

Зоологический журнал, 2020, T. 99, № 1, стр. 104-112
Есть ли запись зимней спячки на поверхности резцов у обыкновенного хомяка (Cricetus cricetus, Rodentia, Cricetidae)?
Г. А. Клевезаль a, *, Е. А. Зайцева b, **, Д. В. Щепоткин a, ***, Н. Ю. Феоктистова b, ****, М. М. Чунков c, *****, А. В. Суров b, ******
a Институт биологии развития им. Н.К. Кольцова РАН
119991 Москва, Россия
b Институт проблем экологии и эволюции им. А.Н. Северцова РАН
119071 Москва, Россия
c Прикаспийский институт биологических ресурсов ДНЦ РАН
367000
Республика Дагестан, Махачкала, Россия
* E-mail: klevezal@bk.ru
** E-mail: zaycevaolena@gmail.com
*** E-mail: dm_shchepotkin@mail.ru
**** E-mail: feoktistovanyu@gmail.com
***** E-mail: chunkov@mail.ru
****** E-mail: surov@sevin.ru
Поступила в редакцию 14.02.2019
После доработки 20.02.2019
Принята к публикации 28.03.2019
Аннотация
Для ряда видов зимоспящих грызунов ранее было показано, что характер спячки отражается на структуре дентина, и это можно видеть как на окрашенных срезах и шлифах резцов, так и на их поверхности. Для обыкновенного хомяка (Cricetus cricetus) характерны факультативная спячка и большая индивидуальная изменчивость динамики температуры тела в зимний период. Чтобы установить характер записи спячки на резцах у этого вида, исследовали нижние резцы особей, перезимовавших с имплантированными термонакопителями в естественных условиях городского парка г. Симферополя (Крым) (2 экз.) и в условиях, приближенным к естественным, в с. Хунзах (Дагестан) (2 экз.), а также резцы 19 особей, обнаруженных погибшими в конце зимы или начале весны в парке г. Симферополь и его окрестностях. Предварительное исследование резцов у хомяков, которых содержали в виварии (8 экз.) и которым вводили тетрациклин, подтвердило, что приросты на поверхности резцов – суточные. Это позволило у особей, зимовавших с термонакопителем, определять тот участок резца, который сформировался за время спячки. У трех из четырех таких особей на этих участках был нарушен рисунок суточных приростов, характерный для остальной части резца. Характер этих нарушений у разных особей был разный, и нарушения нельзя было интерпретировать как запись спячки. Ни у одного из этих хомяков не было характерного признака записи спячки – серии узких, более четких приростов. Такие приросты были выявлены только у одной из 19 особей из природной популяции. Таким образом, мы не обнаружили на резцах обыкновенного хомяка таких изменений рисунка поверхности, которые позволили бы однозначно судить о том, были ли у данной особи периоды гипотермии. Судя по опубликованным ранее данным, у других представителей подсемейства Cricetinae (например, у представителей родов Allocricetulus или Mesocricetus) запись спячки на резцах была обнаружена. Сравнение динамики температуры тела в зимний период у них и у обыкновенного хомяка дает основание предполагать, что на формирование узких приростов на поверхности резца могут влиять длительность и глубина периодов гипотермии, но определяющими факторами являются длительность периодов нормотермии и особенности питания зверьков в период спячки.
Известно, что у сусликов и сурков зимняя спячка отражается не только на характере суточных слоев дентина, которые видны на срезах зуба, но и “записывается” на его поверхности, образуя специфическую “зону спячки” на фоне суточных приростов (Rinaldi, 1999; Goodwin et al., 2005; Goodwin, Ryckman, 2006; Клевезаль, Лобков, 2008; Клевезаль, Щепоткин, 2012; Клевезаль, Ануфриев, 2013). Запись спячки на поверхности резцов обнаружена также у хомяков Радде (Mesocricetus raddei) (Клевезаль и др., 2012, 2018) и хомяков Брандта (Mesocricetus brandti) (Batavia et al., 2013). Она могла быть несколько разной у разных видов, но была одна общая черта – наличие серии приростов, более узких и четких, чем обычные суточные. Именно такая запись спячки обнаружена у представителей рода эверсманновых хомячков (Allocricetulus), монгольского хомячка (A. curtatus) и хомячка Эверсманна (A. eversmanni) (Клевезаль и др., 2015), у которых ранее была описана необычная спячка с глубокими, но непродолжительными и нерегулярными периодами гипотермии и длительными эпизодами нормотермии (Ушакова и др., 2010; Феоктистова и др., 2013).
