

Зоологический журнал, 2020, T. 99, № 11, стр. 1283-1292
Сравнительный анализ роста и продолжительности жизни офиуры Ophiacantha bidentata Retzius 1805 (Echinodermata, Ophiuroidea) в высоких широтах российской Арктики
Е. А. Стратаненко *
Зоологический институт РАН
199034 Санкт-Петербург, Россия
* E-mail: E.Stratanenko@mail.ru
Поступила в редакцию 01.08.2019
После доработки 30.10.2019
Принята к публикации 15.01.2020
Аннотация
Исследование темпов и характера роста выполнено на примере широко распространенного в Арктике вида офиур Ophiacantha bidentata. Для исследований использовался коллекционный материал лаборатории морских исследований ЗИН РАН. Анализируемые образцы офиуры O. bidentata были взяты из двух регионов российской Арктики – пролива Вихрь (Земля Франца-Иосифа) и пролива Вилькицкого. Для определения индивидуального возраста проводился подсчет кольцеобразных меток роста в кальцитовой структуре лучевых позвонков животного. Каждое видимое кольцо рассматривалось как годовое. В качестве основной математической модели, описывающей рост данного вида, использовалось уравнение Гомперца. Для популяции O. bidentata из пролива Вихрь среднее значение теоретически предельного радиуса луча (R∞) составило 399 ± 28 мкм, экспоненциальное замедление удельной скорости роста (g) – 0.58 ± 0.028. Параметры группового роста несколько отличались от вышеприведенных и составили R∞ = 476 ± 28 мкм, g = 0.39 ± 0.063. Аналогичные расчеты, выполненные для популяции офиур O. bidentata из пролива Вилькицкого, показали, что среднее значение теоретически предельного радиуса луча составило 398 ± 25 мкм, а экспоненциальное замедление удельной скорости роста – 0.39 ± 0.018, параметры группового роста R∞ = 495 ± 23 мкм, g = 0.26 ± 0.028. Максимальная продолжительность жизни O. bidentata в обследованной популяции из пролива Вихрь оценена как 7–10 лет, для популяции из пролива Вилькицкого – 10–15 лет.
Офиуры, или змеехвостки, являются одними из самых часто встречаемых представителей иглокожих и зачастую определяют численность и биомассу бентосного сообщества (Dahm, 1993; Piepenburg, Schmid, 1996; Piepenburg, 2000). Крупные скопления змеехвосток в донных сообществах выполняют важную роль: с одной стороны – регулятора численности мелкого зообентоса (Литвинова, 1984), с другой – при недостатке более калорийной пищи – важного источника питания для разных видов рыб и крабов (Комарова, 1939; Templeman, 1982, 1985; Zamarro, 1992; Чучукало, Напаказов, 1999; Пущина, 2005; Парухина, 2005, Живоглядова, 2005; Чучукало и др., 2011).
Несмотря на достаточно большой интерес к биологии змеехвосток, их обилие и частоту встречаемости в бентосных сборах, многие аспекты экологии, в том числе продукционные возможности и основные закономерности популяционной динамики этой группы организмов, остаются малоизученными, поскольку для подобных исследований необходимы знания о характере и особенностях их роста.
В качестве основной методики определения индивидуального возраста змеехвосток применяется метод подсчета кольцеобразных меток роста, формирующихся в структуре лучевых позвонков животного (Gage, 1990, 1990a). На сегодняшний день получены данные о росте змеехвосток из различных регионов Мирового океана. При этом большая часть публикаций посвящена росту офиур из Антарктики и северной Атлантики (Gage, 1990, 1990a; Dahm, 1993; Dahm, Brey, 1998; Dahm, 1999), в Арктике аналогичные исследования были проведены только для двух видов (Анисимова, 2000; Ravelo et al., 2017). Вместе с тем сравнительный анализ роста одного и того же вида из различных географически удаленных местообитаний не проводился, кроме того, не решена проблема зарастания самых ранних колец роста у особей средних и старших возрастов.
