

Зоологический журнал, 2020, T. 99, № 11, стр. 1298-1306
Видоспецифичность строения фронтальной части черепа у соболя (Martes zibellina) и куницы лесной (Martes martes)
В. Г. Монахов *
Институт экологии растений и животных Уральского отделения РАН
620144 Екатеринбург, Россия
* E-mail: mon@ipae.uran.ru
Поступила в редакцию 28.07.2019
После доработки 06.11.2019
Принята к публикации 16.01.2020
Аннотация
Проведен сравнительный анализ трех краниометрических признаков, характеризующих соотношение длины и ширины во фронтальной части черепа у соболя и куницы лесной. Впервые предложен признак Δ (дельта) – расстояние от посторбитального сужения до линии между заглазничными отростками в сагиттальной плоскости. При анализе обучающей (n = 431) и тестовой выборок (n = 154) выявлена высокая дискриминирующая способность данного признака для определения видовой принадлежности краниологического материала. Сопряженный анализ варьирования данного признака с шириной заглазничного сужения впервые выявил видовую специфику их взаимного соотношения, т.е. наличие противоположных трендов в онтогенезе двух видов. Дискриминатный анализ показал высокий (более 97%) уровень правильных классификаций видовой принадлежности на основе предложенного признака.
Соболь (Martes zibellina L.) и куница лесная (Martes martes L.) довольно надежно могут быть различимы по внешним признакам – цвету меха, его шелковистости, седине, горловому пятну, длине хвоста и т.д. (Огнев, 1931; Новиков, 1956; Громов и др., 1963; Anderson, 1970; Аристов, Барышников, 2001; Monakhov, 2011). Значительно сложнее определить видовую принадлежность зверька по тушке без шкурки, еще сложнее – по черепу, особенно в районах, где два вида обитают совместно. Вследствие высокой индивидуальной изменчивости встречаются зверьки с промежуточными внешними признаками, не характерными для вида (Юргенсон, 1947; Павлинин, 1963; Рожнов и др., 2013), в том числе и заложенными в определительные таблицы в упомянутых выше публикациях.
Знание видовой принадлежности животных необходимо не только для научных целей (зоологических, биогеографических, палеонтологических), но и для охотничье-промыслового менеджмента. Потребительная стоимость этих видов, определяемая рыночной конъюнктурой, не одинакова, что хорошо видно по итогам ежегодных аукционных продаж шкурок соболя (Синицын, 2018) и куницы (http://www.sojuzpushnina.ru).
В процессе изучения морфологии видов рода Martes мы обратили внимание на очевидную сложность определения вида по краниальным признакам. Наиболее популярным для этой цели считается применение данных о расположении и величине слуховых барабанов (Огнев, 1931; Бобринский, и др., 1944; Новиков, 1956). Однако на практике исследователи отмечали трудности при работе с данной конструкцией (Павлинин, 1963). Не случайно предложены модернизации разделения видов как на базе bullae ossae (Гашев, Агешин 2003); так и с использованием одонтологических признаков (Гимранов, 2015; Gimranov, Kosintsev, 2015) и признаков нижней челюсти (Паавер, 1965; Гасилин, Косинцев, 2013). Но эти усовершенствования не дают возможности определять виды рода Martes даже с 95%-ной вероятностью. По утверждению Гасилина и Косинцева (2013, с. 227), проводивших тесты на примере 4 видов, “различия между M. zibellina и M. martes во всех группах (признаков – В.М.) наименее выражены”. На значительное морфологическое сходство этих видов обращали внимание и раньше (Юргенсон, 1947; Hagmeier, 1961; Anderson, 1970; Гусев, 1971; Сlark et al., 1987).
