

Зоологический журнал, 2020, T. 99, № 1, стр. 113-120
Экспериментальное исследование охотничьего поведения скальных полевок Alticola strelzowi и Alticola tuvinicus (Rodentia, Cricetidae)
С. Н. Пантелеева a, b, *, Я. В. Левенец a, А. А. Новиковская a, Ж. И. Резникова a, b, Н. В. Лопатина a, Ю. Н. Литвинов a
a Институт систематики и экологии животных Сибирского отделения РАН
630091 Новосибирск, Россия
b Новосибирский национальный исследовательский государственный университет
630090 Новосибирск, Россия
* E-mail: psofia@mail.ru
Поступила в редакцию 14.10.2018
После доработки 28.01.2019
Принята к публикации 28.02.2019
Аннотация
В лабораторных экспериментах у двух видов зеленоядных скальных полевок – тувинской и плоскочерепной – впервые выявлено и детально описано целенаправленное взаимодействие с подвижными насекомыми по типу “хищник–жертва”. Стереотипы охотничьего поведения у этих двух видов полностью сходны между собой. Проявление полного охотничьего стереотипа у скальных полевок не требует предварительного опыта и не совершенствуется, что указывает на его врожденный характер. По сравнению с ранее исследованными видами грызунов, у скальных полевок впервые были выявлены элементы поведения, связанные с попытками “складирования” живой добычи. По характеристикам взаимодействия с добычей они стоят в ряду наиболее успешных и эффективных охотников на подвижных насекомых. По тактике охоты и характеру манипуляций с добычей скальные полевки сходны с наиболее “хищными” из всеядных видов – эверсманновыми хомячками, но отличаются от них факультативным проявлением охотничьего стереотипа. Частота встречаемости стереотипа и успешность охоты у скальных полевок вдвое выше, чем у ранее исследованной зеленоядной узкочерепной полевки. Столь неожиданные для зеленоядных грызунов характеристики охотничьего поведения выявлены у скальных полевок впервые. Расширение рациона за счет охоты на насекомых может объясняться обитанием в засушливых биотопах с дефицитом растительных пищевых ресурсов.
Известно, что беспозвоночные в той или иной мере входят в состав рациона практически всех видов грызунов, в том числе и тех, которых принято считать зеленоядными (Landry, 1970; Башенина, 1977). Высокая ценность беспозвоночных как дополнительных пищевых объектов и их разнообразие в рационе многих видов грызунов неоднократно обсуждались в литературе (обзор см.: Левенец и др., 2016). До недавнего времени охотничье поведение по отношению к подвижной добыче исследовалось, главным образом, на примере хомячков: хищных южного (Onychomys torridus Coues 1874) и северного (O. leucogaster Wied-Neuwied 1841) кузнечиковых хомячков, всеядных белоногого (Peromyscus leucopus Rafinesque 1818) и оленьего (P. maniculatus Wagner 1845) хомячков (Langley, 1986; Kreiter, Timberlake, 1988; Timberlake, Washburne, 1989). Кузнечиковые хомячки обладают рядом морфологических и физиологических адаптаций к взаимодействию даже с ядовитыми беспозвоночными, такими как скорпионы (Rowe, Rowe, 2008; Sarko et al., 2011; Rowe et al., 2013). Недавно было детально описано охотничье поведение по отношению к насекомым у всеядных видов хомячков и были выявлены градации хищнического поведения: представители группы эверсманновых хомячков (Allocricetulus eversmanni Brandt 1859 и A. curtatus Allen 1925) оказались значительно более “насекомоядными”, чем мохноногие (Phodopus sungorus Pallas 1773 и P. сampbelli Thomas 1905), и особенности диеты у этих видов связаны со спецификой стереотипов охотничьего поведения (Левенец и др., 2019).