В связи с этим представляется, что исследования параметров роста и продолжительности жизни конкретного вида из разных районов приполярного региона позволят расширить знания о росте и динамике популяций офиур в Арктике.
В данной статье представлены результаты сравнительного анализа параметров роста и продолжительности жизни арктическо-бореальной офиуры Ophiacantha bidentata (R. 1805) из проливов Вихрь (Земля Франца-Иосифа) и Вилькицкого.
МАТЕРИАЛ И МЕТОДИКА
Для исследования роста и продолжительности жизни O. bidentata использовался коллекционный материал Зоологического института Российской Академии наук (ЗИН РАН).
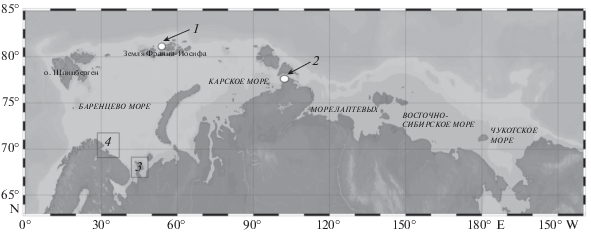
Сбор материала у Земли Франца-Иосифа (ЗФИ) осуществлялся в ходе экспедиция 1936 г. (14 сентября) на судне “Садко” и в проливе Вилькицкого – экспедиция 1993 г. (27 августа) на судне “Polarstern” (рис. 1).
Рис. 1.
Карта района исследований. Цифрами обозначены: 1 – Пролив Вихрь (Земля Франца-Иосифа), 2 – Пролив Вилькицкого, 3 – Северо-Канинское плато, 4 – Мурманское мелководье.
В обоих районах животных отбирали тралом Сигсби с шириной рамы 1 м. Глубина моря в местах отбора проб составляла 50 м (пролив Вихрь – ЗФИ) и 120 м (пролив Вилькицкого). Собранные особи O. bidentata были зафиксированы в 75% спиртовом растворе, занесены в каталог и помещены на хранение в коллекцию лаборатории морских исследований ЗИН РАН (рис. 2). Общее количество O. bidentata, собранных у ЗФИ, составило 208 экз., в проливе Вилькицкого – 91 экз.

В лабораторных условиях у каждого экземпляра из анализируемых проб были измерены диаметр диска и длина лучей. Диаметр диска измерялся с помощью линейки с ценой деления 1 мм, измерение осуществлялось от края диска у основания каждого из лучей до основания противоположного луча (косой диаметр). Длина выпрямленного луча измерялась от его основания у диска до макушки луча.
Для определения индивидуальной массы организмов использовались электронные весы с точностью взвешивания до 0.001 г.
Для анализа роста O. bidentata из каждой пробы были отобраны экземпляры с разными размерами диска. Из сборов экспедиции “Садко” было выбрано 26 офиур, из сборов “Polarstern” – 31.
Определение индивидуального возраста изучаемых офиур выполнялось методом подсчета кольцеобразных ростовых меток на лучевых позвонках животного (Gage, 1990, 1990a; Dahm, 1993; Dahm, Brey, 1998; Dahm, 1999). Для исследования использовались только первые придисковые позвонки. Мягкие ткани и органика удалялись раствором хлорной извести. Далее очищенные и высушенные позвонки подготавливались для работы на сканирующем микроскопе (SEM модель: Quanta-250, maker: FEI Company, Netherlands).
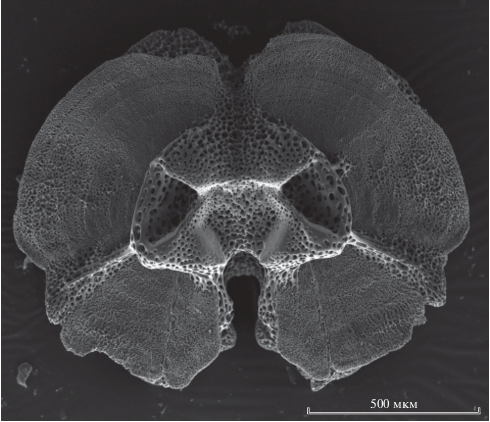
На полученных с помощью сканирующего микроскопа снимках каждого позвонка (рис. 3) измеряли ширину кольцеобразных приростов и по этим величинам рассчитывали параметры уравнений роста Гомперца и Берталанфи.