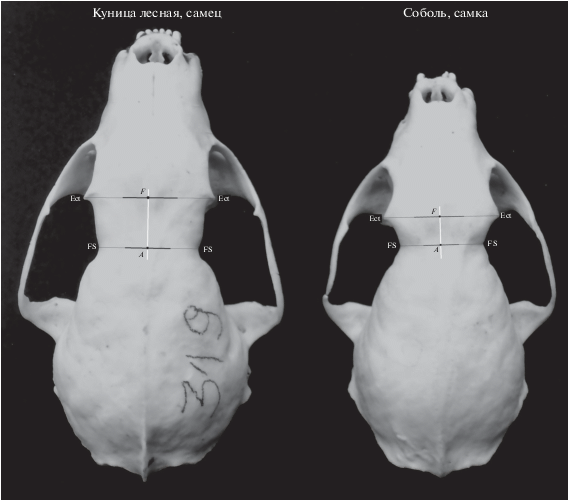
Опыт краниометрической работы с соболем и куницей лесной позволил нам предложить новый способ дифференциации двух видов на основе особенностей строения фронтальной поверхности лицевого отдела черепа (рис. 1). На одну особенность в этой области черепа (латеральные вздутия у куниц между заглазничными отростками и заглазничным сужением) обращали внимание Юргенсон (1947) и Павлинин (1963). Задачей данной работы было нахождение признака или группы признаков, отражающих замеченные различия между видами в морфологии лицевой части черепа. При этом приоритет отдавался возможности формальной (метрической) оценки дискриминирующего признака, который можно было бы анализировать методами основной и многомерной статистики.
МАТЕРИАЛ И МЕТОДЫ
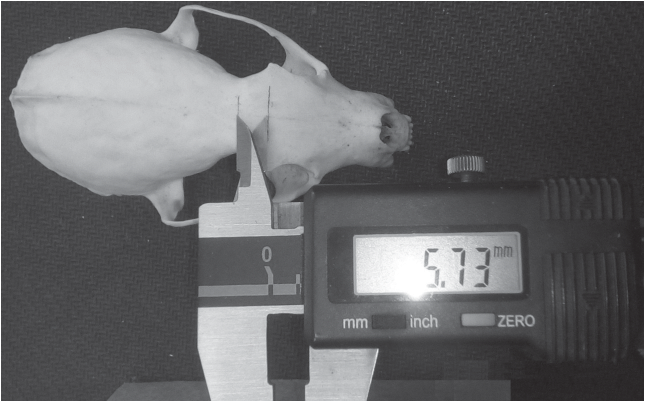
Для поиска формальных межвидовых различий мы изучили комплексный признак, названный нами “дельта” (Δ, рис. 1). Данный промер выполняется штангенциркулем по линии между точками A и F (см. рис. 1). Точка F (frontal midpoint) – расположена на середине линии, соединяющей вершины заглазничных отростков лобной кости между точками Ectorbitale-Ectorbitale (Ect-Ect; Duerst, 1926). Точка А расположена на середине линии между точками Frontostenion-Frontostenion (Fs-Fs; Duerst, 1926), т.е. в центре заглазничного сужения. Измерение делается в сагиттальной плоскости подобно тому, как показано на рис. 2.
Для изучения проявления признака Δ были взяты две краниологические выборки из районов ареалов, где виды обитают “в чистоте”: из окрестностей Баргузинского заповедника (соболь, 217 экз., в т.ч. 98 самцов; добыты в 1983–1989 и 1991–1992 гг.) и Мелеузовского района Башкортостана (куница лесная, 214 экз., в т.ч. 108 самцов; сезон 2012/2013 гг.). Обе коллекции хранятся в музее ИЭРиЖ (Екатеринбург). Данные видовые выборки составили общую обучающую выборку (n = 431). Измерения произведены электронным штангенциркулем с точностью до 0.1 мм. Возраст животных определен по развитию сагиттального и затылочного гребней на лобной и теменной костях черепа (Тимофеев, Надеев, 1955). При помощи данного метода все животные были разделены на сеголетков (до 1 года) и взрослых (старше года). Вместе с признаком Δ измеряли заглазничную ширину (ЗШ) и длину лицевой части черепа (ЛД, по методу: Тимофеев, Надеев, 1955). Предполагалось, что эти признаки, также могут проявлять видоспецифичность.
