

Зоологический журнал, 2020, T. 99, № 10, стр. 1124-1139
Фаунистические обмены между бассейнами Северного Ледовитого океана и Каспия: история и современные процессы
А. А. Махров a, b, *, М. В. Винарский b, **, М. Ю. Гофаров c, ***, Г. А. Дворянкин c, ****, А. П. Новоселов c, *****, И. Н. Болотов b, c, ******
a Институт проблем экологии и эволюции им. А.Н. Северцова РАН
119071 Москва, Россия
b С.-Петербургский государственный университет
199034 Санкт-Петербург, Россия
c Федеральное государственное бюджетное учреждение науки
Федеральный исследовательский центр комплексного изучения Арктики
имени академика Н.П. Лаверова Уральского отделения РАН
163000 Архангельск, Россия
* E-mail: makhrov12@mail.ru
** E-mail: radix.vinarski@gmail.com
*** E-mail: zubr3@yandex.ru
**** E-mail: dga130157@gmail.com
***** E-mail: alexander.novoselov@rambler.ru
****** E-mail: inepras@yandex.ru
Поступила в редакцию 25.04.2020
После доработки 29.04.2020
Принята к публикации 29.04.2020
Аннотация
В обзоре обобщены фаунистические, молекулярно-генетические и палеогеографические данные, позволяющие выявить пути и время пересечения гидробионтами водораздела Каспийского моря и Северного Ледовитого океана. В плиоцене пресноводные фауны водоемов Европы и Северной Азии были в значительной степени сходными, но теплолюбивые виды в Сибири были практически целиком уничтожены плейстоценовыми оледенениями. В межледниковья происходило вселение холодолюбивой пресноводной фауны в бассейн Волги, видимо, как с запада, так и с востока от Урала. Представители морской фауны имели возможность проникнуть в Каспий вместе с соленой водой, поступавшей из Северного Ледовитого океана в конце плиоцена‒начале плейстоцена. В период последнего оледенения возможностей для пересечения водораздела, видимо, не было, но после завершения этого оледенения пресноводные гидробионты, в основном теплолюбивые, в результате перемещения водораздела попали в бассейны Онеги, Северной Двины и Оби. Такие вселения происходят и в настоящее время по каналам, соединившим Волгу и Северную Двину, а также в результате преднамеренного вселения.
Арктика и бассейн Каспийского моря ‒ огромные регионы, населенные множеством эндемичных видов гидробионтов, ряд которых имеет большое практическое значение (рис. 1). Не удивительно, что значительное число работ посвящено как проблеме возникновения пресноводной фауны Арктики (обзоры: Кудеpский, 1987; Weider, Hobaek, 2000; Махров, Болотов, 2006; Makhrov et al., 2019), так и проблеме формирования гидрофауны бассейна Каспийского моря (обзоры: Козлов, 1993; Dumont, 1998; Слынько, Терещенко, 2014; Krijgsman et al., 2019; Wesselingh et al., 2019). Эти фауногенетические процессы были обусловлены как событиями геологического прошлого, так и хозяйственной деятельностью человека, проявлявшейся в историческую эпоху.
Рис. 1.
Северная часть бассейна Каспийского моря и прилегающие бассейны морей Северного Ледовитого океана. I – Каспийское море, 1.1 – р. Кама, 1.2 – р. Чусовая, 1.3 – р. Шексна, 1.4 – оз. Плещеево, 1.5 – оз. Белое. II – Балтийское море, 2.1 – оз. Ладожское, 2.2 – оз. Онежское, 2.3 – оз. Водлозеро. III – Белое море, 3.1 – р. Онега, 3.2 – р. Нюхча, 3.3 – р. Сухона, 3.4 – р. Вычегда, 3.5 – оз. Воже, 3.6 – оз. Лача, 3.7 – оз. Кенозеро, 3.8 – оз. Лекшмозеро, 3.9 – оз. Кубенское, 3.10 – Соловецкие о-ва. IV – Баренцево море. V – Карское море, 5.1 – р. Иртыш, 5.2 – Тургайская ложбина.
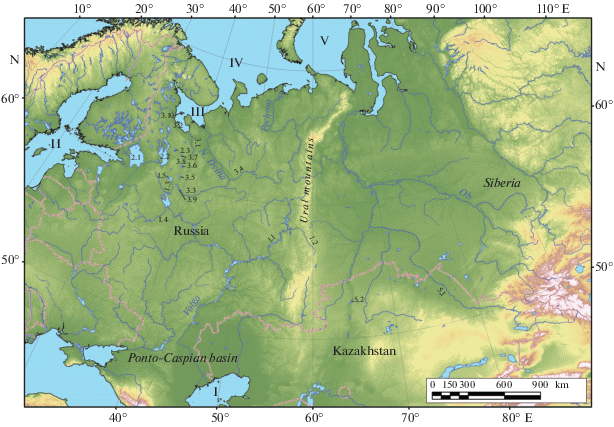
Выяснение масштаба, направлений и механизмов фаунистических обменов имеет большое значение для выявления исходного состава фауны водоемов и водотоков севера Европы и Сибири, а также для понимания особенностей ее формирования на нынешнем этапе ‒ вопроса, давно обсуждаемого в литературе (Зограф, 1895; Рылов, 1921; Домрачев, 1925; Изменение структуры …, 1982; Жаков, 1984; Румянцев, 1996; Смирнов, 2010; Алексеева и др., 2014).
Вселению видов северного происхождения в Каспийский бассейн посвящена обширная литература, начиная с классической монографии Палласа (Pallas, 1814, p. 344). Обобщив все имеющиеся сведения, Берг (1928) опубликовал список таких видов, который с того времени не пополнялся и сейчас значительно устарел.
Недостаточно освещена в современной литературе и проблема миграций в обратном направлении, т.е. из бассейна Каспия в бассейн Северного Ледовитого океана. В обзорной работе (Mordukhai-Boltovskoi, 1964) упоминается только четыре вида, вселившихся из бассейна Волги в бассейн Белого моря: дрейссена (Dreissena polymorpha (Pallas 1771)), длиннопалый рак (Astacus leptodactylus Eschscholtz 1823), стерлядь (Acipenser ruthenus Linnaeus 1758) и ее паразит, Polypodium hydriforme Ussov 1887 (Cnidaria, Polypodiozoa).
Вплоть до недавнего времени реконструкции возможных путей фаунистического обмена в прошлом основывались преимущественно на палеогеографических данных. В самом начале XX в. Э. Зюсс постулировал существование морского пролива, некогда соединявшего Северный Ледовитый океан с Аралом и даже средиземноморским бассейном. Зюсс назвал его Тургайским проливом, “Strasse von Turgai” (Suess, 1901: S. 17). Согласно Зюссу, “этот пролив сделал возможным временное сообщение между средиземноморскими и туранскими водами” (там же, S. 18). Вселение северных видов в Каспий через Тургайский пролив предполагалось многими авторами, начиная с Сарса (Sars, 1927; см. обзор Линдберг, 1972).
Многие авторы полагали, что проникновение холодолюбивых видов из Сибири в Европу через бассейн Каспия происходило по огромной системе приледниковых подпрудных озер (обзоры: Segestrale, 1982; Митенев, 1997; Svärdson, 1998; Гросвальд, 1999; Первозванский, 1999; Румянцев, 2007) либо через соединения низовий рек, расположенных на территории, ныне представляющей собой шельф арктических морей (например, Карского моря). Периодическое осушение шельфа Северного Ледовитого океана в плейстоцене позволяет понять, как мамонты и другие крупные позвоночные оказались на Новосибирских о-вах и о-ве Врангеля (Линдберг, 1972).
Другие исследователи считают, что расселение гидробионтов по приледниковым водоемам было ограничено, и колонизация освобождающихся ото льда водных систем происходила в основном из локальных рефугиумов, располагавшихся в том числе в бассейне Каспийского моря (обзор: Кудеpский, 1987).
Понто-Каспийский бассейн ‒ это родина многих инвазивных видов, распространившихся в историческое время вплоть до Северной Америки (обзор: Орлова, 2000). Активно функционирует “Северный инвазионный коридор”, по которому гидробионты через бассейн Волги проникают на север Европы (Panov et al., 2007). Изучение истории прошлых миграций гидробионтов, несомненно, будет полезно для понимания закономерностей современных инвазий.
В последние годы накопилось немало новой фаунистической, палеонтологической и палеогеографической информации и появились молекулярно-генетические методы, позволяющие реконструировать естественные фаунистические обмены между бассейнами Каспия и Северного Ледовитого океана. Генетические методы представляют собой независимый источник данных. Этот источник позволяет сделать выбор между конкурирующими гипотезами, основанными на палеогеографии. Их использование, в частности, позволило опровергнуть гипотезу о морском происхождении некоторых каспийских эндемиков, например, губок рода Metschnikowia Grimm 1876 (Sokolova et al., 2020).
В статье приведены обзор и анализ этих сведений, а также данных о современных фаунистических обменах между бассейнами Каспия и Северного Ледовитого океана. В обзор включены данные по всем группам гидробионтов, за исключением легко пересекающих водоразделы – в частности, по насекомым и некоторым ракообразным (Bekker et al., 2016, 2018; Kotov et al., 2016).
ЕСТЕСТВЕННЫЕ ВСЕЛЕНИЯ СЕВЕРНЫХ ВИДОВ В БАССЕЙН КАСПИЯ
Уточнение списка северных вселенцев в бассейн Каспия
Полученная за последние десятилетия информация позволила существенно дополнить и уточнить список каспийских видов северного происхождения, впервые составленный Л.С. Бергом (см. табл. 1). Так, Митенев (1997) констатировал, что в бассейне Каспийского моря обитает группа паразитов рыб, относящихся к арктическому пресноводному фаунистическому комплексу. Хозяин большинства из них ‒ налим (Lota lota (Linnaeus 1758)), паразиты этой группы встречаются также на лососевидных (в основном сиговых) рыбах.
Таблица 1.
