

Зоологический журнал, 2020, T. 99, № 10, стр. 1094-1109
Юг Дальнего Востока России и Корея как переходная зона между бореальной и субтропической фаунами ветвистоусых ракообразных (Crustacea, Cladocera)
П. Г. Гарибян a, *, А. Н. Неретина a, **, Н. М. Коровчинский a, ***, А. Ю. Синев a, b, ****, А. В. Чабовский a, *****, А. А. Котов a, c, ******, Н. Н. Смирнов a
a Институт проблем экологии и эволюции имени А.Н. Северцова РАН
119071 Москва, Россия
b Биологический факультет Московского государственного университета
имени М.В. Ломоносова
119991 Москва, Россия
c Казанский (Приволжский) федеральный университет
420008 Казань, Россия
* E-mail: petr.garibyan21@mail.ru
** E-mail: neretina-anna2017@yandex.ru
*** E-mail: nmkor@yandex.ru
**** E-mail: ArtemSinev@yandex.ru
***** E-mail: tiusha2@mail.ru
****** E-mail: alexey-a-kotov@yandex.ru
Поступила в редакцию 25.04.2020
После доработки 01.05.2020
Принята к публикации 11.05.2020
Аннотация
Составлен наиболее полный список видов ветвистоусых ракообразных (Crustacea, Cladocera) юга Дальнего Востока России и Корейского п-ова, насчитывающий 122 таксона, определенных до уровня вида. Для фауны юга Дальнего Востока России отмечено 92 валидных вида; для фауны Южной Кореи – 71 валидный вид. Все выявленные виды отнесены к шести группам: четырем фаунистическим комплексам (широко распространенный евроазиатский, восточноазиатско-американский, эндемичный восточно-азиатский и южный тропический), сборной группе широко распространенных неревизированных видов и видам-вселенцам. При движении с севера на юг происходит: (1) снижение видового богатства, (2) переход структуры доминирования кладоцер по встречаемости видов от ситуации с 2–3 хорошо выраженными доминантами к таковой без доминантов; (3) уменьшение доли таксонов широко распространенного евроазиатского фаунистического комплекса и увеличение доли представителей южного тропического комплекса и, соответственно, (4) смена таксоценозов бореального типа на таксоценозы субтропического типа. В бассейне наиболее крупного водоема региона – озера Ханка, – помимо этих двух таксоценозов, присутствуют еще три дополнительных уникальных таксоценоза. При этом доли таксонов эндемичного восточно-азиатского комплекса сходны в разных районах исследованного региона.
Закономерности пространственного распределения биоразнообразия изучают на примере модельных групп. Для континентальных водоемов такой модельной группой служат ветвистоусые ракообразные (Crustacea, Cladocera), обитающие в водоемах различного типа и размера и часто доминирующие как в планктонных, так и в донных и литоральных сообществах (Dumont, Negrea, 2002; Котов, 2013).
Особой задачей является изучение ветвистоусых ракообразных Дальнего Востока – обширного региона, во многом находящегося под влиянием муссонного климата. При этом наше внимание будет сосредоточено на изучении самой северной части зоны влияния муссона – юга Дальнего Востока России и Корейского п-ова с относительно сухим климатом зимой и влажным и жарким летом во время прихода муссона (Григорьева, 2009; Frederic et al., 2013; Федотов, 2015). Предыдущие исследования микроскопических ракообразных юга Дальнего Востока России (Боруцкий и др., 1952; Барабанщиков, 2000, 2002), Северного Китая (Uéno, 1937, 1939; Xiang et al., 2014; Ji et al., 2015) и Кореи (Yoon, 2010) были нередко поверхностными в плане определения таксономического статуса найденных животных. В течение последнего десятилетия сотрудниками Лаборатории экологии водных сообществ и инвазий ИПЭЭ РАН начаты систематические исследования кладоцер континентальных вод Дальнего Востока России, в том числе юга этого региона (Котов, Синев, 2011; Котов и др., 2011, 2011a; Garibian et al., 2019). Одновременно были начаты систематические исследования кладоцер отдельных районов Южной Кореи (Kotov et al., 2012; Jeong et al., 2014; Kotov et al., 2017). В итоге здесь было описано несколько новых для науки видов, отмечено проникновение в регион теплолюбивых тропических видов, а также наличие многочисленных местных эндемиков (Коровчинский, 2004; Котов, Синев, 2011; Jeong et al., 2014; Kotov, Taylor, 2019).
Исследования различных групп животных и растений свидетельствуют о специфичности обсуждаемого региона. В некоторых схемах биогеографического районирования он даже получает особый биогеографический статус (Берг, 1962). Согласно последней системе регионализации пресных вод (Abell et al., 2008), основанной преимущественно на современных данных по ихтиофауне, пресноводные экосистемы на исследуемой территории относятся к нескольким “экорегионам”. Новые находки кладоцер подтверждают мнение о специфичности фауны данного региона и демонстрируют необходимость интенсификации работ по его изучению.
Разнообразие ветвистоусых ракообразных в регионе явно недооценено, многие определения предыдущих авторов нуждаются в серьезной проверке, что делает преждевременными масштабные биогеографические обобщения (Котов, 2016). Многие области юга Дальнего Востока России и Корейского п-ова еще ждут своего исследования. За прошедшие годы здесь было собрано большое число проб и были выполнены ревизии отдельных таксонов, обитающих на Дальнем Востоке (Popova et al., 2016; Гарибян, 2017; Neretina et al., 2018). Обобщение ранее опубликованной информации по отдельным районам в сочетании с анализом эмпирических данных по другим районам может значительно повысить степень изученности фауны ветвистоусых ракообразных этого интересного в биогеографическом отношении региона, а также позволит гораздо глубже исследовать общие закономерности распределения биоразнообразия во всей зоне влияния умеренного муссонного климата в восточной части Евразии. Соответственно, целью нашей работы было обобщить данные по составу фауны ветвистоусых ракообразных юга Дальнего Востока России и Корейского п-ова и провести ее зоогеографический анализ.
МАТЕРИАЛЫ И МЕТОДЫ
Основным материалом для исследования послужили 387 проб из 170 водоемов юга Дальнего Востока России и 344 пробы из 272 водоемов Южной Кореи (рис. 1a). Большая часть проб собрана авторами в разные годы (с 2009 по 2018 гг.), остальные получены от коллег из России и Южной Кореи. Сбор материала проводили планктонными сетями и сачками различной конструкции с диаметром ячей не более 50 мкм. Почти все водоемы исследованы маршрутным методом в периоды с марта по ноябрь. Только для крупных озер Болонь и Ханка имелись серии проб, собранные в разные сезоны и годы. Пробы фиксировали преимущественно 96% этиловым спиртом, в некоторых случаях – 4% формальдегидом. Для каждой пробы были определены географические координаты.
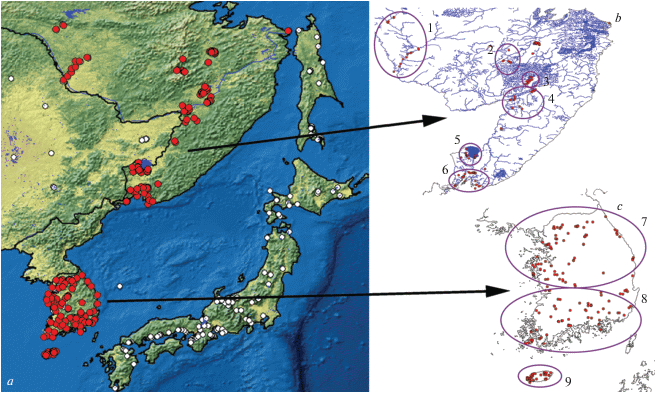
Рис. 1.