Для проверки действенности предлагаемого метода изучены тестовые выборки двух видов, которые взяты из тех же районов, что и основные, вошедшие в обучающую выборку. Так, тестовая выборка соболя из окрестностей Баргузинского заповедника состояла из 73 зверьков, добытых в зимние сезоны 1989/1990 и 1999/2000 гг., в том числе 38 самок. Тестовая выборка куницы лесной из Мелеузовского р-на Башкортостана составлена 40 самцами и 41 самкой, добытых в 2011–2013 гг.
Полученные значения измерений были занесены в электронную базу данных MS Works для вычисления основных статистик. Изменчивость, видоспецифичность и взаимодействие выборочных средних значений параметров, а также статистическую значимость разностей оценивали с помощью дискриминантного и корреляционного анализа, модуля тестов различий в пакете Statistica 6 (StatSoft).
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Результаты измерений соболя и куницы лесной из обучающей выборки приведены в табл. 1, где различия параметров хорошо заметны уже по видовым средним значениям (колонка “вид” табл. 1; p < 0.0001). Средние значения изученных параметров и показатели вариационной статистики для каждого вида с учетом пола и возраста (две группы – сеголетки и взрослые, старше 1 года) также приведены в табл. 1. Все изученные признаки демонстрируют половую изменчивость – отчетливый половой диморфизм (p < 0.0001).
Таблица 1.
Средние значения (мм) изученных краниометрических признаков и ДКФ для видов и возрастно-половых групп соболя и куницы лесной в обучающей выборке
| Признак, показатель |
Всего | Молодые | Взрослые | ||
|---|---|---|---|---|---|
| самцы | самки | самцы | самки | ||
| Соболь | |||||
| Колич. особей в выборке | 217 | 51 | 77 | 47 | 42 |
| Δ | 4.67 ± 0.07 (2.5–8.3) |
4.97 ± 0.08 (4.0–6.1) |
3.86 ± 0.07 (2.5–5.3) |
5.80 ± 0.12 (4.0–8.3) |
4.53 ± 0.07 (3.0–5.4) |
| ЗШ | 15.75 ± 0.09 (12.4–18.6) |
16.58 ± 0.10 (15.1–18.3) |
16.27 ± 0.11 (13.6–18.6) |
15.06 ± 0.15 (12.7–17.9) |
14.56 ± 0.15 (12.4–17.3) |
| ЛД | 32.98 ± 0.13 (24.4–37.4) |
34.56 ± 0.16 (31.7–37.4) |
31.47 ± 0.13 (28.5–34.7) |
34.79 ± 0.15 (32.3–37.3) |
32.09 ± 0.17 (29.8–35.2) |
| Среднее значение ДКФ | –1.797 ± 0.06 (–4.23–0.55) |
–1.13 ± 0.08 (–2.26…–0.32) |
–2.35 ± 0.09 (–4.23…–0.45) |
–1.20 ± 0.10 (–2.63–0.55) |
–2.24 ± 0.09 (–3.38…–0.17) |
| Куница лесная | |||||
| Колич. особей в выборке | 214 | 79 | 85 | 29 | 21 |
| Δ | 7.81 ± 0.09 (5.0–11.3) |
8.59 ± 0.11 (6.5–11.3) |
6.83 ± 0.08 (5.2–8.9) |
8.89 ± 0.18 (6.4–10.8) |
7.34 ± 0.25 (5.0–10.2) |
| ЗШ | 18.24 ± 0.07 (15.9–21.9) |
18.95 ± 0.11 (16.9–21.9) |
18.03 ± 0.08 (16.2–19.6) |
17.92 ± 0.19 (16.2–20.9) |
16.90 ± 0.18 (15.9–18.7) |
| ЛД | 36.17 ± 0.13 (31.7–40.3) |
37.78 ± 0.12 (35.2–40.3) |
34.46 ± 0.12 (31.7–38.4) |
37.45 ± 0.17 (35.7–39.0) |
35.25 ± 0.33 (32.9–39.3) |
| Среднее значение ДКФ | 1.822 ± 0.08 (–1.40–6.41) |
2.48 ± 0.12 (0.36–6.41) |
1.37 ± 0.09 (–0.49–3.79) |
2.05 ± 0.15 (0.33–4.65) |
0.85 ± 0.16 (–1.40–1.92) |
Что касается возрастной изменчивости, то у самцов куницы лесной Δ и ЛД (табл.1) не показали статистически значимых разностей (p > 0.05), у самок же наоборот – все признаки демонстрируют значимые возрастные различия (p < 0.012). У соболя возрастные различия также лучше проявились у самок (p < 0.005) по всем трем признакам. У самцов соболя статистически значимыми возрастные различия оказались для двух признаков – Δ и ЗШ (p < 0.0001).