Для разнообразных зерноядных и зеленоядных грызунов, потребляющих животную пищу, до сих пор оставался неясным вопрос, способны ли эти зверьки расширять свой рацион за счет активной охоты, или они собирают беспозвоночных, находящихся в состоянии неподвижности. В последние годы в экспериментальных исследованиях было обнаружено охотничье поведение у рыжих полевок (Myodes glareolus Schreber 1780) со смешанным зерноядно-зеленоядным типом питания (Sadowska et al., 2008; Konczal et al., 2016), а также у зерноядных полевых мышей (Apodemus agrarius Pallas 1771) (Panteleeva et al., 2013; Reznikova et al., 2017), малой лесной мыши (Sylvaemus uralensis Pallas 1811) и зеленоядной узкочерепной полевки (Lasiopodomys gregalis Pallas 1779) (Левенец и др., 2016, 2016а). Оказалось, что некоторые виды зерноядных и зеленоядных грызунов применяют различные приемы для быстрого и эффективного умерщвления живых насекомых и демонстрируют лаконичные и структурированные охотничьи стереотипы, во многом сходные с таковыми у специализированных хищников (Пантелеева и др., 2011; Левенец и др., 2016а; Левенец, 2017; Reznikova et al., 2017). Хотя общая схема охотничьего поведения в принципе едина у всех хищников, паттерны у разных видов мышевидных грызунов включают различные варианты атаки на добычу и элементы манипуляторной активности, в большей или меньшей степени эффективные для операций с живыми объектами. Кроме того, существенно различаются как частота проявления охотничьих стереотипов в популяциях разных видов, так и степень участия индивидуального опыта (обзор см.: Левенец и др., 2019). У некоторых видов, например, у щетинистого хлопкового хомяка (Sigmodon hispidus Say et Ord 1825), охотничье поведение вообще не проявляется (Timberlake, Washburne, 1989). Облигатное и не требующее предварительного опыта проявление охотничьего стереотипа может свидетельствовать о большей специализации хищнического поведения, как это было выявлено, в частности, у эверсманновых хомячков – эволюционно более молодой группы по сравнению с мохноногими (Левенец и др., 2019). Важным признаком хищнической специализации у грызунов считается характер атаки: ее начало с захвата добычи лапами является одним из показателей эволюционной продвинутости охотничьего поведения по сравнению с захватом зубами (Eisenberg, Leyhausen, 1972; Langley, 1994). Для того чтобы подойти к пониманию эволюции хищнического поведения у грызунов, необходимо исследование широкого спектра видов, которые имеют разные типы питания и обитают в различных биотопах. В этом плане интересными и совершенно неизученными объектами являются представители рода скальных полевок Alticola, в частности, плоскочерепная (Alticola strelzowi Kastschenko 1899) и тувинская (A. tuvinicus Ognev 1950) полевки, сходные по морфологическим признакам, но различающиеся некоторыми экологическими особенностями.
Плоскочерепная полевка населяет обширные пространства Казахского мелкосопочника, Алтая, юго-западной Тувы, Монгольского Алтая и северо-западного Китая и встречается в сухих каменистых участках гор, начиная с высоты 400 м над ур. м. на северо-востоке Казахского нагорья и до 3000 м над ур. м. в Монгольском Алтае (Огнев, 1950; Громов, Ербаева, 1995). Основным фактором, определяющим размещение и плотность поселений, является наличие каменистых убежищ (Шубин, 1959; Капитонов, Кадырбаев, 1972). Образ жизни семейно-колониальный, молодые остаются с родителями до весны следующего года и запасают высушенную траву на зиму. Состав запасаемых растений может включать в разных частях ареала до 68 видов (Капитонов, Кадырбаев, 1972; Юдин и др., 1979). Об употреблении плоскочерепной полевкой животных кормов данных мало. Капитонов (1965) находил в поселениях полевок в Казахском нагорье остатки съеденных бабочек, стрекоз, жуков, саранчовых, моллюсков, реже – мумий рептилий, птиц, млекопитающих со следами резцов грызунов.
Ареал тувинской полевки состоит из разрозненных участков в Хакасии, центральной и южной Туве и северо-западной Монголии (Млекопитающие России, 2012). Заселяет выходы скал с разреженным травянистым покровом, групноглыбчатые осыпи. В Хакасии характерными местообитаниями являются остепненные участки известняковых мелкослоистых горных хребтов с осыпями и трещинами (Кохановский, 1977; Литвинов и др., 2013, 2013а). Для этого вида также характерен семейный образ жизни и запасание травы. В рацион входит около 40 видов растений, данных о животных кормах нет.
Оба вида скальных полевок по типу питания относятся к зеленоядным. Их поведение почти не изучено; лишь недавно было экспериментально показано, что различия между ними в выборе местообитания и в характере активности, проявляющиеся в естественных условиях, отражаются в таких показателях лабораторных тестов, как подвижность и тревожность (Задубровский и др., 2017).
Целью нашей работы является экспериментальное исследование и сравнительный анализ реакций на подвижную добычу у плоскочерепной и тувинской полевок, в сравнении с охотничьим поведением ранее исследованных нами грызунов, обладающих разными типами питания и экологической спецификой.
