

Зоологический журнал, 2019, T. 98, № 9, стр. 1048-1062
Основные этапы формирования фауны жужелиц (Coleoptera, Carabidae) Сихотэ-Алиня на примере эндемиков. 2. Анализ ареалов
Ю. Н. Сундуков *
Федеральный научный центр биоразнообразия наземной биоты
Восточной Азии ДВО РАН
690022 Владивосток, Россия
* E-mail: yun-sundukov@mail.ru
Поступила в редакцию 08.06.2018
После доработки 20.03.2019
Принята к публикации 20.03.2019
Аннотация
Проанализированы ареалы эндемичных жужелиц Сихотэ-Алиня в объеме родственных групп. Выделены “центры эндемизма” и “главные рубежи” на территории Сихотэ-Алиня. Дан анализ особенностей высотно-биотопической приуроченности эндемиков. Выделены главные этапы формирования современной эндемичной фауны жужелиц Сихотэ-Алиня. Выяснено, что отсутствие покровного оледенения в плейстоцене позволило фауне Сихотэ-Алиня в течение длительного периода формироваться без эпох полного уничтожения. Благодаря сохранившейся в плейстоцене лесной растительности на юге и востоке Сихотэ-Алиня, этот район стал важнейшим рефугиумом для третичной фауны Северной Азии.
Это второе, заключительное сообщение, посвященное анализу эндемичной фауны жужелиц Сихотэ-Алиня. В Сообщении 1 были даны сведения по таксономическому составу, распространению, экологии и филогенетическим связям сихотэ-алинских эндемиков (Сундуков, 2019).
В начале Сообщения 2 приведены литературные данные по ископаемым таксонам жужелиц с мелового периода до настоящего времени, населяющим в настоящее время Сихотэ-Алинь. Основная часть сообщения посвящена непосредственно анализу эндемичных таксонов. Здесь рассмотрены их ареалы на уровне родственных групп, выделены “центры эндемизма” и “главные рубежи” на территории Сихотэ-Алиня, дан анализ особенностей высотно-биотопической приуроченности эндемиков и выделены основные этапы формирования современной эндемичной фауны жужелиц Сихотэ-Алиня.
ПАЛЕОНТОЛОГИЧЕСКИЕ ДАННЫЕ
Палеонтологических данных по жужелицам Сихотэ-Алиня практически нет, как и по ископаемым Carabidae Восточной Азии в целом. Поэтому в обзоре приведены данные о наиболее ранних находках таксонов, обитающих в настоящее время на Сихотэ-Алине. Эти данные использовалась нами при оценке возможного времени возникновения эндемичного таксона – рода, подрода, вида или подвида.
Роды жужелиц, известные в настоящее время с Дальнего Востока России, появляются в палеоцене. Например, из позднего палеоцена (58.7–55.8 млн л. н.) Франции описаны ископаемые виды Bembidion Latreille 1802, Agonum Bonelli 1810 и Harpalus Latreille 1802 (Piton, 1940), а из позднего палеоцена северной Аргентины – ископаемый вид Lebia Latreille 1802 (Cockerel, 1936).
В эоценовой фауне современные роды Carabidae встречаются значительно чаще, хотя и представлены только ископаемыми видами. Из раннего (55.8–48.6 млн л. н.) и среднего эоцена (48.6–40.4 млн л. н.) известны Nebria Latreille 1802, ?Amara Bonelli 1810, Carabus Linnaeus 1758 и Poecilus Bonelli 1810 (Manchester, 1992; Omboni, 1886; Pongrácz, 1935; Scudder, 1879), из позднего эоцена (37.2–33.9 млн л. н.) – Nebria, Calosoma Weber 1801, Loricera Latreille 1802, Bembidion, Trechus Clairville 1806, Tachys Dejean 1821, Elaphropus Motschulsky 1839, Pterostichus Bonelli 1810, Platynus Bonelli 1810, Amara, Stenolophus Dejean 1821, Harpalus, Diplocheila Brullé 1837, Dromius Bonelli 1810 и Brachinus Weber 1801 (Arillo et al., 1997; Cai et al., 2017; Erwin, 1971; Germar, 1813; Giebel, 1856; Schmidt, Faille, 2015; Schmidt et al., 2016; Scudder, 1890; Wickham, 1910, 1913).
Олигоценовая фауна жужелиц также представлена только ископаемыми видами. Помимо перечисленных выше родов, из раннего олигоцена (33.9–28.4 млн л. н.) описан ископаемый Anchomenus Bonelli 1810 (Théobald, 1937), а из позднего олигоцена (28.4–23.0 млн л. н.) – представители Chlaenius Bonelli 1810 и Panagaeus Latreille 1802 (Piton, Rudel, 1936; Piton, Théobald, 1936).
В составе миоценовой фауны, помимо ископаемых таксонов, появляются современные и близкие к ним подроды и виды. Например, из бурдигальского яруса (20.4–16.0 млн л. н.) восточного макросклона Сихотэ-Алиня описан Patrobus kudiensis Cockerel 1925 (Cockerel, 1925). Из среднего миоцена (16.0–11.6 млн л. н.) провинции Шаньдун на востоке Китая, помимо ископаемых видов, указан Calosoma cf. maderae (Fabricius 1775) (Zhang, 1989), широко распространенный в настоящее на юго-западе Палеарктики, но не встречающийся в Восточной Азии. Из позднего миоцена (11.6–5.3 млн л. н.) северо-запада Чукотки представлены современные или близкие к ним таксоны (подроды Peryphus Dejean 1821 и Plataphodes Ganglbauer 1891 рода Bembidion, подроды Argutor Dejean 1821, Cryobius Chaudoir 1838 и Eosteropus Tschitschérine 1902 рода Pterostichus, подрод Europhilus Chaudoir 1859 рода Agonum, виды Trachypachus cf. inermis Motschulsky 1850, Notiophilus aquaticus (Linnaeus 1758), Blethisa aurata Fischer von Waldheim 1828, Elaphrus riparius (Linnaeus 1758), Diacheila arctica (Gyllenhal 1810), Pterostichus costatus (Ménétriès 1851), Agonum quinquepunctatum Motschulsky 1844 и Dicheirotrichus cf. cognatus (Gyllenhal 1827)) (Kiselev, Nazarov, 2009). Не менее показательна ископаемая фауна мессинского яруса (7.2–5.3 млн л. н.) Аляски, из которой указаны современные подроды Dyschiriodes Jeannel 1941 (род Dyschirius Bonelli 1810), Trepanedoris Netolitzky 1918 (род Bembidion), Pseudomaseus Chaudoir 1838 (род Pterostichus), виды Carabus truncaticollis Eschscholtz 1823, Patrobus cf. septentrionis Dejean 1828, Pterostichus cf. vermiculosus (Ménétriès 1851) и Pt. cf. corvinus (Dejean 1828) (Hopkins et al., 1971). Большинство этих таксонов и в настоящее время широко распространены на севере Голарктики.
Плиоценовая фауна жужелиц плохо изучена, но она, вероятно, еще более схожа с современной (Пономаренко, Жерихин, 1980). Из раннего плиоцена (5.3–3.6 млн л. н.) северо-востока Якутии, помимо ископаемых видов, указываются Trachypachus cf. inermis и Diacheila cf. arctica (Kiselev, Nazarov, 2009). Из позднего плиоцена (3.6–2.6 млн л. н.) Германии – Calosoma inquisitor (Linnaeus 1758), C. maderae (Fabricius 1775) и C. sycophantha (Linnaeus 1758) (Gersdorf, 1969), Аляски – Notiophilus cf. aeneus (Herbst 1806) и Asaphidion cf. yukonense Wickham 1919 (Matthews et al., 2003), Японии – Elaphrus sp. и Poecilus cf. planicollis Motschulsky 1861 (Hayashi et al., 2003).
Фауна жужелиц плейстоцена практически не отличается от современной. Например, из калабрийского яруса (1.8–0.8 млн л. н.) центрального Хонсю (Япония) приводятся Calosoma ?inquisitor (Linnaeus 1758), Carabus maackii Morawitz 1862, C. maeander Fischer von Waldheim 1820, Poecilus cf. planicollis и Chlaenius sulcicollis (Paykull 1798) (Hayashi, 1996, 1998). Из среднего плейстоцена Японии – Carabus blaptoides (Kollar 1836), C. ?granulatus Linnaeus 1758, C. japonicus Motschulsky 1857, C. maackii, Elaphrus japonicus Uéno 1954, Synuchus sp. (Hayashi et al., 2005, 2008, 2009). Интересны позднеплейстоценовые (126–40 тыс. л. н.) находки на северо-западе США (штат Вашингтон) Trechiama sp. (Cong, Ashworth, 1996) и Microlestes minutulus (Goeze 1777) (Ashworth, Nelson, 2014), а в штате Висконсин, на рубеже плейстоцена-голоцена (12.0–11.7 тыс. л. н.), – Trechus apicalis Motschulsky 1845 (Morgan, Morgan, 1979).