Ближайшим родственником эверсманновых хомячков является обыкновенный хомяк (Cricetus cricetus) (Neumann et al., 2006; Lebedev et al., 2018), характер спячки которого отличается большой индивидуальной изменчивостью (Wassmer, Wollnik, 1997; Wassmer, 2004; Siutz et al., 2017). В зимнее время зверь либо постоянно находится в норе, демонстрируя длительные периоды гипотермии, прерываемые короткими эпизодами нормотермии, либо периодически проявляет наземную активность (Wassmer, 2004; наши наблюдения), и такую спячку правильнее считать факультативной. Ранее, у ископаемых обыкновенных хомяков на окрашенных препаратах и на шлифах дентина была обнаружена “зона спячки” (Трунова, Клевезаль, 1999; Трунова, 2001), однако запись спячки на поверхности резцов у этого вида не была подтверждена. Цель настоящего исследования – установить, возможна ли запись спячки на поверхности резцов у обыкновенного хомяка. В случае положительного решения этого вопроса у исследователей появилась бы возможность методически просто судить о протекании зимнего периода у особей, добытых или найденных весной погибшими в природе, а также у музейных образцов собранных в разные годы в этот период.
МАТЕРИАЛ И МЕТОДИКА
Материал исследования составляли особи из популяции обыкновенного хомяка, обитающей в парке Гагарина в г. Симферополь (Крым). Симферопольская популяция в настоящее время, самая крупная городская популяция данного вида в Европе (Feoktistova et al., 2013; Суров и др., 2015). Двум самцам в октябре–ноябре под анестезией препаратом Zoletil внутрибрюшинно имплантировали термонакопители (Петровский и др., 2008) (интервал измерения температуры 30 мин) и радиопередатчики, которые позволяли определять их местоположение. Обе особи успешно перезимовали в парке. Кроме того, были исследованы 19 особей из природной популяции. Эти хомяки были найдены погибшими в Симферополе и его окрестностях в конце зимы–начале весны 2016 и 2018 гг. Кроме того, в анализе использовали 2 экз. обыкновенного хомяка из Дагестана, которым, также как и животным из парка г. Симферополя, были внутрибрюшинно имплантированы термонакопители. Эти хомяки были пойманы в Кизилюртовском р-не в с. Чонтаул (43°18′18″ с.ш., 46°51′50″ в.д., высота 10 м над ур. м.) и перевезены в с. Хунзах (42°33′17″ с.ш., 46°42″ в.д., высота 1695 м над ур. м.) Хунзахского р-на, где они успешно перезимовали. До начала эксперимента животных содержали поодиночке, при естественном световом и температурном режиме. Пятого октября 2017 г. их поместили в отдельные жестяные короба площадью 100 × 100 см и высотой 130 см, которые были врыты в землю так, чтобы 30 см их высоты оставались на поверхности. В коробах были установлены домики с запасом корма (зерновые и овощи). 14 марта зверьки были выловлены, умерщвлены, термонакопители были извлечены.
Дополнительно 8 взрослых особей, отловленных в разных регионах России, в начале апреля 2018 г. на месяц поместили в виварий в г. Москве. Им вводили тетрациклин (внутримышечно из расчета 20 мг/кг) для маркировки ростовой зоны резца в момент введения.
От всех хомяков были взяты верхние и нижние резцы, но для исследования использовали только нижние. Резцы удаляли из черепа после его отваривания, осторожно очищали от пленок и исследовали под бинокуляром Leica StereoZoom 6 Photo, подбирая такой угол падения света, при котором рельеф поверхности резца выявлялся наиболее полно, после чего фотографировали цифровым аппаратом на фоне линейки с ценой деления 0.5 мм. Иногда для лучшего выявления приростов поверхность зачерчивали мягким грифелем. Приросты измеряли и подсчитывали на медиальной поверхности резцов. Промеры делали в программе Фотошоп по снимкам цифровым аппаратом. Резцы хомяков, содержавшихся в виварии, исследовали и фотографировали также и в ультрафиолетовом свете для обнаружения тетрациклиновой метки по желтой флуоресценции.
