭМ): A – чешуйки одной клетки, B, D–E – радиальные чешуйки, C – пластинчатые чешуйки, сб.в – субапикальное выпячивание, п.пр – поперечные прорези. Остальные обозначения как на рис. 1. Масштаб (мкм): A – 5; B–E – 1.

Материал. Исследованы пластинчатые и радиальные чешуйки трех клеток (СЭМ) из реки Донгнай, детрит и песок под камнем, 02.02.2018. Типовой материал (препараты для СЭМ) хранятся в лаборатории Микробиологии ИБВВ РАН. Типовой рис. 2.
Морфологическое описание. Детали строения протопласта неизвестны. Клетка покрыта листовидными радиальными и яйцевидными пластинчатыми чешуйками. Радиальные чешуйки состоят из полого цилиндрического ствола, базального и латеральных крыльев. Ствол слабо развит, 1.52–2.29 мкм в длину, 0.08 мкм в диаметре. Латеральные крылья тянутся вдоль ствола, дистально сужаясь к заостренному концу, в основании крылья соединяются с базальным крылом, формируя крупное сферическое субапикальное выпячивание 0.30–0.46 мкм в диаметре. Латеральные крылья 0.84–1.19 мкм в самой расширенной части, с поперечными прорезями, края имеют утолщенную маргинальную кайму (0.06–0.07 мкм в диаметре). Пластинчатые чешуйки 1.67–3.08 × 1.35–2.24 мкм, с центральным вдавлением и немного утолщенной маргинальной каймой (0.07–0.08 мкм в диаметре).
Дифференциальный диагноз. Данный вид схож с P. foliacea листовидной формой радиальных чешуек, однако в отличие от исследуемого вида, у P. foliacea в базальной части формируется тонкий “стебелек” вместо достаточно крупного сферического выпячивания. Также латеральные крылья нового вида имеют поперечные прорези. Небольшое округлое базальное выпячивание базального крыла характерно для Pseudoraphidocystis veliformis (Dürrschmidt 1985) Mikrjukov 1997, однако латеральные крылья последнего значительно шире и чешуйка почти округлая в контуре, вместо прорезей латеральные крылья несут радиально расходящиеся ребра, а субапикальное выпячивание слабо развито. Хорошо развитое базальное выпячивание, переходящее в полую ножку, характерно для видов Pterocystis pteracantha (Siemensma 1981) Siemensma et Roijackers 1988 и P. pulchra (Dürrschmidt 1985) Siemensma 1991, но эти виды отличаются от нового вида почти воронковидной формой радиальных чешуек, наличием радиальных ребер на латеральных крыльях и хорошо сформированной полой ножкой, имеющей уплощенное основание. Продольные прорези на латеральных крыльях свойственны виду Raineriophrys pteromorphos (Dürrschmidt 1987) Mikrjukov 2001, который, тем не менее, имеет и другие отличительные признаки: 1) латеральные крылья тянутся не по всей длине ствола; 2) ствол хорошо развит и оканчивается прямо срезанным, а не заостренным дистальным концом; 3) отсутствие базального выпячивания.
Этимология. Вид назван по месту находки – во Вьетнаме.
Raineriophrys erinaceoides (Petersen et Hansen 1960) ssp. undulata Dürrschmidt 1985 (рис. 3A–3D)
Рис. 3.
Морфология чешуек центрохелид рода Raineriophrys (A–D – СЭМ, E–H – ТЭМ): A–D – R. erinaceoides undulata, E–H– R. fortesca; к.в – ковшеобразное выпячивание. Остальные обозначения как на рис. 1. Масштаб (мкм): E – 10; A – 5; B–D, F–H – 1.

Материал. Одна клетка (СЭМ) из небольшого безымянного пруда с опавшей листвой (детрит) близ р. Донгнай, 22.01.2018.
Морфология. Радиальные чешуйки состоят из полого цилиндрического ствола, базального и латеральных крыльев. Ствол 7.55–13.26 мкм в длину, 0.29–0.38 мкм в диаметре, в базальной части изогнут дорсально, дистальная часть плавно сужается в заостренный конец. Латеральные крылья тянутся вдоль ствола на 1/2 его длины, дистально сужаясь, в базальной части соединяются с перпендикулярно ориентированным базальным крылом и формируют ковшеобразное выпячивание. Ширина крыльев в основании 1.36–2.03 мкм, длина 3.17–6.95 мкм. Края латеральных крыльев волнообразные. Яйцевидные пластинчатые чешуйки 4.02–4.29 × 1.96–2.44 мкм, с хорошо выраженным аксиальным гребнем и утолщенной маргинальной каймой.