Для соблюдения методической корректности межвидовые тесты проводили с учетом пола и возраста, между соответствующими внутривидовыми группами (подвыборки репрезентативны: n > 21). Межвидовые различия в разрезе возрастно-половых групп по трем изученным признакам (табл. 1) оказались статистически высоко значимыми (p < 0.0001).
Проведены внутривидовые корреляционные тесты между тремя изученными признаками:
– Δ, ЗШ: r Mz = –0.277, F (1,215) = 17.81, p < < 0.00004, r Mm = 0.247, F(1,212) = 13.83, p < 0.00026;
– Δ, ЛД: r Mz = 0.581, F (1,215) = 109.34, p < 0.00001, r Mm = 0.701, F(1,212) = 204.67, p < 0.00001;
– ЗШ, ЛД: r Mz = 0.057, F (1,215) = 0.690, p = 0.407, r Mm = 0.402, F(1,212) = 40.82, p < 0.00001.
В результате была впервые выявлена видовая специфика в соотношении признаков Δ и ЗШ: значение коэффициента корреляции для соболя –0.277, а для куницы лесной 0.247. Это говорит о различных трендах в онтогенезе видов: у соболя наблюдается увеличение Δ на фоне резкого уменьшения ЗШ, у куницы лесной эти оба тренда менее выражены (табл. 1).
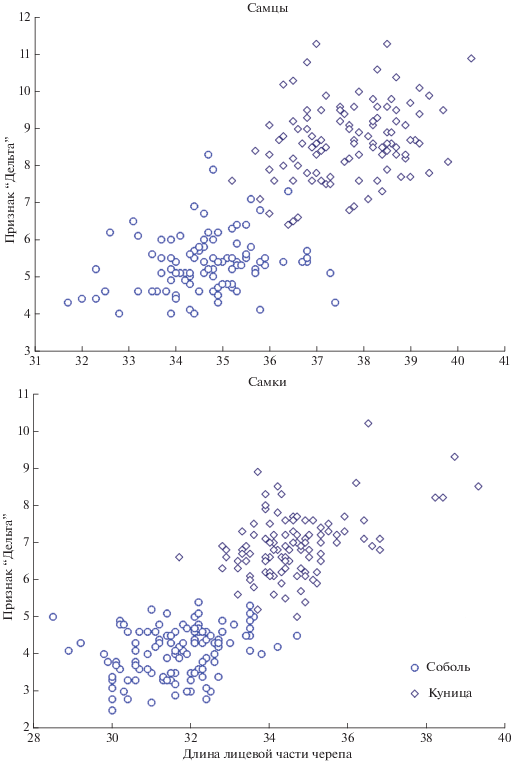
При множественных межвидовых тестах важным показателем является доля перекрываемых значений между совокупностями. Их можно оценить численно по максимальным и минимальным значениям для видов (между соответствующими возрастно-половыми группами), а в графическом виде – по налеганию “облаков” на категоризированной диаграмме рассеивания (рис. 3). В итоге межвидовых сравнений для признака Δ перекрывание оказалось минимальным (0.1 мм, табл. 1) для сеголетков-самок и полностью отсутствовало у самцов-сеголетков. Это свидетельствует о возможности прямого применения (без дополнительных расчетов) предлагаемого признака Δ для видовой дифференциации как минимум сеголетков. Это уже немаловажно, поскольку промысловые пробы состоят главным образом из сеголетков. Как можно видеть на диаграмме рис. 3 особи куниц и соболей одного пола достаточно хорошо разделены.