Рис. 3.
Позвонок офиуры Ophiacantha bidentata R. 1805 из пролива Вилькицкого с видимыми метками роста.
Уравнение Гомперца применялось в следующей форме:
(1)
${{R}_{t}} = {{R}_{\infty }}{{e}^{{\ln ({{{{R}_{0}}} \mathord{\left/ {\vphantom {{{{R}_{0}}} {{{R}_{\infty }}}}} \right. \kern-0em} {{{R}_{\infty }}}}){{e}^{{ - gt}}}}}},$Уравнение Берталанфи применялось в следующей форме:
где Rt – расстояние (мкм) от центра луча до возрастной метки (кольца) в конкретный момент времени t (годы), to – возраст начала роста в соответствии с данным уравнением, R∞ – теоретически предельный радиус луча, e – основание натурального логарифма, k – константа роста. Параметры уравнения подбирались методом Форда-Валфорда для рекуррентных зависимостей Rt + 1 от Rt и методом Берталанфи для зависимостей ln(1 – Rt /R∞) от t.Максимальная продолжительность жизни O. bidentata из анализируемых выборок вычислялась по минимальному значению второй производной для найденных уравнений роста согласно предложению Алимова и Казанцевой (2004).
РЕЗУЛЬТАТЫ
Диаметр дисков исследуемых особей O. bidentata из пролива Вихрь (208 экз.) варьировал от 1 до 17 мм, наибольшая длина выпрямленных неповрежденных лучей составила 47 мм.
В исследуемой популяции O. bidentata (91 экз.) из пролива Вилькицкого размеры дисков животных варьировали от 3 до 15 мм. Максимальная длина лучей превысила 50 мм, составив 56 мм.
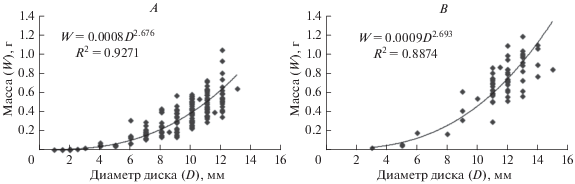
Соотношения массы и размеров офиур в исследуемых популяциях O. bidentatа описывались уравнениями, представленными на рис. 4А–4B.
Рис. 4.
Зависимость массы (W) офиуры Ophiacantha bidentata R. 1805 от диаметра диска (D): в проливе Вихрь (A) и проливе Вилькицкого (B).
Число видимых колец роста, принятых нами за годовые, на поверхности первых лучевых позвонков у анализируемой выборки (26 экз.) из пролива Вихрь (ЗФИ) колебалось от 4 до 8. У большей части офиур с размерами диска от 4 до 12 мм было отмечено по 5–6 видимых меток роста, 4 метки роста наблюдались как у самых маленьких образцов (4 мм), так и у более крупных (10 мм). При этом у самой крупной змеехвостки (17 мм) визуально различимыми на поверхности позвонка были лишь 6 колец (рис. 5A).
Рис. 5.
Зависимость между количеством возрастных меток (N), регистрируемых в структуре позвонка, и диаметром диска (D) офиуры, где A – популяция из пролива Вихрь, B – популяция из пролива Вилькицкого.
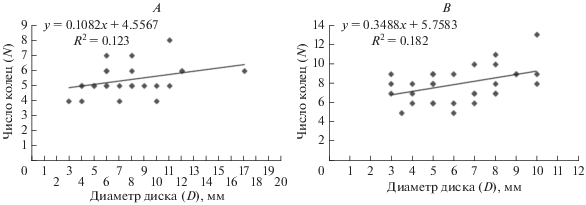
Подсчет меток роста на отснятых с помощью сканирующего микроскопа снимках показал, что индивидуальный возраст O. bidentata (выборка 31 экз.) из пролива Вилькицкого – 5–13 лет. При этом основная доля офиур находилась в возрастной группе 6–9 лет. Как и в предыдущей выборке, анализируемые образцы с разными размерами диска сильно различались по числу видимых колец. Так, 8 видимых кольцевых приростов отмечались как у экземпляров с диаметром диска 3 мм, так и у экземпляров с диаметром диска 10 мм (рис. 5B).