ТЕРМИНОЛОГИЯ
Мы называем базальной часть резца, расположенную в альвеоле, где лежит ростовая зона, и апикальной – свободную часть резца, подверженную стачиванию. Приросты – это поперечные валики на поверхности резца, границы которых видны под определенным углом падения света. Мы называем приросты четкими, если границы их легко различаются и нечеткими, когда разграничить отдельные приросты трудно, как ни меняй угол падения света. Четкие приросты могут быть контрастными, когда рельеф поверхности хорошо выражен, и слабо контрастными, но различимыми, когда рельеф выражен плохо, хотя приросты различимы. Приросты могут быть простыми или сложными, в последнем случае четко ограниченный прирост разделен на несколько субъединиц.
РЕЗУЛЬТАТЫ
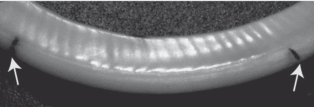
У 7 из 8 содержавшихся в виварии особей обыкновенного хомяка приросты на поверхности резцов были простыми и четкими, заметными даже невооруженным глазом при подборе соответствующего угла падения света, и только у 1 экз. они были сложными. Ширина приростов была по 500–600 мкм, но у разных особей варьировала. Число приростов на участках между метками тетрациклина соответствовало числу суток, прошедших между введениями тетрациклина (рис. 1). Общее же число приростов в нижних резцах варьировало от 46 до 56. Это показывает, что постоянно растущие резцы у обыкновенного хомяка обновляются за полтора–два месяца.
Рис. 1.
Суточные приросты на нижнем резце обыкновенного хомяка, образованные за 21 день (черные метки – положение тетрациклиновых меток от 7.04 и 28.04 при исследовании в ультрафиолетовом свете).
При исследовании поверхности резцов хомяков, которые перезимовали с термонакопителями, мы определяли участок на резце, где следует искать запись спячки, по числу суточных приростов, сформированных после последнего выхода из гипотермии.
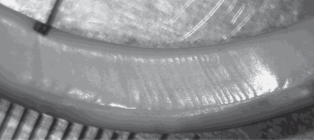
На резцах самца № 48 (последняя нормотермия с 8.02.2017, умерщвлен 7.03.2017) на фоне довольно четких суточных приростов можно было обнаружить два участка, на которых типичные суточные приросты не были заметны. Первый – против начала стачивания резца (рис. 2), и второй – ближе к середине резца. Между этими участками заметно 8–10 нечетких суточных приростов, после второго участка – 26–27 суточных приростов до конца зуба. Участок первый состоял из примерно 18 очень нечетких приростов, ширина которых во много раз меньше ширины суточных приростов. Участок второй – из 2–3 очень нечетких суточных приростов нормальной ширины, включавших множество плохо заметных узких приростов. Число суточных приростов между этими двумя участками соответствовало длительности периода нормотермии после первого эпизода гипотермии, а число приростов после второго участка – числу суток после пробуждения. Однако следует отметить, что если первый участок обнаруживается легко, то для выявления второго участка необходимо было знать, сколько суток прошло после последнего пробуждения. Кроме того, первый участок занимал значительно бóльшую часть резца, чем второй, что не соответствует длительности этих периодов спячки по термографику (см. ниже рис. 7 а). При этом также число узких приростов внутри как первого, так и второго участков не удается связать с изменениями температуры тела в периоды, которым они соответствуют.
Рис. 2.
Апикальная часть нижнего резца обыкновенного хомяка № 48. Черная точка отмечает участок, который мог быть сформирован во время спячки.