Карта исследованного региона (a), а также схемы юга Дальнего Востока РФ (b) и Республики Корея (c) с обозначенными районами взятия проб (см. табл. 1). Красные точки – водоемы, пробы из которых были проанализированы в данной работе; белые точки – пробы из других регионов, проанализированные в сравнительных целях.
Разбор проб и предварительное определение таксонов ветвистоусых ракообразных проводили в лаборатории с использованием стереоскопического микроскопа Olympus SZ51. После этого наиболее мелких, а также отдельных интересных крупных представителей ветвистоусых ракообразных подробно исследовали под микроскопом Olympus CX41. По результатам разбора проб составляли список всех таксонов кладоцер, встреченных в каждой пробе, полученные списки видов вносили в базу данных в формате Microsoft Access.
Для анализа закономерностей географического распределения таксонов по исследуемой территории условно выделили облака точек из районов, для которых имелись репрезентативные выборки, пригодные для количественного анализа (рис. 1b, 1c). Принятое подразделение на районы в целом соответствует выделению пресноводных экорегионов по Abell et al. (2008) (см. также www.feow.org). Для каждого района сбора проб определяли его среднюю широту и долготу (табл. 1). Отдельные пробы из водоемов, которые лежали вне выделенных районов, не были включены в общий анализ (хотя учитывались при составлении общего списка видов).
Таблица 1.
Соответствие выделенных районов сбора проб пресноводным экорегионам по системе Freshwater Ecoregions (www.feow.org) (см. также рис. 1)
| Номер района сбора проб | Условное название района | Номер экорегиона (название), к которому относится выделенный район по системе Freshwater Ecoregions | Средняя широта района, ° с.ш. | Средняя долгота района, ° в.д. |
|---|---|---|---|---|
| 1 | Зея | 617 (“Средний Амур”) | 52.3 | 127.6 |
| 2 | Чукчагир | 616 (“Нижний Амур”) | 52.0 | 136.7 |
| 3 | Болонь | 616 (“Нижний Амур”) | 49.9 | 136.0 |
| 4 | Хабаровск | 616 (“Нижний Амур”) | 48.6 | 135.7 |
| 5 | Ханка | 616 (“Нижний Амур”) | 45.0 | 132.5 |
| 6 | Приморье | 641 (“Сахалин, Хокайдо и побережье”) | 43.4 | 132.2 |
| 7 | Север Южной Кореи | 638 (“Западный дренаж Желтого моря”) | 36.8 | 127.1 |
| 8 | Юг Южной Кореи | 639 (“Юг Корейского п-ова”) | 35.3 | 127.8 |
| 9 | О-в Чеджу | 639 (“Юг Корейского п-ова”) | 33.1 | 126.4 |
Для оценки полноты выявления видового разнообразия кладоцер во всем регионе и его отдельных районах использовали непараметрические методы экстраполяции видового богатства (Chao et al., 2005; Colwell et al., 2004, 2012). Подготовку матриц встречаемости видов в пробах осуществляли при помощи программного модуля, разработанного А.В. Омельченко (Кабинет биоинформатики и моделирования биологических процессов ИПЭЭ РАН). Эмпирическую рандомизированную кривую накопления видов в зависимости от выборочного усилия (числа проанализированных проб) строили в пакете EstimateS 9.1. (Colwell et al., 2012). С его же помощью восстанавливали модельные кривые накопления таксонов при помощи пяти различных алгоритмов: Chao1, Chao2, Jacknife1, Jacknife2, Bootstrap. Выбирали наилучшую из пяти моделей по принципу минимума дисперсии предсказанных значений для каждого шага. Район признавали достаточно полно изученным в случае, если кривая накопления таксонов, предсказанная лучшей эмпирической моделью и эмпирическая кривая накопления видов, выходили на плато при одном и том же значении видового богатства.
Для каждого района построили диаграмму доминирования видов на основе ранжированного распределения таксонов по частоте встречаемости (для удобства представление результатов с диаграмм убраны наиболее редкие виды). Исходили из того, что вероятность обнаружить вид в водоеме (встречаемость) пропорциональна его численности и/или широте распространения и оценивали степень доминирования вида по его встречаемости. Т.е. наш индекс доминирования оценивает относительное обилие и/или широту распространения вида, причем вклад относительного обилия в оценку больше, поскольку обнаружить широко распространенный, но малочисленный вид сложнее, чем обнаружить многочисленный вид.
Все выявленные таксоны отнесли к четырем фаунистическим комплексам, предложенным Котовым (2016):
1) широко распространенный евроазиатский фаунистический комплекс (бореального происхождения) (ШЕ);
2) восточноазиатско-американский (“берингийский”) комплекс (ВА);
3) эндемичный восточно-азиатский комплекс (ЭВ);
4) южный тропический комплекс (южный теплолюбивый комплекс по: Котов, 2016, но мы считаем нужным несколько видоизменить его название, усилив его географическую составляющую и ослабив экологическую) (ЮТ).
Представители трех других фаунистических комплексов (циркумарктического, горного эндемичного и байкальского эндемичного), выделенных Котовым (2016), отсутствуют в рассматриваемом регионе. В соответствии с даными, полученными в последней работе, значительная часть видов отнесена к группе неревизованых широко распространенных видов (ШР). В качестве шестой группы (ВС) рассмотрен единственный вид-вселенец, Alona ossiani herricki Sinev 2013, найденный в Южной Корее (Kotov et al., 2017).
Для каждого района провели подсчет доли представителей каждой из шести групп как от общего числа встреченных таксонов, так и от общего числа находок всех ветвистоусых ракообразных в данном районе и построили соответствующие гистограммы распределения встречаемости представителей каждого фаунистического комплекса. На этих диаграммах районы ранжировали по их номеру, что, в целом, соответствует их расположению в направлении с севера на юг (рис. 1b, 1c).
Одновременно с этим провели регрессионный анализ зависимости встречаемости представителей разных фаунистических комплексов от широты и долготы района. В анализ были включены данные только по трем основным фаунистическим комплексам: эндемичному восточноазиатскому, широко распространенному евроазиатскому и южному тропическому как наиболее представительным. Необходимо отметить, что весь массив проб был взят не строго по широтной трансекте, а несколько диагонально по отношению к широтному и долготному направлению. Поэтому важно было оценить совместное и независимое влияние широты и долготы на распространение фаунистических комплексов. Анализ проводили в пакете Statistica 8.0.
Для выявления географических паттернов в распределении сообществ провели кластерный анализ собранных проб по их видовому составу. В анализ включали только пробы, в которых было не менее пяти видов, и таксоны, для которых имелось не менее пяти находок. Т.е. мы исключили из этого (и только этого!) анализа особо редкие виды, что является распространенной практикой при сравнительном анализе состава сообществ (Krebs, 1999), в частности, поскольку: (1) малочисленные выборки легко приводят к сильному смещению данных на выходе при небольшом смещении данных на входе; (2) редкие виды существенно увеличивают количество переменных (видов) в таблице данных, что затрудняет анализ и может вызвать существенные искажения в его результате. Также из анализа исключили пробы с малым числом видов. Следует специально отметить, что, таким образом, из анализа были исключены все временные водоемы, в которых обычно присутствует мало видов. Целью анализа было выделить кластеры проб, сходные по видовому составу (т.е. со сходными таксоценозами). Сходство проб по видовому составу оценивали по коэффициенту Жаккара (т.е. доле общих видов в двух пробах от общего числа видов в этих двух пробах) (Krebs, 1999; Boyce, Ellison, 2001; Albatineh et al., 2006). Недостатком коэффициента Жаккара (как и остальных бинарных индексов сходства) является чувствительность к неполным выборкам с большим недоучетом видов (Krebs, 1999). Однако в наших выборках недоучет был небольшим (см. ниже). Матрицу различий составили путем вычитания каждого значения коэффициента Жаккара из единицы и провели кластерный анализ методом невзвешенных средних с использованием Евклидовых дистанций. Визуализацию точек, в которых выявлены таксоценозы, относящиеся к разным кластерам, провели в программе DivaGis на основе географических координат каждого водоема.