Северные формы гидробионтов, обитающие в бассейне Каспийского моря, и результаты их исследования молекулярно-генетическими методами
| Таксон | Вид | Использованный генетический маркер | Подтверждение вселения из Арктики (да – +, нет – прочерк) | Ссылка |
|---|---|---|---|---|
| Ракообразные | Limnocalanus grimaldii | ‒ | ‒ | ‒ |
| Saduria entomon | ‒ | ‒ | ‒ | |
| Pseudalibrotus caspia | ‒ | ‒ | ‒ | |
| Pseudalibrotus platyceros | ‒ | ‒ | ‒ | |
| Pontoporeia microphthalma | ‒ | ‒ | ‒ | |
| Gammaracanthus loricatus | Аллозимы, митохондриальный ген COI | + | Väinölä et al., 2001 | |
| Mysis caspia | Несколько маркеров | + | Audzijonytė et al., 2005 | |
| Mysis microphthalma | Несколько маркеров | + | Audzijonytė et al., 2005 | |
| Mysis amblyops* | Несколько маркеров | + | Audzijonytė et al., 2005 | |
| Рыбы | Stenodus leucichthys | Ядерные гены | + | Голованова, 2005 |
| Coregonus albula | Митохондриальные гены ND-1, COI | +/‒ | Боровикова, 2017; Borovikova, Artamonova, 2018 | |
| Salmo trutta | Несколько маркеров | – | Махров, Болотов, 2019; Артамонова и др., 2020 | |
| Acipenser* | Митохондриальные гены, кодирующие цитохром b, NADH5, Д-петля | + | Birstein et al., 2000 | |
| Митохондриальный ген, кодирующий цитохром b | + | Jenneckens et al., 2000 | ||
| Млекопитающие | Pusa caspia* | Несколько маркеров | + | Arnason et al., 2006; Palo, Väinölä, 2006; Fulton, Strobeck, 2010 |
Молекулярно-генетические исследования показали, что в популяции русского осетра (Acipenser gueldenstaedtii Brandt 1833) бассейна Волги представлены гаплотипы митохондриальной ДНК, характерные для сибирского осетра (Acipenser baerii Brandt 1869) (Birstein et al., 2000; Jenneckens et al., 2000). Важно отметить, что в популяции русского осетра бассейна Черного моря эти гаплотипы отсутствуют. В популяции бассейна Азовского моря они присутствуют с небольшой частотой, и, вероятно, их носители попали в этот бассейн в результате искусственного вселения рыб из бассейна Каспия (Тимошкина и др., 2009).
С другой стороны, молекулярно-генетические исследования ряда предполагаемых северных вселенцев в бассейн Каспийского моря привели к неожиданному результату: выявлен высокий уровень генетической дивергенции между родственными видами из бассейна Северного Ледовитого океана и бассейна Каспийского моря. Такие данные получены для рачков родов Mysis Latreille 1802 (Väinölä, 1995; Audzijonytė et al., 2005) и Gammaracanthus Spence Bate 1862 (Väinölä et al., 2001), тюленей Pusa Scopoli 1771 (Arnason et al., 2006; Palo, Väinölä, 2006; Fulton, Strobeck, 2010).
Даже у популяций одного вида, обитающих в разных бассейнах, фиксированы разные гаплотипы митохондриальной ДНК и (или) аллели ядерных генов. Это показано для кумжи (Salmo trutta Linnaeus 1758) (Bernatchez, Osinov, 1995; Осинов, Берначе, 1996), нельмы (Stenodus leucichthys (Güldenstädt 1772)) (Голованова, 2005), сибирского тайменя (Hucho taimen (Pallas 1773)) (Marić et al., 2014). Надо отметить, что европейский таймень (Hucho hucho Linnaeus 1758), как оказалось, живет в Европе очень давно, по крайней мере с конца миоцена (Kovalchuk, 2015). Последовательность контрольного региона митохондриальной ДНК этого вида более сходна с аналогичной последовательностью сычуаньского тайменя (Hucho bleekeri Kimura 1934), чем сибирского (Wang et al., 2011).
В некоторых случаях выяснилось даже, что южные виды или формы древнее северных родственников (которые считались их предками), и, видимо, вселение происходило не с севера на юг, а с юга на север. Так, по целому ряду данных (морфологических, палеонтологических и генетических) центр происхождения кумжи находится в Понто-Каспийском бассейне (обзор: Махров, Болотов, 2019; Артамонова и др., 2020).
Результаты анализа ряда морфологических и генетических исследований до недавнего времени позволяли считать, что в оз. Белое в бассейне Волги и в оз. Водлозеро в бассейне Онежского оз. обитает сибирская ряпушка (Coregonus sardinella Valenciennes 1848) или гибрид европейской (Coregonus albula (Linnaeus 1758)) и сибирской ряпушек (обзор: Боровикова, Махров, 2012), и предполагать вселение сибирской ряпушки в эти водоемы из бассейна Белого моря в ходе последнего оледенения (Кудерский, 1977). Однако углубленный анализ результатов исследования митохондриальной ДНК ряпушек позволяет заключить, что ряпушка уже очень длительное время населяет бассейн Волги (Borovikova, Artamonova, 2018). Возможно, после последнего оледенения ряпушка действительно пересекала водораздел Каспия и Северного Ледовитого океана, но не с севера на юг, а с юга на север (но в межледниковье ряпушка могла вселиться в бассейн Волги из бассейна Печоры).
Таким образом, результаты молекулярно-генетических исследований позволяют исключить из числа северных вселенцев каспийскую кумжу, но включить в их число сибирского осетра. Включение в список сибирского осетра очень важно, поскольку, как будет показано ниже, факт вселения этого вида позволяет уверенно говорить о связи водоемов Сибири и бассейна Каспия.
О возможности вселения северных видов в бассейн Каспия по пресноводным системам
Многие исследователи предполагали, что вселение гидробионтов из бассейна Северного Ледовитого океана и (или) Балтики в бассейн Каспия происходило, вероятно, через приледниковые озера. В качестве возможных мест вселения рассматривались стык бассейнов Балтики, Белого моря и Каспия (Об исследованиях…, 1874; Högbom, 1917; Берг, 1928) и Западная Сибирь (Пирожников, 1937). Существование приледниковых озер в этих районах подтверждают геологические исследования (Об исследованиях …, 1874; Яковлев, 1928; Квасов, 1975; Гросвальд, 1999). Допускалась возможность такого вселения даже через современные соединения водоразделов Северной Двины и Камы (Подлесный, 1941).
Однако, нам представляется, что факт фиксации разных гаплотипов и (или) аллелей у родственных форм из бассейнов Северного Ледовитого океана и бассейна Каспийского моря, описанный в предыдущем разделе, не согласуется с гипотезой расселения через водораздел по приледниковым озерам в период последнего оледенения, и тем более с предположением о вселении после завершения последнего оледенения.
Видимо, приледниковые озера представляли собой мало пригодный для большинства гидробионтов биотоп. Судя по всему, это были очень холодные водоемы с мутной водой и с очень бедной фауной (Ashley, 2002).
Для пресноводных гидробионтов наиболее вероятным представляется вселение из Сибири в бассейн Каспия в плейстоцене через речные системы. Один из возможных сценариев – вселение через р. Чусовая (приток Камы). Еще в начале четвертичного периода значительная часть бассейна этой реки принадлежала к бассейну р. Оби (Щукина, 1948). При включении верховьев реки в бассейн Волги в этот бассейн могли попасть и населявшие верховья Чусовой рыбы.
Исследование морфологических признаков некоторых рыб р. Чусовая выявило их сходство с сибирскими родственниками и позволило высказать гипотезу об их вселении из бассейна Оби в плейстоцене (Костарев, 1973). В то же время современные морфологические исследования рыб Чусовой не позволяют однозначно говорить об их сходстве с сибирскими родственниками (Зиновьев, Богданов, 2017).
Однако представляется весьма вероятным, что именно при переходе верховьев р. Чусовая из бассейна Оби в бассейн Волги в последний попали нельма, сибирский осетр и сибирский таймень. Вселение ряпушки в бассейн Волги с учетом генетических данных (Borovikova, Artamonova, 2018) более вероятным представляется из бассейна Печоры.
Другие северные вселенцы (рачки родов Mysis и Gammaracanthus, тюлени Pusa), которые генетически сильнее обособлены от оставшихся в Северном Ледовитом океане родственников и редко встречаются в реках, вероятно, попали в Каспий раньше, возможно, в ходе прямого соединения морских бассейнов. По другой гипотезе (Dumont, 1998), все эти виды проникли в Каспий через былое соединение бассейнов Волги и Сухоны.
Морские проливы, соединявшие Каспийское море и Северный Ледовитый океан, и их зоогеографическое значение
В позднем мезозое–палеогене на территории современной Западной Сибири и Центрального Казахстана располагался узкий Тургайский пролив, который тянулся в меридиональном направлении и соединял Западно-Сибирское эпиконтинентальное и Среднеазиатское (Туранское) моря (Найдин, 2003; Ахметьев, 2011). Не исключено, что именно по этому проливу проникли в Понто-Каспийский бассейн предки лососей рода Salmo Linnaeus 1758, хотя их вселение через Тетис более вероятно (Артамонова и др., 2020).
Неоднократно высказывались предположения о проникновении морских вод в Каспий с севера Европы в период трансгрессий океанов (обзор: Линдберг, 1972). Предположения некоторых биологов об относительно недавних морских трансгрессиях (Полищук, 1989; Удра, 2009) не нашли подтверждения в современных геологических исследованиях. Однако в настоящее время практически доказано, что морская вода из Северного Ледовитого океана поступала через водораздел в конце плиоцена‒начале плейстоцена. Повышение уровня, отмеченное в этот период, сопровождалось повышением солености и изменением состава фауны Акчагыльского бассейна, существовавшего в конце плиоцена на месте современного Каспия (Krijgsman et al., 2019). Это хорошо объясняет, почему в осадках, отлагавшихся в Акчагыльском бассейне, присутствуют остатки морских организмов северного происхождения ‒ фораминифер и динофлагеллят (обзор: Richards et al., 2018).
Вероятно, с этим же потоком воды из Северного Ледовитого океана проникли в бассейн Каспийского моря его современные обитатели ‒ тюлени Pusa и рачки родов Mysis и Gammaracanthus. Это согласуется с данными о достаточно высокой генетической дивергенции этих таксонов от их арктических родственников (Väinölä, 1995; Väinölä et al., 2001; Audzijonytė et al., 2005; Arnason et al., 2006; Palo, Väinölä, 2006; Fulton, Strobeck, 2010).