Применение уравнения Берталамфи в качестве математической модели роста O. bidentata позволило подобрать параметры роста для всей выборки из пролива Вихрь (ЗФИ) и только для 30 экз. из пролива Вилькицкого (у одного из образцов наклон линии регрессии Форда-Валфорда превышал 45°). Теоретически предельный размер радиуса луча (R∞) в среднем для 26 экз. из пролива Вихрь составил 498 ± 39 мкм, константа роста (k) – 0.27 ± 0.022, для пролива Вилькицкого данные показатели были следующими: R∞ = 590 ± 45 мкм, k = 0.14 ± 0.006.
Параметры группового роста популяций O. bidentata из исследуемых районов, вычисленные по общей реккурентной регрессии для уравнения Берталанфи, существенно отличались от усредненных и характеризовались следующими значениями: R∞ = 1007 ± 129 мкм, k = 0.07 ± 0.027 (в проливе Вихрь) и R∞ = 1800 ± 331 мкм, k = 0.03 ± ± 0.012 (в проливе Вилькицкого).
Применение уравнения Гомперца позволило подобрать параметры роста для всех исследуемых экземпляров O. bidentata. Среднее значение теоретически предельного радиуса позвонков (R∞) для исследуемой популяции из пролива Вихрь достигало 399 ± 28 мкм, усредненная константа экспоненциального замедления удельной скорости роста (g) – 0.58 ± 0.028. Для выборки офиур из пролива Вилькицкого аналогичные, упомянутым выше, расчеты дали следующие результаты: теоретически предельный радиус луча (R∞) в среднем составил 398 ± 25 мкм, а экспоненциальное замедление удельной скорости роста – 0.39 ± ± 0.018. Вычисленные с помощью уравнения Гомперца параметры группового роста несколько отличались от таковых усредненных, но не столь значительно, как в случае с уравнением Берталанфи – R∞ = 476 ± 28 мкм, g = 0.39 ± 0.063 (для пролива Вихрь) и R∞ = 495 ± 23 мкм, g = 0.26 ± ± 0.028 (для пролива Вилькицкого).
В связи с возникшими сложностями применения уравнения Берталанфи, для дальнейших расчетов максимальной продолжительности жизни O. bidentata в каждом из рассматриваемых районов использовались параметры уравнения Гомперца. Расчеты проводились по усредненным для исследуемой выборки параметрам R∞ и g и отдельно по значениям параметров, вычисленных для группового роста.
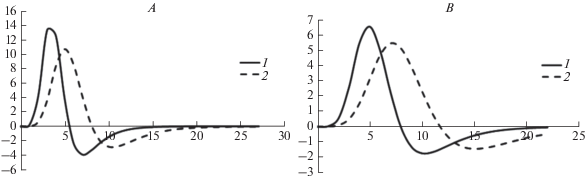
Анализ кривых вторых производных показал, что O. bidentata из популяции в проливе Вихрь (ЗФИ) может доживать до 7–10 лет (рис. 6A), а из популяции в проливе Вилькицкого – до 10–15 лет (рис. 6B).
ОБСУЖДЕНИЕ
Размеры диаметров дисков анализируемых выборок O. bidentata из исследуемых регионов Арктики незначительно разнились между собой. Наиболее крупные экземпляры были характерны для выборки из популяции, обитающей в проливе Вилькицкого. В районе Земли Франца-Иосифа офиуры крупнее 11–12 мм отмечены единично.