МАТЕРИАЛЫ И МЕТОДЫ
Исследования проводились в 2015–2017 гг. в лаборатории на двух видах полевок: плоскочерепной (A. strelzowi) (n = 33) и тувинской (A. tuvinicus) (n = 53). Часть особей плоскочерепной (n = 6) и тувинской полевки (n = 4) отловлена в естественных условиях, остальные животные являются их потомками первого и второго поколения. К началу экспериментов возраст 16 плоскочерепных и трех тувинских полевок составлял 30 дней, трех и одной, соответственно, 45 дней, возраст остальных варьировал от 3 до 18 месяцев. Точки отлова полевок: A. strelzowi – Республика Алтай, Кош-Агачский р-н, хребет Сайлюгем, верховья р. Уландрык; A. tuvinicus – Республика Хакасия, Ширинский р-н, 5 км ЮЗ п. Колодезный. Рожденные в лаборатории животные до исследования не контактировали с насекомыми. Зверьков содержали парами или семейными группами в клетках Ferplast Mini Duna Multy 55 × 39 × 27 см при постоянном доступе к воде и пище. В рацион входили свежее разнотравье или кормовое сено, сочные овощи, сухофрукты, творог, зерносмесь. Все эксперименты проведены с соблюдением правил проведения научных исследований с использованием экспериментальных животных, утвержденных распоряжением Президиума АН СССР от 2 апреля 1980 12 000-№ 496 и приказом Минвуза СССР от 13 сентября 1984 № 22.
Для исследования спонтанного проявления охотничьего поведения скальных полевок мы применили схему, ставшую стандартной (Левенец и др., 2016, 2016а, Левенец и др., 2019; Reznikova et al., 2017): животным предлагали подвижных мраморных (Nauphoeta cinerea Olivier 1789) и туркменских (Shelfordella lateralis Walker 1868) тараканов; средняя длина тела 27.93 ± 0.22 мм. Зверьков по одному помещали в прозрачную арену (45 × 45 × 50 см), передерживали 5 мин, после чего предъявляли добычу (аккуратно сбрасывали на арену сверху). Действия животных фиксировались с помощью видеокамер Sony Handycam DCR-SR68 и HDR–AS200V.
В данной работе мы применили два режима тестирования. В первом режиме плоскочерепных (n = 20) и тувинских полевок (n = 16) тестировали по одному разу. Если животное охотилось, то на арену помещали еще две единицы добычи, каждую после окончания манипуляций с предыдущей. Таким образом, зверькам в рамках одного теста предъявлялось максимум по три единицы добычи.
Во втором режиме каждого зверька (13 плоскочерепных и 37 тувинских полевок) тестировали один раз в день, 10 дней подряд (с двухдневным перерывом после 5-го дня). В каждом тесте животным предъявляли только по одному таракану, а не до трех, как в первом режиме тестирования. Проведение длительных серий тестов с каждым животным позволяло оценить роль индивидуального опыта в проявлении охотничьего поведения. В обоих режимах, если зверек не проявлял интерес к насекомому в течение 10 мин, тест прекращали.
Для описания поведения в качестве элементарной единицы нами выделяются элементарные двигательные акты и позы – “элементы поведения” (Tinbergen, 1951). Поведенческой последовательностью мы называем произвольный набор последовательно совершаемых элементов поведения. Внутри поведенческих последовательностей, полученных для каждого вида, мы выделяем поведенческий стереотип, состоящий из устойчиво повторяющихся “цепочек” элементов поведения (подробно см.: Reznikova et al., 2017). Обработка видеозаписей проводилась с 25- и 5-кратным замедлением в программе The Observer XT 12.5 (Noldus Information Technology). Используя полученный “алфавит” из 15 элементов, мы преобразовали демонстрируемое поведение в последовательности букв, где каждая буква соответствовала одному элементу поведения. Для анализа отбирались только случаи успешной охоты, закончившиеся поимкой добычи. Началом охоты считалось активное взаимодействие с добычей (преследование, захват насекомого зубами или лапами), окончанием – элемент поведения, предшествующий либо поеданию добычи, либо прекращению манипуляций с ней. Используя полученный “алфавит” из 15 элементов поведения, мы преобразовали демонстрируемое поведение в последовательности букв, где каждая буква соответствовала одному элементу поведения. Полученные последовательности символов через пробел перемещались в отдельные для каждого вида “суммарные” текстовые файлы (в формате .txt). Как и ранее (Reznikova et al., 2017; Левенец и др., 2019), каждый суммарный текстовый файл включал все зафиксированные поведенческие последовательности, в том числе полученные при повторных тестированиях зверьков. Это позволило выявить все связи между элементами, выделить среди них наиболее вероятные и описать поведенческие стереотипы для исследуемых видов.