Для основных кластеров построили диаграммы доминирования таксонов на основе их ранжированного распределения в таксоценозах по частоте встречаемости.
РЕЗУЛЬТАТЫ
Всего получено 3184 определения таксонов из 387 проб с юга Дальнего Востока России РФ и 3182 определения из 344 проб из Южной Кореи. Всего выявлено 142 таксона Cladocera (табл. 2). Среди них 21 таксон был определен лишь до уровня рода, в частности, в тех случаях, когда популяции были представлены исключительно единичными ювенильными особями, точное определение которых затруднено или невозможно (Котов, 2013). Всего для фауны юга Дальнего Востока России отмечено 92 валидных вида; 50 таксонов, определенных до вида, были найдены только здесь, для ряда видов это наиболее южный район их обитания. Для фауны Южной Кореи отмечен 71 валидный вид; 23 таксона были встречены только на территории Южной Кореи (включая остров Чеджу), для ряда видов это наиболее северный район их обитания. С продвижением с юга на север видовое богатство в пробах значимо возрастало (R2 = 0.65, F1,7 = 13.08, p = 0.009, рис. 2а), но не менялось в зависимости от долготы (R2 = = 0.14, F1,7 = 1.16, p = 0.3).
Таблица 2.
Присутствие отдельных таксонов ветвистоусых ракообразных на юге Дальнего Востока России и на прилегающих территориях: районы сбора проб
| № | Таксон | Районы | Фаунистический комплекс |
||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | |||
| 1 | Acroperus africanus Neretina et Kotov 2015 | 1 | ?ЮТ | ||||||||
| 2 | Acroperus angustatus Sars 1863 | 1 | 1 | 1 | 1 | 1 | ШЕ | ||||
| 3 | Acroperus harpae Baird 1834 | 1 | 1 | 1 | 1 | 1 | 1 | ШЕ | |||
| 4 | Acroperus sp. | 1 | 1 | ? | |||||||
| 5 | Alona affinis Leydig 1860 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | ШЕ |
| 6 | Alona cf. affinis Leydig 1860 | 1 | 1 | 1 | ? | ||||||
| 7 | Alona guttata Sars 1862 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | ШР | ||
| 8 | Alona kotovi Sinev 2012 | 1 | ЮТ | ||||||||
| 9 | Alona ossiani herricki Sinev 2013 | 1 | 1 | ВС | |||||||
| 10 | Alona quadrangularis (O.F. Müller 1776) | 1 | 1 | 1 | 1 | ШЕ | |||||
| 11 | Alona sp. | 1 | 1 | 1 | 1 | 1 | 1 | 1 | ? | ||
| 12 | Alonella excisa Fischer 1854 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | ШР |
| 13 | Alonella exigua Lilljeborg 1853 | 1 | 1 | ШР | |||||||
| 14 | Alonella nana Baird 1843 | 1 | 1 | 1 | 1 | ШР | |||||
| 15 | Alonella sp. | 1 | 1 | 1 | ? | ||||||
| 16 | Anchistropus emarginatus Sars 1862 | 1 | ШЕ | ||||||||
| 17 | Bosmina fatalis Burckhardt 1924 | 1 | 1 | 1 | 1 | ЭВ | |||||
| 18 | Bosmina longirostris O.F. Müller 1776 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | ШР |
| 19 | Bosmina longispina Leydig 1860 | 1 | ШЕ | ||||||||
| 20 | Bosmina sp. | 1 | 1 | ? | |||||||
| 21 | Bosminopsis deitersi Richard 1895 | 1 | 1 | 1 | 1 | 1 | ШР | ||||
| 22 | Bunops serricaudata Daday 1884 | 1 | ШЕ | ||||||||
| 23 | Camptocercus fennicus Stenroos 1898 | 1 | ШЕ | ||||||||
| 24 | Camptocercus lilljeborgi Schödler 1862 | 1 | 1 | ШЕ | |||||||
| 25 | Camptocercus rectirostris Schödler 1862 | 1 | ШР | ||||||||
| 26 | Camptocercus sp. | 1 | 1 | 1 | 1 | ? | |||||
| 27 | Camptocercus streletskayae Smirnov 1998 | 1 | ВА | ||||||||
| 28 | Camptocercus uncinatus Smirnov 1971 | 1 | 1 | 1 | 1 | 1 | 1 | ВА | |||
| 29 | Camptocercus vietnamensis Thanh et al. 1980 | 1 | 1 | ЮТ | |||||||
| 30 | Ceriodaphnia affinis Lilljeborg 1901 | 1 | ШР | ||||||||
| 31 | Ceriodaphnia cornuta Sars 1885 | 1 | 1 | ЮТ | |||||||
| 32 | Ceriodaphnia laticaudata P.E. Müller 1867 | 1 | ШР | ||||||||
| 33 | Ceriodaphnia megops Sars 1862 | 1 | ШР | ||||||||
| 34 | Ceriodaphnia pulchella Sars 1862 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | ШР | ||
| 35 | Ceriodaphnia quadrangula O.F. Müller 1785 | 1 | 1 | 1 | 1 | ШР | |||||
| 36 | Ceriodaphnia reticulata Jurine 1820 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | ШР | ||
| 37 | Ceriodaphnia rotunda Straus 1820 sensu Sars 1862 | 1 | ШР | ||||||||
| 38 | Ceriodaphnia sp. | 1 | 1 | 1 | 1 | 1 | 1 | ? | |||
| 39 | Chydorus irinae Smirnov et Sheveleva 2010 | 1 | 1 | 1 | ЭВ | ||||||
| 40 | Chydorus sp. | 1 | 1 | 1 | 1 | 1 | ? | ||||
| 41 | Chydorus sphaericus O.F. Müller 1776 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | ШР |
| 42 | Coronatella cf. trachystriata | 1 | ? | ||||||||
| 43 | Coronatella rectangula Sars 1862 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | ШР |
| 44 | Coronatella trachystriata Chen Shou-zhong, Zhang E et Liu Huanzhang 1994 | 1 | 1 | 1 | 1 | 1 | ЭВ | ||||
| 45 | Daphnia cristata Sars 1862 | 1 | 1 | 1 | ШЕ | ||||||
| 46 | Daphnia cucullata Sars 1862 | 1 | 1 | ШЕ | |||||||
| 47 | Daphnia curvirostris Eylmann 1887 | 1 | 1 | 1 | ШЕ | ||||||
| 48 | Daphnia galeata Sars 1864 | 1 | 1 | 1 | 1 | 1 | 1 | ШЕ | |||
| 49 | Daphnia sp. n. 1 | 1 | ЭВ | ||||||||
| 50 | Daphnia cf. longispina (O.F. Müller 1776) | 1 | ШР | ||||||||
| 51 | Daphnia sp. n. 3 | 1 | ЭВ | ||||||||
| 52 | Daphnia sp. n. 2 | 1 | ЭВ | ||||||||
| 53 | Daphnia obtusa Kurz 1874 | 1 | 1 | ШР | |||||||
| 54 | Daphnia pulex Leydig 1860 | 1 | 1 | 1 | ШР | ||||||
| 55 | Daphnia sinensis Gu, Xu, Li, Dumont et Han 2013 | 1 | 1 | 1 | 1 | 1 | 1 | ШЕ | |||