Наибольшее квалификационное значение для оценки валидности предлагаемого метода мы, вслед за Дзевериным и Лашковой (2006), придаем результатам дискриминантного анализа. Поскольку в исследовании использованы три переменные (для двух видовых выборок), классификация проводится по одной дискриминантной функции (ДКФ), и ее собственное значение для зверька в выборке (подобно значению первой главной компоненты факторного анализа) является интегральным размерным показателем при отнесении этого зверька к одному из двух видов. Основные итоги дискриминантного анализа изученных признаков животных обучающей выборки представлены в табл. 2.
Таблица 2.
Итоги дискриминантного анализа краниометрических признаков соболя и куницы лесной в обучающей выборке
| Признак | Wilks’ Lambda | Partial Lambda | F3,427 | p | Коэффициент дискриминантной функции | Стандартизованный коэффициент для ДКФ |
|---|---|---|---|---|---|---|
| Δ | 0.397 | 0.591 | 296.0 | 0.0000 | –0.966 | –0.855 |
| ЗШ | 0.338 | 0.692 | 189.7 | 0.0000 | –0.655 | –0.557 |
| ЛД | 0.246 | 0.953 | 21.0 | 0.0000 | 0.299 | 0.155 |
| Все признаки | 0.234 | – | 465.0 | 0.0000 | – | – |
Как видно из данных табл. 2, наибольшую (из трех исходных признаков) дискриминирующую виды способность имеет признак Δ, что подтверждает вывод, основанный на анализе выборочных средних. Признак ЗШ не может считаться самостоятельным признаком для дифференциации видов из-за высокой возрастной изменчивости, но все же он вносит определенный вклад в разделение животных, поскольку проявляет своеобразную вариабельность у двух видов.
В результате дискриминантного анализа общая доля правильных классификаций в обучающей выборке составила 97.45% (табл. 3), что подтверждает возможность использования изученного параметра (Δ) для определения видовой принадлежности животных.
Таблица 3.
Результаты классификации животных по итогам дискриминантного анализа краниометрических признаков соболя и куницы лесной в обучающей выборке
| Вид | Доля правильных классификаций, % | Соболь | Куница | Значение ДКФ (для вида) |
|---|---|---|---|---|
| Соболь | 98.62 | 214 | 3 | –1.797 ± 0.060 |
| Куница лесная | 96.26 | 8 | 206 | 1.822 ± 0.075 |
| Всего (n = 431) | 97.45 | 222 | 209 | – |
Мы выяснили, какие именно из 431 животного были ошибочно определены как другой вид. Среди соболей это были три самца, имевшие значения Δ 6.0–8.3 мм, т.е. максимальные показатели для самцов соболя, вследствие чего программа переводила их в разряд другого вида, у которого значения этого признака выше. Наоборот, из числа куниц программа определила соболями исключительно самок с Δ 5.0–7.0 мм – с минимальными значениями для куницы лесной (табл. 1).
При подключении к процедуре дискриминантного анализа тестовой выборки (табл. 4) были получены следующие результаты. Из 73 соболей все они без исключения определены программой как соболи. Среди 81 образца куницы лесной все 40 самцов распознаны как куницы, а из 41 самки правильно отнесены к куницам 34, а остальные 7 самок – к соболю. Общая доля ошибочных определений по совмещенной тестовой выборке (n = 154) оказалась 4.5%.