Замечания. Исследованные нами чешуйки в значительной степени соответствуют первоописанию данного подвида (Dürrschmidt, 1987a). Однако, по данным Дюршмит, пластинчатые чешуйки имеют слабо развитые радиальные прорези, а размеры чешуек в первоописании значительно меньше (длина радиальных чешуек 4.3–5.0 мкм, длина пластинчатых чешуек 2.2–2.8 мкм). Отличием данного подвида от остальных (R. erinaceoides erinaceoides, R. erinaceoides sculpta) являются волнообразные края латеральных крыльев. Благодаря этому признаку, подвид схож с видом R. kilianii. Отличием считается наличие у последнего: 1) радиальных ребер на латеральных крыльях радиальных чешуек, 2) прямо срезанного дистального конца ствола радиальных чешуек, 3) слабо развитого аксиального утолщения пластинчатых чешуек.
Распространение. Для подвида R. erinaceoides undulata: пресные воды Чили, Шри-Ланки (Dürrschmidt, 1987a). Для всего вида: пресные воды европейской части России (Златогурский, 2014; Leonov, 2010; Mikrjukov, 1993), Украины (Gaponova, 2008), Эстонии (Mikrjukov, 1993a), Монголии (Kosolapova, Mylnikov, 2015), Канады (Nicholls, 1983), Чили (Dürrschmidt, 1985), Австралии (Croome, 1986); солоноватые воды европейской части России, 2‰ (Plotnikov, Gerasimova, 2017), ~ 20‰ (Mikrjukov, 2001).
Raineriophrys fortesca (Nicholls 1983) Mikrjukov 2001 [bas.: Acanthocystis fortesca Nicholls 1983; syn.: Acanthocystis pantopodeoides Nicholls 1983; Acanthocystis cuneiformis Dürrschmidt 1985] (рис. 3E–3H)
Материал. Четыре клетки (ТЭМ) из морских бентосных проб в окрестностях г. Нячанг 31.01.2018.
Морфология. Радиальные чешуйки состоят из полого цилиндрического ствола, базального и латеральных крыльев. Ствол 8.52–14.12 мкм в длину, 0.28–0.32 мкм в диаметре, в базальной части изогнут дорсально, дистальный конец несет 4–6 зубцов. Латеральные крылья тянутся вдоль ствола на 2.41–3.29 мкм, дистально сужаясь, в базальной части соединяются с перпендикулярно ориентированным базальным крылом и формируют ковшеобразное выпячивание. Ширина расширенной части крыльев 2.08–2.41 мкм. Базальное крыло несет радиально отходящие от ствола ребра. Яйцевидные пластинчатые чешуйки 3.33–4.72 × 2.06–2.74 мкм, с вогнутой центральной частью и аксиальным гребнем.
Замечания. Морфология исследованных чешуек соответствует предыдущим описаниям, однако ранее были указаны значительно более длинные радиальные чешуйки: 15–24 мкм (Mikrjukov, 1993; Nicholls, 1983). Морские образцы из Черного моря характеризовались значительно меньшими размерами: длина ствола радиальных чешуек 2.9–4.1 мкм, диаметр 0.11–0.13 мкм, длина пластинчатых чешуек 1.8–2.2 мкм (Prokina et al., 2019).
Распространение. Пресные воды европейской части России (Mikrjukov, 1993), Нидерланд, Швеции (Siemensma, Roijackers, 1988), Германии, Японии ([как A. cuneiformis] Dürrschmidt, 1985), Канады ([как A. fortesca и A. pantopodeoides] Nicholls, 1983), Чили (Dürrschmidt, 1985; Prokina, Mylnikov, 2019), Австралии ([как A. pantopodeoides] Croome, 1986). Белое море, 24‰ (Leonov, 2010), Черное море, 16–18‰ (Prokina et al., 2019).
Pseudoraphidocystis dentata Prokina, Radaykina, Mylnikov sp. n. (рис. 4)
Рис. 4.
Морфология чешуек Pseudoraphidocystis dentata sp. n. (A–D – ТЭМ, E–G – СЭМ): A – чешуйки одной клетки, B, D–G – радиальные чешуйки, C – пластинчатые чешуйки, зб – зубчики, нж – ножка. Остальные обозначения как на рис. 1. Масштаб (мкм): A – 5; B–G – 1.