| 56 | Daphnia sinevi Kotov, Ishida et Taylor 2006 | 1 | 1 | 1 | 1 | ЭВ | |||||
| 57 | Daphnia sp. | 1 | 1 | 1 | 1 | 1 | 1 | 1 | ? | ||
| 58 | Diaphanosoma amurensis Korovchinsky et Sheveleva 2009 | 1 | 1 | 1 | 1 | ЭВ | |||||
| 59 | Diaphanosoma brachyurum Liévin 1848 s. l. | 1 | 1 | ШЕ | |||||||
| 60 | Diaphanosoma cf. brevireme Sars 1901 | 1 | ЭВ | ||||||||
| 61 | Diaphanosoma chankensis Ueno 1939 | 1 | ЭВ | ||||||||
| 62 | Diaphanosoma dubium Manujlova 1964 | 1 | 1 | 1 | 1 | 1 | 1 | ЭВ | |||
| 63 | Diaphanosoma macrophtalma Korovchinsky et Mirabdullaev 1995 | 1 | 1 | 1 | 1 | 1 | 1 | ЭВ | |||
| 64 | Diaphanosoma orghidani Negrea 1982 | 1 | 1 | ШЕ | |||||||
| 65 | Diaphanosoma pseudodubium Korovchinsky 2000 | 1 | 1 | ЭВ | |||||||
| 66 | Diaphanosoma sp. | 1 | 1 | 1 | 1 | 1 | 1 | ? | |||
| 67 | Diaphanosoma transamurensis Korovchinsky 1986 | 1 | 1 | 1 | 1 | 1 | ЭВ | ||||
| 68 | Disparalona chappuisi Brehm 1934 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | ЮТ | ||
| 69 | Disparalona ikarus Kotov et Sinev 2011 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | ЭВ | |
| 70 | Disparalona sp. | 1 | 1 | 1 | ? | ||||||
| 71 | Dunhevedia crassa King 1853 | 1 | ЮТ | ||||||||
| 72 | Ephemeroporus barroisi Richard 1894 | 1 | ЮТ | ||||||||
| 73 | Eurycercus macracanthus Frey 1973 | 1 | 1 | 1 | 1 | 1 | 1 | ?ВА | |||
| 74 | Flavalona costata Sars 1862 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | ШР |
| 75 | Flavalona rustica (Scott 1895) | 1 | ШР | ||||||||
| 76 | Graptoleberis testudinaria Fischer 1848 | 1 | 1 | 1 | 1 | 1 | ШР | ||||
| 77 | Holopedium gibberum Zaddach 1855 | 1 | 1 | ШЕ | |||||||
| 78 | Ilyocryptus cuneatus Štifter 1988 | 1 | 1 | 1 | 1 | ШЕ | |||||
| 79 | Ilyocryptus cf. raridentatus Smirnov 1989 | 1 | 1 | 1 | 1 | ЮТ | |||||
| 80 | Ilyocryptus sp. | 1 | 1 | 1 | ? | ||||||
| 81 | Ilyocryptus spinifer Herrick 1882 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | ЮТ | ||
| 82 | Ilyocryptus yooni Jeong, Kotov et Lee 2012 | 1 | 1 | 1 | 1 | 1 | ЭВ | ||||
| 83 | Korealona karanovici Jeong, Sinev, Brancelj, Chang et Kotov 2017 | 1 | ЭВ | ||||||||
| 84 | Kurzia latissima Kurz 1875 | 1 | 1 | 1 | ШЕ | ||||||
| 85 | Kurzia longirostris Daday 1898 | 1 | ЮТ | ||||||||
| 86 | Lathonura rectirostris O.F. Müller 1776 | 1 | 1 | 1 | ШР | ||||||
| 87 | Leptodora kindtii Focke 1844 | 1 | 1 | 1 | ШЕ | ||||||
| 88 | Leptodora richardi Korovchinsky 2009 | 1 | ЭВ | ||||||||
| 89 | Leptodora sp. | 1 | 1 | ? | |||||||
| 90 | Leydigia acanthocercoides Fischer 1854 | 1 | ШЕ | ||||||||
| 91 | Leydigia ciliata Gauthier 1939 | 1 | ЮТ | ||||||||
| 92 | Leydigia leydigi Schödler 1863 | 1 | ШЕ | ||||||||
| 93 | Leydigia louisi Kotov, Elías-Gutiérrez et Nieto 2003 | 1 | ЮТ | ||||||||
| 94 | Leydigia sp. | 1 | ? | ||||||||
| 95 | Limnosida frontosa Sars 1862 | 1 | 1 | 1 | ШЕ | ||||||
| 96 | Macrothrix laticornis Jurine 1820 | 1 | ШЕ | ||||||||
| 97 | Macrothrix pennigera Shen Chia-Jui, Sung Ta-hsiang et Chen Kou-hsiao 1961 | 1 | ЭВ | ||||||||
| 98 | Macrothrix rosea Jurine 1820 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | ШЕ | ||
| 99 | Macrothrix sp. | 1 | 1 | 1 | 1 | ? | |||||
| 100 | Macrothrix vietnamensis Silva-Briano, Dieu et Dumont 1999 | 1 | 1 | ЮТ | |||||||
| 101 | Moina affinis Birge 1893 | 1 | 1 | ШР | |||||||
| 102 | Moina belli Gurney 1904 | 1 | 1 | ШР | |||||||
| 103 | Moina brachiata Jurine 1820 | 1 | ШЕ | ||||||||
| 104 | Moina lipini Smirnov 1976 | 1 | 1 | ШЕ | |||||||
| 105 | Moina macrocopa Straus 1820 | 1 | 1 | 1 | ШЕ | ||||||
| 106 | Moina micrura Kurz 1875 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | ШР | ||
| 107 | Moina sp. | 1 | 1 | 1 | 1 | 1 | ? | ||||
| 108 | Moina weismanni Ishikawa 1896 | 1 | 1 | 1 | 1 | ШЕ | |||||
| 109 | Monospilus daedalus Kotov et Sinev 2011 | 1 | 1 | ЭВ | |||||||
| 110 | Monospilus sp. | 1 | 1 | 1 | ? | ||||||
| 111 | Nedorchynchotalona chiangi Kotov et Sinev 2011 | 1 | 1 | 1 | ЭВ | ||||||
| 112 | Nicsmirnovius eximius Kiser 1948 | 1 | ЮТ | ||||||||
| 113 | Ophryoxus gracilis Sars 1862 | 1 | ШЕ | ||||||||
| 114 | Ophryoxus zini Sars 1862 | 1 | ЭВ | ||||||||
| 115 | Oxyurella tenuicaudis Sars 1862 | 1 | 1 | ШЕ | |||||||
| 116 | Picripleuroxus denticulatus Birge 1879 | 1 | 1 | 1 | 1 | ?ЮТ | |||||
| 117 | Picripleuroxus laevis Sars 1862 | 1 | 1 | 1 | 1 | ШЕ | |||||
| 118 | Picripleuroxus quasidenticulatus Smirnov 1996 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | ЮТ | ||
| 119 | Picripleuroxus sp. | 1 | 1 | 1 | ? | ||||||
| 120 | Picripleuroxus striatus Schödler 1862 | 1 | 1 | 1 | 1 | ШЕ | |||||
| 121 | Pleuroxus aduncus Jurine 1820 | 1 | 1 | 1 | 1 | 1 | 1 | ШР | |||
| 122 | Pleuroxus jejuensis Jeong, Kotov et Lee 2013 | 1 | ЭВ | ||||||||
| 123 | Pleuroxus sp. | 1 | 1 | 1 | ? | ||||||
| 124 | Pleuroxus truncatus O.F. Müller 1785 | 1 | 1 | 1 | ШЕ | ||||||
| 125 | Pleuroxus сf. trigonellus O.F. Müller 1776 | 1 | 1 | ВА | |||||||
| 126 | Polyphemus pediculus Linnaeus 1761 | 1 | 1 | 1 | 1 | 1 | ШР | ||||
| 127 | Pseudochydorus globosus Baird 1843 | 1 | 1 | 1 | 1 | 1 | 1 | ШЕ | |||
| 128 | Pseudosida szalayi Daday 1898 | 1 | 1 | ЮТ | |||||||