Как показали молекулярно-генетические исследования рода Mysis, каспийские представители этого рода филогенетически ближе к пресноводным арктическим формам, чем к формам, обитающим в Северном Ледовитом океане (Audzijonytė et al., 2005). Возможно, они попали в Каспий из какого-то пресного озера, оказавшегося на пути потока морских арктических вод.
Вандендорп с соавторами (Vandendorpe et al., 2019) недавно показали, что фаунистический обмен между водоемами Каспийского бассейна и Северного Ледовитого океана в позднем плиоцене и раннем плейстоцене мог затронуть и моллюсков. Именно так авторы объясняют родственную связь между современными понто-каспийскими моллюсками рода Ecrobia Stimpson 1865 (Gastropoda, Hydrobiidae) и родственными им формами из северо-западной Атлантики. Предполагается, что этот фаунистический обмен обеспечивался как морскими трансгрессиями, так и существованием на территории Западной Сибири крупных озерных систем, служивших для моллюсков “промежуточными станциями” в их движении на север (Vandendorpe et al., 2019). Интересно, что распространение видов рода Ecrobia в значительной степени совпадает с распространением филогенетических линий лососей рода Salmo (Артамонова и др., 2020). Поэтому не исключено, что представители Ecrobia, как и представители рода Salmo, возникли в Понто-Каспийском бассейне и попали в Атлантический океан через Гибралтар.
Однако вселение понто-каспийских видов на север Европы через Гибралтар, видимо, случалось редко. Большинство таких вселенцев, как мы покажем в следующих разделах, попали из одного пресноводного бассейна в другой в результате изменения положения водораздела, постройки каналов или преднамеренного вселения.
ЕСТЕСТВЕННЫЕ ВСЕЛЕНИЯ ИЗ КАСПИЯ В БАССЕЙН СЕВЕРНОГО ЛЕДОВИТОГО ОКЕАНА
Вселенцы из бассейна Каспия в Сибири
Специалистами-систематиками неоднократно высказывалось мнение, что до эпохи плейстоценовых оледенений пресноводная фауна Сибири была весьма богата и тесно связана с европейской фауной. В частности, из Сибири в водоемы Европы вселились водяные ослики Asellus Geoffroy 1762 (Бирштейн, 1985; Vekhoff, 1994).
Богачев (1961), изучив плиоценовую фауну моллюсков юга Западной Сибири, выделил группу видов рода Unio Philipsson 1788, очень сходных с обитавшими в этот период в Европе, включая западное Приуралье. По его словам, “…в плиоценовую эпоху и сам Урал не был так поднят, и озера были значительно развиты, представляя таким образом пути переселения из системы рек западного склона в систему рек восточного, сибирского, склона” (Богачев, 1961, С. 145). Более поздними исследователями высокая степень морфологического сходства между западносибирскими плиоценовыми унионидами и современными им европейскими формами объясняется параллельной эволюцией, и большинство видов этих унионид были отнесены к современным или ископаемым родам, свойственным сино-индийской, а не европейской фауне (Старобогатов, 1970; Толстикова, 1980; Зыкин, 2012).
Однако в плиоценовых отложениях юга Западной Сибири имеется определенное число видов и родов пресноводных моллюсков, современный ареал которых ограничен Центральной и Восточной Европой. Таковы, например, представители родов Borysthenia Lindholm 1914 (Valvatidae) и Marstoniopsis van Regteren Altena 1936 (Amnicolidae) (Зыкин, 2012). Род Borysthenia известен также из среднего миоцена Зайсанской впадины (Толстикова, 1980). В плиоцене–плейстоцене, по мере прогрессирующего похолодания климата, теплолюбивая малакофауна сино-индийского типа на юге Западной Сибири постепенно заменилась фауной бореально-европейского облика, представители которой проникали на восток, по-видимому, по территории Южного Урала и современного западного Казахстана (Старобогатов, 1970; Зыкин, 2012; Винарский, 2014).
В современной малакофауне водоемов юго-западной части Иртышского бассейна представлены виды рода Caspiohybrobia Starobogatov 1970 (семейство Hydrobiidae) (Андреева, 1987; Шишкоедова, 2011), очевидно имеющего понто-каспийское происхождение.
Ихтиофауна разных регионов умеренного пояса Евразии до конца неогена была очень сходна; в частности, в неогеновых водоемах Западной Сибири обитали осетр (вид точно не определен), плотва (Rutilus rutilus (Linnaeus 1758)), сом (Silurus glanis Linnaeus 1758), лещ (Abramis brama (Linnaeus 1758)), красноперка (Scardinius erythrophtalmus (Linnaeus 1758)), уклейка (Alburnus alburnus (Linnaeus 1758)), судак (Stizostedion lucioperca (Linnaeus 1758)) (Яковлев, 1964). Только первые два вида обитают в Сибири сейчас. Плотва отмечена в этом регионе и в плиоцене (Штылько, 1934).
В водоемах Монголии в плиоцене еще обитали рыбы родов Scardinius Bonaparte 1837, Blicca Heckel 1843, Abramis Cuvier 1816, Stizostedion Rafinesque 1820 (Сычевская, 1983), но эти теплолюбивые формы, видимо, не пережили плейстоценового похолодания.
Однако, судя по молекулярно-генетическим данным, это похолодание в каком-то рефугиуме пережил сибирский осетр. Барминцева и Мюге (2017) предполагают, что этот рефугиум располагался в районе современного Байкала. Сибирский осетр, хотя и близкий родственник русского осетра, но генетически ‒ вполне самостоятельный вид (Birstein et al., 2000; Jenneckens et al., 2000; Sergeev, 2016; Luo et al., 2019). Как упоминалось выше, относительно недавно произошло возвратное вселение этого вида в бассейн Каспия (Birstein et al., 2000; Jenneckens et al., 2000; Тимошкина и др., 2009). Интересно, что из всех рыб Сибири только у осетровых, возможно, есть паразиты понто-каспийского происхождения (Трофименко, 1969).
Некоторые виды гидробионтов, по-видимому, являются относительно недавними вселенцами из Европы в Сибирь, причем в ряде случаев можно предполагать, что они таким образом восстановили некогда утраченную часть своего былого ареала. Так, ограниченные генетические данные по сравнению популяций стерляди бассейнов Каспийского моря и Сибири (Кузьмин, Кузьмина, 2005; Pobedintseva et al., 2019) показывают, что, в отличие от сибирского и русского осетров, сибирские и волжские популяции стерляди слабо дифференцированы генетически.
Плотва, судя по всему, вымерла в Сибири в результате плейстоценового оледенения. Исследование разнообразия митохондриального гена cyt b (Levin et al., 2017) показало, что современные популяции плотвы Сибири генетически очень сходны с популяциями бассейна Волги (имеют один и тот же наиболее распространенный гаплотип), что, вероятно, объясняется относительно недавним вселением этого вида в Сибирь.
Исследование разнообразия другого митохондриального гена, COI, утиной беззубки (Anodonta anatina Linnaeus 1758) также выявило значительное сходство выборок из Европы и района Байкала (Klishko et al., 2018). Нами было показало, что этот вид широко распространен в Сибири и населяет бассейны Оби, Таза, Енисея и Лены, причем все сибирские популяции сформированы представителями единственной генетической линии, которая также обнаружена в Европе, включая бассейн Волги (Bolotov et al., 2020; Tomilova et al., 2020).
Вероятно, все перечисленные выше виды использовали для расселения обширную реку. Эта река соединяла приледниковые озера, находившиеся в Западной Сибири, с бассейном Каспия через Тургайскую ложбину (Гросвальд, 1999). Видимо, в районе Тургайской ложбины река достаточно прогревалась, и упомянутые виды смогли подняться выше будущего водораздела.
Миграцией по гигантской системе приледниковых озер объяснялось генетическое сходство налима Сибири и Европы (Van Houdt et al., 2005). Однако детальное исследование выявило многочисленные гаплотипы мтДНК, “эндемичные” для налима водных систем Сибири (Хрунык и др., 2017). Отметим также, что ископаемые остатки представителя рода Lota Oken 1817 найдены в нескольких десятках километров севернее современного Аральского моря в отложениях, датируемых серединой третичного периода (Рыбы Казахстана, 1986). Это свидетельствует о весьма долгой эволюционной истории налима.
Существование южного “коридора”, соединявшего юг Восточной Европы и юго-запад Западной Сибири через водотоки Южного Урала и южного Зауралья было постулировано Винарским (2014). В фауне пресноводных гастропод Западной Сибири прослеживается сокращение доли видов-вселенцев из Европы в северо-восточном направлении; на северо-востоке региона они почти целиком замещены видами сибирского происхождения, в то время как в южном Зауралье на их долю приходится около 90% всей фауны (Винарский и др., 2012; Винарский, 2014).
Миграция моллюсков на восток через южноуральский “коридор” продолжается и поныне. Свидетельством тому может служить обнаружение в р. Уй (крайняя западная часть бассейна Иртыша) двух видов двустворчатых моллюсков подсемейства Unioninae (Andreyeva et al., 2009), ранее не отмечавшихся для этого бассейна. Физико-географические особенности Южного и Среднего Урала таковы, что в некоторых районах истоки рек Камского и Иртышского, а также Уральского и Иртышского бассейнов, практически смыкаются, что дает возможность гидробионтам мигрировать из одной реки в другую. Известны находки Unio и из верхней части бассейна Иртыша; возможно, их следует объяснять не недавним вселением, а существованием рефугиума, располагавшегося в верховьях Иртышского бассейна (Bolotov et al., 2020).