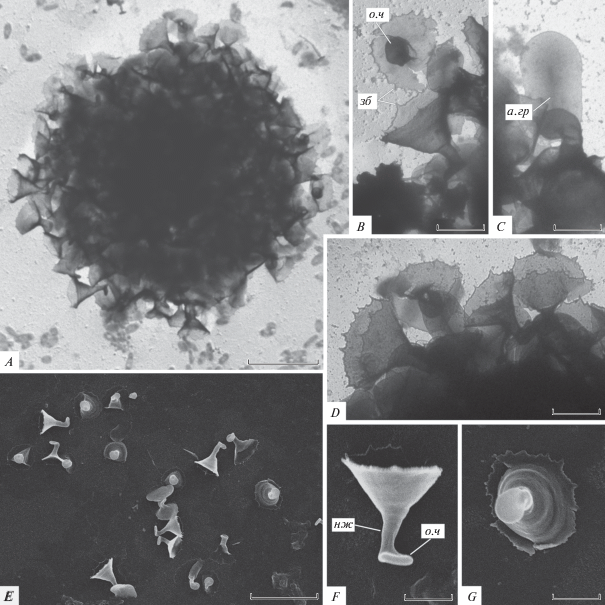
Материал. Исследованы пластинчатые и радиальные чешуйки 5 клеток (ТЭМ) и 1 скопление чешуек (СЭМ) из морских бентосных проб в окрестностях г. Нячанг, 31.01.2018. Типовой материал (препараты для СЭМ) хранятся в лаборатории Микробиологии ИБВВ РАН. Типовой рис. 4.
Диагноз. Детали строения протопласта неизвестны. Клетка покрыта воронковидными радиальными и овальными пластинчатыми чешуйками. Радиальные чешуйки 1.88–3.15 мкм в длину, с конически расширенной воронковидной дистальной частью (1.93–2.55 мкм в длину) и полой цилиндрической ножкой (1.05–1.85 мкм в длину). Дистальные края расширенной части чешуйки неровные, с заостренными зубчиками разной длины, диаметр расширенной части 2.17–3.03 мкм. Ножка диаметром 0.25–0.68 мкм в зауженной части, базальная часть расширяется в уплощенное основание 0.65–0.98 мкм в диаметре. Пластинчатые чешуйки 2.4–2.75 × 1.32–1.36 мкм, со слабо выраженным аксиальным гребнем.
Дифференциальный диагноз. Род Pseudoraphidocystis насчитывает 3 вида. P. glutinosa (Penard 1904) Mikrjukov 1997 имеет бесфактурные радиальные чешуйки с ровными дистальными краями воронковидной части чешуйки. P. flabellata (Dürrschmidt 1987) Mikrjukov 1997 и P. umbraculiformis (Dürrschmidt 1987) Mikrjukov 1997 отличаются наличием радиальных ребер, а также слабо развитой и короткой ножкой, а у подвида Ps. flabellata ssp. novae-zelandiae ножка полностью отсутствует. P. umbraculiformis также отличается наличием короткого стебелька. Небольшие воронковидные радиальные чешуйки характерны также для представителей рода Raphidocystis, однако отличаются наличием второго типа радиальных чешуек – длинных трубковидных или пипетковидных, а также морфологией пластинчатых чешуек, имеющих ретикулярную структуру и широкую полую маргинальную кайму.
Этимология. Вид назван по форме дистальных краев воронковидной части радиальной чешуйки, несущих многочисленные зубцы.
***Panacanthocystida Shishkin et Zlatogursky 2018
****Acanthocystidae Claus 1874 emend. Cavalier-Smith et von der Heyden 2007
Acanthocystis pectinata Penard 1889 emend. Nicholls 1983 [syn.: Acanthocystis radiosa Roskin 1929] (рис. 5A–5D)
Рис. 5.
Морфология живых клеток и чешуек центрохелид родов Acanthocystis и Raphidocystis (A, C–D, J–K – СЭМ; B, G H – ТЭМ; E–F, I – световая микроскопия): A–D – A. pectinata, E–H – A. turfacea, I–K – R. symmetrica; мб – мембрана, гр – гранулы, акс – аксоподии, п.р – продольное ребро, чш – чешуйки. Остальные обозначения как на рис. 1. Масштаб (мкм): E–F, I–J – 10; A–D, G–H, J–K – 2.