У самца № 218 (последняя нормотермия с 8.03.2018, умерщвлен 16.03.2018) на поверхности нижних резцов в апикальной и средней частях видны довольно четкие суточные приросты, хотя и неравномерные, а иногда сложные. Судя по данным терпонакопителя, последний перед гибелью эпизод нормотермии у зверька длился 8 суток. Следовательно, запись спячки на резце можно искать в базальной его части. В базальной трети после широких, не очень контрастных суточных приростов был виден участок почти без приростов, площадь которого примерно одинакова на обоих нижних резцах. На этом участке были едва заметны более узкие приросты: 8–9 на одном резце и не менее 20 на другом. Восемь суточных приростов, которые должны были бы сформироваться за 8 суток, прошедших после пробуждения, на поверхности резцов не были видны вовсе.
Самец из Дагестана (последняя нормотермия с 26.02.2018, умерщвлен 15.03.2018) прожил 16 полных суток после пробуждения. На поверхности всего резца были видны суточные приросты, местами сложные, все не очень контрастные, но хорошо заметные после зачерчивания поверхности зуба грифелем. Никакого отличающегося участка, который мог бы быть связан со спячкой, нами обнаружено не было (рис. 3).
Рис. 3.
Базальная часть нижнего резца самца обыкновенного хомяка из Дагестана. Поверхность резца зачерчена грифелем.

У самки из Дагестана с 26.02.2018 была нормотермия 8 суток, затем температура тела понизилась до 8°С и через сутки восстановилась до нормы почти на трое полных суток, после чего 15.03.2018 самка была умерщвлена. На поверхности резца были видны четкие суточные приросты на протяжении всего резца, кроме базальной его части. В базальной части, где можно было бы ожидать запись спячки, приросты были не четкие и сложные. Последние примерно 7 приростов были одинаково не четкие, но не сложные и заметные.
На резцах 18 из 19 хомяков из природной популяции суточные приросты либо были четко видны по всему резцу, либо кроме них были заметны разного размера участки с менее четкими и более сложными приростами. Два независимых мнения относительно наличия и положения на резце таких участков совпали только в половине случаев. И только у одного хомяка, труп которого был обнаружен в конце февраля, на фоне широких (по 500–600 мкм) суточных приростов средней четкости выделялись 15–18 более четких приростов (рис. 4). Этот рисунок был похож на рисунок “зоны спячки” на резцах хомяка Радде (Клевезаль и др., 2012, 2018).
ОБСУЖДЕНИЕ
То, что приросты (валики), заметные на поверхности резцов грызунов, формируются с суточной периодичностью, было ранее показано при исследовании сурка Marmota flaviventris (Rinaldi, 1999), сусликов нескольких видов (Goodwin et al., 2005; Клевезаль, Лобков, 2008) и эверсманновых хомячков (Клевезаль и др., 2015). Наши данные, полученные при содержании в неволе обыкновенных хомяков, показали, что на резцах этого вида приросты также суточные, и что эти постоянно растущие резцы обновляются за полтора–два месяца. Зная дату пробуждения от спячки (по данным термонакопителя), можно было отметить участки резца, сформированные в период спячки. На таких участках ни у одной из четырех особей мы не обнаружили характерных признаков “зоны спячки” – серии приростов, более четких и более узких, чем суточные приросты.
В соответствующих участках резцов у двух обыкновенных хомяков из г. Симферополя был нарушен рисунок суточных приростов, характерный для остальной части резца: суточные приросты не видны, поверхность участка либо без выраженного рельефа, либо со слабо угадывающимися узкими приростами. Характер этого нарушения у двух особей был разным. Еще более загадочная картина обнаружена на резцах двух особей из Дагестана. Они прожили зиму в одинаковых условиях, приближенных к естественным. У самки на участке резца, образованном в период спячки, рисунок суточных приростов был явно нарушен, аналогично тому, как это было у симферопольских хомяков. У самца11 вообще не было никакого нарушения рисунка суточных приростов. Если при этом не иметь записи динамики реальной температуры тела, можно было бы сказать, что у этого самца вообще не было эпизодов гипотермии, но на самом деле это не так. В последние месяцы жизни у него было 15 эпизодов гипотермии с понижением температуры тела до 4–5°С.
Из 19 особей из природной популяции только у одного хомяка, найденного мертвым в феврале, было на резце изменение характера приростов, похожее на запись спячки.