| 129 | Scapholeberis cf. kingii Sars 1903 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | ЮТ | |
| 130 | Scapholeberis mucronata O.F. Müller 1776 | 1 | 1 | 1 | 1 | 1 | 1 | ШЕ | |||
| 131 | Scapholeberis sp. | 1 | 1 | 1 | 1 | 1 | 1 | ? | |||
| 132 | Sida crystallina (O.F. Müller 1776) | 1 | 1 | 1 | 1 | 1 | 1 | 1 | ШЕ | ||
| 133 | Sida ortiva Korovchinsky 1979 | 1 | 1 | 1 | 1 | 1 | 1 | ВА | |||
| 134 | Sida sp. | 1 | 1 | 1 | 1 | 1 | ? | ||||
| 135 | Simocephalus congener Koch 1841 | 1 | 1 | 1 | 1 | ШР | |||||
| 136 | Simocephalus exspinosus De Geer 1778 | 1 | 1 | 1 | 1 | 1 | ШР | ||||
| 137 | Simocephalus mixtus Sars 1903 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | ШР | |
| 138 | Simocephalus serrulatus Koch 1841 | 1 | 1 | 1 | 1 | 1 | 1 | ШР | |||
| 139 | Simocephalus sp. | 1 | 1 | 1 | 1 | 1 | 1 | 1 | ? | ||
| 140 | Simocephalus vetuloides Sars 1899 | 1 | ВА | ||||||||
| 141 | Simocephalus vetulus (O.F. Müller 1776) | 1 | 1 | 1 | 1 | 1 | ШР | ||||
| 142 | Streblocerus serricaudatus Fischer 1849 | 1 | ШР | ||||||||
Примечания. 1 – р. Зея, 2 – Чукчагир, 3 – оз. Болонь, 4 – Хабаровск, 5 – оз. Ханка, 6 – юг Приморского края, 7 – северная часть Южной Кореи, 8 – южная часть Южной Кореи, 9 – О-в Чеджу. Фаунистические комплексы: ШЕ – широко распространённый евроазиатский, ЭВ – эндемичный восточноазиатский, ВА – восточноазиатский-американский, ЮТ – южный тропический, ШР – неревизованые широко распространенные виды, ВС – виды-вселенцы.
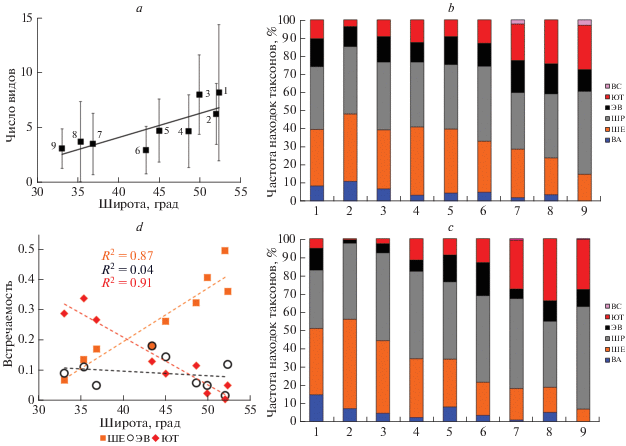
Рис. 2.
Результаты фаунистического анализа ветвистоусых ракообразных региона: a – изменение среднего (± ст. откл.) числа видов с широтой (цифрами обозначены номера районов сбора проб – см. табл. 1; статистика – см. табл. 4); b – частота встречаемости таксонов различных фаунистичеcких комплексов; c – частота встречаемости находок таксонов различных фаунистических комплексов; d – встречаемость представителей южного тропического (ЮТ), широко распространённого евроазиатского (ШЕ) и эндемичного восточно-азиатского (ЭВ) фаунистических комплексов в зависимости от средней широты района сбора проб (см. статистику в табл. 4). Для b–c по осям абсцисс – номера районов, соответствующие направлению с севера на юг (см. табл. 1).
Анализ полноты выявления таксонов в регионе в целом и в его отдельных районах показал, что почти во всех случаях модель Chao1 оказывалась наилучшей. Эмпирическая рандомизированная кривая накопления видов и модельная кривая (Chao1) для всех определений из всех проб с юга Дальнего Востока России, включая таксоны, определенные только до рода, но без учета “пустых” проб, близки к выходу на плато, при этом итоговое разнообразие оценивается моделью как 168 видов при обнаруженных 143 таксонах (из которых только 122 – валидные виды). Из следующего анализа исключили определения до уровня рода, что значительно улучшило сходность модельной и эмпирической кривых. При этом только для районов “Ханка” (5) и “Южная часть Южной Кореи” (8) можно говорить о полноте выявления видового состава ветвистоусых ракообразных, для прочих районов выявлен серьезный недоучет видов (табл. 3).
Таблица 3.
Непараметрическая оценка видового богатства отдельных районов
| Номер района | Название района | Эмпирическое число видов | Лучшая модель | Предсказанное число видов |
|---|---|---|---|---|
| 1 | Зея | 61 | Chao 1 | 74 |
| 2 | Чукчагир | 28 | Chao 1 | 34 |
| 3 | Болонь | 46 | Chao 1 | 55 |
| 4 | Хабаровск | 77 | Chao 1 | 95 |
| 5 | Ханка | 93 | Chao 1 | 101 |
| 6 | юг Приморского края | 63 | Bootstrap | 76 |
| 7 | северная часть Южной Кореи | 62 | Chao 1 | 73 |
| 8 | южная часть Южной Кореи | 69 | Chao 1 | 72 |
| 9 | О-в Чеджу | 43 | Chao 1 | 59 |
При движении с севера на юг наблюдается переход структуры доминирования кладоцер по встречаемости видов от ситуации с 2–3 хорошо выраженными доминантами к таковой без выраженных доминантов (рис. 3). Предварительный анализ наших данных показывает, что, чем южнее расположен район взятия проб, тем больше в нем отмечается представителей южного тропического комплекса (ЮТ) и тем меньше – представителей широко распространенного евроазиатского комплекса (ШЕ), при этом доля представителей эндемичного восточноазиатского комплекса (ЭВ) остается приблизительно одинаковой (10–20%) (рис. 2b). Тенденция к увеличению частоты присутствия южных видов и уменьшения доли северных видов также хорошо заметна и для общего числа находок представителей каждого фаунистического комплекса в каждом районе (рис. 2c). Эти тенденции подтверждаются результатами регрессионного анализа (рис. 2d, табл. 4): с увеличением широты (т.е. при движении с юга на север) значимо снижается доля представителей южного тропического комплекса (ЮТ) и увеличивается участие представителей широко распространенного евроазиатского комплекса (ШЕ), тогда как представленность эндемичного восточноазиатского комплекса (ЭВ) остается неизменной. При этом долгота на встречаемость представителей этих комплексов значимо не влияет (табл. 4).
Рис. 3.
Частота встречаемости отдельных таксонов в более северных районах (1, 5 на юге Дальнего Востока РФ) и в более южных районах (7 и 9 в Южной Корее). Черным цветом выделены доминирующие (по встречаемости) виды.