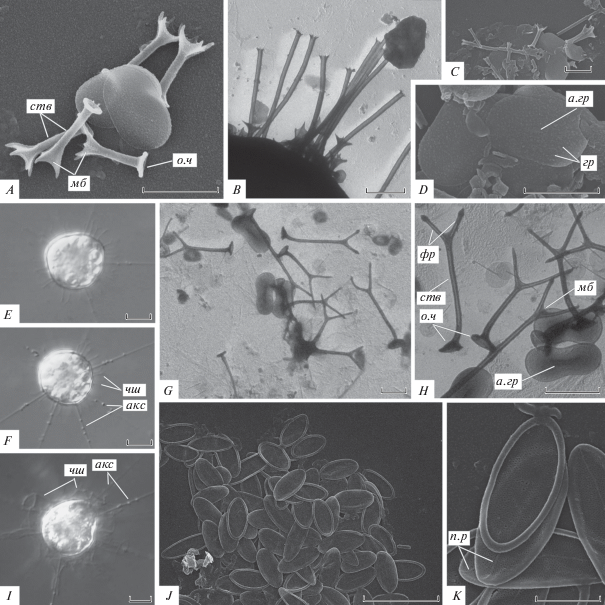
Материал. Два скопления чешуек (СЭМ) из р. Донгнай, детрит и песок под камнем, 02.02.2018; одна клетка (ТЭМ) из небольшого безымянного пруда с опавшей листвой и детритом в окрестностях р. Донгнай, 22.01.2018.
Морфология. Радиальные чешуйки двух типов, состоят из полого цилиндрического ствола 0.21–0.29 мкм в диаметре, воронковидного основания 0.67–0.85 мкм в диаметре и расширенного дистального конца, несущего фурки. Длинные радиальные чешуйки 6.28–7.97 мкм в длину, конец ствола незначительно расширяется (0.96–1.25 мкм в диаметре) и несет 6–9 коротких фурок. Короткие радиальные чешуйки 1.93–2.38 мкм в длину, конец ствола расширен в чашевидную структуру (0.35–0.49 мкм в диаметре) и несет 5–6 длинных фурок, соединенных мембраной. Овальные пластинчатые чешуйки 2.48–3.22 × × 1.47–1.66 мкм, с многочисленными мелкими гранулами на поверхности.
Замечания. Морфология исследованных чешуек в основном схожа с указанными ранее, однако некоторые авторы описывают 2–3 фурки на дистальных концах ствола длинных радиальных чешуек (Леонов, Плотников, 2009; Plotnikov, Ermolenko, 2015; Plotnikov, Gerasimova, 2017; Siemensma, Roijackers, 1988).
Распространение. Пресные воды европейской части России (Леонов, Мыльников, 2012; Леонов, Плотников, 2009; Leonov, 2010; Mikrjukov, 1993; Plotnikov, Ermolenko, 2015; Prokina et al., 2017), Эстонии (Mikrjukov, 1993a), Швеции, Нидерланд (Siemensma, Roijackers, 1988), Шри-Ланки ([как A. pectinata ssp. ceylanica] Dürrschmidt, 1987), Чили (Prokina, Mylnikov, 2019), солоноватые внутренние воды европейской части России, 2‰ (Plotnikov, Gerasimova, 2017).
Acanthocystis turfacea Carter 1863 (рис. 5E–5H)
Материал. Одно скопление чешуек (ТЭМ) и две живые клетки из морских бентосных проб в окрестностях г. Нячанг 31.01.2018.
Морфология. Протопласт живой клетки 21–24 мкм в диаметре. Радиальные чешуйки двух типов, состоят из полого цилиндрического ствола 0.15–0.22 мкм в диаметре, и воронковидного основания 0.79–1.11 мкм в диаметре, дистальный конец делится на 2 конически заостренные фурки. Ствол длинных радиальных чешуек 4.97–5.29 мкм в длину, фурки 0.98–1.22 мкм в длину. Ствол коротких радиальных чешуек 2.14–3.78 мкм в длину, фурки 1.05–1.90 мкм в длину. Овальные пластинчатые чешуйки 2.32–2.75 × 0.89–1.14 мкм, с немного зауженной центральной частью и аксиальным гребнем.
Замечания. В отличие от деталей строения чешуек, указанных в предыдущих описаниях данного вида, исследованные радиальные чешуйки имели характерные резко конически заостренные концы фурок, а также достаточно длинные фурки у длинных радиальных чешуек относительно длины ствола. В литературе встречаются значительные колебания размерностей чешуек и морфологических признаков. Чешуйки могут достигать огромных размеров: длина радиальных чешуек 35.6–65 мкм (Croome, 1986; Mikrjukov, 1993; Nicholls, 1983; Prokina et al., 2017), длина пластинчатых чешуек 4.9–12 мкм (Croome, 1986; Mikrjukov, 1993; Nicholls, 1983). Морфология радиальных чешуек также подвержена значительным изменениям: указываются как заостренные фурки (Croome, 1986; Croome et al., 1987; Dürrschmidt, 1985; Prokina et al., 2017), так и фурки с 2–3 маргинальными зубцами (Leonov, 2010; Mikrjukov, 1993; Nicholls, 1983; Plotnikov, Ermolenko, 2015; Plotnikov, Gerasimova, 2017; Prokina et al., 2017). Мембрана, соединяющая фурки, меняется от значительно выраженной (Леонов, Плотников, 2009; Leonov, 2010; Plotnikov, Ermolenko, 2015; Prokina et al., 2019), до практически полностью редуцированной (Croome, 1986; Dürrschmidt, 1985; Leonov, 2010; Prokina et al., 2019). Вполне вероятно, что это могут оказаться разные виды и для решения этого вопроса необходимы комплексные исследования моноклональных культур из различных местообитаний с применением методов молекулярной филогении.