Таблица 4.
Средние значения (мм) изученных краниометрических признаков для возрастно-половых групп соболя и куницы лесной в тестовой выборке
| Признак | Молодые | Взрослые | ||
|---|---|---|---|---|
| самцы | самки | самцы | самки | |
| Соболь (n = 73) | ||||
| Колич. особей в выборке | 18 | 19 | 17 | 19 |
| Δ | 4.77 ± 0.19 (3.0–6.3) |
3.83 ± 0.11 (2.9–4.7) |
5.56 ± 0.16 (4.2–6.7) |
4.48 ± 0.17 (3.0–6.1) |
| ЗШ | 16.67 ± 0.22 (14.9–18.3) |
16.10 ± 0.22 (14.4–17.5) |
15.05 ± 0.29 (13.1–16.6) |
14.67 ± 0.29 (11.0–17.1) |
| ЛД | 34.97 ± 0.19 (33.5–36.4) |
31.02 ± 0.16 (29.6–33.4) |
35.22 ± 0.15 (34.3–36.6) |
32.04 ± 0.25 (30.4–34.0) |
| Куница лесная (n = 81) | ||||
| Колич. особей в выборке | 23 | 24 | 17 | 17 |
| Δ | 8.29 ± 0.20 (6.9–10.5) |
6.41 ± 0.16 (5.1–8.2) |
8.91 ± 0.26 (6.9–10.7) |
6.60 ± 0.13 (5.7–7.5) |
| ЗШ | 19.25 ± 0.19 (17.9–21.2) |
18.11 ± 0.13 (17.1–19.8) |
18.05 ± 0.16 (16.9–19.9) |
16.91 ± 0.19 (15.2–17.8) |
| ЛД | 37.27 ± 0.23 (35.6–39.3) |
34.10 ± 0.17 (32.4–35.2) |
37.69 ± 0.29 (35.5–40.1) |
34.08 ± 0.22 (32.6–35.7) |
Здесь хотелось бы уточнить, какое место в видовой размерной иерархии занимают каждая из взятых для анализа популяций. Согласно данным, приведенным нами (Монахов, 2015) баргузинский соболь относится к мелким представителям вида с кондилобазальной длиной черепа у самцов 81.12 ± 0.28, самок – 74.28 ± 0.24 мм (Монахов, 2006). Куницы лесные Башкортостана (для характеристики которых была взята выборка из Мелеузовского р-на) также относятся к мелким зверькам с кондилобазальной длиной черепа 81.99 ± 0.18 у самцов и 75.34 ± 0.24 мм у самок (Монахов, 2009). Самыми мелкими являются куницы Пермской и Кировской областей. Таким образом, обе изученные популяции составлены из зверьков с мелким черепом, однако куницы несколько, но статистически значимо, крупнее соболей.
На наш взгляд, можно повысить точность распознавания видов на основе предлагаемого признака. Для этого необходимо предварительно разделить обучающую выборку на самцов и самок и затем провести распознавание с помощью дискриминантной процедуры (отдельно самцов и самок). Как правило, принадлежность к полу – это исходная информация при сборе биоматериала. Однако даже если эта информация отсутствует, то пол зверька определить не составляет труда по причине хорошо выраженного полового диморфизма.
В результате разделения обучающей выборки по признаку пола мы получили из исходного файла две выборки, состоящие из 206 самцов и 225 самок обоих видов. К ним соответственно были добавлены 75 самцов и 79 самок из тестовой выборки двух видов. Для каждой из разделенных по половому признаку новых выборок провели дискриминантный анализ, результаты которого отражены в табл. 5. Значение показателя “частная лямбда Уилкса” для признака Δ по самцам равно 0.564 (F (1,202) = 156.1; p < 0.00001), для самок 0.490 (F (1,221) = 229.9; p < 0.00001), что довольно близко к результату предыдущего дискриминантного анализа (табл. 2).