АНАЛИЗ СИХОТЭ-АЛИНСКИХ ЭНДЕМИКОВ
Группы ареалов
Изучение ареалов сихотэ-алинских эндемиков, их биотопической приуроченности и родственных связей на уровне минимальных таксономических групп (Сундуков, 2019) позволило выделить на Сихотэ-Алине 6 групп эндемиков, связанных общими закономерностями распространения и эволюционной историей:
(1) “Восточноазиатские эндемики” (15 таксонов) имеют дизъюнктивные ареалы, ограниченные в своем северном распространении горами Приамурья и Южной Сибири, и родственные связи с современными таксонами юга и востока Восточноазиатской зоогеографической области. По характеру распространения и происхождению выделяется две подгруппы:
(а) “Гумидные восточноазиатские эндемики” – представители рода Xestagonum Habu 1978 (2 вида), подродов Eonebria Semenov et Znojko 1928 (2 вида), Epaphiama Jeannel 1962 (1 вид), Leptepaphiama Jeannel 1962 (2 вида) и видовых групп “densicornis” (3 вида с 5 подвидами) и “sundukovi” (1 вид) рода Trechus (Сундуков, 2019: рис. 3A–3F).
Оптимальными условиями обитания на Сихотэ-Алине для этих таксонов является верхняя зона пояса гумидных темнохвойных лесов (“умеренные дождевые леса”) на высотах 800–1600 м. Большинство из них адаптированы к гипогейному образу жизни, что выражается в редукции плеч и задних крыльев, уменьшении глаз, удлинении конечностей и хет, уплощении тела и депигментации покровов. В современное время все указанные группы – эндемики или субэндемики Восточной Азии. Их наиболее архаичные или мало специализированные формы сохранились на юге Восточной Азии, а островные и материковые формы на севере области находятся в различной степени дивергенции.
(б) “Восточноазиатские реликты” – 1 эндемичный вид – Masuzoa ussuriensis Lafer 1989 (Сундуков, 2019: рис. 1C).
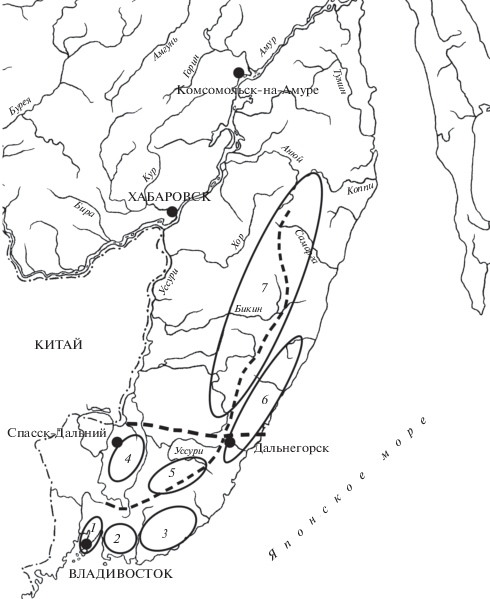
Рис. 1.
Важнейшие “рубежи” и “центры эндемизма” на Сихотэ-Алине. Центры эндемизма: 1 – п-ов Муравьева-Амурского, 2 – Ливадийский хребет, 3 – Партизанский хребет, 4 – Синий хребет, 5 – группа горных вершин в истоках р. Уссури, 6 – восточный макросклон Сихотэ-Алиня к северу от линии Спасск-Дальний–Дальнегорск, 7 – высокогорья северного Сихотэ-Алиня. Рубежи обозначены пунктирной линией.
Дизъюнктивный ареал рода схож с ареалами следующей группы, но относится нами к группе “Восточноазиатские эндемики” на основании фрагмента ареала Masuzoa на о-ве Хоккайдо. По ряду морфологических, в том числе апоморфных, признаков, Masuzoa изолирован от других известных родов Trechini и может быть рассмотрен как реликт одной из архаичных форм филетической линии Epaphiopsis (Сундуков, 2019).
Схожая дизъюнкция ареала и, вероятно, эволюционная история в палеогене наблюдается у палеарктических представителей семейства Grylloblattidae (Wipfler et al., 2014).
(2) “Эндемики гор Южной Сибири и Приамурья” (4 таксона) – виды подрода Leistus Frölich 1799 (2 эндемичных вида), а так же видовых групп “montanus” (1 вид) и “almonius” (1 вид) рода Trechus (Сундуков, 2019: рис. 1A, 1D, 2A).
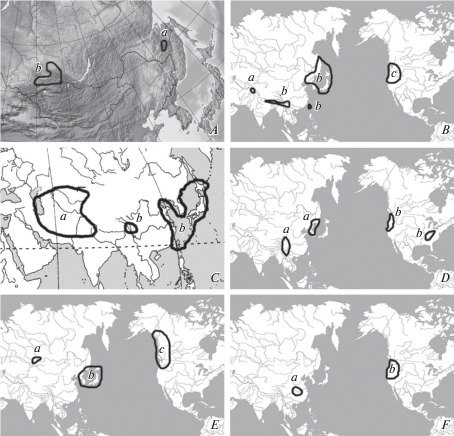
Рис. 2.
Ареалы таксономических групп насекомых (Insecta): A – видовая группа “Trechus mordkovitschi” (Carabidae) (a – Trechus badzhalensis Plutenko 2004; b – T. mordkovitschi Shilenkov 1982); B – монофилетические роды трибы Broscini, Carabidae (a – Kashmirobroscus Schmidt, Wrase et Sciaky 2013; b – Eobroscus Kryzhanovskij 1951; c – Zacotus LeConte 1869); C – род Craspedonotus Schaum 1863, Carabidae (a – C. margellanicus Kraatz 1884 и C. himalayanus Semenov 1910; b – C. tibialis Schaum 1863); D – род Cryptocercus Scudder 1862, Cryptocercidae (a – восточноазиатские виды, b – североамериканские виды); E – роды семейства Grylloblattidae (a – Grylloblattella Storozhenko 1988; b – Grylloblattina Bey-Bienko 1951 и Galloisiana Caudell 1924; c – Grylloblatta Walker 1914); F – роды трибы Metriini, Carabidae (a – Sinometrius Wrase et Schmidt 2006; b – Metrius Eschscholtz 1829).
Для этих таксонов на Сихотэ-Алине характерно обитание у верхней границы темнохвойных лесов (Trechus) или в высокогорьях (Leistus).
Ареал группы сходен с распространением рода Masuzoa. Наиболее заметное отличие от современного распространения группы “Восточноазиатские эндемики” заключается в почти полном отсутствии на восточных окраинах Восточной Азии и, наоборот, значительном таксономическом разнообразии не только в горах западного Китая, но и в целом на западе Палеарктики.
Схожую и, вероятно, общую по происхождению дизъюнкцию ареала проявляет видовая группа “mordkovitschi” рода Trechus (рис. 2A), в которую мы включаем 2 вида: описанный из верхнего пояса Баджальского хребта T. badzhalensis Plutenko 2004 и обитающий в Саянских горах T. mordkovitschi Shilenkov 1982 (Белоусов, Кабак, 1994; Плутенко, 2004; Shilenkov, 1982). Оба вида населяют горные лиственничники и тундры, встречаясь во влажной моховой подстилке, и имеют очень схожие строение эдеагуса, внутреннее вооружение эндофаллуса и строение переднеспинки, усиков и надкрылий.
(3) “Транспалеарктические неморальные эндемики” (13 таксонов) имеют трансевразиатские азиадизъюнктивные или суббореальные ареалы по классификации Городкова (1984). Сюда включены эндемики Сихотэ-Алиня из подродов Eosteropus (1 вид и 1 подвид), Phonias Gozis 1886 (1 вид) и Feroperis Lafer 1979 (7 видов с 4 подвидами) рода Pterostichus и один вид подрода Cymindis Latreille 1805 (Сундуков, 2019: рис. 4B; 5A–5C).
Рис. 3.
Положение палео- (серая заливка) и современной суши (оконтурено) в Восточной Азии в миоцене (А – 15.5 млн л., B – 8.0–5.5 млн и C – 5.4–4.3 млн лет назад; по: Sun, Wang, 2005), минделе (D) и риссе (E; по: Minato, Ijiri, 1976).