Полученные соотношения индивидуальных масс и размеров особей O. bidentatа в исследуемых популяциях в целом не имели существенных различий. Значения показателей степени в данных уравнениях были почти тождественны и составили 2.676 ± 0.152 для района ЗФИ и 2.693 ± 0.236 для пролива Вилькицкого, таким образом, в обоих случаях можно говорить о наличии тенденции к уплощению диска офиур по мере их роста (коэффициент существенно меньше 3).
Результаты подсчета возрастных меток на поверхности лучевых позвонков офиур из пролива Вихрь оказались близки к результатам исследований продолжительности жизни популяции O. bidentata в прибрежных водах Мурмана (Баренцево море), выполненных Анисимовой (2000) на основе метода размерно-возрастного распределения. Согласно представленным в публикации данным, в мурманской популяции змеехвосток было выделено 8 возрастных групп с размерами диска от 3 до 12 мм (Анисимова, 2000). В нашей выборке максимальный возраст анализируемых организмов не превысил 8 лет.
Подсчет визуально различимых меток роста в структуре лучевых позвонков офиур, обитающих в проливе Вилькицкого, показал, что индивидуальный возраст отдельных экземпляров O. bidentata превышает на 2–3 года таковой у офиур из пролива Вихрь. Максимальное число кольцевых приростов отмечено у 10 мм офиуры и составило 13.
Сопоставление размеров дисков (тела животного) и количества видимых меток роста на позвонках O. bidentata из рассматриваемых районов не показывает четкой взаимозависимости, поскольку во многих случаях одно и то же количество возрастных меток соответствовало организмам с разными размерами диска (см. рис. 5A, 5B). С подобной проблемой сталкивались и другие авторы, занимающиеся исследованием роста этих животных (Dahm, 1993; Dahm, Brey, 1998). В своих публикациях они указывают, что данный метод определения индивидуального возраста хорошо работает только для экземпляров, у которых видны все кольца роста, и дает искаженные результаты для особей более старших возрастных групп, так как по мере роста животного и уплотнения кальцитовой структуры первые годовые кольца становятся неотличимыми от общей структуры (Dahm, 1993; Dahm, Brey, 1998). Для решения этой проблемы Dahm & Brey (1998) предлагают вычислять максимально возможный радиус первого кольца роста у самой маленькой особи с дальнейшим вычислением валидного возраста. Следуя данной процедуре, авторы полагают, что можно определить возраст гораздо более точно, чем с помощью подсчета колец на снимке. Однако не всегда в исследуемых выборках присутствуют мелкие особи, и использование данного приема становится невозможным. В нашем случае как в популяции из пролива Вихрь, так и из пролива Вилькицкого наиболее мелкие экземпляры были отмечены единично, в связи с чем подобный анализ не проводился.
В качестве альтернативы применялся математический анализ кривых роста, при использовании которого зафиксированные значения размеров радиуса луча каждого из исследуемых экземпляров накладывались на построенную для него кривую роста. Применение такого рода анализа позволило определить, что радиус начальных меток роста на позвонках большинства офиур приходится на возраст 3–4 года (рис. 7). Подобные результаты представляются вполне адекватными, поскольку совпадают с результатами зарубежных исследователей, изучавших рост других видов офиур в экспериментальных условиях (Dahm, 1998).
Рис. 7.
Пример индивидуальной кривой роста Ophiacantha bidentatа R. 1805, аппроксимирующей уравнение Гомперца, с нанесенными на нее исходными значениями радиусов на позвонке (обозначены точками).

В ходе математического анализа было обнаружено, что уравнение Берталанфи, которое наиболее часто используется для описания роста животных, не совсем точно отображает рост данного вида змеехвосток. Основной его недостаток проявился при расчете параметров группового роста. В обоих случаях (как в проливе в Вихрь, так и в проливе Вилькицкого) теоретическое значение максимального радиуса позвонка, рассчитанное для группового роста, более чем в 2 раза превысило усредненные значения этого параметра, чего быть не должно.