Следовательно, приходится признать, что на резцах обыкновенного хомяка мы не обнаружили такого изменения поверхности, которое однозначно можно было бы связать с периодом спячки. Однако такая запись была обнаружена на резцах его близких родственников – эверсманновых хомячков (Клевезаль и др., 2015) и хомяка Радде (Клевезаль и др., 2012, 2018), живших в условиях приближенных к естественным и в природе, а также у хомяка Брандта в условиях лабораторного эксперимента (Batavia et al., 2013).
Сравнение изменения температуры тела во время спячки у обыкновенного хомяка и хомяка Радде показало, что спячка у всех Cricetus cricetus протекает сходным образом, но резко отличается у таковой, обнаруженной у хомяка Радде. У обыкновенного хомяка длительность периодов гипотермии не превышала 6 суток и менялась не направленно, хаотично (рис. 5а–5г), температура тела (у исследованных нами особей) опускалась до 4–5°С. Также хаотично менялась и продолжительность периодов нормотермии, которая в большинстве случаев была не меньше суток. У хомяка Радде в Дагестане, перезимовавшего с термонакопителем (Клевезаль и др., 2018), продолжительность периодов гипотермии плавно увеличивалась от 2 до 12 суток ко второй половине спячки и потом снижалась, а температура тела опускалась до 1°С. Параллельно этому длительность периодов нормотермии уменьшалась от полуторасуток до 12 ч и затем увеличивалась до 2–3 суток (рис. 5д). Сходным образом менялась длительность периодов гипо- и нормотермии у другой особи хомяка Радде (рис. 5е), которая была перевезена из Дагестана, но которая зимовала в естественной норе в Подмосковье и погибла в конце марта во время спячки (Клевезаль и др., 2012).
Рис. 5.
Длительность эпизодов гипо- и нормотермии во время спячки у обыкновенных хомяков из г. Симферополя (а – № 48, б – № 218), Дагестана (в – самка, г – самец), хомяков Радде из Дагестана (д – по: Клевезаль и др., 2018) и Подмосковья (е – по: Клевезаль и др., 2012) и арктического суслика (ж – по: Соломонов и др., 2012).
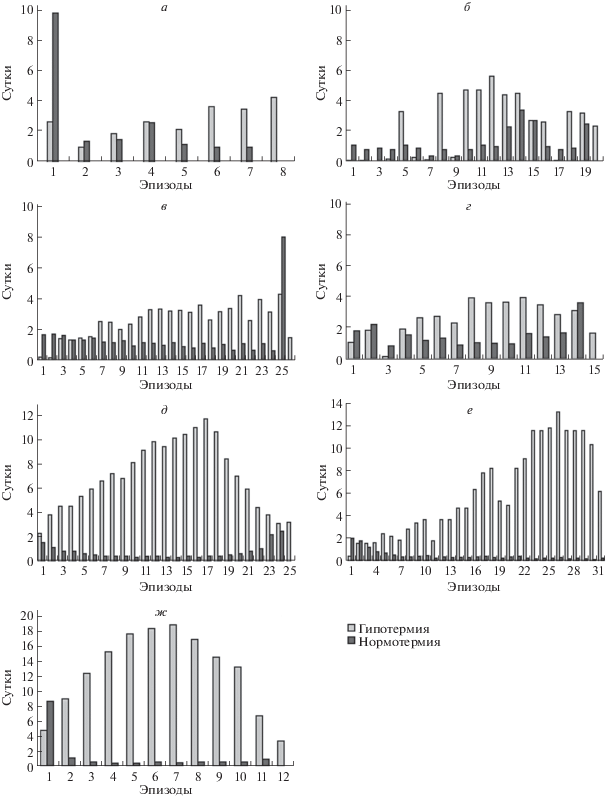
Особенно интересно сравнить изменения температуры тела во время спячки у самцов обыкновенного хомяка и хомяка Радде из Дагестана, зимовавших в сходных климатических условиях, хотя и в разные годы (рис. 5г и 5д). У хомяка Радде из 25 периодов гипотермии 20 длились более 4 суток; у самца обыкновенного хомяка из 5 периодов гипотермии только два длились 4 суток, остальные – меньше. У хомяка Радде периоды нормотермии только в 5 случаях, в начале и в конце спячки, были больше суток, тогда как у обыкновенного хомяка только в одном случае нормотермия длилась чуть меньше суток, в остальных случаях – больше. Интересно, что ход зимней спячки у хомяка Радде очень сходен с таковым у арктического суслика (сравни рис. 5д и 5ж). Тот конкретный экземпляр, чей термографик приведен на рис. 5ж, жил в искусственно созданных условиях зимовки, и у него на резце четко была видна “запись спячки” (Клевезаль, Ануфриев, 2013). Продолжительность периодов гипотермии доходила у него до 18 суток. У обыкновенного хомяка периоды гипотермии были как минимум в два раза короче, чем у хомяка Радде, и регулярной записи на резцах не оказалось вообще. Это наводит на мысль, что образование узких приростов на резце в период спячки может зависеть от длительности не только периодов нормотермии, но и периодов гипотермии, а также и от глубины гипотермии.