Вселенцы из бассейна Волги в бассейнах Белого и Баренцева морей
В ходе раскопок послеледниковых поселений человека в бассейне р. Онега (юго-западная часть бассейна Белого моря) обнаружены кости таких типичных южных рыб, как красноперка, жерех (Aspius aspius (Linnaeus 1758)), стерлядь, синец (Abramis ballerus (Linnaeus 1758)), сом, лещ (Никольский, 1943) и линь (Tinca tinca (Linnaeus 1758)) (Цепкин, 1999; Гимранов, Косорукова, 2019). До недавнего времени считалось, что все они в настоящее время здесь вымерли, но недавно в Кенозере, в бассейне р. Онега, обнаружены красноперка и синец (Дворянкин, 2016). Синец обитает также в бассейне р. Нюхчи, впадающей в Онежский залив Белого моря (Новоселов, 2000; Петрова, 2003). В бассейне Онеги обитает другой типичный южный вид, густера (Blicca bjoerkna (Linnaeus 1758)) (Радченко, 2002; Дворянкин, 2016).
В бассейне р. Северная Двина, занимающем юго-восточную часть бассейна Белого моря, из рыб понто-каспийского происхождения (без учета недавних вселенцев) присутствуют только лещ, голавль (Leuciscus cephalus (Linnaeus 1758)), густера (Кучина, 1967) и красноперка (Соловкина, 1969).
Кроме того, молекулярно-генетические исследования показывают, что в популяциях плотвы бассейна Северной Двины встречаются гаплотипы, принадлежащие к “Понто-Каспийской кладе” (Ponto-Caspian clade). Однако, не исключено, что носители гаплотипов этой клады проникли из бассейна Волги в бассейн Северной Двины по каналам (Levin et al., 2017).
В расположенном восточнее бассейне Печоры из понто-каспийских рыб (без учета недавних вселенцев) встречается только лещ (Никольский и др., 1947; Зверева и др., 1953; Лукин и др., 2000; Захаров, Бознак, 2019). В этом бассейне также значительно меньше паразитов рыб, относящихся к понто-каспийской экологической группе бореального равнинного фаунистического комплекса, чем в бассейне Северной Двины (Митенев, 1997; Радченко, 2002; Доровских, 2011).
Сходная тенденция прослежена при анализе разнообразия водных олигохет севера Европы: “Число южных видов заметно уменьшается по направлению на восток и северо-восток ‒ в Печорской провинции их не более 4–5” (Попченко, 1988, С. 88).
Таким образом, на европейском севере России хорошо выражено снижение числа гидробионтов понто-каспийского происхождения с запада на восток. Весьма вероятно, что вселенцы из бассейна Волги попадали в бассейн Белого моря где-то в районе стыка бассейнов Онеги и Северной Двины.
Пути вселения каспийских видов в бассейн Белого моря
Уже во второй половине 19 века И.С. Поляков писал: “… соединялись несомненно озера Волжского бассейна с озерами Балтийского и Беломорского, как на то указывают мощные пласты послеледниковых осадков и орография” (Об исследованиях …, 1874, с. 319). Многие исследователи считали, что рыбы и другие гидробионты пересекали водоразделы Каспийского, Балтийского и Белого морей по системам приледниковых озер (обзор: Кудерский, 1987).
Ряд холодолюбивых видов рыб, действительно, после ледника проник из бассейна Балтики в бассейн Белого моря. Этим путем, вероятно, прошли европейская жемчужница (Margaritifera margaritifera (Linnaeus 1758)), которая до сих пор отсутствует в бассейне Северной Двины (Беспалая и др., 2007; Makhrov et al., 2014), сиг (Coregonus lavaretus (Linnaeus 1758)) (Боровикова, Малина, 2018), кумжа (Makhrov et al., 2002), европейский хариус (Thymallus thymallus (Linnaeus 1758)) (Ponomareva et al., 2019), атлантический лосось (Salmo salar Linnaeus 1758) (Makhrov et al., 2005), арктический голец (Salvelinus alpinus (Linnaeus 1758)) (Махров и др., 2019). Европейская корюшка (Osmerus eperlanus (Linnaeus 1758)), возможно, также прошла этим путем, хотя она могла и пережить оледенение в рефугиуме в современном бассейне Печоры (Артамонова и др., 2020а).
В то же время, встречного послеледникового вселения рыб сибирского происхождения в бассейны Балтийского и Каспийского морей, видимо, не было. Распространение недавних сибирских вселенцев в Европе в основном ограничено бассейнами Баренцева и Белого морей (Махров, Лайус, 2018; Skurikhina et al., 2018; Новоселов и др., 2019; Ponomareva et al., 2019), куда они проникли, видимо, по морю. Сиг поллан, эндемик Ирландии, сходный морфологически и генетически с арктическим омулем (Coregonus autumnalis (Pallas 1776)) (Ferguson, 1978), видимо, также обогнул Скандинавию по океану. Обнаружение вариантов генов, характерных для этого вида, в бассейне Балтики (Sendek et al., 2017), скорее всего ‒ следствие преднамеренного вселения (Кудерский, 2015). Другие сибирские виды сиговых в этом регионе отсутствуют (Боровикова, Махров, 2009).
Предполагается сибирское происхождение некоторых пресноводных моллюсков, обитающих в Скандинавии (Старобогатов, 1970), но эта гипотеза пока еще не проверялась молекулярными методами.
Хотя холодолюбивые рыбы после последнего оледенения пересекали водораздел Белого моря и Балтики, по каким-то причинам пересечение границ волжского бассейна в этот период для них было затруднено. В бассейне Волги, видимо, исходно не было сига, он появился там в результате преднамеренного вселения, начатого еще в 1870‑е годы (Скаткин, 1962); до этого сиг только изредка попадал в Волгу по каналам (Кесслер, 1870). А кумжа до сих пор отсутствует в бассейне Северной Двины (Махров, 2013). Волжские вселенцы в бассейнах Онеги и Северной Двины, описанные в предыдущем разделе, ‒ в основном теплолюбивые рыбы, вероятно, пересекшие водораздел уже после отступления ледника. Скорее всего, эта группа видов обитала изначально в большом Сухонском озере, которое занимало значительную часть современного бассейна Северной Двины (Lunkka et al., 2001), но до среднего голоцена принадлежало к бассейну Волги (Квасов, 1975). Интересно, что эта группа рыб, видимо, заселила не только бассейн р. Онега, но и проникла в бассейн Онежского озера (Кудеpский, 2005).
Как отмечено выше, в бассейне Печоры почти отсутствуют вселенцы из Понто-Каспийского бассейна. Это хорошо соответствует палеогеографическим данным ‒ во время последнего оледенения водораздел Волги и Печоры не подвергался оледенению, и, соответственно, здесь не возникали возможности для соединения разных бассейнов (Mangerud et al., 2004).
Завершая описание естественных миграций гидробионтов через водораздел бассейнов Каспия и Северного Ледовитого океана, надо отметить, что этот водораздел удалось преодолеть относительно немногим видам и только в нескольких местах. Как будет показано ниже, гораздо успешнее оказались вселения через водораздел, обусловленные деятельностью человека.
ФАУНИСТИЧЕСКИЕ ОБМЕНЫ В ИСТОРИЧЕСКОЕ ВРЕМЯ
Первый канал между бассейнами Каспия и Белого моря был построен еще при Иване Грозном, во второй половине 16 века; он соединил р. Шексна в бассейне Волги и р. Сухона в верхней части бассейна Северной Двины (Гапеев, Кононов, 2009). Видимо, уже в результате постройки этого канала началась антропогенная трансформация фауны бассейна Северного Ледовитого океана.
В 1774 г. в р. Сухона был обнаружен длиннопалый рак (Georgi, 1775); в 19 в. он выявлен в среднем течении Северной Двины (Baer, 1837), в 1930-х годах ‒ в нижнем течении этой реки (Бирштейн, Виноградов, 1934).
В 1896 г. в бассейне Северной Двины впервые обнаружена дрейссена (Скориков, 1903). Судя по распространению гаплотипов митохондриального гена COI, в верхнюю часть этого бассейна она проникла через каналы. Но в популяции дрейссены устья Северной Двины обнаружен с высокой частотой гаплотип, не типичный для бассейна Волги; происхождение этой популяции неясно (Ворошилова, 2008; Voroshilova et al., 2011). Как и во многих других регионах, в северодвинской популяции обнаружены сосальщики Phyllodistomum macrocotyle (Lühe 1909) (Digenea), для которых дрейссена служит специализированным промежуточным хозяином (Travina et al., 2019).
В бассейн Северной Двины по каналам из бассейна Волги вселилось также несколько видов водных олигохет, в частности Tubifex newaensis (Michaelsen 1902) (Тимм, 1987).
В настоящее время в бассейне Северной Двины встречаются несколько видов рыб, проникших сюда, скорее всего, через каналы, ‒ жерех, белоглазка (Abramis sapa (Pallas 1814)), чехонь (Pelecus cultratus (Linnaeus 1758)), обыкновенная щиповка (Cobitis taenia Linnaeus 1758) и стерлядь (обзоры: Берг, 1945; Новоселов, 2000; Захаров, Бознак, 2011). Как отмечалось выше, не исключено, что носители “Понто-Каспийской клады” гаплотипов плотвы также вселились в бассейн Северной Двины из Волги через каналы (Levin et al., 2017).
Берг (1949) полагал, что через каналы в бассейн Северной Двины проникли верховка (Leucaspius delineatus (Heckel 1843)) и густера. Однако недавние находки этих видов и в бассейне р. Онега (Радченко, 2002; Дворянкин, 2016) показывают, что эти виды, скорее всего, пересекли водораздел естественным путем.
Кроме того, в бассейн Северного Ледовитого океана с 19 в. искусственно вселялись полезные человеку гидробионты, в том числе из бассейна Каспийского моря. Судак вселен в бассейны рек Онега и Северная Двина (Новоселов, 2000) и ряд водоемов беломорской Карелии (Стерлигова, Ильмаст, 2009). Стерлядь вселена в р. Онега (Новоселов, 2000), в Печору (Захаров и др., 1998) и на Соловецкие о-ва (Алексеева и др., 2014).