Именно в связи с возникшими при применении уравнения Берталанфи сложностями в качестве альтернативы для построения более адекватной модели роста исследуемых популяций O. bidentata было использовано уравнение Гомперца. В отличие от уравнения Берталанфи, уравнение Гомперца прекрасно работало для всей выборки (26 экз. из пролива Вихрь и 31 экз. из пролива Вилькицкого), полученные параметры группового роста были сопоставимы с усредненными параметрами индивидуального роста (399 ± ± 28 и 476 ± 28 мкм для пролива Вихрь и 398 ± 25 и 495 ± 23 мкм для пролива Вилькицкого, соответственно). В результате мы пришли к выводу о том, что в данном случае рост анализируемых офиур более точно описывает уравнение Гомперца (рис. 8A–8H).
Рис. 8.
Ophiacantha bidentata R. 1805. Пролив Вихрь: A – реккурентная зависимость параметров роста для уравнения Берталанфи (Rt – расстояние (мкм) от центра луча до возрастной метки в конкретный момент времени t); B – реккурентная зависимость параметров роста для уравнения Гомперца (Rt – расстояние (мкм) от центра луча до возрастной метки в конкретный момент времени t); C – функция Берталанфи для группового роста с нанесенным 95% доверительным интервалом (R∞ = 1007 ± 129 мкм, k = 0.07 ± 0.027); D – функция Гомперца для группового роста с нанесенным 95% доверительным интервалом (R∞ = 476 ± 28 мкм, g = 0.39 ± 0.063). Пролив Вилькицкого: E – реккурентная зависимость параметров роста для уравнения Берталанфи (Rt – расстояние (мкм) от центра луча до возрастной метки в конкретный момент времени t); F – реккурентная зависимость параметров роста для уравнения Гомперца (Rt – расстояние (мкм) от центра луча до возрастной метки в конкретный момент времени t); G – функция Берталанфи для группового роста с нанесенным 95% доверительным интервалом (R∞ = 1800 ± 331 мкм, k = 0.03 ± 0.012); H – функция Гомперца для группового роста с нанесенным 95% доверительным интервалом (R∞ = 495 ± 23 мкм, g = 0.26 ± 0.028).
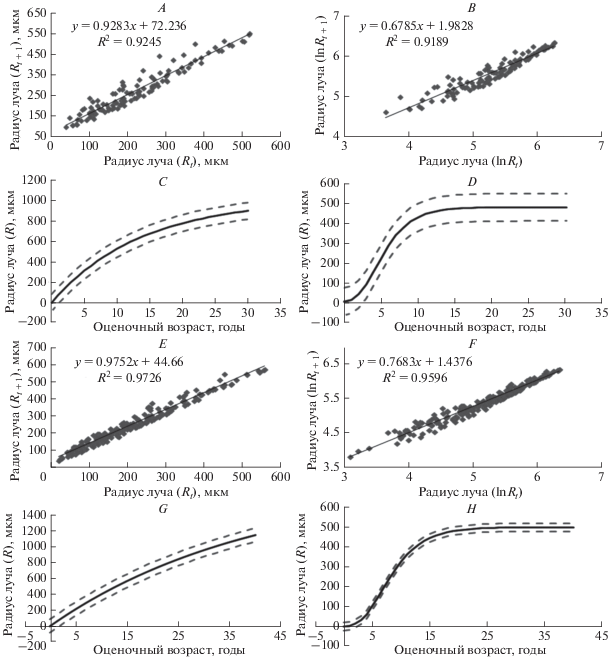
В целом кривые роста, построенные с помощью уравнения Гомперца для исследованных популяций O. bidentata из проливов Вихрь и Вилькицкого, имеют схожую сигмоидную форму. Для большинства из них характерна активная фаза роста до возраста 6–8 лет, которая плавно переходит в фазу замедления (см. рис. 8D, 8H). При этом максимальные значения продолжительности жизни, вычисленные методом Алимова и Казанцевой (2004), в рассматриваемых регионах значительно различались, достигая 7–10 лет в районе ЗФИ и 10–15 лет в проливе Вилькицкого.