Распространение. Пресные воды европейской части России (Леонов, Плотников, 2009; Leonov, 2010; Mikrjukov, 1993; Plotnikov, Ermolenko, 2015; Prokina et al., 2017), Швейцарии (Златогурский, 2014), Японии (Takamura et al., 2000), Канады (Nicholls, 1983), Австралии (Croome, 1986), Чили (Dürrschmidt, 1985; Prokina, Mylnikov, 2019); Балтийское море, 5–6‰ (Vørs, 1992), Черное море, 16–18‰ (Prokina et al., 2019), Белое море, 24‰ (Leonov, 2010), морские воды Антарктики, ~35‰ (Croome et al., 1987), соленые континентальные воды европейской части России, 30–42.2‰ (Plotnikov, Gerasimova, 2017).
****Raphidocystidae Zlatogursky in Zlatogursky et al. 2018
Raphidocystis symmetrica (Penard 1904) Zlatogursky in Zlatogursky et al. 2018 [bas.: Raphidiophrys symmetrica Penard 1904; syn.: Polyplacocystis symmetrica (Penard 1904) Mikrjukov 1996] (рис. 5I–5K)
Материал. Три клетки (СЭМ) из р. Донгнай, детрит и песок под камнем, 02.02.2018.
Описание. Протопласт живой клетки 29 мкм в диаметре. Овальные пластинчатые чешуйки 4.64–7.9 × 1.8–2.68 мкм, с полой маргинальной каймой 0.19–0.31 мкм в диаметре. Отношение длины к ширине 1.86–4.39. Внутренняя часть чешуйки имеет ретикулярную структуру, вдоль продольной оси чешуйки располагается ребро.
Замечания. Размеры и морфология исследованных чешуек соответствуют предыдущим описаниям. Чешуйки этого вида схожи с чешуйками R. ambigua (Penard 1904) Zlatogursky in Zlatogursky et al. 2018 и R. tubifera Penard 1904. R. ambigua отличается наличием также зауженных и длинных пластинчатых чешуек; R. tubifera отличается наличием радиальных воронковидных чешуек.
Распространение. Пресные воды европейской части России (Leonov, 2010; Prokina et al., 2017), Швеции, Нидерландов (Siemensma, Roijackers, 1988a), Канады (Nicholls, Dürrschmidt, 1985), Гренландии (Ikävalko et al., 1996).
ОБСУЖДЕНИЕ
Исследованы покровные элементы (неорганические чешуйки) центрохелидных солнечников из разнотипных местообитаний Вьетнама. Всего выявлено 10 видов из 6 родов и 4-х семейств (Choanocystidae, Pterocystidae, Acanthocystidae, Raphidocystidae). Все обнаруженные виды – вероятно, новые для Вьетнама, 2 вида – новые для науки. Родовой коэффициент (отношение количества видов к количеству родов) низкий (1.67), что может характеризовать выявленную фауну центрохелид как недостаточно изученную. Наибольшее количество видов обнаружено в роде Pterocystis (3), Raineriophrys и Acanthocystis (по 2). Остальные роды – по одному виду. Наиболее богаты видами семейства Pterocystidae (6) и Acanthocystidae (2). Остальные семейства насчитывают по 1 виду.
Наиболее часто встречаемые виды – Pterocystis foliacea и Acanthocystis pectinata – обнаружены в двух биотопах. Остальные виды были выявлены только в одном биотопе. Распределение видов по исследованным биотопам неравномерное. Наибольшим видовым богатством характеризуется р. Донгнай (5 видов, 50% от выявленных). Меньше видов в безымянном пруду близ р. Донгнай и в морских местообитаниях в окрестностях г. Нячанг (по 3 вида, по 30%). Минимальное количество видов найдено в озере Бау Сау (1 вид, 10%). Среднее значение видового разнообразия водных объектов 3.0.