Недавно в бассейне р. Печора обнаружены верховка и уклейка (Захаров, Бознак, 2011) – видимо, это случайные вселенцы. А в озере Святое на Соловецких о-вах во второй половине 20 в. пойман длиннопалый рак (Новосельцев, Попченко, 1972), видимо, вселенный человеком. К началу 21 в. этот вид широко распространился по пресным водоемам Большого Соловецкого о-ва (Borovikova et al., 2016). Длиннопалый рак также был преднамеренно вселен в бассейн Оби и широко там распространился (Бирштейн, Виноградов, 1934). Успешным также было вселение в этот бассейн ряда рыб из бассейна Каспия ‒ здесь натурализовались судак, лещ, верховка, уклейка (Интересова, 2016).
Отметим, что, согласно обзорным работам (Орлова, 2000; Слынько и др., 2010; Karabanov et al., 2018; Жохов и др., 2019), в бассейне Каспия в последние годы появился только один северный вселенец ‒ трехиглая колюшка (Gasterosteus aculeatus Linnaeus 1758). Однако и эта рыбка попала в бассейн Волги не из бассейна Белого моря, а из бассейна Онежского озера (Артамонова и др., 2015). По данным Решетникова и Поповой (2012), изредка в бассейн Волги попадает нельма из бассейна Северной Двины.
Успешное искусственное переселение ряда гидробионтов из бассейна Каспия как в европейскую, так и в азиатскую части бассейна Северного Ледовитого океана свидетельствует, что водораздел двух бассейнов уже длительный период времени представляет собой непреодолимую естественную преграду для расселения многих гидробионтов. Поэтому высока вероятность, что постройка каналов, случайные и преднамеренные вселения понто-каспийских видов в бассейн Северного Ледовитого океана приведут к возникновению многочисленных популяций.
Можно предсказать, что целенаправленные или случайные переселения гидробионтов приведут к вспышкам численности переселенцев, а в случае патогенных организмов ‒ к масштабным вспышкам заболеваний. Примером может быть недавнее вселение в реки Норвегии и бассейна Белого моря опасного паразита Gyrodactylus salaris Malmberg 1957 (Monogenea), который ранее был эндемиком бассейна Балтийского моря (Kudersky et al., 2003).
ЗАКЛЮЧЕНИЕ
Как видно из представленных в работе данных, миграция видов через водораздел Каспия и Северного Ледовитого океана происходила в ограниченное время и на ограниченных участках, причем вселение с юга на север и в обратном направлении имело место, видимо, не одновременно и в разных местах. Исключением являются широко распространенные, легко расселяющиеся и экологически крайне пластичные виды, такие, как дафнии и насекомые.
Обмен пресноводной фауной между Сибирью и бассейном Волги имел место в плиоцене (он шел, видимо, в обоих направлениях), в плейстоцене происходили вселения холодноводных гидробионтов в бассейн Волги из бассейнов Оби и Печоры, после оледенения относительно теплолюбивые гидробионты проникали из бассейна Каспия в бассейны рек Северной Двины и Онеги, а также в бассейн Оби. Морские виды проникли в Каспий вместе с соленой водой из Северного Ледовитого океана, вероятно, в акчагыльское время.
Мы должны констатировать, что красивая теория расселения гидробионтов от Сибири до Балтики в период последнего оледенения по цепи приледниковых озер не имеет фактических доказательств. Гипотеза о вселении сибирских видов в Европу вдоль арктического побережья подтверждается, но только один из этих видов (арктический омуль) обогнул Скандинавию и достиг Западной Европы.
Поскольку физические барьеры в значительной степени задерживали продвижение гидробионтов на север Европы и в Сибирь после отступления оледенения, велика вероятность, что случайные и направленные вселения гидробионтов из бассейна Каспийского моря в южные части бассейна Северного Ледовитого океана в будущем окажутся успешными.
Список литературы
Алексеева Я.И., Андреева А.П., Груздева М.А., Дворянкин Г.А., Кузищин К.В., Махров А.А. и др., 2014. Пресноводная ихтиофауна Соловецких островов (Белое море, Европейский Север России): история формирования и современное состояние // Российский журнал биологических инвазий. № 2. С. 2–14.
Андреева С.И., 1987. Моллюски рода Caspiohydrobia Starobogatov, 1970 (Gastropoda, Pyrgulidae) водоемов Казахстана // Моллюски. Результаты и перспективы их исследований. Восьмое Всесоюзное совещание по изучению моллюсков. Ленинград, апрель 1987 г. Авторефераты докладов. Л. С. 178–180.
Артамонова В.С., Афанасьев С.А., Бардуков Н.В., Голод В.М., Кокодий С.В., Кулиш А.В. и др., 2020. Центр происхождения и пути расселения благородных лососей Salmo (Salmonidae, Actinopterigii) // Доклады РАН. Науки о жизни. Т. 493. С. 333–340.
Артамонова В.С., Махров А.А., Попов И.Ю., Спицын В.М., 2020а. Европейская корюшка Osmerus eperlanus (Linnaeus, 1758) на острове Колгуев (Баренцево море) и причины, ограничивающие распространение этого вида в Арктике // Сибирский экологический журнал. № 2. С. 160–166.
Артамонова В.С., Махров А.А., Семенов Д.Ю., 2015. Источник заселения трехиглой колюшкой (Gasterosteus aculeatus) бассейна Каспийского моря // Программа и материалы международной научной конференции, посвящённой 50-летию Зоологического музея им. М.И. Глобенко Таврической академии Крымского федерального университета им. В.И. Вернадского (Симферополь, 16–18 сентября 2015 г.). Симферополь. С. 12–13.
Ахметьев М.А., 2011. Проблемы стратиграфии и палеогеографии палеогена средних широт центральной Евразии // Геология и геофизика. Т. 52. № 10. С. 1367–1387.
Барминцева А.Е., Мюге Н.С., 2017. Природный генетический полиморфизм и филогеография сибирского осетра Acipenser baerii Brandt, 1869 // Генетика. Т. 53. № 3. С. 345–355.
Берг Л.С., 1928. О происхождении северных элементов в фауне Каспия // Доклады АН СССР. № 7. С. 107–112.
Берг Л.С., 1945. О стерляди в бассейне Белого моря // Природа. № 6. С. 66–67.
Берг Л.C., 1949. Рыбы пресных вод СССР и сопредельных стран. Часть 3. 4-е Изд., испр. и доп. М.-Л.: Изд-во АН СССР. С. 929–1382.
Беспалая Ю.В., Болотов И.Н., Махров А.А., 2007. Состояние популяции европейской жемчужницы Margaritifera margaritifera (L.) (Mollusca, Margaritiferidae) на северо-восточном краю ареала (р. Солза, бассейн Белого моря) // Экология. № 3. С. 204–211.
Бирштейн Я.А., 1985. Генезис пресноводной, пещерной и глубоководной фаун. М.: Наука. 247 с.
Бирштейн Я.А., Виноградов Л.Г., 1934. Пресноводные Decapoda СССР и их географическое распространение // Зоологический журнал. Т. 13. Вып. 1. С. 39–70.
Богачев В.В., 1961. Материалы к истории пресноводной фауны Евразии. Киев: Изд. АН УССР. 403 с.
Боровикова Е.А., 2017. Особенности генетической структуры и происхождения популяции ряпушки Coregonus albula озера Плещеево // Известия РАН. Серия биол. № 3. С. 228–233.
Боровикова Е.А., Малина Ю.И., 2018. Филогеография сига (Coregonus lavaretus L.) водоемов северо-запада европейской территории России // Сибирский экологический журнал. № 3. С. 311–324.
Боровикова Е.А., Махров А.А., 2009. Систематическое положение и происхождение сигов (Coregonus, Cоregonidae, Osteichthyеs) Европы. Генетический подход // Успехи современной биологии. Т. 129. № 1. С. 58–66.
Боровикова Е.А., Махров А.А., 2012. Изучение популяций переходной зоны между европейской и сибирской ряпушками (Coregonus): роль среды обитания в видообразовании // Принципы экологии. Т. 1. № 4. С. 5–18.
Винарский М.В., 2014. Легочные моллюски (Mollusca: Gastropoda: Lymnaeiformes) водоемов Урала и Западной Сибири. Дис. … докт. биол. наук. Омск: Омский государственный педагогический университет. 547 с.
Винарский М.В., Андреев Н.И., Андреева С.И., Каримов А.В., Лазуткина Е.А., 2012. Широтная изменчивость разнообразия пресноводных брюхоногих моллюсков (Mollusca: Gastropoda) водоемов Западной Сибири // Биология внутренних вод. № 1. С. 75–83.
Ворошилова (Сергеева) И.С., 2008. Происхождение и популяционная структура периферических популяций Dreissena polymorpha (Pallas, 1771) северо-восточной границы ареала вида. Автореф. дис. … канд. биол. наук. Борок: ИБВВ РАН. 24 с.
Гапеев А.М., Кононов В.В., 2009. Водно-транспортные соединения России в XVII–XIX вв. // Журнал университета водных коммуникаций. № 1. С. 6–21.
Гимранов Д.О., Косорукова Н.В., 2019. Данные о рыболовстве в бассейне озера Воже по материалам археологического памятника Караваиха 4 (средний голоцен) // Динамика экосистем в голоцене. К 100-летию Льва Григорьевича Динесмана. Матер. V Всерос. науч. конф. с межд. участием. Москва, 11–15 ноября 2019 г. М.: Медиа-ПРЕСС. С. 73–75.
Голованова Т.С., 2005. Анализ генетической изменчивости белорыбицы и нельмы Stenodus leucichthys (Güldenstädt, 1772) в связи с задачами искусственного воспроизводства. Автореф. дис. … канд. биол. наук. М.: Межведомственная Ихтиологическая Комиссия. 24 с.
Гросвальд М.Г., 1999. Евразийские гидросферные катастрофы и оледенение Арктики. М.: Научный мир. 120 с.
Дворянкин Г.А., 2016. Рыбы Кенозерского национального парка. Архангельск: ФГБУ Нац. парк “Кенозерский”. 100 с.
Домрачев П.Ф., 1925. К вопросу о ледниковых реликтах в фауне озерных рыб // Известия отдела прикладной ихтиологии и научно-промысловых исследований. Т. 3. Вып. 2. С. 210–214.
Доровских Г.Н., 2011. Зоогеография паразитов рыб главных рек северо-востока Европы. Сыктывкар: Изд-во Сыктывкарского гос. ун-та. 142 с.