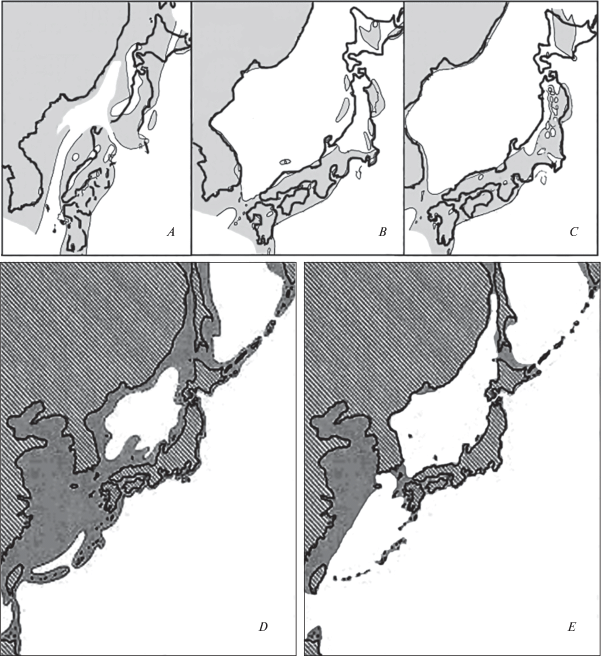
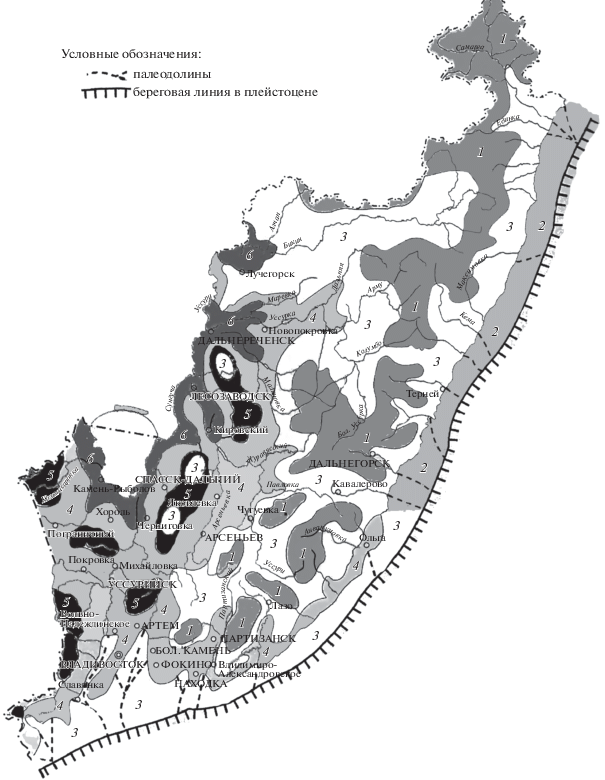
Рис. 4.
Виды ландшафтов Южного Сихотэ-Алиня в климатический минимум плейстоцена (около 18 тыс. л. н.) (по: Короткий и др., 1996): 1 – горные тундры, 2 – тундры прибрежных равнин, 3 – лесотундры и редколесья, 4 – березово-лиственничные леса, 5 – еловые и березово-лиственничные леса, 6 – редкостойные леса на заболоченных равнинах.
Все таксоны этой группы связаны с лесами, обитая на Сихотэ-Алине под пологом широколиственных и смешанных лесов (Cymindis), горных темнохвойных лесов (Pt. datshenkoae Sundukov 2013) или во всех типах леса (Eosteropus, Feroperis). Дизъюнкция их ареалов может быть объяснена плиоцен-плейстоценовым разрывом в распространении неморальных лесов в Сибири, поэтому мы считаем, что происхождение этих таксонов связано именно с неморальными лесами.
Примеры подобных разрывов ареалов нередки. Среди насекомых это рода Rhysodes Germar 1822 (Rhysodidae), Lucanus Scopoli 1763 (Lucanidae) и Rosalia Audinet-Serville 1833 (Cerambycidae), жужелицы подрода Calosoma Weber 1801 и рода Blemus Dejean 1821. Среди других групп животных это голубая сорока, обыкновенная ворона, черноголовая гаичка, благородный олень, рыба обыкновенный вьюн и другие.
(4) “Бореальные эндемики” (8 таксонов) – основная часть ареала монофилетических групп, в которые входят эндемичные для Сихотэ-Алиня таксоны, расположена в бореальном поясе. По характеру их распространения и происхождению можно выделить две подгруппы:
(а) “Восточноазиатские бореальные эндемики” – эндемичные таксоны из подродов Cryobius (3 вида), Lenapterus O. Berlov 1996 (2 вида с 1 подвидом) и Petrophilus Chaudoir 1838 (1 вид) рода Pterostichus (Сундуков, 2019: рис. 2C; 4C–4F).
Становление этих групп связывается нами с формированием хвойных таежных лесов на северо-востоке Азии во второй половине миоцена– плиоцене, а дифференциация на видовом уровне – в плиоцен–плейстоценовое время (см. ниже раздел Основные этапы…).
(б) “Транспалеарктические бореальные эндемики” – 1 эндемичный вид, Cymindis laferi Sundukov 1999 (Сундуков, 2019: рис. 5D ).
Становление этого вида можно связать с одним из холодных периодов плейстоцена, во время которых на востоке и юге Сихотэ-Алиня происходили сокращения лесной растительности до небольших рефугиумов и появлялась возможность изоляции от ареала C. vaporariorum (Linnaeus 1758). Как было показано ранее, значительная вариабельность формы вершины ламеллы у C. vaporariorum дает возможность предположить, что C. laferi, имеющий отчетливо выраженный крючок на вершине ламеллы, мог в относительно короткое время дивергировать по этому признаку (Сундуков, 2011).
(5) “Стенопейские эндемики” (8 таксонов) – эндемики из подрода Aulonocarabus Reitter 1896 (2 вида с 6 эндемичными подвидами) рода Carabus и подрода Metallophilus Chaudoir 1838 (1 вид с 2 эндемичными подвидами) рода Pterostichus (Сундуков, 2019: рис. 1B, 2B, 4A).
Становление этих таксонов, на уровне подрода или видовой группы, происходило в миоценовых лесах материковой части Стенопейской подобласти (см. ниже раздел Основные этапы …).
(6) “Эндемики неясного происхождения” (2 таксона) – 2 вида подрода Curtonotus Stephens 1827 (Сундуков, 2019: рис. 2D), населяющие каменистые россыпи альпийской зоны Сихотэ-Алиня.
Филогенетические связи этих видов остаются невыясненными. Во-первых, это сильно дивергировавшая группа, не имеющая морфологических аналогов в подроде Curtonotus и, возможно, заслуживающая выделения в самостоятельный подрод (Сундуков, 2001). Во-вторых, в Сибири и на Дальнем Востоке нет высокогорных петрофильных видов этого подрода, а ближайшие альпийские группы Curtonotus из гор Средней Азии, Тибета и Сино-Тибетских гор, как и виды японского подрода Armatoleirides Tanaka 1957 (горы островов Хонсю и Сикоку), не демонстрируют апоморфных признаков с видами группы “larisae”.
“Рубежи” и “центры эндемизма” на Сихотэ-Алине
Анализ распределения эндемичных таксонов на Сихотэ-Алине показывает, что на территории этой горной страны можно выделить главные “рубежи” и “центры эндемизма”.
В качестве “центров эндемизма” выделяются: п-ов Муравьева-Амурского, Ливадийский хребет, Партизанский хребет, Синий хребет, группа высоких вершин в истоках р. Уссури, восточный макросклон Сихотэ-Алиня к северу от линии Спасск-Дальний – Дальнегорск, высокогорья Сихотэ-Алиня к северу от хр. Дальний (рис. 1). Ареалы большинства эндемиков приурочены либо к наиболее древним (Бровко и др., 2005) сооружениям Сихотэ-Алиня (хребты Ливадийский и Партизанский), либо к высокогорным районам, либо к его восточным приморским участкам, что подтверждает вывод о существовании главных рефугиумов аркто-третичной флоры на восточном макросклоне Сихотэ-Алиня (Крестов, 2010).
Помимо эндемичных таксонов, к этим же “центрам” на юге Сихотэ-Алиня приурочены реликты третичной (Eobroscus Kryzhanovskij 1951) и аркто-бореальной фаун (Nebria baicalopacifica Dudko et Shilenkov 2002, Miscodera arctica (Paykull 1798), Patrobus assimilis Chaudoir 1844, Harpalus lederi Tschitschérine 1899).
Важнейшим “рубежом” являются долины рек Уссури, Журавлёвка и Рудная, образующие на карте линию, соединяющую города Спасск-Дальний и Дальнегорск (рис. 1). Этот “рубеж” отделяет группу высоких вершин и хребтов южного Сихотэ-Алиня от остальной территории этой горной страны. Это предел распространения на север почти всех видов группы “восточноазиатские эндемики” и линия демаркации для ряда сестринских таксонов (виды Leistus, Lenapterus, Curtonotus, подвиды Carabus gossarei Haury 1879 и некоторые другие). Не столь явным, но достаточно показательным “рубежом” является главный водораздел Сихотэ-Алиня в пределах Приморского края. Он разграничивает ряд плиоцен-плейстоценовых эндемиков из рода Trechus, подродов Eosteropus и Feroperis, подвиды Carabus canaliculatus Adams 1812 и Pterostichus interruptus (Dejean 1828).
Особенности высотно-биотопической приуроченности эндемиков Сихотэ-Алиня
Анализ высотно-биотопической приуроченности сихотэ-алинских эндемиков выявил ряд взаимосвязей между их экологией, морфологическими адаптациями и происхождением. Наиболее характерными для этой группы можно считать следующие закономерности:
а) Все “восточноазиатские эндемики” в настоящее время на Сихотэ-Алине приурочены к узкой зоне верхнего пояса гумидных темнохвойных лесов (“умеренные дождевые леса”) или нижнего пояса высокогорий. Их реликтовые популяции прошли значительную адаптацию в условиях достаточно холодного климата в направлении гигрофильного гипогейного образа жизни. К таким же условиям адаптировались сохранившиеся до настоящего времени на Сихотэ-Алине реликты третичной фауны и эндемики гор Южной Сибири.
б) Эндемики неморального или автохтонного генезиса, адаптированные к обитанию на поверхности почвы или в верхнем слое подстилки. Большинство из них относится к эврибионтам лесного пояса, что согласуется с расселением реликтовой неморальной флоры из рефугиумов, сложившейся впоследствии в уникальный биом “уссурийская тайга” (Крестов и др., 2009).