Полученные значения усредненных параметров индивидуального и группового роста O. bidentata, обитающих в исследуемых районах российской Арктики, очень хорошо согласуются между собой по величинам теоретически предельного радиуса луча. При этом значения константы роста для поселения из пролива Вилькицкого несколько меньше, чем в проливе Вихрь, где офиуры растут быстрее, но живут меньше.
И действительно, по данным гидрологических наблюдений северное побережье Земли Франца-Иосифа подвержено влиянию более теплых атлантических вод, что в целом сказывается положительно на распространении фауны и ее функциональных характеристиках (Зенкевич, 1963; Голиков, Аверинцев, 1977). Пролив Вилькицкого достаточно удален от зоны активного влияния атлантических вод, и здесь, в большей мере, чем у берегов ЗФИ, проявляются свойства арктических вод. Поэтому обнаруженная в ходе данного исследования тенденция увеличения продолжительности жизни при уменьшении значения константы роста скорее всего обусловлена более низкими среднегодовыми температурами. Схожая зависимость константы роста в уравнениях роста Берталанфи от температурных условий обитания была отмечена в свое время для исландского гребешка (Денисенко, 1989).
Для другого вида офиур (Ophiura sarsii L. 1855) в Чукотском и Баренцевом морях независимо выполненные друг от друга исследования роста с использованием подсчета ростовых меток (Анисимова, 2000; Ravelo et al., 2017) также дали сильно различающиеся результаты по оценкам продолжительности жизни. Оказалось, что эти офиуры в Чукотском море (северо-восточная часть) живут гораздо дольше, чем в Баренцевом море (Северо-Канинское плато и Мурманское мелководье), до 27 и 10–12 лет, соответственно, но баренцевоморские офиуры значительно уступают чукотским по максимальным размерам, 23 мм против 32 мм. Последнее опять же хорошо согласуется с положительной корреляцией константы роста и температуры воды и негативной корреляцией константы роста и теоретически предельных размеров (Денисенко, 1989).
Таким образом, выявленные различия характера роста и продолжительности жизни O. bidentata, собранных в различающихся районах Арктики, по всей видимости, обусловлены в первую очередь климатическими особенностями и доступностью пищи в рассматриваемых районах. Результаты выполненной работы в полной мере согласуются с данными, полученными другими исследователями, а оценки максимальной продолжительности жизни O. bidentata в проливах Вихрь и Вилькицкого укладываются в диапазон колебаний продолжительности жизни, рассчитанной для отдельных видов офиур из Антарктики, северной Атлантики и Пацифики (Gage, 1990, 1990a; Dahm, 1993, 1999; Dahm, Brey, 1998; Quiroga, Sellanes, 2009; Ravelo et al., 2017).
Список литературы
Алимов А.Ф., Казанцева Т.И., 2004. Основные количественные характеристики популяций и связь между ними // Журнал общей биологии. Т. 65. № 1. С. 3–18.
Анисимова Н.А., 2000. Экология массовых видов иглокожих // Современный бентос Баренцева и Карского моря. Апатиты: Изд. КНЦ РАН. С. 275–300.
Голиков А.Н., Аверинцев В.Г., 1977. Биоценозы верхних отделов шельфа архипелага Земли Франца-Иосифа и некоторые закономерности их распределения // Биоценозы шельфа Земли Франца-Иосифа и фауна сопредельных акваторий. Исследования фауны морей. Т. XIV (XXII). С. 5–54.
Денисенко С.Г., 1989. Экология и ресурсы исландского гребешка в Баренцевом море. Апатиты: Изд. КНЦ РАН. 137 с.
Живоглядова Л.А., 2005. Питание равношипого краба Lithodes aequispinus, Benedict северо-западной части Охотского моря // Известия ТИНРО. Т. 143. С. 196–202.
Зенкевич Л.А., 1963. Биология морей СССР. М.: Изд. АН СССР. 739 с.
Комарова И.В., 1939. Питание камбалы-ерша (Hipoglossoides platessoides) в Баренцевом море в связи с кормовыми ресурсами // Труды ВНИРХО. Т. IV. С. 297–320.