Жаков Л.А., 1984. Формирование и структура рыбного населения озер северо-запада СССР. М.: Наука. 144 с.
Жохов А.Е., Пугачёва М.Н., Молодожникова Н.М., Беречикидзе И.А., 2019. Чужеродные виды паразитов рыб в бассейне Волги: обзор данных по числу видов и распространению // Российский журнал биологических инвазий. Т. 10. № 1. С. 38–55.
Захаров А.Б., Бознак Э.И. 2011. Современные изменения рыбного населения крупных рек Европейского северо-востока России // Российский журнал биологических инвазий. Т. 2. № 1. С. 23–33.
Захаров А.Б., Бознак Э.И., 2019. Рыбное население водоемов Тимана. Сыктывкар: ФНЦ Коми НЦ УрО РАН. 184 с.
Захаров А.Б., Крылова В.Д., Осипова О.С., 1998. Итоги и перспективы интродукции северодвинской стерляди Acipenser ruthenus в бассейн Печоры // Вопросы ихтиологии. Т. 38. № 6. С. 825–829.
Зверева О.С., Кучина Е.С., Остроумов Н.А., 1953. Рыбы и рыбный промысел среднего и нижнего течения Печоры. М.: Изд-во АН СССР. 230 с.
Зиновьев Е.А., Богданов В.Д., 2017. О возможности наличия сибирских форм рыб в бассейне Камы // Фауна Урала и Сибири. № 2. С. 62–68.
Зограф Н., 1895. Опыт объяснения происхождения фауны озер Европейской России // Известия Императорской Академии Наук. Т. 3. № 2. С. 173–191.
Зыкин В.С., 2012. Стратиграфия и эволюция природной среды и климата в позднем кайнозое юга Западной Сибири. Новосибирск: Гео. 487 с.
Изменение структуры рыбного населения эвтрофируемого водоема, 1982. М.: Наука. 248 с.
Интересова Е.А., 2016. Чужеродные виды рыб в бассейне Оби // Российский журнал биологических инвазий. Т. 7. № 1. С. 83–100.
Квасов Д.Д., 1975. Позднечетвертичная история крупных озер и внутренних морей Восточной Европы. Л.: Наука. 278 с.
Кесслеp К.Ф., 1870. Об ихтиологической фауне реки Волги // Труды С.-Петербургского общества естествоиспытателей. Т. 1. Вып. 2. С. 236–310.
Козлов В.И., 1993. Экологическое прогнозирование ихтиофауны пресных вод (на примере Понто-Каспийского региона). М.: ВНИРО. 252 с.
Костарев Г.Ф., 1973. О генезисе ихтиофауны р. Чусовой // Вопросы ихтиологии. Т. 13. Вып. 4. С. 611–617.
Кудерский Л.А., 1977. О путях проникновения сибирской ряпушки в водоемы Балтийского и Каспийского бассейнов // Известия ГосНИОРХ. Т. 111. С. 54–62.
Кудеpский Л.А., 1987. Роль ихтиологических данных в изучении путей формирования водных систем Северо-Запада и Севера Европейской части СССР // Сборник научных трудов ГосНИОРХ. Вып. 264. С. 147–156.
Кудерский Л.А., 2005. Пути формирования ихтиофауны Онежского озера // Биогеография Карелии. Труды Карельского научного центра РАН. Вып. 7. С. 147–163.
Кудерский Л.А., 2015. Избранные труды. Исследования по ихтиологии, рыбному хозяйству и смежным дисциплинам. Том 4. Акклиматизация рыб в водоемах России. М.-СПб.: Товарищество научных изданий КМК. 290 с.
Кузьмин Е.В., Кузьмина О.Ю., 2005. Популяционный анализ электрофоретической изменчивости альбуминов сыворотки крови европейской (Acipenser ruthenus Linnaeus) и сибирской (A. ruthenus marsiglii Brandt) стерляди // Генетика. Т. 41. № 2. С. 246–253.
Кучина Е.С., 1967. Общая характеристика ихтиофауны, стерлядь и лососевые р. Северной Двины // Известия Коми Филиала Всесоюзного географического общества. Т. 2. № 1. С. 92–105.
Линдберг Г.У., 1972. Крупные колебания уровня океана в четвертичный период. Биогеографические обоснования гипотезы. Л.: Наука. 548 с.
Лукин А.А., Даувальтер В.А., Новоселов А.П., 2000. Экосистема Печоры в современных условиях. Апатиты: Изд. Кольского научного центра РАН. 192 с.
Махров А.А., 2013. Кумжа (Salmo trutta L.) на северо-восточном краю ареала // Принципы экологии. Т. 2. № 1. С. 5–19.
Махров А.А., Болотов И.Н., 2006. Пути расселения и видовая принадлежность пресноводных животных севера Европы (обзор молекулярно-генетических исследований) // Генетика. Т. 42. № 10. С. 1319–1334.
Махров А.А., Болотов И.Н., 2019. Экологические причины высокой морфологической пластичности представителей таксона, обитающих в центре его происхождения (на примере благородных лососей Salmo) // Известия РАН. Серия биологическая. № 1. С. 43–51.
Махров А.А., Болотов И.Н., Спицын В.М., Гофаров М.Ю., Артамонова В.С., 2019. Жилые и проходные формы арктического гольца (Salvelinus alpinus) Европейского Севера России – пример высокой экологической пластичности без видообразования // Доклады РАН. Т. 485. № 2. С. 242–246.
Махров А.А., Лайус Д.Л., 2018. Послеледниковое вселение рыб и миноги из Тихого океана в моря севера Европы // Сибирский экологический журнал. № 3. С. 265–279.
Митенев В.К., 1997. Паразиты пресноводных рыб Кольского Севера. Мурманск: Изд-во ПИНРО. 199 с.
Найдин Д.П., 2003. Тургайский пролив в системе меридионального соединения позднемеловых морей северного полушария // Бюллетень Московского общества испытателей природы. Отдел геологический. Т. 78. Вып. 4. С. 49–55.
Никольский Г.В. 1943. К истории ихтиофауны бассейна Белого моря // Зоологический журнал. Т. 22. Вып. 1. С. 27–32.
Никольский Г.В., Громчевская Н.А., Морозова Г.И., Пикулева В.А., 1947. Рыбы бассейна Верхней Печоры. М.: МОИП. 224 с.
Новоселов А.П., 2000. Современное состояние рыбной части сообществ в водоемах европейского северо-востока России. Дис. … докт. биол. наук. Архангельск: СевПИНРО. 390 с.
Новоселов А.П., Кондаков А.В., Гофаров М.Ю., Болотов И.Н., 2019. Обыкновенная малоротая корюшка Hypomesus olidus (Osmeridae) – новый вид для фауны Баренцева моря // Вопросы ихтиологии. Т. 59. № 1. С. 28–32.
Новосельцев Г.Е., Попченко В.И., 1972. Донная фауна озер Большого Соловецкого острова // Труды СевНИОРХ. Т. 6. С. 67–84.
Об исследованиях в верховьях Волги И.С. Полякова, 1874. Известия Императорского Русского географического общества. Т. 10. № 8. С. 318–321.
Орлова М.И., 2000. Каспийский бассейн как регион-донор и регион-реципиент биоинвазий водных беспозвоночных // Виды-вселенцы в европейских морях России. Апатиты: Изд-во Кольского НЦ РАН. С. 58–75.
Осинов А.Г., Берначе Л., 1996. “Атлантическая” и “дунайская” филогенетические группы кумжи Salmo trutta complex: генетическая дивергенция, эволюция, охрана // Вопросы ихтиологии. Т. 36. № 6. С. 762–786.
Пеpвозванский В.Я., 1999. Формирование современной ихтиофауны внутренних водоемов Карелии в поздне- и послеледниковое время: новый взгляд на проблему // Биологические основы изучения, освоения и охраны животного и растительного мира, почвенного покрова Восточной Фенноскандии. Тез. докл. междунар. конф. и выездной сессии Отделения общей биологии РАН, г. Петрозаводск, 6–10 сентября 1999 г. Петрозаводск. С. 151.
Петрова Л.П., 2003. Рыбы водоемов национального парка “Водлозерский”. Петрозаводск: НП “Водлозерский”. 44 с.
Пирожников П.Л., 1937. К вопросу о происхождении северных элементов в фауне Каспия // Доклады АН СССР. Т. 15. № 8. С. 513–516.
Подлесный А.В., 1941. Географическое распространение белорыбицы Stenodus leucichthys (Güldenstädt) и ее происхождение в бассейне Каспия // Зоологический журнал. Т. 20. Вып. 3. С. 433–445.
Полищук В.В., 1989. Особенности изменений гидрографии Понто-Каспийско-Аральской солоноватой области за историческое время и их биогеографическая интерпретация // Труды Зоологического института АН СССР. Т. 199. С. 115–143.
Попченко В.И., 1988. Водные малощетинковые черви [Oligochaeta limicola] Севера Европы. Л.: Наука. 287 с.
Радченко Н.М., 2002. Паразиты рыб озера Воже (Систематика, фауна, экология, зоогеография). Вологда: Вологодский институт развития образования. 160 с.
Решетников Ю.С., Попова О.А., 2012. Лососеобразные и корюшковые рыбы в водохранилищах Волги // Бассейн Волги в XXI-м веке: структура и функционирование экосистем водохранилищ. Ижевск. С. 236–239.
Румянцев Е.А., 1996. Эволюция фауны паразитов рыб в озерах. Петрозаводск: Карельский НЦ РАН. 188 с.
Румянцев Е.А., 2007. Паразиты рыб в озерах Европейского Севера (фауна, экология, эволюция). Петрозаводск: Изд-во ПетрГУ. 252 с.
Рыбы Казахстана, 1986. В 5-ти т. Т. 1. Алма-Ата: Наука. 272 с.
Рылов В.М., 1921. К вопросу о ледниковых реликтах в пресноводной фауне // Известия Российского гидрологического института. № 1-3. С. 97–113.
Скаткин П.Н., 1962. Биологические основы искусственного рыборазведения. М.: Изд-во АН СССР. 244 с.
Скориков А.С., 1903. Распространение Dreissensia polymorpha (Pallas) в России. Зоогеографический очерк. Саратов: Паровая скоропечатня губернского правления. 48 с.