в) Все высокогорные эндемики Сихотэ-Алиня, независимо от происхождения, приурочены к каменистым россыпям, что связывается нами с обширным развитием гольцовых ландшафтов в горных районах Азии во время плейстоценовых ледниковых периодов (Короткий, Бородкина, 2007), приведших к адаптациям этих таксонов к петрофильному образу жизни.
ОСНОВНЫЕ ЭТАПЫ ФОРМИРОВАНИЯ СОВРЕМЕННОЙ ЭНДЕМИЧНОЙ ФАУНЫ ЖУЖЕЛИЦ СИХОТЭ-АЛИНЯ
Время, прошедшее до наступления кайнозойской эры, не оставило следов в формировании современной фауны жужелиц Сихотэ-Алиня. Мезозойская карабидофауна еще полностью состоит из ископаемых родов и видов (Мартынов, 1936; Пономаренко, Волков, 2013; Handlirsch, 1906; Heer, 1852; Hong, 1981, 1982, 1984, 1985, 1987, 1995; Hong, Wang, 1990; Jia et al., 2011; Ren, 1995; Wang et al., 2009; Zhang, 1997 и другие), а Сихотэ-Алинь как суша начинает существовать лишь с середины мелового периода, когда 97 млн лет назад из моря поднялись древнейшие горстовые хребты – Ливадийский и Партизанский (Бровко и др., 2005). Поэтому интересующий нас исторический период начинается с палеогенового периода. Ниже выделены основные исторические события, сыгравшие наибольшую роль в формировании современной эндемичной фауны жужелиц Сихотэ-Алиня.
(1) Первая половина палеогенового периода
В палеоценовой фауне жужелиц планеты уже известны ископаемые виды современных родов, населяющих в настоящее время Дальний Восток России (Bembidion, Agonum, Harpalus, Lebia), а эоценовая фауна представлена значительным числом современных родов. По данным молекулярно-генетических исследований, в палеоцене начинается радиация основных групп в роде Carabus (Prüser, 1996), а к концу эоцена уже сформировались линии трибы Trechini – Epaphius Leach 1819 и Trechus (Faille et al., 2013). Второй половиной эоцена датируется минимальный возраст для рода Pterostichus (Schmidt et al., 2012). Вероятно, в палеоцене должны были сформироваться род Masuzoa и клады филетической линии Epaphiopsis, представляющие наиболее архаичные типы трибы Trechini (Uéno, 1982). А в эоценовую эпоху предковые формы нынешних сихотэ-алинских эндемиков из родов Masuzoa, Epaphiopsis, Trechiama и реликтов третичной фауны Eobroscus и Amerizus Chaudoir 1868 могли широко расселиться в Восточной Азии.
В этот же период происходил межконтинентальный обмен элементами формирующейся третичной фауны между Азией и Северной Америкой. Например, на основе молекулярного датирования, возраст возникновения на “американо-восточноазиатской территории” предковой группы Megaxyela Ashmead 1898 (Xyelidae) определяется серединой палеоцена, а ископаемые виды этого рода известны из олигоцена Северной Америки и миоцена Китая (Blank et al., 2017). Среди жужелиц, сходные дизъюнкции ареала у сравнимых по возрасту таксонов известны в трибах Metriini (2 рода: Metrius Eschscholtz 1829 в североамериканских Кордильерах и Sinometrius Wrase et Schmidt 2006 в горах южного Китая) и Broscini (3 монофилетических рода: Eobroscus и Kashmirobroscus Schmidt, Wrase et Sciaky 2013 в Гималаях и Восточной Азии и Zacotus LeConte 1869 в Кордильерах от Калифорнии до южной Аляски) (Roig-Juñent, 2000; Schmidt et al., 2013; Wrase, Schmidt, 2006) (рис. 2B, 2F).
(2) Граница эоцена–олигоцена
Происходят значительные изменения в составе морской и наземной флоры и фауны, известные в отношении европейской фауны как “великий перелом” (Алексеев, 1989). Современные исследования показывают, что в этот период произошло быстрое изменение климата Земли. Резкое похолодание в Северной Америке, установление засушливого климата в Азии и образование ледяного щита Антарктиды произошли всего в течение 400 тысяч лет (Bowen, 2007; Dupont-Nivet et al., 2007; Zanazzi et al., 2007). Эти события нередко связывают со “столкновением” Индии с Азией и начавшимся подъемом Гималаев и Тибета (например, Sun et al., 2014).
Вероятно, в этот период начала формироваться дизъюнкция ареала рода Craspedonotus Schaum 1863. В настоящее время 2 вида этого рода (C. margellanicus Kraatz 1884 и C. himalayanus Semenov 1910) обитают на песчаных берегах рек Средней Азии, Ирана, Пакистана и южных предгорий Западных Гималаев, а третий (C. tibialis Schaum 1863) – на морских побережьях Восточной Азии и в провинции Сычуань на юге Китая (рис. 2C). Возможно, в эоцене предковая форма этого рода населяла песчаные биотопы умеренной Азии вдоль побережий Тургайского моря и Тихого океана. С этим же периодом можно связать начало дивергенции предковой формы на роды Eobroscus и Kashmirobroscus (рис. 2В).
Усиление аридизации климата в центральном и северном Китае стало причиной разрыва единых ареалов гумидной восточноазиатской фауны на “южную” и “северную” части.
(3) Олигоцен–начало миоцена
Начало образования Японского моря (рис. 3A) запускает процесс разделения материковых и островных таксонов. Климат в этот период становится еще холоднее, достигнув минимума в его конце (Павлюткин, Голозубов, 2010). Продолжающееся похолодание вызывает дальнейшую аридизацию (Sun, Wang, 2005; Xu et al., 2015), что приводит к постепенному разрыву американо-азиатских и восточноазиатских ареалов, закреплению уже образовавшихся дизъюнкций и возникновению новых. Генетические исследования показали, что поздний олигоцен – ранний миоцен являются временем обособления восточноазиатского рода Grylloblattina Bey-Bienko 1951 и западно-американского Grylloblatta Walker 1914 (Grylloblattidae) (рис. 2E) (Schoville, Kim, 2011). Подобный же разрыв наблюдается в ареале рода Cryptocercus (Cryptocercidae), 7 видов которого обитают в Восточной Азии и 5 видов – в США (рис. 2D) (Grandcolas et al., 2005; Hossain, Kambhampati, 2001). Также олигоценом датируется дизъюнкция между восточноазиатским Callipogon relictus (Cerambycidae) и неотропическими видами этого рода (Kim et al., 2018).
Можно предположить, что в среднем миоцене произошло разделение ареала у жужелиц рода Amerizus на три части – североамериканскую, южно-азиатскую и североазиатскую. При этом мы не согласны с предположением о “юго-восточноазиатском происхождении” этого рода и его проникновением на Алтай “восточным путем”, то есть вдоль тихоокеанского побережья Восточной Азии и, далее, по горам Южной Сибири (Дудко, 2011). По нашему мнению, в олигоцене Amerizus был широко расселен на территории Азии и Северной Америки. Аридизация климата в центральном Китае, достигшая максимума в начале миоцена, привела к разрыву ареала Amerizus на южную (Гималаи, Тибет, Сино-Тибетские горы) и северную части (Северная Азия), что стало причиной дивергенции рода на подроды Amerizus s. str. и Tiruka Andrewes 1935.
Схожий сценарий эволюции таксонов, приведший к формированию современных восточноазиатских ареалов мы предполагаем и для сихотэ-алинских эндемиков из родов Epaphiopsis, Trechiama и Trechus из филетической линии Epaphius. Возможно, какие-то из перечисленных групп в этот период населяли Северную Америку, в пользу чего говорит находка Trechiama в позднеплейстоценовых отложениях на северо-западе США (Cong, 1997).
Вероятно, фрагментация ареалов сопровождалась адаптацией таксонов палеофауны Восточной Азии к новым условиям. К концу периода усиление климатических и флористических различий между материковыми и приокеаническими районами Азии приводит к внутриродовой дифференциации и возникновению современных подродов в крупных родах. Например, молекулярно-генетические исследования показывают, что дивергенция комплекса Leptocarabus Géhin 1885 рода Carabus началась на материке около 25 млн л. н. (Osawa et al., 2004). Отделение подрода Ethira Andrewes 1936 от рода Pterostichus датируется началом миоцена, а его проникновение в Гималаи и Сино-Тибетские горы происходило из бореальной Азии (Schmidt et al., 2012). Вероятно, в этот же период в приокеанических лесах Восточной Азии могло происходить формирование подрода Aulonocarabus.
(4) Средний миоцен
В среднем миоцене происходит восстановление климата до эоценового (Короткий и др., 1996). На юге Дальнего Востока России в это время развились хвойно-широколиственные леса, напоминающие современную растительность Южного Китая (Крестов, Верхолат, 2002; Лопатина, 2004). В это же время в центре Китая и на Тибете наблюдается увеличение площади лесостепных ландшафтов под влиянием зимнего Восточно-Азиатского муссона (Jacques et al., 2013; Liu et al., 2016; Zhang et al., 2012).