Для построения схемы стереотипа на основе суммарных текстовых файлов рассчитывались матрицы вероятностей перехода от одного поведенческого элемента к другому (Марковский процесс первого порядка) (Casarrubea et al., 2008). Матрица размера m × m позволяет отразить связанность последующего элемента поведения с предшествующим, где m – размер “алфавита”, из элементов которого состоит анализируемая поведенческая последовательность. Значение вероятности появления элемента i при условии, что предыдущим элементом был элемент j, указано в ячейке на пересечении i-го столбца и j-й строки. На основании полученных матриц выделяются схемы охотничьих стереотипов, в которых отражены порядок совершения элементов поведения и вероятности перехода от одного элемента к другому (их связанность).
Для оценки различий в охотничьем поведении между видами мы сравнили стереотипы охоты, полученные при первом режиме тестирования и первых трех тестов при втором. С помощью точного теста Фишера сравнивали доли охотившихся и не охотившихся особей у разных видов, успешных и неуспешных атак у разных особей, а также доли различных типов элементов поведения. Сравнения количества элементов в стереотипах и “скорости охоты” (количества элементов поведения в единицу времени) проведены с помощью H-критерия Краскела-Уоллиса. При сравнении представлены медиана, первый и третий квартили: Me (Q1–Q3).Чтобы оценить возможную роль индивидуального опыта, мы сравнивали охотничье поведение животных в первых трех и семи последующих тестах при втором режиме тестирования.
РЕЗУЛЬТАТЫ
Элементы охотничьего поведения у тувинской и плоскочерепной полевки оказались одинаковыми. Мы разделили их на 3 типа. 1) “Ключевые”, связанные с обнаружением, преследованием и захватом добычи, без которых совершение стереотипа невозможно: сближение с добычей бегом (Q) или спокойным шагом (S), укус (W), захват добычи лапами (E). 2) “Дополнительные”, связанные с “приготовлениями” к охоте и с “обработкой” добычи и присутствующие не во всех стереотипах: принюхивание (D), перенос добычи в зубах (G), перехват (R), откусывание конечностей насекомого (H). 3) “Шумовые”, не связанные непосредственно с процессом охоты и не влияющие на совершение стереотипа: поворот корпуса на 90° (V), разворот корпуса 180° (B), вертикальная стойка (I), стойка с опорой на арену (Y), движения назад (U), прыжок (J) и чистка (X). Отдельно отмечались элементы, выходящие за рамки охотничьего стереотипа: поедание добычи (T) и опускание добычи на дно арены (Z).
У обоих видов скальных полевок охотничий стереотип проявлялся факультативно (не у всех животных). При первом предъявлении подвижной добычи доли особей, проявивших охотничье проведение, у двух видов достоверно не различались (точный тест Фишера, p > 0.05): у плоскочерепных полевок – 49% (16 из 33 особей), у тувинских полевок – 68% (36 из 53).
Успешная охота заканчивалась поимкой добычи, после чего животные переходили к поеданию либо переносили таракана в зубах в угол арены и опускали на пол. Это поведение, ранее не отмеченное у других грызунов, мы называем “складирование”. Характерно, что во многих случаях зверек подталкивал добычу носом, как бы “прикапывая” ее. За все время эксперимента отмечено 50 охот, закончившихся “складированием” добычи: 1 случай (3% от всех охот) у плоскочерепной и 49 случаев (22%) у тувинской полевки, а поедание отмечалось в 28 (97%) и 178 (78%) случаях соответственно. “Складированный” таракан мог убежать из угла арены; если полевка находила его, она могла снова его поймать и либо начать поедать, либо повторно “складировать”. В 26 случаях тувинские полевки (n = 6) возвращали добычу в угол арены: в восьми случаях 1 раз, в двух случаях 2 раза, в трех случаях 3 раза, в одном случае 6 раз. В 27 случаях тувинские полевки (n = 7) после сближения с добычей брали ее в зубы и сразу без каких-либо манипуляций переносили в угол арены, где опускали на пол. У плоскочерепных полевок подобное поведение не наблюдалось, но они могли уже после поедания насекомого “складировать” его остатки.
Успешность охоты на первых трех предложенных тараканов у обоих видов оказалась сходной: 28 из 52 (54%) охот у плоскочерепных и 104 из 179 (58%) у тувинских полевок закончились поимкой добычи. Длины стереотипов (количество элементов в них) и скорость охоты (количество совершаемых элементов в секунду) также сходны: у плоскочерепных полевок – 9 (6–11) элементов, 1.6 (1.3–1.9) элемента в секунду, у тувинских полевок 6 (5–10) и 1.5 (1.1–2.0) соответственно (H = 1.0, NS; H = 0.5, NS).