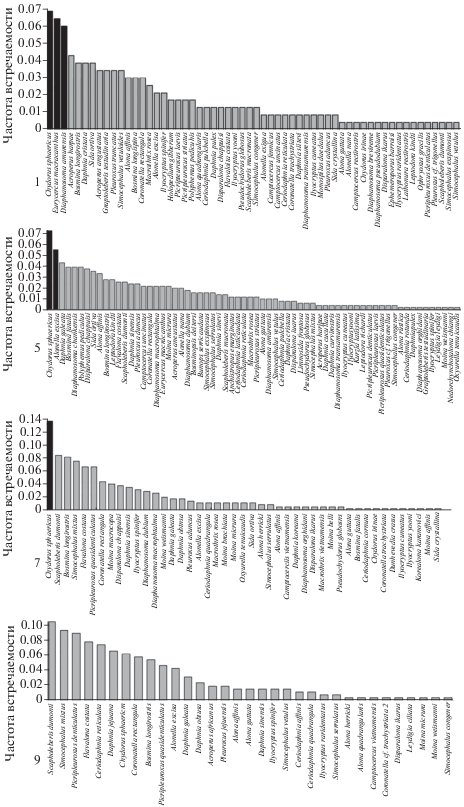
Таблица 4.
Влияние широты и долготы на обилие видов, а также встречаемость эндемичного восточноазиатского (ЭВ), широко распространенного евроазиатского (ШЕ) и южного тропического (ЮТ) комплексов по отдельности
| Эффект | Параметры | |||
|---|---|---|---|---|
| среднее число видов (F1,6, p) | доля ШЕ (F1,6, p) | доля ЭВ (F1,6, p) | доля ЮТ (F1,6, p) | |
| Долгота | 1.6, 0.3 | 1.0, 0.4 | 0.5, 0.5 | 0.9, 0.4 |
| Широта | 12.8, 0.01 | 18.0, 0.005 | 0.01, 0.9 | 28.3, 0.002 |
Кластерный анализ разделяет весь пул проб (и таксоценозов) на два больших (А, B) и три малых (C, D, E) кластера (рис. 4а). Таксоценозы наиболее обширного кластера А распространены преимущественно в более северных районах и полностью отсутствуют на юге Корейского п-ова (рис. 4в) (к сожалению, все пробы с о-ва Чеджу были бедны видами и не попали в наш анализ). Наоборот, таксоценозы кластера B большей частью встречаются в Корее, но редки на юге Дальнего Востока. При этом в таксоценозах кластера А преобладают виды в основном из числа широко распространенных палеарктических видов (ШЕ), а в таксоценозах кластера B – южные тропические виды (ЮТ) (рис. 5).
Рис. 4.
Результаты кластеризации проб (метод невзвешенных средних) по сходству видового состава проб (a) и их распределение по исследованному региону (b). Белые точки – местообитания, таксоценозы которых не попали в анализ вследствие их малочисленности (менее 5 видов Cladocera на водоем). A–E – основные кластеры таксоценозов.
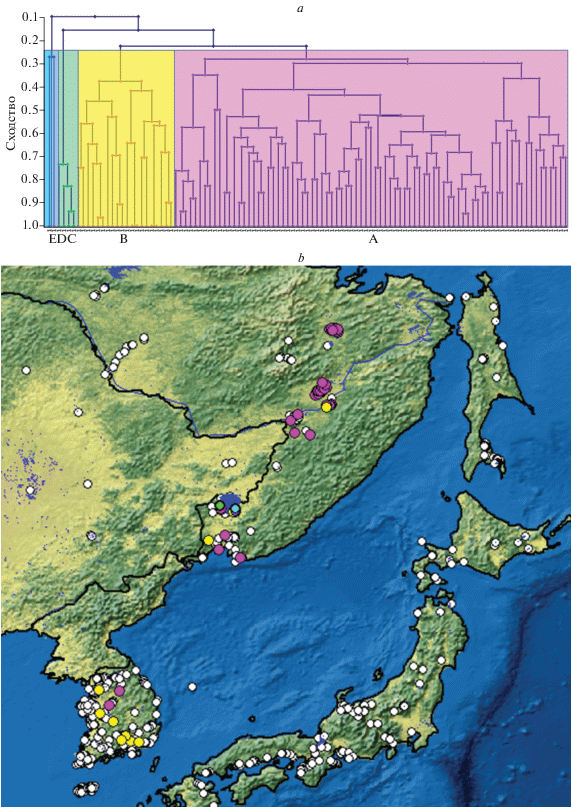
Рис. 5.
Частота находок отдельных таксонов в таксоценозах кластеров А (a) и B (b). На верхнем графике оранжевым цветом помечены таксоны широко распространённого евроазиатского (ШЕ), на нижнем графике красным цветом помечены таксоны южного тропического комплекса (ЮТ).
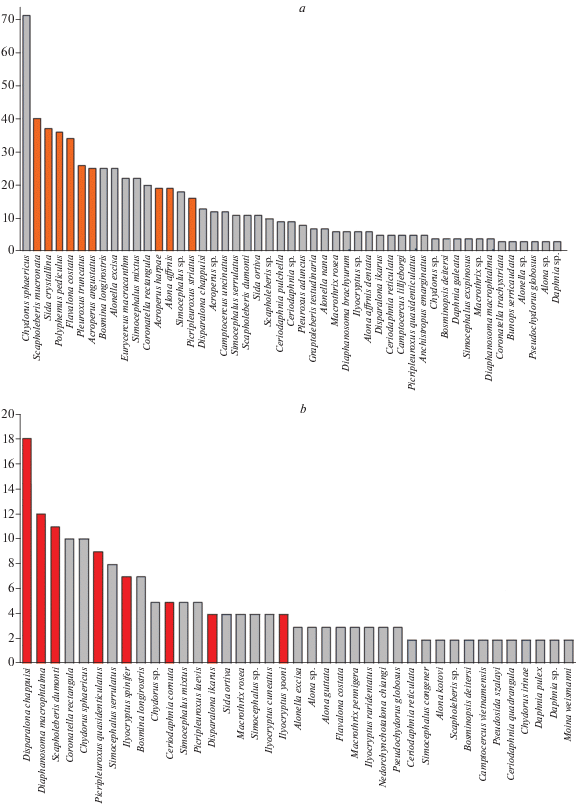
Наконец, таксоценозы из кластеров C, D и E найдены в единственном районе, бассейне озера Ханка, где, при этом, встречаются и таксоценозы типов А и B. Т.е. разнообразие таксоценозов в бассейне оз. Ханка превосходит суммарное разнообразие таксоценозов всех остальных водоемов.
ОБСУЖДЕНИЕ
Фауна ветвистоусых ракообразных обсуждаемого региона ныне исследована гораздо более полно по сравнению с предыдущими работами, что позволило сделать некоторые обобщения, касающиеся вопросов биогеографии, которые ранее были невозможны. Отдельные эндемичных виды из данного региона регистрировались и ранее (Uéno, 1937, 1939; Мануйлова, 1964; Смирнов, Шевелева, 2010), однако их находки носили по большей части бессистемный характер. Котов (2016) выделил их в особый фаунистический комплекс и предположил их широкое распространение в регионе. В частности, было продемонстрировано, что эндемичные таксоны обильны в бассейнах р. Зея и оз. Болонь (Котов, 2016; Garibian et al., 2019). Важным результатом нашей работы стало обнаружение таксонов эндемичного восточно-азиатского фаунистического комплекса на значительно большей территории – во всех изученных районах региона. Более того, доля их встречаемости сходна в разных районах. Таким образом, все изученные районы лежат в пределах зоны эндемизма ветвистоусых ракообразных, существование которой было предсказано Коровчинским (2004; Korovchinsky, 2006). Точные границы этой зоны неизвестны, к настоящему времени лишь понятно, что выявленные эндемики отсутствуют в Якутии, Монголии и южном Китае. Данный регион одновременно является зоной эндемизма многих других пресноводных беспозвоночных (Старобогатов, 1970; Timm, 1997; Тиунова, 2007; Тесленко, 2012; Lopes-Lima et al., 2020) и рыб (Никольский, 1956; Новомодный и др., 2004).