В пресных водах выявлено наибольшее количество видов – 7, в то время как в морских – всего 3. При этом не выявлено видов, общих для морской и пресноводной фаун. Большинство видов, найденных в исследованных биотопах Вьетнама, известны из различных континентов и климатических зон. Наиболее редкими (кроме новых видов, описываемых впервые) можно считать Pterocystis tropica (известен из Европы, Ю. Америки и Азии) и Raphidocystis symmetrica (известен из Европы, С. Америки и Гренландии). Больше половины (т.е. 6 видов, Acanthocystis pectinata (2‰), A. turfacea (5–42.2‰), Choanocystis perpusilla (2–35‰), Pterocystis foliacea (16–18‰), Raineriophrys fortesca (16–24‰), R. erinaceoides (2–20‰)) из обнаруженных здесь видов – были ранее выявлены как в пресных, так и в соленых континентальных и в морских местообитаниях. При этом вид Acanthocystis pectinata ранее был выявлен в наименее соленых местообитаниях, а Choanocystis perpusilla и A. turfacea имеют широкую границу вариабельности солености биотопов. Два вида (Pterocystis tropica и Raphidocystis symmetrica), известные только из пресных биотопов, подтвердили свою приуроченность к пресным водам в нашем исследовании.
Морфология чешуек исследованных видов в целом соответствует известным описаниям. Четыре вида (Choanocystis perpusilla, Raineriophrys fortesca, Pterocystis foliacea и Raphidocystis symmetrica) не имели значимых отличий в морфологии и размерах чешуек. Другие четыре вида из известных ранее имели некоторые особенности. Для вида Acanthocystis pectinata часто указывали наличие 2–3 зубцов на верхушке ствола радиальных чешуек (Леонов, Плотников, 2009; Plotnikov, Ermolenko, 2015; Siemensma, Roijackers, 1988; Plotnikov, Gerasimova, 2017), в то время как мы обнаружили 5–6 зубцов. В отличие от предыдущих описаний, радиальные чешуйки Acanthocystis turfacea имели характерные конически заостренные концы фурок, а также большую длину фурок длинных радиальных чешуек относительно длины ствола. В литературе описаны значительные колебания размерностей чешуек, а также морфологических признаков (как заостренные фурки, так и с 2–3 маргинальными зубцами; с хорошо выраженной мембраной между фурок – и с полностью редуцированной). У вида Pterocystis tropica известно 2 подвида, при этом исследованные пластинчатые чешуйки больше соответствуют таким чешуйкам подвида P. tropica paucistriata (без радиальных прорезей), а радиальные чешуйки больше схожи с таковыми подвида P. tropica tropica (сильно выраженные плечи, большее количество радиальных ребер на латеральных и базальном крыльях) (Dürrschmidt, 1987a). Исследованные пластинчатые чешуйки вида Raineriophrys erinaceoides ssp. undulata не имееют радиальных прорезей в отличие от того, что указано в первоописании для этого подвида, а также в первоописании указаны значительно меньшие размеры чешуек (Dürrschmidt, 1987a).
Два новых вида центрохелидных солнечников имеют уникальную морфологию чешуек, отличимую от чешуек всех известных ранее видов. Pterocystis vietnamica sp. n. характеризуется схожими с P. foliacea листовидными радиальными чешуйками, однако имеет характерные только для него признаки – поперечные прорези на латеральных крыльях и крупное округлое субапикальное выпячивание. Pseudoraphidocystis dentata sp. n. характеризуется наличием маргинальных зубцов на краях воронковидных радиальных чешуек, что также является уникальной особенностью, не известной ранее для этого рода. Для обоих видов было выявлено несколько клеток, что позволяет нам с уверенностью утверждать об отсутствии мутации уже известных видов, а также позволяет исследовать вариабельность морфологических и морфометрических признаков новых видов. Поскольку выделить в клональные культуры данные организмы не удалось, детали строения живой клетки неизвестны, а также нет молекулярных данных. В дальнейшем мы планируем провести эти исследования и совместно с морфологическим анализом определить правильность родовой принадлежности видов и выявить филогенетические связи с другими известными видами центрохелид.
Полученные морфологические данные неорганических чешуек всех исследованных видов дополняют наши знания о внутривидовой изменчивости центрохелидных солнечников. Выявленные новые виды центрохелидных солнечников подтверждают предположение о существовании огромного, еще не известного многообразия этих протистов. Выявленные виды центрохелид, известные ранее из различных местообитаний на многих континентах, в некоторой степени заполняют пробелы в биогеографии солнечников и в дальнейшем помогут решить вопросы о космополитизме данных организмов, а также о вероятном единстве пресноводной и морской фауны.