Слынько Ю.В., Дгебуадзе Ю.Ю., Новицкий Р.А., Христов О.А., 2010. Инвазии чужеродных рыб в бассейнах крупнейших рек Понто-Каспийского бассейна: состав, векторы, инвазионные пути и темпы // Российский журнал биологических инвазий. Т. 2. № 4. С. 74–89.
Слынько Ю.В., Терещенко В.Г., 2014. Рыбы пресных вод Понто-Каспийского бассейна (Разнообразие, фауногенез, динамика популяций, механизмы адаптаций). М.: Изд-во ПОЛИГРАФ-ПЛЮС. 328 с.
Смирнов Н.Н., 2010. Историческая экология пресноводных зооценозов. М.: Товарищество научных изданий КМК. 225 с.
Соловкина Л.Н., 1969. О находках серебряного карася [Carassius auratus gibelio (Bloch)] и красноперки [Scardinius erythrophthalmus (L.)] на Европейском Северо-Востоке СССР // Вопросы ихтиологии. Т. 9. Вып. 5. С. 945−948.
Старобогатов Я.И., 1970. Фауна моллюсков и зоогеографическое районирование континентальных водоемов земного шара. Л.: Наука. 372 с.
Стерлигова О.П., Ильмаст Н.В., 2009. Виды-вселенцы в водных экосистемах Карелии // Вопросы ихтиологии. Т. 49. № 3. С. 372–379.
Сычевская Е.К., 1983. История формирования ихтиофауны Монголии и проблема фаунистических комплексов // Рыбы Монгольской Народной Республики. М.: Наука. С. 225–249.
Тимм Т., 1987. Малощетинковые черви (Oligochaeta) водоемов Северо-Запада СССР. Таллин: Валгус. 300 с.
Тимошкина Н.Н., Барминцева А.Е., Усатов А.В., Мюге Н.С., 2009. Внутривидовой генетический полиморфизм русского осетра (Acipenser gueldenstaedtii) // Генетика. Т. 45. № 9. С. 1250–1259.
Толстикова Н.В., 1980. Моллюски // Палеолимнология Зайсана. Л.: Наука. С. 69–88.
Трофименко В.Я., 1969. К вопросу о генезисе гельминтофауны пресноводных рыб азиатской Субарктики // Известия АН СССР. Серия биологическая. № 6. С. 912–918.
Удра И.Ф., 2009. Биогеографическое подтверждение морских трансгрессий и внутриматериковых подтоплений на территории Евразии в антропогене // Известия РАН. Серия географическая. № 2. С. 46–59.
Хрунык Ю.Я., Богданов В.Д., Ялковская Л.Э., Копориков А.Р., Ракитин С.Б., Сибиряков П.А., Бородин А.В., 2017. Генетическое разнообразие налима (Lota lota L., 1758) Западной Сибири по результатам анализа полиморфизма мтДНК // Генетика. Т. 53. № 2. С. 214–222.
Цепкин Е.А., 1999. Ихтиофауна бассейна реки Онеги – четкий индикатор изменений климата в голоцене // Вопросы ихтиологии. Т. 39. № 1. С. 117–119.
Шишкоедова О.С., 2011. К фауне и распространению моллюсков рода Caspiohydrobia (Mollusca: Gastropoda: Hydrobiidae) в Зауралье и Западной Сибири // Экология: сквозь время и расстояние. Материалы конференции молодых ученых, 11–15 апреля 2011 г. Екатеринбург: Гощицкий. С. 201–205.
Штылько Б.А., 1934. Неогеновая фауна пресноводных рыб западной Сибири // Труд. Всесоюзного геолого-разведочного объединения НКТП СССР. Вып. 359. 96 с.
Щукина Е.Н., 1948. Четвертичные отложения Среднего Урала // Труды Института геологических наук. Вып. 94. С. 1–83.
Яковлев В.Н., 1964. История формирования фаунистических комплексов пресноводных рыб // Вопросы ихтиологии. Т. 4. Вып. 1. С. 10–22.
Яковлев С.А., 1928. О связи Балтийского моря с бассейном р. Волги в позднеледниковое время // Доклады АН СССР. Т. А. № 3. С. 41–46.
Andreeva S.I., Vinarski M.V., Karimov A.V., 2009. The first record of Unio species (Bivalvia: Unionidae) in the Irtysh River basin (Western Siberia, Russia) // Mollusca. V. 27. P. 87–91.
Arnason U., Gullberg A., Janke A., Kullberg M., Lehman N., Petrov E.A., Väinölä R., 2006. Pinniped phylogeny and a new hypothesis for their origin and dispersal // Molecular Phylogenetics and Evolution. V. 41. P. 345–354.
Ashley G.M., 2002. Glaciolacustrine environments // Modern and past clacial environments. Ed. J. Menzies. Butterworth-Heinemann: Elsevier. P. 335–359.
Audzijonytė A., Damgaard J., Varvio S.-L., Vainio J.K., Väinölä R., 2005. Phylogeny of Mysis (Crustacea, Mysida): history of continental invasions inferred from molecular and morphological data // Cladistics. V. 21. P. 575–596.
Baer K., 1837. Expédition a Novaïa Zemlia et en Laponie // Bulletin scientifique publé par L’Academie Impériale des Sciences de Saint-Pétersbourg. V. 2. P. 315–319.
Bekker E.I., Karabanov D.P., Galimov Y.R., Kotov A.A., 2016. DNA Barcoding Reveals High Cryptic Diversity in the North Eurasian Moina Species (Crustacea: Cladocera) // PLoS ONE. V. 11. № 8. e0161737.
Bekker E.I., Karabanov D.P., Galimov Y.R., Haag C.R., Neretina T.V., Kotov A.A., 2018. Phylogeography of Daphnia magna Straus (Crustacea: Cladocera) in Northern Eurasia: Evidence for a deep longitudinal split between mitochondrial lineages // PLoS ONE. V. 13. № 3. e0194045.
Bernatchez L., Osinov A., 1995. Genetic diversity of trout (genus Salmo) from its most eastern native range based on mitochondrial DNA and nuclear gene variation // Molecular Ecology. V. 4. P. 285–297.
Birstein V.J., Doukakis P., DeSalle R., 2000. Polyphyly of mtDNA lineages in the Russian sturgeon, Acipenser gueldenstaedtii: forensic and evolutionary implications // Conservation Genetics. V. 1. P. 81–88.
Bolotov I.N., Kondakov A.V., Konopleva E.S., Vikhrev I.V., Aksenova O.V., Aksenov A.S. et al., 2020. Integrative taxonomy, biogeography and conservation of freshwater mussels (Unionidae) in Russia // Scientific Reports. V. 10. P. 1–20.
Borovikova E.A., Alekseeva Ja.A., Bagirov N.E., Makhrov A.A., Popov I.Yu., 2016. Genetic identification of a crayfish (Astacus) species at the northern edge of their distribution area (Solovetsky Islands, White Sea) // Biochemical Systematics and Ecology. V. 65. P. 205–208.
Borovikova E.A., Artamonova V.S., 2018. Morphological specificities of vendace (Salmoniformes: Salmonidae: Coregoninae: Coregonus albula) population in Lake Pleshcheyevo (the Volga River basin): relationships of two phylogenetic lineages in a new zone of secondary contact // Organisms Diversity & Evolution. V. 18. P. 355–366.
Dumont H.J., 1998. The Caspian Lake: history, biota, structure, and function // Limnology and Oceanography. V. 43. P. 44–52.
Ferguson A., 1978. Systematics of the Irish pollan (Coregonus pollan Thompson): an electrophoretic comparision with other Holarctic Coregoninae // Journal of Fish Biology. V. 12. P. 221–233.
Fulton T.L., Strobeck C., 2010. Multiple fossil calibrations, nuclear loci and mitochondrial genomes provide new insight into biogeography and divergence timing for true seals (Phocidae, Pinnipedia) // Journal of Biogeography. V. 37. P. 814–829.
Georgi J.G., 1775. Bemerkungen einer Reise im Russischen Reich in der Jahren 1773 und 1774. V. 2. St. Peterburg: Kaiserl. Academie der Wissenschaften. 506 s.
Högbom A.G., 1917. Über die arktischen Elemente in der aralokaspischen Fauna, ein tiergeographisches Problem // Bulletin of the Geological Institution of University of Upsala. V. 14. P. 241–260.
Jenneckens I., Meyer J.-N., Debus L., Pitra C., Ludwig A., 2000. Evidence of mitochondrial DNA clones of Siberian sturgeon, Acipenser baerii, within Russian sturgeon, Acipenser gueldenstaedtii, caught in the River Volga // Ecology Letters. V. 3. P. 503–508.
Karabanov D.P., Pavlov D.D., Bazarov M.I., Borovikova E.A., Gerasimov Yu.V., Kodukhova Yu.V. et al., 2018. Alien species of fish in the littoral of Volga and Kama reservoirs (results of complex expeditions of IBIW RAS in 2005–2017) // Transactions of Institute for biology of Inland Waters RAS. V. 82. P. 67–80.
Klishko O.K., Lopes-Lima M., Bogan A.E., Matafonov D.V., Froufe E., 2018. Morphological and molecular analyses of Anodontinae species (Bivalvia, Unionidae) of Lake Baikal and Transbaikalia // PLoS ONE. V. 13. № 4. e0194944.
Kotov A.A., Karabanov D.P., Bekker E.I., Neretina T.V., Taylor D.J., 2016. Phylogeography of the Chydorus sphaericus Group (Cladocera: Chydoridae) in the Northern Palearctic // PLoS ONE. V. 11. № 12. e0168711.
Kovalchuk O.M., 2015. The first appearance of Hucho (Salmonidae) in the fossil record of Eastern Europe // Vestnik Zoologii. V. 49. P. 413–420.
Krijgsman W., Tesakov A., Yanina T., Lazarev S., Danukalova G., Van Baak C.G.C. et al., 2019. Quaternary time scales for the Pontocaspian domain: Interbasinal connectivity and faunal evolution // Earth-Science Reviews. V. 188. P. 1–40.