Хотя непараметрическая оценка биоразнообразия и указала на явный недоучет видового состава ряда выделенных районов, на данный момент наш список кладоцер – наиболее полный для региона. При этом понятно, что в наших рассуждениях не следует концентрировать внимание на различиях между отдельными районами, поскольку зачастую они мнимые и могут быть объяснены неполнотой сравниваемых списков. Однако подобное обстоятельство не может помешать нам проанализировать состав фауны кладоцер в целом и общие тенденции ее изменения при движении с севера на юг статистическими методами, которые по определению работают с неполными выборками.
На трансекте длиной около 2500 км (от нижнего течения Амура (52° с.ш.) до корейского о-ва Чеджу (33° с.ш.)) наблюдаются ярко выраженные изменения состава и структуры фауны Cladocera. При движении с севера на юг сокращается доля таксонов широко распространенного евроазиатского фаунистического комплекса, для ряда таксонов которого методами филогеографии продемонстрировано бореальное происхождение (Xu et al., 2010; Kotov, Taylor, 2019). Одновременно происходит увеличение доли представителей южного тропического комплекса, для которого предполагается тропическое происхождение, хотя прямых филогеографических доказательств этого к настоящему времени не получено (Kotov et al., 2012; Гарибян, 2017). Помимо смены фаунистических комплексов, происходят изменения в структуре таксономического разнообразия: видовое богатство снижается, а структура доминирования видов по встречаемости становится более равномерной – в южных таксоценозах, в отличие от северных, нет ярко выраженных доминантов. “Скошенное” распределение видов по обилию с несколькими доминантами характерно для фауны умеренных широт, более равномерное без очевидных доминантов – для фаун тропических широт (Ричардс, 1961; Dumont, 1994; Dumont, Segers, 1996). Широтная смена происходит и с таксоценозами: таксоценозы типа A c преобладанием видов бореального генезиса сменяются при движении на юг на таксоценозы типа B с преобладанием видов тропического генезиса, что также подтверждает переходный статус исследованного региона.
При этом бассейн оз. Ханка отличается большим разнообразием таксоценозов, только здесь встречаются редкие таксоценозы типов C, D и E. По-видимому, это связано с необычайно высоким разнообразием биотопов в бассейне этого наибольшего по площади водоема региона, но данное предположение нуждается в проверке.
Исследованный регион на разных схемах биогеографического районирования принадлежит к разным зонам. Уоллес (Wallace, 1876), проанализировавший распределение различных представителей наземной фауны и флоры, целиком относил его к Палеарктической зоне. Старобогатов (1970), исследовавший пресноводных моллюсков, напротив, считал регион частью Ориентальной зоны (“Сино-Индийской области”). По Бергу (1962), оперировавшему ихтиологическими данными, по рассматриваемому региону проходит граница между двумя основными областями: Голарктической на севере и Амурской (Маньчжурской) на юге. Полученные результаты наиболее хорошо соотносятся с последней схемой биогеографического районирования: регион является переходной зоной между бореальной и субтропической фаунами, что продемонстрировано здесь на примере ветвистоусых ракообразных.
Список литературы
Барабанщиков Е.И., 2000. Качественный состав зоопланктона озера Ханка // Геохимические и эколого-биогеохимические исследования в Приамурье. № 10. С. 130–139.
Барабанщиков Е.И., 2002. Данные по фауне веслоногих ракообразных (Copepoda) бассейнов озера Ханка, реки Сунгачи и реки Уссури // Материалы Всероссийской Интернет-конференции молодых ученых. Владивосток: Изд. ТИНРО-центра. С. 92–96.
Боруцкий Е.В., Ключарева О.А., Никольский Г.В., 1952. Донные беспозвоночные (зообентос) Амура и их роль в питании амурских рыб // Труды Амурской ихтиологической экспедиции. Т. 3. С. 5–39.
Берг Л.С., 1962. Разделение территории Палеарктики и Амурской области на зоогеографические области на основании распространения пресноводных рыб. Избранные труды. М.: Изд. АН СССР. Т. 5. С. 320–360.
Гарибян П.Г., 2017. Nicsmirnovius eximius (Kiser, 1948) (Cladocera: Chydoridae) из Приморского края: первая находка рода для фауны России // Зоологический журнал. Т. 96. № 11. С. 1359–1363.
Григорьева Е.А., 2009. Динамика временных границ вегетационного периода в южной части Дальнего Востока // Региональные проблемы. № 12. С. 29–33.
Коровчинский Н.М., 2004. Ветвистоусые ракообразные отряда Ctenopoda мировой фауны (морфология, систематика, экология, зоогеография). М.: Товарищество научных изданий КМК. 410 с.
Котов А.А., 2013. Морфология и филогения Anomopoda (Crustacea: Cladocera). М.: Товарищество научных изданий КМК. 638 с.
Котов А.А., 2016. Фаунистические комплексы Cladocera (Crustacea, Branchiopoda) Восточной Сибири и Дальнего Востока России // Зоологический журнал. Т. 95. № 7. С. 748–768.
Котов A.A., Коровчинский Н.М., Синев А.Ю., Смирнов Н.Н., 2011. Cladocera (Crustacea: Branchiopoda) бассейна реки Зеи (Амурская Область, Российская Федерация). 3. Систематико-фаунистический и зоогеографический анализ // Зоологический журнал. Т. 90. № 4. С. 402–411.
Котов A.A., Синев А.Ю., 2011. Cladocera (Crustacea, Branchiopoda) бассейна реки Зеи (Амурская Область, Российская Федерация). 2. Описания новых таксонов // Зоологический журнал. Т. 90. № 3. С. 272–284.
Котов A.A., Синев А.Ю., Коровчинский Н.М., Смирнов Н.Н., Беккер Е.И., Шевелева Н.Г., 2011а. Cladocera (Crustacea, Branchiopoda) бассейна реки Зеи (Амурская Область, Российская Федерация). 1. Новые таксоны для фауны России // Зоологический журнал. Т. 90. № 2. С. 131–142.
Мануйлова E.Ф., 1964. Ветвистоусые рачки фауны СССР // Определители по фауне СССР, издаваемые Зоологическим институтом Академии наук СССР. Т. 88. С. 1–327 с.
Никольский Г.В., 1956. Рыбы бассейна Амура // Итоги Амурской ихтиологической экспедиции 1945–1949. М.: Изд. Академии Наук СCCР. 551 с.
Новомодный Г.В., Золотухин С.Ф., Шаров П.О., 2004. Рыбы Амура: богатство и кризис. М.: Апельсин. 30 с.
Ричардс П.У., 1961. Тропический дождевой лес. М.: Иностранная литература. 448 с.
Смирнов Н.Н., Шевелева Н.Г., 2010. Chydorus irinae sp. n. (Anomopoda, Chydoridae, Chydorinae) из р. Томь (Бассейн Амура, Россия) // Зоологический журнал. Т. 89. С. 635–638.
Старобогатов Я.И., 1970. Фауна моллюсков и зоогеографическое районирование континентальных водоемов земного шара. Л.: Наука. 372 с.
Тесленко В.А., 2012. Фауна веснянок (Insecta, Plecoptera) Еврейской автономной области // Евроазиатский энтомологический журнал. Т. 11. С. 29–36.