Близкими родственниками обыкновенных хомяков считаются не средние, а эверсманновы хомячки (род Allocricetulus) (Neumann et al., 2006, Lebedev et al., 2018). И как показывают результаты экспериментов, именно они, а конкретно – монгольские хомячки, более сходны с обыкновенным хомяком по характеру зимней спячки. У тех четырех особей монгольского хомячка (A. curtatus), которые зимовали в клетках при естественном световом и температурном режимах г. Москвы со вшитыми термонакопителями, продолжительность периодов гипотермии варьировала, но не превышала двое суток, тогда как продолжительность периодов нормотермии была очень разнообразной (Клевезаль и др., 2015). Характер их спячки, судя по данным термографиков, сходен с таковой обыкновенного хомяка своей полной неупорядоченностью изменений длительности эпизодов гипо- и нормотермии и тем, что температура тела при гипотермии не опускалась у них ниже 4–5°С. Но есть и существенные различия. У эверсманновых хомячков длительность эпизодов гипотермии была существенно меньше, а нормотермии – существенно больше, чем у обыкновенных хомяков. Возможно, именно эти различия и определили особенности в характере записи спячки на поверхности их резцов. У монгольских хомячков запись спячки, в виде серий более узких и четких приростов, была видна только в тех случаях, когда имел место ряд эпизодов с нормотермией не более полутора суток. Такие серии эпизодов нормотермии были и у обыкновенного хомяка (например, у самки из Дагестана), но четких узких приростов на резце отмечено при этом не было. Нельзя исключить, что эти различия связаны и с различиями в условиях зимовки хомячков и хомяков: клеточное содержание в первом случае и природные условия или вольерное содержание во втором.
Хотя длительность и глубина периодов гипотермии во время спячки, судя по нашим данным, может влиять на формирование узких приростов на поверхности резца, наиболее вероятно, что определяющим фактором является длительность периодов нормотермии, во время которых животное может питаться.
Хомяки Радде (Mesocricetus raddei) делают запасы, часто значительные, но в течение зимы питаются мало (или не питаются вовсе) и основную массу запасов (более 90%) используют в весенний период (апрель–май) после выхода из спячки (Яковлев, Колесников, 1954; Магомедов, Омаров, 1995; Магомедов и др., 2001). У того хомяка, на данные по которому мы ссылаемся в этой статье, была возможность питаться: в начале зимовки корм был в наличии, а после того как хомяк проснулся, корм не был обнаружен (Клевезаль и др., 2018), но не известно, когда он был съеден. За месяц до начала спячки зверек весил 325 г, а через 5 дней после пробуждения – 227 г, скорее всего, запасы были съедены в течение месяца перед погружением в спячку, а во время спячки он не питался.
Обыкновенные хомяки питаются своими запасами в зимний период во время пробуждения. Об этом свидетельствуют результаты зимних раскопок их нор, результаты наблюдений в природе (хомяки появлялись на поверхности в период спячки и потребляли корм), а также данные экспериментальных исследований (Калабухов, 1985; Wassmer, Wollnik, 1997; Wassmer, 2004; наши наблюдения в городе Симферополе в зимний период). Показано и прямое влияние доступности и состава пищи на характер спячки (Suitz, Millesi, 2017; Siutz et al., 2017). Один из исследованных нами симферопольских хомяков практически не потерял в весе за время спячки (№ 48 – 710 г до и 720 г после спячки), второй потерял в веса очень незначительно (№ 218 – 490 и 425 г соответственно), следовательно, оба питались, хотя и с разной интенсивностью. Не потеряли в весе и обыкновенные хомяки из Дагестана. Вес самки до спячки был 311 г, после пробуждения – 306 г; а вес самца – 539 и 531 г соответственно.