Для того чтобы составить представление о том, как оба вида полевок взаимодействуют с добычей, мы сравнили доли различных типов элементов в охотничьих стереотипах и количество элементов поведения, связанных с атакой и обработкой добычи. У плоскочерепных и тувинской полевок доли ключевых (75 и 72%), дополнительных (20 и 22%) и шумовых элементов (5 и 6%) были сходными. Не различалось и количество элементов поведения “укус” (W), “захват лапами” (E), “перехват” (R) и “откусывание конечностей” (H), приходящихся на один стереотип (H ≤ 3, NS для всех случаев). Результаты представлены в таблице. Для того чтобы понять, каким образом полевки повреждают добычу, мы исследовали повторяемость элемента поведения “укус” (W). Вся совокупность этих элементов была разделена на три группы: одиночные, двойные (два последовательно совершаемых “укуса”) и множественные (три и более “укусов” подряд). Для обоих исследованных видов были характерны, главным образом, одиночные укусы, реже двойные (табл. 1).
Таблица 1.
Количество элементов поведения, приходящихся на один охотничий стереотип у исследованных видов полевок
| Элементы поведения | Плоскочерепная полевка | Тувинская полевка |
|---|---|---|
| Укус (W) | 3 (1–3) | 2 (1–4) |
| Захват лапами (E) | 2 (1–3) | 2 (1–2) |
| Перехват (R) | 0 (0–1) | 0 (0–1) |
| Откусывание конечностей (H) | 0 (0–0) | 0 (0–0) |
| Одиночный укус (W1) | 1 (1–2) | 1 (1–2) |
| Двойной укус (W2) | 0 (0–1) | 0 (0–0.5) |
| Множественные укусы (Wn) | 0 (0–0) | 0 (0–0) |
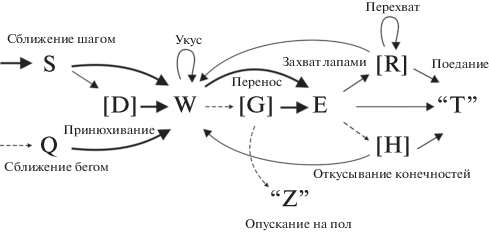
На основании матриц вероятностей переходов между элементами поведения построены схемы охотничьих стереотипов (рис. 1), которые у обоих видов скальных полевок оказались сходны. Охотничий стереотип начинался со сближения с добычей спокойным шагом (S), крайне редко бегом (Q), затем могло следовать принюхивание (D). Атака почти всегда начиналась с захвата зубами (“укус”, W), затем следовал захват добычи передними лапами (E). Крайне редко зверьки предпринимали попытки захвата таракана лапами, но всегда упускали его. После поимки грызуны могли переносить добычу в зубах (G) или начать с ней манипулировать: чаще всего перехватывать (R), существенно реже – откусывать конечности (H). После чего переходили к поеданию добычи (“T”). В отдельных случаях после манипуляций повторно следовал захват добычи зубами – “укус” (W), перенос (G) в угол арены и опускание добычи на дно (“Z”), в таком случае охотничий стереотип переходил в “складирование”.
Рис. 1.
Схема охотничьего стереотипа плоскочерепной и тувинской полевки. Тонкой пунктирной линией обозначены некоторые неустойчивые связи между элементами (p < 0.2). Простой линией обозначены устойчивые связи (0.2 ≤ p < 0.5). Жирной линией обозначены высоко устойчивые связи между элементами (p ≥ 0.5). В квадратных скобках указаны дополнительные элементы, в кавычках – элементы, не входящие в охотничий стереотип (поедание и “складирование” добычи).
При втором режиме тестирования тувинских полевок, то есть после проведения 10 тестов, охотничье поведение проявилось у 32 из 34 особей (из 37 животных 3 погибли), таким образом к 29 зверькам, которые охотились во время первых трех тестов, прибавились 3. Успешность охоты в целом по выборке увеличилась незначимо: с 63 до 67%. У плоскочерепных полевок число охотников и успешность охоты не изменились на протяжении 10 тестов. Схемы охотничьего стереотипа у обоих видов оставались неизменными. Не изменились на протяжении 10 тестов и такие параметры как длина стереотипа, скорость охоты, доли ключевых, дополнительных и шумовых элементов поведения, количество приходящихся на один стереотип “укусов” (W), “захватов лапами” (E), “перехватов” (R) и “откусывания конечностей” (H) между первыми тремя и последующими семью тестами не различались у обоих видов.