Совпавшее с потеплением климата начало активного горообразования в Азии предоставляет возможность адаптированным к холодному климату представителям гумидной фауны проникать в горы, что закрепляет образовавшиеся в центре Азии разрывы ареалов, а автохтонам лесов умеренной Азии позволяет распространиться к северу.
На основе молекулярного датирования возраст многих современных подродов рода Carabus, в том числе восточноазиатских Ohomopterus Reitter 1896, Diocarabus Reitter 1896, Tomocarabus Reitter 1896 и Leptocarabus s. str. определяется началом среднего миоцена (Andújar et al., 2012; Osawa et al., 2004). Мы предполагаем, что в это же время произошли окончательное обособление и начало адаптации к горным условиям таких групп сихотэ-алинских эндемиков, как Leptepaphiama, Epaphiama, Eonebria, Xestagonum и гумидные Trechus.
Сильное потепление климата отодвинуло границы сформировавшихся в конце олигоцена на северо-востоке Азии темнохвойных лесов к северу, что стало причиной распространения к северу и дальнейшего видообразования в таких группах, как Aulonocarabus, Lenapterus, Cryobius.
Расширение Японского моря в начале среднего миоцена (рис. 3B, 3C) приводит к окончательному разрыву материковых и островных ареалов гумидных групп жужелиц Trechiama, Epaphiopsis, Masuzoa. Причем к этому времени Masuzoa мог быть монотипическим родом, на что указывают одинаковые строение эдеагуса и внешняя морфология его современных видов.
(5) Конец миоцена–плиоцен
Климатический оптимум миоцена сменился похолоданием (Величко и др., 2015). В отличие от других районов Евразии, климат южного Китая становится теплее и влажнее, что связано (Zhang et al., 2012) с увеличением высоты Гималаев и Тибета, отгородивших Южный Китай от зимнего муссона.
Произошедшая около 8–7 млн лет назад еще большая аридизация климата на большей части центрального и северного Китая, в Тибете и Сибири (Wang et al., 2014), могла послужить причиной для становления и широкого распространения в горно-таежных районах Северной Азии видовых групп “almonius”, “montanus” и “chalybeus” рода Trechus.
В середине этого периода на большей части Сибири начинают доминировать леса из лиственницы, а в приокеанических районах северо-восточной Азии развивается темнохвойная тайга (Крестов и др., 2009). Это могло стать причиной возникновения сибирско-дальневосточных дизъюнкций у Masuzoa, Epaphiama и Grylloblattidae, разрыва связей между азиатской и американской частью ареалов Amerizus и лесных видов Lenapterus и Eosteropus. В это время могла формироваться и видовая группа “motdkovischi” рода Trechus. Интересно, что современные американо-азиатские связи в роде Trechus сохранились у одного вида – T. apicalis Motschulsky 1845, что косвенно подтверждает возможность возникновения габитуально и экологически близких групп “montanus” и “chalybeus” в темнохвойных лесах обоих континентов.
Палеонтологические данные показывают, что Якутия, Чукотка и Аляска в это время были населены рядом современных аркто-бореальных подродов и даже видов (Kiselev, Nazarov, 2009; Hopkins et al., 1971). Существование этих таксонов в Арктике позднего миоцена – раннего плиоцена до появления первых тундровых сообществ указывает, что формирование современных арктических таксонов надвидового ранга происходило в лесной зоне северной Азии, а дальнейшая радиация шла вслед за образованием зональных тундр. Например, для восточноазиатского по происхождению подрода Aulonocarabus, можно предложить следующую схему: а) возникновение в лесах Восточной Азии в раннем–среднем миоцене; б) адаптация к горным темнохвойным лесам в среднем миоцене; в) адаптация к высокогорным биотопам в конце миоцена–начале плиоцена; г) расселение в середине плиоцена адаптированных к высокогорным ландшафтам таксонов на северо-востоке Азии и северо-западе Северной Америки на участках палеотундры; д) активное заселение в позднем плиоцене тундровых и степных сообществ, господствующих на территории Берингии и Восточной Сибири; е) распространение в зональных тундрах аркто-бореальных таксонов в плейстоцене. Близкий сценарий становления подрода Lenapterus в бореальных лесах северо-восточной Азии, после его обособления от общего с Eosteropus предка в миоцене, ранее нами уже рассматривался (Сундуков, 2005).
Оценивая таксономический состав арктической карабидофауны в целом, логично предположить, что ее становление происходило исключительно за счет аллохтонных элементов надвидового (а, возможно, и видового) ранга, прошедших начальные стадии формирования в миоцен-плиоценовых ландшафтах и последующие адаптации к арктическим условиям в плейстоцене-голоцене. На это указывают и палеонтологические данные, и геологическая молодость зональной тундры, и высокая доля в арктической фауне аркто-умеренных таксонов, и современное распространение ее таксонов за пределы Арктики в горных областях, и присутствие в составе арктических биоценозов специализированных петробионтов, и легкость, с которой некоторые современные виды аркто-бореальных групп осваивают высокогорные гольцово-тундровые ландшафты. Вполне возможно, что морфологическая нестабильность, которая присуща многим арктическим таксонам и которую часто объясняют суровыми природными условиями Арктики, является результатом современного видообразования в этой природной зоне.
(6) Конец плиоцена–плейстоцен
Сильное похолодание конца плиоцена–плейстоцена (особенно позднего плейстоцена) оказало наиболее существенное влияние на современные состав и распространение фауны жужелиц Сихотэ-Алиня. Горный рельеф, долготное направление, приокеаническое положение и отсутствие покровного оледенения, позволили Сихотэ-Алиню сохранить на своей территории элементы почти всех фаун, развивающихся с момента его образования. Именно в силу хорошей изученности геологических событий этого периода, детальный анализ времени изоляции, видообразования и современной локации эндемичных таксонов этой горной страны чрезвычайно сложен. В рамках этого сообщения мы ограничимся попыткой выявить общие закономерности изменений, происходящие с сихотэ-алинскими эндемиками в плейстоцене.
С исчезновением широколиственных лесов в центре Сибири в первой половине этого периода связаны разрывы ареалов в группе “транспалеарктических неморальных эндемиков”.
Несомненно, что в плейстоцене неоднократно создавались условия для разобщения представителей гумидной фауны Сихотэ-Алиня и Маньчжуро-Корейских гор. В настоящее время это проявляется в обитании в этих горах близкородственных видов как миоценовых (Eonebria, Xestagonum), так и плиоцен-плейстоценовых (Cymindis s. str., Feroperis, Eosteropus) изолятов. Эти же условия явились причиной разделения видов C. venustus Morawitz 1862 и C. wulffiusi Morawitz 1862, подвидов C. gossarei, C. smaragdinus Fischer von Waldheim 1823, C. hummeli Fischer von Waldheim 1823, C. canaliculatus или стали преградой для проникновения на Сихотэ-Алинь C. jankowskyi Oberthür 1883, C. fraterculus Reitter 1895, Pristosia vigil (Tschitschérine 1895) и других. Вероятно, это происходило в среднем плейстоцене, когда лесная растительность сокращалась до горно-приморских рефугиумов (Крестов и др., 2009). На примере C. gossarei можно предположить, что некоторые дизъюнкции образовывались не позже минделя (300 тыс. л. н.), когда о-в Уллындо в последний раз соединялся с Корейским п-овом (рис. 3D, 3E) (Ломтев, 2012; Minato, Ijiri, 1976), и на нем обособилась самостоятельная популяция C. g. ullungensis Deuve et Li 2007. Во время последних и наиболее сильных похолоданий, случившихся в конце плейстоцена, флористические рефугиумы приморских склонов Сихотэ-Алиня и Маньчжуро-Корейских гор стали убежищами и местами формирования современных эндемичных видов Feroperis, подвидов C. canaliculatus, Pt. tuberculiger (Tschitschérine 1897), Pt. interruptus и некоторых других.
Вероятно, лесные эндемики Сихотэ-Алиня (Pt. sutschanensis Jedlička 1962, Cymindis laferi), широко распространенные на его территории и имеющие родственные связи с бореальными таксонами, должны были пройти свой эволюционный путь в достаточной изоляции от близких видов. Такие возможности предоставлялись во время сильных трансгрессий раннего (2.0–1.8 млн л. н.) и среднего (700–375 тыс. л. н.) плейстоцена, когда Сихотэ-Алинь мог становиться островом (Берсенев, Безверхий, 1991). Дальнейшая эволюция этих видов происходила в горных темнохвойных лесах, сохраняющихся на Сихотэ-Алине в течение всего плейстоцена, включая его конец. Возможно, ранний плейстоцен стал и временем образования горно-таежного C. gossarei, ограниченного в своем распространении на север долиной р. Амур. В этот же период могли начать формирование на Сихотэ-Алине популяции высокогорных эндемиков Leistus, Curtonotus, Lenapterus и лесного Trechus sikhotealinus Uéno et Lafer 1994. Возможно, с этим же временем связано разделение близких видов – материкового C. canaliculatus и островного C. kurilensis Lapouge 1913.