Список литературы
Ермоленко Е.А., Плотников А.О., 2013. Экология центрохелидных солнечников в водоемах Южного Предуралья // Вестник Оренбургского Государственного университета. Т. 10. № 159. С. 119–121.
Златогурский В.В., 2014. Разнообразие и эволюция покровных структур у центрохелидных солнечников (Protista: Centrohelida). Дис… канд. биол. наук. СПбГУ, Санкт-Петербург.
Леонов М.М., 2010. Новые виды центрохелидных солнечников рода Acanthocystis (Centroheliozoa) // Зоологический журнал. Т. 89. № 5. С. 507–513.
Леонов М.М., Мыльников А.П., 2012. Центрохелидные солнечники из Южной Карелии // Зоологический журнал. Т. 91. № 5. С. 515–515.
Леонов М.М., Плотников А.О., 2009. Центрохелидные солнечники (Centroheliozoa) Центрального Черноземья и Южного Урала: видовой состав, морфология, распространение // Зоологический журнал. Т. 88. № 6. С. 643–653.
Микрюков К.А., 2002. Центрохелидные солнечники (Centroheliozoa). М.: Товарищество научных изданий КМК. 136 с.
Шатилович А.В., Мыльников А.П., Ступин Д.В., 2010. Фауна и морфология гетеротрофных жгутиконосцев и солнечников позднеплейстоценовых ископаемых нор сусликов (Колымская низменность) // Зоологический журнал. Т. 89. № 4. С. 387–397.
Adl S.M., Bass D., Lane C.E., Lukeš J., Schoch C.L., Smirnov A., et al., 2019. Revisions to the classification, nomenclature, and diversity of Eukaryotes // Journal of Eukaryotic Microbiology (in press).
Cavalier-Smith T., von der Heyden S., 2007. Molecular phylogeny, scale evolution and taxonomy of centrohelid heliozoan // Molecular Phylogenetics and Evolution. V. 44. № 3. P. 1186–1203.
Croome R., 1986. Observations of the heliozoan genera Acanthocystis and Raphidocystis from Australia // Archiv für Protistenkunde. V. 131. № 3–4. P. 189–199.
Croome R., 1987. Observations of the genera Acanthocystis, Raphidiophrys, Clathrulina and Pompholyxophrys (Protozoa, Sarcodina) from Australian freshwaters // Archiv für Protistenkunde. V. 133. № 3–4. P. 237–243.
Croome R.L., van den Hoff J., Burton H.R., 1987. Observations of the heliozoean genera Pinaciophora and Acanthocystis (Heiliozoea, Sarcodina, Protozoa) from Ellis Fjord, Antarctica // Polar Biology. V. 8. № 1. P. 23–28.
Dürrschmidt M., 1985. Electron microscopic observations on scales of species of the genus Acanthocystis (Centrohelidia, Heliozoa) from Chile, I // Archiv für Protistenkunde. V. 129. № 1–4. P. 55–87.
Dürrschmidt M., 1987. An electron microscopical study of freshwater Heliozoa (genus Acanthocystis, Centrohelidia) from Chile, New Zealand, Malaysia and Sri Lanka. II // Archiv für Protistenkunde. V. 133. № 1–2. P. 21–48.
Dürrschmidt M., 1987a. An electron microscopical study on freshwater Heliozoa (genus Acanthocystis, Centrohelidia) from Chile, New Zealand, Malaysia and Sri Lanka. III // Archiv für Protistenkunde. V. 133. № 1–2. P. 49–80.
Gaponova L., 2008. The Centrohelid Heliozoans (Protista, Centrohelida) of algal mats from the water body in Ukrainian Polissya // Vestnik zoologii. V. 42. № 2. P. e38–e41.
Ikävalko J., Thomsen H.A., Carstens M., 1996. A preliminary study of NE Greenland shallow meltwater ponds with particular emphasis on loricate and scale-covered forms (Choanoflagellida, Chrysophyceae sensu lato, Synurophyceae, Heliozoea), including the descriptions of Epipyxis thamnoides sp. nov. and Pseudokephyrion poculiforme sp. nov. (Chrysophyceae) // Archiv für Protistenkunde. V. 147. № 1. P. 29–42.
Kosolapova N.G., Mylnikov A.P., 2015. First records of Centrohelid Heliozoans (Centrohelida) in Mongolia // Inland Water Biology. V. 8. № 3. P. 232–235.
Leonov M.M., 2010. Heliozoans (Heliozoa, Sarcodina, Protista) of fresh and marine waters of the European part of Russia: species composition, morphology, and distribution // Inland Water Biology. V. 3. № 4. P. 344–355.