ОБСУЖДЕНИЕ
У двух видов зеленоядных скальных полевок – тувинской и плоскочерепной – впервые выявлено и детально описано целенаправленное взаимодействие с подвижной добычей по типу “хищник – жертва”, с демонстрацией полного стереотипа охотничьего поведения. Частота проявления и успешность охотничьего поведения у них не различалась (49 и 52% у плоскочерепной, 68 и 54% у тувинской полевок соответственно). По этим параметрам скальные полевки оказались сходны с эврифагом серой крысой (51.8 и 61.9% соответственно) и зерноядной полевой мышью (65.4 и 69.2%) (Левенец и др., 2016, 2016а; Reznikova et al., 2017) По сравнению с зеленоядной же узкочерепной полевкой (18.5 и 25.4%), у скальных полевок успешность охоты существенно выше и сопоставима с тем же показателем у насекомоядного вида – обыкновенной бурозубки (62.3%) (Reznikova et al., 2017). Скорость охоты оказалась универсальным показателем, сходным у скальных полевок как со специализированными хищниками (бурозубкой и кузнечиковыми хомячками), так и с ранее исследованными нами серой крысой, полевой мышью, узкочерепной полевкой и хомячком Кэмпбелла (Левенец и др., 2016, 2016а; Левенец и др., 2019; Reznikova et al., 2017).
Несмотря на некоторые различия в выборе местообитаний и характере активности тувинской и плоскочерепной полевок, у них полностью совпадают схемы охотничьего стереотипа и все параметры поведения по отношению к добыче: соотношение долей разных элементов (в частности, элементов “атаки” и “обработки” добычи), длины стереотипов, скорость охоты.
Тактика охоты у скальных полевок оказалась такой же, как у облигатных “хищников” – эверсманновых хомячков (Левенец и др., 2019): манипуляции с удерживаемой передними лапами добычей и ее быстрое повреждение чаще одним, реже двумя–тремя точными укусами. Это существенно отличает паттерны скальных полевок и эверсманновых хомячков от других грызунов, исследованных нами ранее. Так, полевая мышь и узкочерепная полевка повреждают добычу продолжительной серией укусов, которые они беспорядочно наносят в разные части тела насекомого, хомячок Кэмпбелла обездвиживает добычу с помощью специфического приема, быстро обкусывая ее конечности, серая крыса поедает пойманную добычу без предварительного повреждения (Левенец, 2017). Можно считать, что манипуляции с добычей у скальных полевок, как и у эверсманновых хомячков, носят более специализированный характер, чем у полевой мыши и узкочерепной полевки: они не “искусывают насмерть”, а наносят насекомому быстрые и точные повреждения. Способ атаки на добычу у скальных полевок более примитивный, чем у эверсманновых хомячков, и сходен с таковым у серой крысы, полевой мыши, узкочерепной полевки и джунгарского хомячка (Левенец и др., 2016, 2016а; Левенец и др., 2019): они захватывают насекомое зубами и только затем перехватывают лапами. Это важно отметить, так как начало атаки с захвата добычи лапами является одним из показателей хищнической специализации (Eisenberg, Leyhausen, 1972; Langley, 1994). Кузнечиковые хомячки начинают атаку на добычу с захвата лапами почти всегда (Langley, 1986, 1994; Rowe et al., 2013), эверсманновы хомячки – в четверти случаях (Левенец и др., 2019), у остальных же исследованных нами видов грызунов, в том числе скальных полевок, такой способ атаки не встречается.
Проявление полного охотничьего стереотипа у “наивных” взрослых и неполовозрелых зверьков обоих видов скальных полевок, как оказалось, не требовало предварительного опыта. После проведения 10 тестов (при втором режиме тестирования) у зверьков обоих видов не выявлено существенных различий между первыми тремя и последующими семью охотами. Животные не совершенствуют охотничий стереотип, в стереотипе не возникают новые элементы поведения, не происходит усиление связей между элементами. Это указывает на врожденный и высоко специфичный характер стереотипа охотничьего поведения у исследованных видов. По этому признаку скальные полевки сходны со специализированными хищными кузнечиковыми (Onychomys spp.) (Langley 1986, 1994) и эверсманновыми хомячками (Левенец и др., 2019), а также с хищником-генералистом серой крысой (Левенец, 2017).
Следует отметить, что у скальных полевок впервые были выявлены элементы поведения, связанные с попытками “складирования” живой добычи, не встречавшиеся у ранее исследованных видов. Эти реакции проявлялись как у взрослых особей, так и у 30–35-дневных детенышей, и при этом из 50 наблюдаемых случаев на долю тувинской полевки приходится 49, и лишь один случай зафиксирован для плоскочерепной. Можно предположить, что у тувинской полевки мы наблюдали попытки запасания насекомых, а “прикапывание” живых тараканов – это “сбой программы”, который нередко происходит у разных видов животных в неволе. В литературе имеются указания на “запасание насекомых” хомячком эверсманна (обзор см.: Феоктистова, 2008; Левенец и др., 2019), охотничье поведение которого как раз и оказалось сходным с таковым у скальных полевок. Пока остается неизвестным, поедают ли животные высохших насекомых из своих запасов в естественных условиях. Интересно отметить, что при высокой степени сходства охотничьего поведения обоих видов скальных полевок, запасание живой добычи отмечено главным образом у тувинской полевки и, видимо, может оказаться характерным видовым признаком.