Таким образом, есть все основания полагать, что наличие записи спячки на поверхности резцов у одних представителей хомяков и хомячков и отсутствие у других определяется характером их питания в период спячки. Что касается использования резцов обыкновенных хомяков, добытых или найденных весной в природе для получения данных о характере протекания их зимнего периода, мы можем сказать только одно. Утверждать, что какой-то определенный хомяк впадал в зимнюю спячку, мы можем только, если на поверхности резца обнаружены приросты, выделяющиеся на фоне суточных приростов как более узкие и четкие, как у одной из 19 особей симферопольской природной популяции (рис. 4). Дополнительные исследования, оптимально с введением тетрациклина или других биомаркеров, позволят выяснить, что именно отражают другие, не похожие на запись спячки, нарушения рисунка суточных ритмов на поверхности резцов, обнаруженные у многих перезимовавших особей этого вида.
Список литературы
Калабухов Н.И., 1985. Спячка млекопитающих М.: Наука. 260 с.
Клевезаль Г.А., Ануфриев А.И., 2013. О росте резцов сусликов (род Spermophilus) в период зимней спячки // Зоологический журнал. Т. 92. № 4. С. 481–489.
Клевезаль Г.А., Лобков В.А., 2008. Суточные приросты и “зона спячки” на поверхности резцов сусликов рода Spermophilus // Зоологический журнал. Т. 87. № 12. С. 495–1503.
Клевезаль Г.А., Ушакова М.В., Чунков М.М., Феоктистова Н.Ю., Суров А.В., 2012. Запись зимней спячки на поверхности резцов хомяка Радде (Mesocricetus raddei) // Зоологический журнал. Т. 91. № 6. С. 714–720.
Клевезаль Г.А., Щепоткин Д.В., 2012. Скорость роста резцов грызунов и запись полного годового цикла в резцах серого сурка (Marmota baibacina centralis) // Зоологический журнал. Т. 91. № 1. С. 111–118.
Клевезаль Г.А., Феоктистова Н.Ю., Щепоткин Д.В., Суров А.В., 2015. Особенности записи зимней спячки на поверхности резцов хомячков рода Allocricetulus // Зоологический журнал. Т. 94. № 2. С. 259–272.
Клевезаль Г.А., Чунков М.М., Омаров К.З., Щепоткин Д.В., 2018. Запись зимней спячки на поверхности резцов хомяка Радде (Mesocricetus raddei, Rodentia, Cricetidae) из Дагестана // Зоологический журнал. Т. 97. № 5. С. 591–598
Магомедов М.-Р., Омаров К.З., 1995. Особенности питания и состояния природной популяции хомяка Радде (Mesocricetus raddei avaricus) в агроландшафтах горного Дагестана // Зоологический журнал. Т. 74. № 3. С. 123–133.
Магомедов М-Р.Д., Омаров К.З., Гасанова С.М., 2001. Трофические и территориально зависимые механизмы регуляции плотности населения хомяка Радде Mesocricetus raddei (Rodentia, Cricetidae). 2. Регуляция зимней смертности // Зоологический журнал. Т. 80. № 1. С. 90–96.
Петровский Д.В., Новиков Е.А., Мошкин М.П., 2008. Динамика температуры тела обыкновенной слепушонки (Ellobius talpinus, Rodentia, Cricetidae) в зимний период // Зоологический журнал. Т. 87. № 12. С. 1504–1508.
Соломонов Н.Г., Ануфриев А.И., Охлопков И.М., 2012. Ритмы зимней спячки арктического суслика Spermophilus parryi при температуре тела ниже нуля // Наука и образование. № 1. С. 60–64.