Помимо этого, можно отметить ряд закономерностей в современном распространении эндемичных жужелиц на Сихотэ-Алине. Например, виды трибы Trechini из группы “восточноазиатские эндемики” (Masuzoa, Epaphiama, Trechiama) в настоящее время ограничены только югом Сихотэ-Алиня. При этом их ареалы невелики и не перекрываются. То есть, если на Сихотэ-Алине и сохранялось разнообразие третичной фауны жужелиц, то в этот период оно было сведено к минимуму. Современное обитание Epaphiopsis semenovi Jeannel 1962 только на Синем хребте можно объяснить тем, что в самый холодный период плейстоцена именно на этом хребте и Шкотовском плато сохранились достаточно большие массивы темнохвойных лесов (рис. 4) (Короткий и др., 1996). А разделение предковой формы Leptepaphiama на два вида, вероятно, произошло в результате регрессии моря и последовавшей за этим интенсивной денудацией в раннем эоплейстоцене, разделившей соседние Ливадийский и Партизанский хребты глубокой долиной р. Партизанская (Берсенев, Безверхий, 1991).
Еще одна закономерность проявляется в распространении высокогорных эндемиков. Группы Leistus, Lenapterus и Curtonotus имеют по одному эндемичному виду в горах к югу и северу от “рубежа” Спасск-Дальний – Дальнегорск. Видовой статус этих эндемиков и специализация к петрофильному образу жизни указывают на их ранне- или средне-плейстоценовый возраст. Вероятно, видообразование в этих группах проходило в два этапа: 1) образование общего эндемичного таксона в раннем или среднем плейстоцене и 2) разделение таксона на два близких вида в межледниковые периоды минделя, рисса или вюрма. Возможно, эту же схему можно применить к эволюции в группе “Trechus densicornis”.
(7) Голоцен
В голоцене оформились современный рельеф и растительность Сихотэ-Алиня, что окончательно определило современный состав фауны и распространение эндемичных таксонов.
Современная приуроченность высокогорных эндемиков (Masuzoa ussuriensis, Leistus janae Farkač et Plutenko 1992, Pterostichus galae Farkač et Plutenko 1996, Pt. tokmakovae Sundukov 2013) к высочайшим вершинам южного Сихотэ-Алиня явилась результатом климатического оптимума голоцена (8–4.5 тыс. л. н.), сопровождавшегося сокращением площади высокогорных местообитаний. Например известно, что в этот период на вершине горы Ольховая (1669 м) росли кедрово-широколиственные леса и полностью отсутствовали высокогорные ландшафты (Короткий и др., 2009). Вероятно, в голоцене сформировался реликтовый ареал Harpalus lederi, распространенный в горных районах Магаданской области, северного Забайкалья и на единственной вершине южного Сихотэ-Алиня – г. Снежная. Возможно, в плейстоценовое время этот вид был шире распространен на Сихотэ-Алине, но исчез на значительной территории при потеплении. С голоценовым оптимумом так же можно связать начало образования подвидов Pt. galae и возникновение небольших современных ареалов Trechus tardokijanensis Lafer 1989 и T. badzhalensis Plutenko 2004.
Другим проявлением потепления в голоцене стало расширение ареалов видов и возникновение зон интерградации между подвидами Feroperis, C. canaliculatus, Pt. interruptus. Сохраняясь в плейстоцене в лесных рефугиумах, в настоящее время они широко расселились в лесной зоне и даже осваивают высокогорья. Некоторые из них в настоящее время находятся в стадии активного видообразования.
Нельзя не отметить усилившееся влияние человека на состав локальных фаун. Формирование антропогенных ландшафтов и активные перемещения человека по планете, особенно в последнее столетие, приводят к исчезновению аборигенных видов и интродукции чужеродных элементов. Для относительно слабо освоенного человеком Сихотэ-Алиня наибольшую роль играют дороги, довольно густой сетью покрывающие всю его территорию. Виды родов Cicindela Linnaeus 1758, Poecilus Bonelli 1810, Dolichus Bonelli 1810, Amara, Anisodactylus Dejean 1829, Harpalus широко используют дороги, просеки и другие свободные пространства, проникая в самые отдаленные лесные районы.
Влияние человека на локальную фауну рассматривалось нами на примере жужелиц острова Шикотан (Сундуков, 2014). Доля видов, заселивших населенные пункты и интразональные местообитания этого острова за последние 40 лет, составляет не менее 30%, что указывает на возрастающее значение адвентивных элементов в формировании его современной фауны.
Вероятно, уже можно говорить о возникающей зависимости некоторых групп от человека. Например, многие виды подрода Pseudoophonus Motschulsky 1844 в настоящее время населяют почти исключительно культурные ландшафты, и для них остаются невыясненными естественные места обитания. Особенно много этих видов на востоке Китая, где естественная растительность была уничтожена или изменена несколько тысячелетий назад.
История Восточной Азии, и Сихотэ-Алиня в частности, уникальна тем, что ее фауна перенесла плейстоценовое похолодание без покровного оледенения, избежав эпох полного уничтожения, имевших место в Европе, Сибири и Северной Америке. Благодаря сохранению высокой влажности в период позднеплейстоценовой аридизации климата, восточный макросклон и крайний юг Сихотэ-Алиня стали важнейшими рефугиумами фауны, сформировавшейся на восточной окраине Евразии в плиоцене. Сохранение здесь эндемиков и реликтов третичной гумидной фауны было одной из ключевых причин становления в голоцене уникального биома “уссурийская фауна” на современной территории Приморья и Приамурья.
Список литературы
Алексеев А.С., 1989. Глобальные биотические кризисы и массовые вымирания в фанерозойской истории // Меннер В.В. (ред.). Биотические события на основных рубежах фанерозоя. Москва: МГУ. С. 22–47.
Белоусов И.А., Кабак И.И., 1994. К познанию жужелиц рода Trechus Clairv. (Coleoptera, Carabidae) Саяно-Алтайской горной системы // Selevinia. № 3. С. 10–22.
Берсенев И.И., Безверхий В.Л., 1991. Глава 5. Японское море и тихоокеанский шельф Японских островов // Атлас палеографических карт шельфы Евразии в мезозое и кайнозое. Т. 1. Текст. Москва: Геологический институт АН СССР. С. 5(1–11).
Бровко П.Ф., Берсенев Ю.И., Петренко В.С., Пшеничников Б.Ф., Пшеничникова Н.Ф. и др., 2005. Шкотовский район. Владивосток: Дальневосточный государственный университет. 186 с.
Величко А.А., Фаустова М.А., Писарева В.В., Тумской В.Е., Борисова О.К. и др., 2015. Формирование экосистем Арктической зоны в условиях зарождения и развития криосферы в высоких широтах Евразии. Москва: Институт географии РАН. 17 с.
Городков К.Б., 1984. Типы ареалов насекомых тундры и лесных зон Европейской части СССР // Ареалы насекомых Европейской части СССР, карты 179–221. Ленинград: Наука. С. 3–20.
Дудко Р.Ю., 2011. О реликтовых жесткокрылых (Coleoptera: Carabidae, Agyrtidae) с алтайско-восточноазиатским дизъюнктивным ареалом // Евразиатский энтомологический журнал. Т. 10. С. 349–360 + + 348 + VI.
Короткий А.М., Бородкина М.А., 2007. Динамика многолетней мерзлоты в четвертичное время (Юг Дальнего Востока) // V Всероссийское совещание по изучению четвертичного периода. Фундаментальные проблемы квартера: итоги изучения и основные направления дальнейших исследований. 7–9 ноября 2007 г. Москва. С. 190–193.
Короткий А.М., Гребенникова Т.А., Пушкарь В.С., 1996. Климатические смены на территории юга Дальнего Востока в позднем кайнозое (миоцен–плейстоцен). Владивосток: ДВГУ. 58 с.
Короткий А.М., Коробов В.В., Скрыльник Г.П., 2009. Изменение растительности в верхнем поясе гор под действием экзогенных процессов (поздний плейстоцен – голоцен) // Бюллетень Ботанического сада-института ДВО РАН. Вып. 4. С. 41–49.
Крестов П.В., 2010. Рефугиумы – “окна” в историю растительного покрова // Наука в России. № 5. С. 105–112.
Крестов П.В., Баркалов В.Ю., Омелько А.М., Якубов В.В., Накамура Ю., Сато К., 2009. Реликтовые комплексы растительности современных рефугиумов северо-восточной Азии // Комаровские чтения. Владивосток: Дальнаука. Вып. 56. С. 5–63.
Крестов П.В., Верхолат В.П., 2002. Редкие растительные сообщества Приморья и Приамурья. Владивосток: ДВО РАН. 200 с.
Ломтев В.Л., 2012. Новое в строении котловины и трога Уллындо (Японское море) // Вестник КРАУНЦ. Науки о Земле. № 1. Вып. 19. С. 1–10.
Лопатина Д.А., 2004. Растительность и климат эоцена–миоцена Восточного Сихотэ-Алиня по данным палеоботанического анализа // Тихоокеанская геология. Т. 23. № 3. С. 98–112.
Мартынов А.В., 1936. О некоторых новых материалах членистоногих животных из Кузнецкого бассейна // Известия Академии наук СССР, серия биологическая. Вып. 6. С. 1251–1264.
Павлюткин Б.И., Голозубов В.В., 2010. Палеоботанические свидетельства времени возникновения Японского моря // Вестник КРАУНЦ. Науки о Земле. № 2. Вып. 16. С. 19–26.