Kudersky L.A., Ieshko E., Shulman B., 2003. Distribution range formation history of the monogenean Gyrodactylus salaris Malmberg, 1957 – a parasite of juvenile Atlantic salmon Salmo salar Linnaeus, 1758 // Atlantic salmon: biology, conservation and restoration. Petrozavodsk: Institute of Biology KRC RAS. P. 77–83.
Levin B.A., Simonov E.P., Ermakov O.A., Levina M.A., Interesova E.A., Kovalchuk O.M. et al., 2017. Phylogeny and phylogeography of the roaches, genus Rutilus (Cyprinidae), at the Eastern part of its range as inferred from mtDNA analysis // Hydrobiologia. V. 788. P. 33–46.
Lunkka J.P., Saarnisto M., Gey V., Demidov I., Kiselova V., 2001. Extent and age of the Last Glacial Maximum in the southeastern sector of the Scandinavian Ice Sheet // Global and Planetary Change. V. 31. P. 407–425.
Luo D., Li Y., Zhao Q., Zhao L., Ludwig A., Peng Z., 2019. Highly Resolved Phylogenetic Relationships within Order Acipenseriformes According to Novel Nuclear Markers // Genes. V. 10, 38.
Makhrov A.A., Bolotov I.N., Artamonova V.S., Borovikova E.A., 2019. Mountain regions are the putative place of origin of many Arctic animal and plant forms // Zoologicheskiy Zhurnal. V. 98. № 11. P. 1291–1303.
Makhrov A., Bespalaya J., Bolotov I., Vikhrev I., Gofarov M., Alekseeva Ya., Zotin A., 2014. Historical geography of pearl harvesting and current status of populations of freshwater pearl mussel Margaritifera margaritifera (L.) in the western part of Northern European Russia // Hydrobiologia. V. 735. P. 149–159
Makhrov A.A., Skaala O., Altukhov Yu.P., 2002. Alleles of sAAT-1,2* isoloci in brown trout: potential diagnostic marker for tracking routes of post-glacial colonization in northern Europe // Journal of Fish Biology. V. 61. P. 842–846.
Makhrov A.A., Verspoor E., Artamonova V.S., O’Sullivan M., 2005. Atlantic salmon colonization of the Russian Arctic coast: pioneers from North America // Journal of Fish Biology. V. 67. Supplement A. P. 68–79.
Mangerud J., Jacobsson M., Alexanderson H., Astakhov V., Clarke G.K.C., Henriksen M. et al., 2004. Ice-dammed lakes and rerouting of the drainage of northern Eurasia during the Last Glaciation // Quaternary Science Review. V. 23. P. 1313–1332.
Marić S., Alekseyev S., Snoj A., Askeyev O., Askeyev I., Weiss S., 2014. First mtDNA sequencing of Volga and Ob basin taimen Hucho taimen: European populations stem from a late Pleistocene expansion of H. taimen out of western Siberia and are not intermediate to Hucho hucho // Journal of Fish Biology. V. 85. P. 530–539.
Mordukhai-Boltovskoi Ph.D., 1964. Caspian fauna beyond the Caspian Sea // International Revue der Gesamten Hydrobiologie. V. 49. P. 139–176.
Pallas P.S., 1814. Zoographia rosso-asiatica, sistens omnium animalium in extenso Imperio Rossico et adjacentibus maribus observatorum recensionem, domicilia, mores et descriptiones anatomen atque icones plurimorum. V. 3. Petropoli: Academia Scientiarum. 428 s.
Palo J.U., Väinölä R., 2006. The enigma of the landlocked Baikal and Caspian seals addressed through phylogeny of phocine mitochondrial sequences // Biological Journal of the Linnean Society. V. 88. P. 61–72.
Panov V.E., Dgebuadze Yu.Yu., Shiganova T.A., Filippov A.A., Minchin D., 2007. A risk assessment of biological invasions in the inland waterways of Europe: the Northern invasion corridor case study // Biological invaders in inland waters: Profiles, distribution, and threats. F. Gherardi, Ed. New York etc.: Springer. P. 639–656.
Pobedintseva M.A., Makunin A.I., Kichigin I.G., Kulemzina A.I., Serdyukova N.A., Romanenko S.A. et al., 2019. Population genetic structure and phylogeography of sterlet (Acipenser ruthenus, Acipenseridae) in the Ob and Yenisei river basins // Mitochondrial DNA Part A. V. 30. P. 156–164.
Ponomareva E., Volkov A., Ponomareva M., Makeenko G., Shubina E., 2019. European grayling (Thymallus thymallus) mtDNA control region haplotypes diversity and postglacial colonization of Russian European North // XVI European Congress of Ichthyology. September 2–6, 2019, Aquatis Hotel, Aquatis Aquarium, Lausanne, Switzerland. P. 194–198.
Richards K., van Baak C.G.C., Athersuch J., Hoyle T.M., Stoica M., Austin W.E.N. et al., 2018. Palynology and micropalaeontology of the Pliocene–Pleistocene transition in outcrop from the western Caspian Sea, Azerbaijan: Potential links with the Mediterranean, Black Sea and the Arctic Ocean? // Palaeogeography, Palaeoclimatology, Palaeoecology. V. 511. P. 119–143.
Sars G.O., 1927. Notes on the Crustacean Fauna of the Caspian Sea // Сборник в честь профессора Николая Михайловича Книповича (1885-1925). М.: Изд. народного комиссариата земледелия РСФСР. С. 315–329.
Segestrale S.G., 1982. The immigration of glacial relicts into Northern Europe in the light of recent geological research // Fennia. V. 160. P. 303–312.
Sendek D.S., Bochkarev N.A., Zuykova E.I., Politov D.V., Wanzenböck J., Himberg M., Titov S.F. 2017. Signs of introgression of Baikal omul (Coregonus migratorius) or Arctic cisco (C. autumnalis) into European whitefish (C. lavaretus) in the eastern Baltic Sea // Fundamental and Applied Limnology. V. 189. P. 209–225.
Sergeev A.A., 2016. Evolutionary relations and population differentiation of Acipenser gueldenstaedtii Brandt, Acipenser persicus Borodin, and Acipenser baerii Brandt [version 2; referees: 2 approved] // F1000Research. V. 5. P. 1–11.
Skurikhina L.A., Oleinik A.G., Kukhlevsky A.D., Kovpak N.E., Frolov S.V., Sendek D.S., 2018. Phylogeography and demographic history of the Pacific smelt Osmerus dentex inferred from mitochondrial DNA variation // Polar Biology. V. 41. P. 877–896.
Sokolova A.M., Palatov D.M., Itskovich V.B., 2020. Genetic analysis confirms the freshwater origin of the endemic Caspian sponges (Demospongiae, Spongillida, Metschnikowiidae) // ZooKeys. V. 915. P. 1–16.
Suess E., 1901. Das Antlitz der Erde. Prag-Wien-Leipzig: F. Tempsky & G. Freytag. Bd. 3. № 1. S. 1–508.
Svärdson G., 1998. Postglacial dispersal and reticulate evolution of Nordic coregonids // Nordic Journal of Freshwater Research. V. 74. P. 3–32.
Tomilova A.A., Lyubas A.A., Kondakov A.V., Vikhrev I.V., Gofarov M.Y., Kolosova Y.S., Vinarski M.V., Palatov D.M., Bolotov I.N., 2020. Evidence for Plio-Pleistocene Duck Mussel Refugia in the Azov Sea River Basins // Diversity. V. 12. P. 1–14.
Travina O.V., Bespalaya Y.V., Aksenova O.V., Shevchenko A.R., Sokolova S.E., 2019. Infection of Dreissena polymorpha (Bivalvia: Dreissenidae) with Phyllodistomum macrocotyle (Digenea: Gorgoderidae) in the Northern Dvina River Basin, Northern Russia // Biharean Biologist. V. 13. P. 49–51.
Väinölä R., 1995. Origin and recent endemic divergence of a Caspian Mysis species flock with affinities to the “glacial relict” crustaceans in boreal lakes // Evolution. V. 49. P. 1215–1223.
Väinölä R., Vainio J.K., Palo J.U., 2001. Phylogeography of “glacial relict” Gammaracanthus (Crustacea, Amphipoda) from boreal lakes and the Caspian and White seas // Canadian Journal of Fisheries Aquatic Sciences. V. 58. P. 2247–2257.
Van Houdt J.K.J., De Cleyn L., Perretti A., Volckaert F.A.M., 2005. A mitogenic view on the evolutionary history of the Holarctic freshwater gadoid, burbot (Lota lota) // Molecular Ecology. V. 14. P. 2445–2457.
Vandendorpe J., van Baak C.G.C., Stelbrink B., Delicado D., Albrecht C., Wilke T., 2019. Historical faunal exchange between the Pontocaspian Basin and North America // Ecology & Evolution. V. 9. P. 10 816–10 827.
Vekhoff N.V., 1994. Waterlice from the extra-tundra areas of Siberia and the Far East of Russia, with notes on systematics and zoogeography (Crustacea, Isopoda, Asellidae) // Arthropoda Selecta. V. 3. P. 21–31.
Voroshilova I.S., Artamonova V.S., Yakovlev V.N., 2011. The Origin of populations of Dreissena polymorpha near the North-Eastern boundary of its distribution area // Mussels: Anatomy, Habitat and Environmental Impact. Ed. L.E. McGevin. New York: Nova Science Publishers, Inc. P. 453–468.
Wang Y., Guo R., Li H., Zhang X., Du J., Song Z., 2011. The complete mitochondrial genome of the Sichuan taimen (Hucho bleekeri): Repetitive sequences in the control region and phylogenetic implications for Salmonidae // Marine Genomics. V. 4. P. 221–228.
Weider L.J., Hobæk A., 2000. Phylogeography and arctic biodiversity: a review // Annales Zoologici Fennici. V. 37. P. 217–231.
Wesselingh F.P., Neubauer T.A., Anistratenko V.V., Vinarski M.V., Yanina T., ter Poorten J.J. et al., 2019. Mollusc species from the Pontocaspian region – an expert opinion list // ZooKeys. V. 827. P. 31–124.
Дополнительные материалы отсутствуют.
Инструменты
Зоологический журнал