В целом, зеленоядные плоскочерепная и тувинская полевки по показателям охотничьей активности существенно отличаются от зеленоядной же узкочерепной полевки, а по сравнению с исследованными ранее видами зерноядных и всеядных грызунов и даже специализированными хищниками (Timberlake, Washburne, 1989), они стоят в ряду наиболее успешных и эффективных охотников на подвижных насекомых. По тактике охоты, включающей манипуляторную активность и точное повреждение жертвы, скальные полевки сходны с наиболее “хищными” из всеядных видов – эверсманновыми хомячками. От всех “хищных” видов скальные полевки, так же как и исследованные нами ранее зерноядные и всеядные грызуны, отличаются факультативным проявлением охотничьего стереотипа, но при этом важно отметить, что частота встречаемости стереотипа у скальных полевок почти вдвое выше, чем у зеленоядной узкочерепной полевки. Столь неожиданные для зеленоядных грызунов характеристики охотничьего поведения выявлены у скальных полевок впервые. Стоит отметить, что скальные полевки обитают в засушливых местах со скудной растительностью, в условиях нехватки пищевых ресурсов, так же, как и эверсманновы хомячки (обзор см.: Левенец и др., 2019). Возможно, сходство условий местообитания отчасти объясняет использование этими видами насекомых как дополнительного пищевого ресурса. В совокупности наши данные по видам грызунов с разными пищевыми предпочтениями позволяют предположить широкое распространение стереотипа охотничьего поведения. У систематически далеких групп этот стереотип имеет специфические черты не только различий, но и сходства. Это позволяет предположить общее происхождение элементов стереотипа у разных видов от далеких предковых форм.
Список литературы
Башенина Н.В., 1977. Пути адаптации мышевидных грызунов. М.: Наука. 354 с.
Громов И.М., Ербаева М.А., 1995. Млекопитающие фауны России и сопредельных территорий. Зайцеобразные и грызуны. СПб.: Определители по фауне России, издаваемые Зоологическим институтом РАН. Вып. 167. 522 с.
Задубровский П.А., Степанова А.В., Лопатина Н.В., Литвинов Ю.Н., 2017. Поведение неполовозрелых скальных полевок двух видов (Alticola strelzowi и A. tuvinicus) в тесте “открытое поле” // Сибирский экологический журнал. Т. 24. № 3. С. 257–263.
Капитонов В.И., 1965. Плоскочерепная полевка на севере Казахского нагорья // Материалы научной конференции по природной очаговости и профилактике чумы. Алма-Ата. 668 с.
Капитонов В.И., Кадырбаев Х.К., 1972. Плоскочерепная полевка (Alticola strelzowi Kastsch.) в Калбинском Алтае // Бюллетень Московского общества испытателей природы, отд. биол. Т. 72. № 3. С. 54–57.
Кохановский Н.А., 1977. К экологии высокогорной серебристой полевки // Экология, методы изучения и организация охраны млекопитающие горных областей. Свердловск: УНЦ АН СССР. С. 87–88.
Левенец Я.В., 2017. Оценка изменчивости и сложности поведенческих стереотипов на примере охотничьего поведения мелких млекопитающих. Автореф. дис. … канд. биол. наук. Новосибирск: Институт систематики и экологии животных СО РАН. 22 с.
Левенец Я.В., Пантелеева С.Н., Резникова Ж.И., 2016. Сравнительный анализ стереотипного поведения на примере процесса охоты у мелких млекопитающих // Экспериментальная психология. Т. 9. № 4. С. 68–78.
Левенец Я.В., Пантелеева С.Н., Резникова Ж.И., 2016а. Экспериментальное исследование питания насекомыми у грызунов // Евразиатский энтомологический журнал. Т. 6. № 6. С. 550–554.
Левенец Я.В., Пантелеева С.Н., Резникова Ж.И., Гуреева А.В., Феоктистова Н.Ю., Суров А.В., 2019. Экспериментальный сравнительный анализ охотничьего поведения четырех видов хомячков подсемейства Сricetinae // Зоологический журнал. Т. 96. № 6. С. 1–11.
Литвинов Ю.Н., Абрамов С.А., Лопатина Н.В., Чертилина О.В., 2013. Сообщества грызунов горностепных ландшафтов юга Сибири (фауно-экологический анализ) // Зоологический журнал. Т. 92. № 1. С. 99–105.