Плутенко А.В., 2004. Новые данные о видах рода Trechus (Coleoptera, Carabidae) Дальнего Востока России // Евразиатский энтомологический журнал. Т. 3. С. 33–36.
Пономаренко А.Г., Волков А.Н., 2013. Ademosynoides asiaticus Martynov, 1963 – древнейший представитель ныне существующего семейства жесткокрылых (Insecta, Coleoptera, Trachypachidae) // Палеонтологический журнал. № 6. С. 41–46.
Пономаренко А.Г., Жерихин В.В., 1980. Надотряд Scarabaeidae. Жесткокрылообразные // Историческое развитие класса насекомых, Труды Палеонтологического института АН СССР, Т. 175. Москва: Наука. С. 75–84.
Сундуков Ю.Н., 2001. Два новых вида рода Curtonotus Stephens (Coleoptera, Carabidae) с юга Дальнего Востока России // Энтомологическое обозрение. Т. 80. С. 436–442.
Сундуков Ю.Н., 2005. Обзор видов подрода Lenapterus (Coleoptera, Carabidae, Pterostichus), с описанием новых вида и подвида с юга Сихотэ-Алиня // Зоологический журнал. Т. 84. С. 803–825.
Сундуков Ю.Н., 2011. Обзор рода Cymindis Latreille 1806 (Coleoptera, Carabidae, Lebiini) Восточной Азии // Амурский зоологический журнал. Т. 3. С. 315–344.
Сундуков Ю.Н., 2014. Особенности формирования современной фауны жужелиц (Coleoptera, Carabidae) острова Шикотан, Курильские острова // Чтения памяти А.И. Куренцова. Вып. 25. С. 25–33.
Сундуков Ю.Н., 2019. Основные этапы формирования фауны жужелиц (Coleoptera, Carabidae) Сихотэ-Алиня на примере эндемиков. 1. Характеристика таксонов // Зоологический журнал. Том 98. № …
Andújar C., Gómez-Zurita J., Rasplus J.-Y., Serrano J., 2012. Molecular systematics and evolution of the subgenus Mesocarabus Thomson, 1875 (Coleoptera: Carabidae: Carabus), based on mitochondrial and nuclear DNA // Zoological Journal of the Linnean Society. V. 166. P. 787–804.
Arillo A., Ortuño V.M., Nel A., 1997. Description of an enigmatic insect from Baltic amber // Bulletin de la Société Entomologique de France. V. 102. P. 11–14.
Ashworth A.C., Nelson R.E., 2014. The paleoenvironment of the Olympia beds based on fossil beetles from Discovery Park, Seattle, Washington, U.S.A. // Quaternary International. V. 341. P. 243–254.
Blank S.M., Kramp K., Smith D.R., Sundukov Y.N., Wei M., Shinohara A., 2017. Big and beautiful: the Megaxyela species (Hymenoptera, Xyelidae) of East Asia and North America // European Journal of Taxonomy. № 348. P. 1–46.
Bowen G.J., 2007. Palaeoclimate: When the world turned cold // Nature. Vol. 445. P. 607–608.
Cai C., Liu Y., Huang D., 2017. A new species of Loricera Latreille from Eocene Baltic amber (Coleoptera: Carabidae: Loricerinae) // Alcheringa. V. 41. Iss. 3. P. 315–320.
Cockerell T.D.A., 1925. Tertiary insects from Kudia River, Maritime Province, Siberia // Proceedings of the United States National Museum. V. 68. № 5. P. 1–16.
Cockerell T.D.A., 1936. The fauna of the Sunchal (or Margas Verdes) formation, Northern Argentina // American Museum Novitates. V. 886. P. 1–9.
Cong S., 1997. Fossils of an undescribed blind trechine (Coleoptera: Carabidae) from near Kalaloch, Olympic Peninsula, Washington // The Coleopterists Bulletin. V. 51. № 3. P. 208–211.
Cong S.G., Ashworth A.C., 1996. Palaeoenvironmental interpretation of Middle and Late Wisconsinan fossil coleopteran assemblages from western Olympic Peninsula, Washington, USA // Journal of Quaternary Science. V. 11. P. 345–356.
Dupont-Nivet G., Krijgsman W., Langereis C.G., Abels H.A., Dai S., Fang X., 2007. Tibetan plateau aridification linked to global cooling at the Eocene–Oligocene transition // Nature. V. 445. P. 635–638.
Erwin T.L., 1971. Fossil tachyine beetles from Mexican and Baltic amber // Entomologica Scandinavica. V. 2, № 3. P. 233–236.
Faille A., Casale A., Balke M., Ribera I., 2013. A molecular phylogeny of Alpine subterranean Trechini (Coleoptera: Carabidae) // BMC Evolutionary Biology. V. 13. № 248. P. 1–16.
Germar E.F., 1813. Insecten in Bernstein eingeschlossen, beschrieben aus dem academischen Mineralien-Cabinet zu Halle // Magazin der Entomologie. Bd. 1. S. 11–18.
Gersdorf E., 1969. Käfer (Coleoptera) aus dem Jungtertiär Norddeutschlands // Geologisches Jahrbuch. Bd. 87. S. 295–332.
Giebel C.G., 1856. Die Insecten und Spinnen der Vorwelt mit steter Berücksichtigung der lebenden Insekten und Spinnen // Die Fauna der Vorwelt. Bd. 2. S. 1–511.
Grandcolas P., Legendre F., Park Y.C., Bellés X., Murienne J., Pellens R., 2005. The genus Cryptocercus in East Asia: distribution and new species (Insecta, Dictyoptera, Blattaria, Polyphagidae) // Zoosystema. V. 27. P. 725–732.
Handlirsch A., 1906. Die Fossilen Insekten und die Phylogenie der Rezenten Formen, parts I–IV // Ein Handbuch fur Palaontologen und Zoologen. S. 1–640.
Hayashi M., 1996. Insect fossil assemblage and paleoenvironments from the Early Pleistocene Bushi Formation in Saitama Prefecture, Japan // Earth Science (Chikyu Kagaku). V. 50. P. 223–237.
Hayashi M., 1998. Early Pleistocene ground beetles (Coleoptera: Carabidae) from the Ookui Formation in Nagano Prefecture, central Japan, and their biogeographical and paleoenvironmental significance // The Quaternary Research. V. 37. P. 117–129.
Hayashi M., Yahiro K., Kitabayashi E., 2003. Fossil insects and their paleoenvironments of the Pliocene Tsubusagawa Formation in Ajimu Basin, Oita Prefecture, Kyushu, Japan // Bulletin of the Mizunami Fossil Museum. V. 30. P. 167–173.
Hayashi M., Yahiro K., Kitabayashi E., 2005. Middle Pleistocene fossil insects from the Hirabaru Formation of Kitsuki City, Oita Prefecture, Kyushu, Japan // Bulletin of the Mizunami Fossil Museum. V. 32. P. 227–234.
Hayashi M., Yahiro K., Kitabayashi E., 2008. Middle Pleistocene fossil insects from the Yoshino Formation of Kawachimachi, Kumamoto City, Kyushu, Japan // Bulletin of the Mizunami Fossil Museum. V. 34. P. 95–98.
Hayashi M., Yahiro K., Kitabayashi E., 2009. Middle Pleistocene fossil insects from the Nogami Formation of Kokonoe, Oita Prefecture, Kyushu, Japan // Bulletin of the Mizunami Fossil Museum. V. 35. P. 105–110.
Heer O., 1852. Die Lias-Insel des Aargau’s // Zwei Geologische Vorträge Gehalten im März. 1852(1). S. 1–15.
Hong Y.C., 1981. Discovery of new Early Cretaceous insects from Xishan, Beijing // Bulletin of the Tianjin Institute of Geology and Mineral Resources. V. 4. P. 87–96.
Hong Y.C., 1982. Mesozoic Fossil Insects of Jiuquan Basin in Gansu Province. Beijing: Geological Punlishing House. 187 p.
Hong Y.C., 1984. Insecta // Palaeontological Atlas of North China. II. Mesozoic Volume. P. 128–185.
Hong Y.C., 1985. Early Jurassic fossil insects from Daye of Hubei Province // Professional Papers of Stratigraphy and Palaeontology. V. 15. P. 181–187.
Hong Y.C., 1987. The study of Early Cretaceous insects of Kezuo, west Liaoning // Professional Papers of Stratigraphy and Palaeontology. V. 18. P. 76–87.
Hong Y.C., 1995. Fossil insects of the northern Ordos Basin // Acta Geologica Gansu. V. 4. № 2. P. 1–9.
Hong Y.C., Wang W.L., 1990. Fossil insects from the Laiyang Basin, Shandong Province // Stratigraphy and Palaeontology of Laiyang Basin, Shandong Province. P. 44–189.
Hopkins D.M., Matthews J.V., Wolfe J.A., Silberman M.L., 1971. A Pliocene flora and insect fauna from the Bering Strait region // Palaeogeography, Palaeoclimatology, Palaeoecology. V. 9. P. 211–231.
Hossain S., Kambhampati S., 2001. Phylogeny of Cryptocercus species (Blattodea: Cryptocercidae) inferred from nuclear ribosomal DNA // Molecular Phylogenetics and Evolution. V. 21. Iss. 1. P. 162–165.