Таблица 5.
Результаты классификации животных по итогам дискриминантного анализа краниометрических признаков раздельных выборок самцов и самок соболя и куницы лесной
| Выборка по полу | Вид | Доля правильных классификаций, % | Соболь | Куница лесная |
|---|---|---|---|---|
| Обучающая выборка | ||||
| Самцы (n = 206) | Соболь | 100.0 | 98 | 0 |
| Куница | 98.15 | 2 | 106 | |
| Всего | 99.03 | 100(2) | 106(0) | |
| Самки (n = 225) | Соболь | 99.16 | 118 | 1 |
| Куница | 99.06 | 1 | 105 | |
| Всего | 99.11 | 119(1) | 106(1) | |
| Итого (n = 431) | 99.07 | 217(3) | 214(1) | |
| Тестовая выборка | ||||
| Самцы (n = 75) | Соболь | 100.0 | 35 | 0 |
| Куница | 100.0 | 0 | 40 | |
| Всего | 100.0 | 35(0) | 40(0) | |
| Самки (n = 79) | Соболь | 100.0 | 38 | 0 |
| Куница | 97.56 | 1 | 40 | |
| Всего | 98.73 | 39(1) | 40(0) | |
| Итого (n = 154) | 99.35 | 74(1) | 80(0) | |
Итоговые показатели точности определений для обучающей и тестовой выборок оказались улучшены (по сравнению с первичной объединенной обучающей выборкой), соответственно, до 99.07 и 99.35% (табл. 5). Животные, определенные ошибочно в однополых обучающих выборках, имели значения Δ максимальные (самка соболя) либо минимальные (2 самца и самка куницы). В тестовой выборке единственное неправильное определение принадлежало самке куницы со значением Δ 5.7 мм (близкое к минимальному в группе).
Результаты проведенных тестов свидетельствуют о наличии видовой специфики в пропорциях фронтальной части черепа, что в итоге и удалось выявить с помощью предложенного и изученного признака Δ.
ЗАКЛЮЧЕНИЕ
Проведенное сравнительное исследование особенностей строения черепа у соболя и куницы лесной подтвердило видовую специфичность его фронтальной части. Она проявляется в различной величине расстояния от заглазничного сужения до линии между заглазничными отростками в сагиттальной плоскости (признак Δ): среднее значение для соболя 4.67 ± 0.07, для куницы лесной 7.81 ± 0.09 (p < 0.00001). Два модельных вида также демонстрируют различные тенденции в соотношении продольных и поперечных пропорций изученной черепной конструкции в онтогенезе. Это выражается коэффициентами корреляции между величинами признаков Δ и ЗШ: у соболя rΔ-ЗШ = ‑0.277, p < 0.00004; у куницы лесной rΔ-ЗШ = 0.247, p < 0.00026 (разность значима при p < 0.00001). Отмечается, что предлагаемый признак Δ позволяет с высокой долей вероятности определять соболя и куницу лесную по краниологическим выборкам. Точность определений до 97.45% с возможным увеличением до 99% (при дискриминантном анализе однополых выборок).
Задачей дальнейших исследований в этом направлении мы видим как верификацию метода, так и применение его для коллекций из районов близкого или совместного обитания двух видов. Не исключено также нахождение других краниальных (как метрических, так и фенетических) признаков, способных показать высокую результативность межвидовой диагностики.
Список литературы
Аристов А.А., Барышников Г.Ф., 2001. Млекопитающие фауны России и сопредельных территорий. Хищные и ластоногие. СПб.: ЗИН РАН. 560 с.
Бобринский Н.А., Кузнецов Б.А., Кузякин А.П. 1944. Определитель млекопитающих СССР. М.: ГоИз “Советская наука”. 440 с.
Гасилин В.В., Косинцев П.А., 2013. Диагностика видов подрода Martes str. (Carnivora, Mustelidae) по метрическим признакам нижней челюсти // Зоологический журнал. Т. 92. № 2. С. 221–230.