Тиунова Т.М., 2007. Современное состояние изученности поденок (Ephemeroptera) Дальнего Востока России и сопредельных территорий // Евроазиатский энтомологический журнал. Т. 6. С. 181–194.
Федотов В.И., 2015. География России. Почвенно-климатические условия, сельское хозяйство и отрасли пищевой и легкой промышлености // Вестник Воронежского государственного университета. № 3. С. 86–111.
Abell R., Thieme M.L., Revenga C., Bryer M., Kottelat M., Bogutskaya N., Coad B., Mandrak N., Balderas S.C., Bussing W., Stiassny M.L. J., Skelton P., Allen G.R., Unmack P., Naseka A., Rebecca N., Sindorf N., Robertson J., Armijo E., Higgins J.V., Heibel T.J., Wikramanayake E., Olson D., Lópes H.L., Reis R E., Lundberg J.G., Sabaj Péres M.H., Petry P., 2008. Freshwater ecoregions of the world: a new map of biogeographic units for freshwater biodiversity conservation // BioScience. V. 58. P. 403–414.
Albatineh A.N., Niewiadomska-Bugaj M., Mihalko D., 2006. On similarity indices and correction for chance agreement // Journal of Classification. V. 23. P. 301–313.
Boyce R.L., Ellison P.C., 2001. Choosing the best similarity index when performing fuzzy set ordination on binary data // Journal of Vegetation Science. V. 12. P. 711–720.
Chao A., Chazdon R.L., Colwell R.K., Shen T.J., 2005. A new statistical approach for assessing compositional similarity based on incidence and abundance data // Ecology Letters. V. 8. P. 148–159.
Colwell R.K., Chao A., Gotelli N.J., Lin S.Y., Mao C.X., Chazdon R.L., Longino J.T., 2012. Models and estimators linking individual-based and sample-based rarefaction, extrapolation, and comparison of assemblages // Journal of Plant Ecology. V. 5. P. 3–21.
Colwell R.K., Mao C.X., Chang J., 2004. Interpolating, extrapolating, and comparing incidence-based species accumulation curves // Ecology. V. 85. P. 2717–2727.
Dumont H.J., 1994. On the diversity of the Cladocera in the tropics // Hydrobiologia. V. 272. P. 27–38.
Dumont H.J., Negrea S.V., 2002. Introduction to the class Branchiopoda // Guides to the identification of the microinvertebrates of the continental waters of the world. Dumont H.J. (ed.). V. 19. Leiden: Backhuys Publishers. 398 p.
Dumont H.J., Segers H., 1996. Estimating lacustrine zooplankton species richness and complementarity // Hydrobiologia. V. 341. P. 125–132.
Frédéric J.M.B., Su T., Huang Y.J., Wang L., Zhou Z.K., 2013. A global-scale test for monsoon indices used in paleoclimatic reconstruction // Paleoworld. V. 22. P. 93–100.
Garibian P.G., Chertoprud E.S., Sinev A.Yu., Korovchinsky N.M., Kotov A.A., 2019. Cladocera and Copepoda (Crustacea: Branchiopoda) of the Lake Bolon and its basin (Far East of Russia) // Arthropoda Selecta. V. 28. № 1. P. 37–63.
Jeong H.G., Kotov A.A., Lee W., 2014. Checklist of the freshwater Cladocera (Crustacea: Branchiopoda) of South Korea // Proceedings of the Biological Society of Washington. V. 127. P. 216–228.
Jeong H.G., Kotov A.A., Lee W., Jeong R., Cheon S., 2015. Diversity of freshwater Cladoceran species (Crustacea: Branchiopoda) in South Korea // Journal of Ecology and Environment. V. 38. P. 361–366.
Ji G.H., Xiang X.F., Chen S.Z., Yu G.L., Kotov A.A., Dumont H.J., 2015. Annotated checklist of Chinese Cladocera (Crustacea: Branchiopoda). Part II. Order Anomopoda (families Macrothricidae, Eurycercidae and Chydoridae) // Zootaxa. V. 4044. P. 241–269.
Korovchinsky N.M., 2006. The Cladocera (Crustacea: Branchiopoda) as a relict group // Zoological Journal of the Linnean Society. V. 147. P. 109–124.
Kotov A.A., Jeong H.J., Lee W., 2012. Cladocera (Crustacea: Branchiopoda) of the south-east of the Korean Peninsula, with twenty new records for Korea // Zootaxa. V. 3368. P. 50–90.
Kotov A.A., Sinev A.Y., Garibian P.G., Neretina A.N., Jeong H.G., Lee W., Min G.S., 2017. Recent progress in studies of the Cladocera (Crustacea: Branchiopoda) of South Korea, with seven new records for the Korean Peninsula // Journal of Species Research. V. 6. P. 227–246.
Kotov A.A., Taylor D.J., 2019. Contrasting endemism in pond-dwelling cyclic parthenogens: the Daphnia curvirostris species group (Crustacea: Cladocera) // Scientific Reports. V. 9. № 6812. https://doi.org/10.1038/s41598-019-43281-9
Krebs C.J., 1999. Ecological Methodology. 2nd Edition. Menlo Park: Benjamin Cummings. 620 p.
Lopes-Lima M., Hattori A., Kondo T., Lee J.H., Kim S.K., Shirai A. et al., 2020. Freshwater mussels (Bivalvia: Unionidae) from the rising sun (Far East Asia): phylogeny, systematics, and distribution // Molecular Phylogenetics and Evolution. V. 146. № 106 755.
Neretina A.N., Garibian P.G., Sinev A.Y., Kotov A.A., 2018. Diversity of the subgenus Disparalona (Mixopleuroxus) Hudec, 2010 (Crustacea: Cladocera) in the New and Old World // Journal of Natural History. V. 52. P. 155–205.
Popova E.V., Petrusek A., Kořinek V., Mergeay J., Bekker E.I., Karabanov D.P., Galimov Y.R., Neretina T.V., Taylor D.J., Kotov A.A., 2016. Revision of the Old World Daphnia (Ctenodaphnia) similis group (Cladocera: Daphniidae) // Zootaxa. V. 4161. P. 1–40.
Timm T., 1997. Freshwater Oligochaeta of some urban water-courses in the Russian Far East // International Revue der gesamten Hydrobiologie. V. 82. P. 437–467.
Uéno M., 1937. Cladocera of Manchoukuo // Internationale Revue der gesamten Hydrobiologie und Hydrographie. Bd. 35. P. 199–216.
Uéno M., 1939. Manchurian fresh-water Cladocera // Annotationes Zoologicae Japonenses. V. 18. P. 219–231.
Wallace A.R., 1876. The geographical distribution of animals. In 2 volumes. New York: Harper. 553 p.
Xiang X.F., Ji G.H., Chen S.Z., Yu G.L., Xu L., Han B.P., Kotov A.A., Dumont H.J., 2014. Check-List of Chinese Cladocera (Crustacea: Branchiopoda). Part 1. Haplopoda, Ctenopoda, Onychopoda and Anomopoda (families Daphniidae, Moinidae, Bosminidae, Ilyocryptidae) // Zootaxa. V. 3904. P. 1–27.
Xu L., Han B.P., Van Damme K., Vierstraete A., Vanfleteren J.R., Dumont H.J., 2010. Biogeography and evolution of the Holarctic zooplankton genus Leptodora (Crustacea: Branchiopoda: Haplopoda) // Journal of Biogeography. V. 38. P. 359–370.
Yoon S.M., 2010. Arthropoda: Branchiopoda: Anostraca, Notostraca, Spinicaudata, Laevicaudata, Ctenopoda, Anomopoda, Haplopoda // Invertebrate fauna of Korea. V. 21. № 2. P. 1–156.
Дополнительные материалы отсутствуют.
Инструменты
Зоологический журнал