Jacques F.M.B., Shi G., Wang W.M., 2013. Neogene zonal vegetation of China and the evolution of the winter monsoon // Bulletin of Geosciences. V. 88. № 1. P. 175–193.
Jia T., Liang H., Chang H., Ren D., 2011. A new genus and species of fossil Eodromeinae from the Yixian Formation of western Liaoning, China (Coleoptera: Adephaga: Trachypachidae) // Zootaxa. № 2736. P. 63–68.
Kim S., de Medeiros A.S., Byun B.-K., Lee S., Kang J.-H., et al., 2018. West meets East: How do rainforest beetles become circum-Pacific? Evolutionary origin of Callipogon relictus and allied species (Cerambycidae: Prioninae) in the New and Old Worlds // Molecular Phylogenetics and Evolution. V. 125. P. 163–176.
Kiselev S.V., Nazarov V.I., 2009. Late Cenozoic insects of northern Eurasia // Paleontological Journal. V. 43. № 7. P. 732–850.
Liu J., Li J.J., Song C.H., Yu H., Peng T.J. et al., 2016. Palynological evidence for late Miocene stepwise aridification on the northeastern Tibetan Plateau // Climate of the Past. V. 12. P. 1473–1484.
Manchester S.R., 1992. Flowers, fruits, and pollen of Florissantia, an extinct Malvalean genus from the Eocene and Oligocene of western North America // American Journal of Botany. V. 79. P. 996–1008.
Matthews J.V., Westgate J.A., Ovenden L., Carter L.D., Fouch T., 2003. Stratigraphy, fossils, and age of sediments at the upper pit of the Lost Chicken gold mine: new information on the late Pliocene environment of east central Alaska // Quaternary Research. V. 60. P. 9–18.
Minato M., Ijiri S., 1976. Japanese Archipelago. Tokyo: Iwanami-shinnsho. 209 p. [In Japanese].
Morgan A.V., Morgan A., 1979. The fossil Coleoptera of the Two Creeks Forest Bed, Wisconsin // Quaternary Research. V. 12. P. 226–240.
Omboni G., 1886. Di alcuni insetti fossili del Veneto // Atti del Reale Istituto Veneto di Scienze, Lettere ed Arti, Serie Sesta. V. 4. P. 1421–1436.
Osawa S., Su Z.-R., Imura Y., 2004. Molecular phylogeny and evolution of Carabid ground beetles. Springer Japan KK. XIV + 191 p.
Piton L., 1940. Paléontologie du Gisement Éocéne de Menat (Puy-de-Dôme) (Flore et Faune) // Mémoire de la Société d’Histoire Naturelle d’Auvergne. V. 1. P. 1–303.
Piton L., Rudel M.A., 1936. Sur de nouveaux gisements d’insectes fossiles dans l’Oligocène de la Limagne // Bulletin Mensuel de la Société Linnéenne de Lyon. V. 5. № 5. P. 78–81.
Piton L., Théobald N., 1936. Les insectes fossiles du Puy-de-Mur (Limagne) // Bulletin Mensuel de la Société des Sciences de Nancy. V. 9. P. 202–214.
Pongrácz A., 1935. Die eozäne Insektenfauna des Geiseltales // Nova Acta Leopoldina. V. 2. P. 485–572.
Prüser F., 1996. Variabilität mitochondrialer DANN-Sequenzen und die Phylogenie der Gattung Carabus Linné 1758 (Coleoptera: Carabidae) // Dissertation Universität Bremen. S. 1–173.
Ren D., 1995. Insecta // Ren D. (Ed.): Faunae and stratigraphy of Jurassic-Cretaceous in Beijing and the adjacent areas. Beijing: Seismology Publishing House. P. 47–121.
Roig-Juñent S.A., 2000. The subtribes and genera of the tribe Broscini (Coleoptera: Carabidae): cladistic analysis, taxonomic treatment and biogeographical considerations // Bulletin of the American Museum of Natural History. V. 255. P. 1–90.
Schmidt J., Faille A., 2015. Confirmation of occurrence of Trechus Clairville, 1806 in the Baltic amber forests, with description of a flightless edaphic species, and remarks on Trechoides Motschulsky 1856 (Coleoptera: Carabidae: Trechini) // Zootaxa. № 4028. P. 539–550.
Schmidt J., Hoffmann H., Michalik P., 2016. Blind life in the Baltic amber forests: description of an eyeless species of the ground beetle genus Trechus Clairville, 1806 (Coleoptera: Carabidae: Trechini) // Zootaxa. № 4083. P. 431–443.
Schmidt J., Opgenoorth L., Höll S., Bastrop R., 2012. Into the Himalayan exile: The phylogeography of the ground beetle Ethira clade supports the Tibetan origin of forest-dwelling himalayan species groups // PLoS ONE. V. 7. № 9. P. 1–15.
Schmidt J., Wrase D.W., Sciaky R., 2013. Description of Kashmirobroscus gen. nov. with two new species from the Northwest Himalaya, and remarks on the East Asian genus Eobroscus Kryzhanovskij, 1951 (Coleoptera: Carabidae: Broscini) // Journal of Natural History. P. 1–19.
Schoville S.D., Kim B.W., 2011. Philogenetic relationships and relictualism of rock-crawlers (Grylloblattodea: Grylloblattidae) in cave and mountain habitats of Korea // Annals of the Entomological Society of America. V. 104. P. 337–347.
Scudder S.H., 1879. Appendix A. The fossil insects collected in 1877, by Mr. G.M. Dawson, in the interior of British Columbia // Geological Survey of Canada, Report of Progress for 1877–1878. P. 175–185.
Scudder S.H., 1890. The tertiary Insects of North America // Report of the United States Geological Survey of the Territories. V. 13. P. 1–734.
Shilenkov V.G., 1982. Some notes on South Siberian Trechini (Coleoptera, Carabidae) with descriptions of three new species // Annales Historico-Naturales Musei Nationalis Hungarici. V. 74. P. 85–91.
Sun J., Xu Q., Liu W., Zhang Z., Xue L., Zhao P., 2014. Palynological evidence for the latest Oligocene – early Miocene paleoelevation estimate in the Lunpola Basin, central Tibet // Palaeogeography, Palaeoclimatology, Palaeoecology. V. 399. P. 21–30.
Sun X., Wang P., 2005. How old is the Asian monsoon system? – Palaeobotanical records from China // Palaeogeography, Palaeoclimatology, Palaeoecology. V. 222. Iss. 3–4. P. 181–222.
Théobald N., 1937. Les insectes fossiles des terrains oligocènes de France // Bulletin Mensuel (Mémoires) de la Société des Sciences de Nancy. V. 1. P. 1–473.
Uéno Sh.-I., 1982. Origin and dispersal of the Trechina in East Asia (Coleoptera: Carabidae) // Entomologia Generalis. V. 8. P. 71–78.
Wang B., Zhang H.C., Fang Y., 2009. Paleogeographical distribution of Mesozoic Palaeontinidae (Insecta, Hemiptera) in China with description of new taxa // Acta Geologica Sinica. V. 84. P. 31–37.
Wang P.X., Wang B., Cheng H., Fasullo J., Guo Z.T. et al., 2014. The global monsoon across timescales: coherent variability of regional monsoons // Climate of the Past. V. 10. P. 2007–2052.
Wickham H.F., 1910. New fossil Coleoptera from Florissant, with notes on some already described // American Journal of Science, Series 4. V. 29. P. 47–51.
Wickham H.F., 1913. Fossil Coleoptera from the Wilson Ranch near Florissant, Colorado // Bulletins from the Laboratory of Natural History of the State University of Iowa. V. 6. № 4. P. 3–29.
Wipfler B., Bai M., Schoville S., Dallai R., Uchifune T. et al., 2014. Ice Crawlers (Grylloblattodea) – the history of the investigation of a highly unusual group of insects // Journal of Insect Biodiversity. V. 2. № 2. P. 1–25.
Wrase D.W., Schmidt J., 2006. A first representative of the tribe Metriini in the Palaearctic: Sinometrius turnai gen. nov., spec. nov. (Insecta: Coleoptera: Carabidae: Metriini) // Hartmann M., Weipert J. (Eds): Biodiversität und Naturausstattung im Himalaya. Bd. 2. S. 315–324.
Xu Q., Qiu J., Zhou Z., Jin J., 2015. Eocene Podocarpium (Leguminosae) from South China and its biogeographic implications // Frontiers in Plant Science. V. 6. Iss. 938. P. 1–14.
Zanazzi A., Kohn M.J., MacFadden B.J., Terry D.O., 2007. Large temperature drop across the Eocene–Oligocene transition in central North America // Nature. V. 445. P. 639–642.
Zhang H.C., 1997. Early Cretaceous insects from the Dalazi Formation of the Zhixin basin, Jilin Province, China // Palaeoworld. V. 7. P. 75–103.
Zhang J.F., 1989. Fossil insects from Shanwang, Shandong, China // Shandong Science and Technology Publishing House. P. 1–459.
Zhang Q.-Q., Ferguson D.K., Mosbrugger V., Wang Y.-F., Li C.-S., 2012. Vegetation and climatic changes of SW China in response to the uplift of Tibetan Plateau // Palaeogeography, Palaeoclimatology, Palaeoecology. № 363–364. P. 23–36.
Дополнительные материалы отсутствуют.
Инструменты
Зоологический журнал