Гашев С.Н., Агешин Н.Н., 2003. К вопросу о дифференциации представителей рода Martes Тюменской области // Вестник Тюменского госуниверситета. Т. 2. С. 15–22.
Гимранов Д.О., 2015. Морфотипическая изменчивость зубов Martes martes и Martes zibellina (Carnivora Mustelidae) // Зоологический журнал. Т. 94. № 5. С. 570–579.
Громов И.М., Гуреев А.А., Новиков Г.А., и др., 1963. Млекопитающие фауны СССР. Ч. 1. М.– Л.: Изд. АН СССР. 640 с.
Гусев О.К., 1971. Восстановление соболя в СССР // Природа. № 11. С. 68–74.
Дзеверин И.И., Лашкова Е.И., 2006. Возможности и ограничения некоторых алгоритмов дискримианантного анализа в идентификации близких видов на примере лесных мышей Sylvaemus (Rodentia, Muridae) // Вестник зоологии. Т. 40 (1). С. 63–69.
Монахов В.Г., 2006. Динамика размерной и фенетической структуры соболя в ареале. Екатеринбург: НИСО УрО РАН, Банк культурной информации. 202 с.
Монахов В.Г., 2009. Особенности размерной структуры популяций лесной куницы (Martes martes Linnaeus, 1758) в ареале // Доклады Академии Наук. Т. 427. № 3. С. 420–422.
Монахов В.Г., 2015. Географическая изменчивость соболя в ареале и филогеография // Экология. № 3. С. 219–228.
Новиков Г.А., 1956. Хищные млекопитающие фауны СССР. М.– Л.: АН СССР. 294 с.
Огнев С.И., 1931. Звери Восточной Европы и Северной Азии. Т. 2. М.–Л.: ГОИЗ. 776 с.
Паавер К.Л., 1965. Формирование териофауны и изменчивость млекопитающих Прибалтики в голоцене. Тарту: АН ЭстССР. 493 с.
Павлинин В.Н., 1963. Тобольский соболь. Свердловск: УФАН СССР. 112 с.
Рожнов В.В., Пищулина С.Л., Мещерский И.Г., Симакин Л.В., 2013. О соотношении фенотипа и генотипа соболя и лесной куницы в зоне симпатрии на северном Урале // Вестник МГУ. Сер. 16. Биол. № 3. С. 23–26.
Синицын А.A., 2018. Динамика численности соболя и эксплуатация его запасов в России за последние 70 лет // Вестник охотоведения. Т. 15. № 3. С. 149–165.
Тимофеев В.В., Надеев В.Н., 1955. Соболь. М.: Заготиздат. 404 с.
Юргенсон П.В., 1947. Кидас – гибрид соболя и куницы // Труды Печоро-Илычского заповедника. Вып. 5. С. 145–178.
Anderson E., 1970. Quaternary evolution of the genus Martes (Carnivora, Mustelidae) // Acta Zoologica Fennica. V. 130. P. 1–132.
Clark T.W., Anderson E., Douglas C., Strickland M., 1987. Martes americana // Mammalian Species. V. 289. P. 1–8.
Duerst U. 1926. Kraniometrie // Handbuch der biologischen Arbeitsmethoden. Wien. Bd. 7, Heft 2. S. 233–342.
Gimranov D., Kosintsev P., 2015. Differentiation of three Martes species (M. martes, M. zibellina, M. foina) by tooth morphotypes // Comptes Rendus Palevol. V. 14. № 8. P. 647–656.
Hagmeier E.M., 1961. Variation and relationships in North American marten // Canadian Field-Naturalist. V. 75. P. 122–138.
Monakhov V.G., 2011. Martes zibellina (Carnivora: Mustelidae) // Mammalian Species. V. 43. Issue 1. P. 75–86.
Дополнительные материалы отсутствуют.
Инструменты
Зоологический журнал