Литвинов Ю.Н., Чертилина О.В., Абрамов С.А., Лопатина Н.В., 2013а. Адаптивные стратегии размножения полевок открытых пространств гор юга Сибири и Северного Казахстана // Известия Иркутского университета. Серия “Биология. Экология”. Т. 6. № 2. С. 74–78.
Млекопитающие России. 2012. Систематико-географический справочник / ред. Павлинов И.Я., Лисовский А.А. М.: Товарищество научных изданий КМК. 604 с.
Огнев С.И., 1950. Звери СССР и прилежащих стран. Т. VII. М.: Изд-во Академии наук СССР. 760 с.
Пантелеева С.Н., Выгоняйлова О.Б., Резникова Ж.И., 2011. Рыжие лесные муравьи как потенциальная массовая добыча полевых мышей: результаты лабораторных экспериментов // Евразиатский энтомологический журнал. Т. 10. С. 99–103.
Феоктистова Н.Ю., 2008. Хомячки рода Phodopus. Систематика, филогеография, физиология, поведение, химическая коммуникация. М.: Товарищество научных изданий КМК. 414 с.
Шубин И.Г., 1959. Экология полевки Стрельцова в Казахском нагорье // Труды Института зоологии АН КазССР. Т. 10. С. 87–113.
Юдин Б.С., Галкина Л.И., Потапкина А.Ф., 1979. Млекопитающие Алтае-Саянской горной страны. Новосибирск: Наука. 293 с.
Casarrubea M., Sorbera F., Crescimanno G., 2008. Multivariate analysis of the modifications induced by an environmental acoustic cue on rat exploratory behavior // Physiology & Behavior. V. 93. № 4. P. 687–696.
Eisenberg J.F., Leyhausen P., 1972. The phylogenesis of predatory behavior in mammals // Zeitschrift für Tierpsychologie. V. 30. № 1. P. 59–93.
Konczal M., Koteja P., Orlowska-Feuer P., Radwan J., Sadowska E.T., Babik W., 2016. Genomic response to selection for predatory behavior in a mammalian model of adaptive radiation // Molecular Biology and Evolution. V. 33. № 9. P. 2429–2440.
Kreiter N., Timberlake W., 1988. Form and development of predation on crickets in adults of Peromyscus maniculatus bairdi and P. leucopus noveboracensis // Journal of Comparative Psychology. V. 102. № 3. P. 269–278.
Landry S.O., 1970. The Rodentia as omnivores // Quarterly Review of Biology. V. 45. P. 351–372.
Langley W.M., 1986. Development of predatory behavioral in the southern grasshopper mouse (Onychomys torridus) // Behaviour. V. 99. P. 275–295.
Langley W.M., 1994. Comparison of predatory behaviors of deer mice (Peromyscus maniculatus) and grasshopper mice (Onychomys leucogaster) // Journal of Comparative Psychology. V. 108. № 4. P. 394–400.
Panteleeva S., Reznikova Z., Vygonyailova O., 2013. Quantity judgments in the context of risk/reward decision making in striped field mice: first “count,” then hunt // Frontiers in Psychology. V. 4. P. 45–53.
Reznikova Z., Levenets J., Panteleeva S., Ryabko B., 2017. Studying hunting behaviour in the striped field mouse using data compression // Acta ethologica. V. 20(2). P. 165–173.
Rowe A.H., Rowe M.P., 2008. Physiological resistance of grasshopper mice (Onychomys spp.) to Arizona bark scorpion (Centruroides exilicauda) venom // Toxicon. V. 52. № 5. P. 597–605.
Rowe A.H., Xiao Y., Rowe M.P., Cummins T.R., Zakon H.H., 2013. Voltage-gated sodium channel in grasshopper mice defends against bark scorpion toxin // Science. V. 342. № 6157. P. 441–446.
Sadowska E.T., Baliga, Klimczyk K., Chrzascik K.M., Koteja P., 2008. Laboratory model of adaptive radiation: a selection experiment in the bank vole // Physiological and Biochemical Zoology. V. 81. № 5. P. 627–640.
Sarko D.K., Leitch D.B., Girard I., Sikes R.S., Catania K.C., 2011. Organization of somatosensory cortex in the Northern grasshopper mouse (Onychomys leucogaster), a predatory rodent // Journal of Comparative Neurology. V. 519. № 1. P. 64–74.
Timberlake W., Washburne D.L., 1989. Feeding ecology and laboratory predatory behavior toward live and artificial moving prey in seven rodent species // Animal Learning & Behavior. V. 17. P. 2–11.
Tinbergen N., 1951. The Study of Instinct. L.: Oxford University Press. 228 p.
Дополнительные материалы отсутствуют.
Инструменты
Зоологический журнал