Суров А.В., Поплавская Н.C., Богомолов П.Л., Кропоткина М.В., Товпинец Н.Н., и др., 2015. Синурбизация обыкновенного хомяка (Cricetus cricetus L., 1758) // Российский журнал биологических инвазий. № 4. С. 105–117.
Трунова Ю.Е., 2001. Метка спячки в резцах ископаемых грызунов // Доклады Академии Наук. Т. 377. № 4. С. 568–570.
Трунова Ю.Е., Клевезаль Г.А., 1999. Межвидовые различия в регистрации зимней спячки дентином резцов грызунов // Зоологический журнал. Т. 78. № 12. С. 1455–1464.
Ушакова М.В., Феоктистова Н.Ю., Петровский Д.В., Гуреева А.В., Найденко С.В., Суров А.В., 2010. Особенности зимней спячки хомячка эверсмана (Allocricetulus eversmanni Brandt, 1859) из Саратовского Заволжья // Поволжский экологический журнал. № 4. С. 415–422.
Феоктистова Н.Ю., Найденко С.В., Суров А.В., Менчинский К.М., 2013. Эколого-физиологические особенности сезонной биологии монгольского хомячка (Allocricetulus curtatus, Allan 1940, Cricetinae, Rodentia) // Экология. № 1. С. 60–64.
Яковлев М.Г., Колесников И.М., 1954. Некоторые новые данные по распространению и экологии предкавказского хомяка в Ростовской области // Зоологический журнал. Т. 33. № 3. С. 693–700.
Batavia M., Nguyen G., Zucker I., 2013. The effects of day length, hibernation, and ambient temperature on incisor dentin in the Turkish hamster (Mesocricetus brandti) // Journal of Comparative Physiology. P. B. V. 183. P. 557–566.
Goodwin H.T., Michener G.R., Gonzalez D., Rinaldi C.E., 2005. Hibernation is recorded in lower incisors of recent and fossil ground squirrels (Spermophilus) // Journal of Mammalogy. V. 86. № 2. P. 323–332.
Goodwin H.T., Ryckman E.M., 2006. Lower incisors of prairie dogs (Cynomys) as biorecorders of hibernation and season of death // Journal of Mammalogy. V. 87. № 5. P. 1002–1012.
Feoktistova N.Yu., Surov A.V., Tovpinetz N.N., Kropotkina M.V., Bogomolov P.L. et al., 2013. The common hamster as a synurbist: a history of settlement in European cities // Zoologica Poloniae. V. 58. № 3-4. P. 116–129.
Lebedev V.S, Bannikova A.A., Neumann K., Ushakova M.V., Ivanova N.V., Surov A.V., 2018. Molecular phylogenetics and taxonomy of dwarf hamsters Cricetulus Milne-Edwards, 1867 (Cricetidae, Rodentia): description of a new genus and reinstatement of another // Zootaxa. V. 4387. № 2. P. 331–349.
Neumann K., Neumann S., Gattermann R., Michaux J., Lebedev et al., 2006. Molecular phylogeny of the Cricetinae subfamily based on the mitochondrial cytochrome b and 12S rRNA // Molecular Phylogenetics and Evolution. V. 39. P. 135–148.
Rinaldi C., 1999. A record of hibernation in the incisor teeth of recent and fossil marmots (Marmota flaviventris) // Proc. 11th International Symposium on Dental Morphology. Oulu, Finland: Oulu University Press. P. 112–119.
Suitz C., Millesi E., 2017. Torpor patterns in common hamsters with and without access to food stores // Journal of Comparative Physiology. B. V. 187. P. 881–888.
Siutz C., Nemeth M., Wagner K.-H., Quint R., Ruf T., Millesi E., 2017. Effects of food store quality on hibernation performance in common hamsters // PLoS ONE. V. 12. https://doi.org/10.1371/journal.pone.0185913
Wassmer T., 2004. Body temperature and above-ground patterns during hibernation in European hamsters (Cricetus cricetus L.) // Journal of Zoology. V. 262. P. 281–288.
Wassmer T., Wollnik F., 1997. Timing of torpor bouts during hibernation in European hamsters (Cricetus cricetus L.) // Journal of Comparative Physiology. B. V. 167. P. 270–279.
Дополнительные материалы отсутствуют.
Инструменты
Зоологический журнал