Литвинова Н.М., 1984. Питание и поведение шельфовых и глубоководных офиур и их место в трофических сетях. Автореф. дис. … канд. биол. наук. М. 141 с.
Парухина Л.В., 2005. О питании и суточном рационе трески Белого моря Gadus morhua marisalbi Derjugin // Материалы IX международной конференции 11–14 октября 2004 г., г. Петрозаводск. С. 253–257.
Пущина О.И., 2005. Питание и пищевые взаимоотношения массовых видов донных рыб в водах приморья в весенний период // Известия ТИНРО. Т. 2. С. 246–269.
Чучукало В.И., Напаказов В.В., 1999. К методике определения суточных рационов питания и скорости переваривания пищи у хищных и бентосоядных рыб // Известия ТИНРО. Т. 126. С. 160–171.
Чучукало В.И., Надточий В.А., Кобликов В.Н., Борилко О.Ю., 2011. Питание и некоторые черты экологии массовых промысловых видов крабов в водах северо-западной части Японского моря в ранневесенний период // Известия ТИНРО. Т. 166. С. 123–137.
Dahm C., 1993. Growth, production and ecological significance of Ophiura albida and O. ophiura (Echinodermata: Ophiuroidea) in the German Bight // Marine Biology. V. 116 (3). P. 431–437.
Dahm C., 1999. Ophiuroids (Echinodermata) of Southern Chile and the Antarctic: taxonomy, biomass, diet and growth of dominant species // Scientia Marina. V. 63 (1). P. 427–432.
Dahm C., Brey T., 1998. Determination of growth and age of slow growing brittle stars (Echinodermata: Ophiuroidea) from natural growth bands // Journal of the Marine Biological Association of the United Kingdom. V. 78 (3). P. 941–151.
Gage J.D., 1990. Skeletal growth bands in sea brittle stars: microstructure and significance as age markers // Journal of the Marine Biological Association of the United Kingdom. V. 70 (1). P. 209–224.
Gage J.D., 1990 a. Skeletal growth markers in the deep-sea brittle stars Ophiura ljungmani and Ophiomusium lymani // Marine Biology. V. 104 (3). P. 427–435.
Piepenburg D., 2000. Arctic Brittle stars (Echinodermata: Ophiuroidea) // Oceanography and Marine Biology: an Annual review 2000. V. 38. P. 189–256.
Piepenburg D., Schmid M.D., 1996. Brittle star fauna (Echinodermata: Ophiuroidea) of the arctic northwestern Barents sea: composition, abundance, biomass and spatial distribution // Polar Biology. V. 16. P. 383–392.
Quiroga E., Sellanes J., 2009. Growth and size-structure of Stegophiura sp. (Echinodermata: Ophiuroidea) on the continental slope off central Chile: a comparison between cold seep and non-seep sites // Journal of the Marine Biological Association of the United Kingdom. V. 89 (2). P. 421–428.
Ravelo A., Konar B., Bluhm B., Iken K., 2017. Growth and production of the brittle stars Ophiura sarsii and Ophiocten sericeum (Echinodermata: Ophiuroidea) // Continental shelf research. V. 139. P. 9–20.
Sparre P., Venema S., 1998. Introduction to tropical fish stock assessment. Part l. Manual. FAO Fisheries Technical Paper. №.306.1, Rev. 2. Rome FAO. 407 p.
Templeman W., 1982. Stomach Contents of the Thorny Skate, Raja radiata, from the Northwest Atlantic // Journal of Northwest Atlantic Fishery Science. V. 3 (2). P. 123–126.
Templeman W., 1985. Stomach Contents of Atlantic Wolffish (Anarhichas lupus) from the Northwest Atlantic // NAFO Scientific Council Studies. № 8. P. 49–51.
Zamarro J., 1992. Feeding Behavior of the American Plaice (Hippoglossoides platessoides) on the Southern Grand Bank of Newfoundland // Netherlands Journal of Sea Research. V. 29 (1–3). P. 229–238.
Дополнительные материалы отсутствуют.
Инструменты
Зоологический журнал