Mikrjukov K.A., 1993. Observations on Centroheliozoa of the Volga basin (Protozoa: Sarcondina) // Zoosystematica Rossica. V. 2. P. 201–209.
Mikrjukov K.A., 1993a. On the Centrohelid and Rotosphaerid Heliozoa from the environs of the Võrtsjärv limnological station in Estonia // Proceedings of the Estonian Academy of ciences. V. 42. №. 2. P. 154–160.
Mikrjukov K.A., 1999. Interesting findings of heliozoan (Protista) in the Black Sea and Crimea: data on the similarity of sea and freshwater fauna of these organisms // Russian Journal of Zoology. V. 3. № 2. P. 240–249.
Mikrjukov K.A., 2001. Heliozoa as a component of marine microbenthos: a study of heliozoan of the White Sea // Ophelia. V. 54. № 1. P. 51–73.
Moestrup Ø., Thomsen H.A., 1980. Preparations of shadow cast whole mounts // Handbook of Phycological Methods. Cambridge: Cambridge Univ. Press. P. 385–390.
Nicholls K.H., 1983. Little-known and new Heliozoeans: the centrohelid genus Acanthocystis, including descriptions of nine new species // Canadian Journal of Zoology. V. 61. № 6. P. 1369–1386.
Nicholls K.H., Dürrschmidt M., 1985. Scale structure and taxonomy of some species of Raphidocystis, Raphidiophrys, and Pompholyxophrys (Heliozoea) including descriptions of six new taxa // Canadian Journal of Zoology. V. 63. № 8. P. 1944–1961.
Plotnikov A.O., Ermolenko E.A., 2015. Centrohelid heliozoa (Chromista, Hacrobia) of Southern Cis-Ural Region // Biology Bulletin. V. 42. № 8. P. 683–695.
Plotnikov A.O., Gerasimova E.A., 2017. Heliozoa (Centrohelea, Haptista, Hacrobia) of saline and brackish water bodies and watercourses of Russia // Inland Water Biology. V. 10. № 2. P. 121–129.
Prokina K.I., Mylnikov A.P., 2019. Centrohelid heliozoans from freshwater habitats of South Patagonia and Tierra del Fuego, Chile // Inland Water Biology. V. 12. № 1. P. 10–20.
Prokina K.I., Zagumyonnyi D.G., Mylnikov A.P., 2019. Marine centrohelid heliozoans (Centroplasthelida Febvre-Chevalier et Febvre, 1984) from bays of Sevastopol (the Black Sea shore) // Russian Journal of Marine Biology (in press).
Prokina K.I., Zagumyonnyi D.G., Philippov D.A., 2017. Centrohelids in the mires of Northern Russia // Protistology. V. 11. № 1. P. 3–19.
Siemensma F.J., Roijackers R.M.M., 1988. A study of new and little-known acanthocystid heliozoeans, and a proposed division of the genus Acanthocystis (Actinopoda, Heliozoea) // Archiv für Protistenkunde. V. 135. P. 1–4. P. 197–212.
Siemensma F.J., Roijackers R.M.M., 1988a. The genus Raphidiophrys (Actinopoda, Heliozoea): scale morphology and species distinctions // Archiv für Protistenkunde. V. 136. № 3. P. 237–248.
Takamura N., Shen Yu., Xie P., 2000. Species richness of protozoa in Japanese lakes // Limnology. V. 1. P. 91–106.
Tikhonenkov D.V., Mylnikov A.P., 2011. Choanocystis antarctica sp. n., a new heliozoan (Centrohelida) species from the littoral zone of King George Island, South Shetland Islands, Antarctica // Biology Bulletin. V. 38. № 7. P. 663–666.
Vørs N., 1992. Heterotrophic amoebae, flagellates and heliozoan from the Tvärminne area, gulf of Finland, in 1988 – 1990 // Ophelia. V. 36. № 1. P. 1–109.
Wujek D.E., Ogundipe O.T., 2002. Heliozoa from Nigeria // Tropical Freshwater Biology. V. 11. № 1. P. 1–9.
Zlatogursky V.V., 2015. Two new species of Centrohelid Heliozoans: Acanthocystis costata sp. nov. and Choanocystis symna sp. nov. // Acta Protozoologica. V. 53. № 4. P. 313–324.
Zlatogursky V.V., Gerasimova E.A., Plotnikov A.O., 2017. A new species of Centrohelid Heliozoan Acanthocystis amura n. sp. isolated from two remote locations in Russia // Journal of Eukaryotic Microbiology. V. 64. № 4. P. 434–439.
Дополнительные материалы отсутствуют.
Инструменты
Зоологический журнал