

Зоологический журнал, 2020, T. 99, № 11, стр. 1268-1282
Новые и малоизвестные свeрчки подсемейства Phalangopsinae (Orthoptera, Gryllidae). 13. Подтриба Phaeophilacridina subtrib. n.
А. В. Горохов *
Зоологический институт РАН
199034 С.-Петербург, Россия
* E-mail: orthopt@zin.ru
Поступила в редакцию 31.07.2019
После доработки 01.10.2019
Принята к публикации 05.11.2019
Аннотация
Из тропической Африки описаны восемь новых надвидовых таксонов трибы Phalangopsini: Phaeophilacridina subtrib. n.; Xenadenacris gen. n.; подроды Arachnacris subgen. n., Bukobacris subgen. n., Taenacris subgen. n., Djacris subgen. n., Hymenacris subgen. n. и Korupacris subgen. n. в составе рода Phaeophilacris. Три бывших рода восстановлены из синонимов Phaeophilacris также в качестве подродов этого рода: Сavernacris stat. n., Guasacris stat. n. и Pholeogryllus stat. n. Кроме того, описаны семь новых видов и подвидов: Phaeophilacris (A.) koilophila sp. n.; Ph. (A.) troglophila beroni subsp. n.; Ph. (C.) microps beshkovi subsp. n.; Ph. (K.) korup sp. n.; Ph. (D.) dja sp. n.; Ph. (T.) anikini sp. n.; X. macrorachis sp. n. Кратко рассмотрены Ph. (Speluncacris) ?harzi и Ph. (C.) m. microps из Кении, а также Ph. (B.) bukobiana из Уганды.
Данная публикация продолжает частичную ревизию африканских представителей подсемейства пауковидных сверчков (Phalangopsinae), начатую в этой же серии публикаций (Горохов, 2015, 2015а). В первом из цитированных сообщений рассматривался афромадагаскарский род Paragryllodes Karny из трибы Paragryllini, а также обсуждался вопрос о подтрибальной принадлежности этого рода и трех родов трибы Phalangopsini: Phaeophilacris Walk. из Африки, Phalangacris Bol. и Seychellesia Bol. с Сейшельских о-вов. Во втором из этих сообщений рассматривался род Phaeophilacris – он был условно включен вместе с Phalangacris и Seychellesia в подтрибу Heterogryllina, которая до этого включала лишь американских представителей. Включение в эту подтрибу основывалось на том, что строение гениталий самца у этих родов и у Heterogryllina довольно примитивно по сравнению с таковым у других подтриб трибы Phalangopsini. Однако такое сходство может быть не синапоморфией этих таксонов, а их симплезиоморфией. Кроме того, недавние молекулярные исследования показали явную обособленность Heterogryllina от рода Phaeophilacris (Chintauan-Marquier et al., 2016). Все это приводит к заключению, что для последнего рода и его ближайших родственников необходимо описать особую подтрибу.
Описания этой подтрибы и ряда более низких таксонов в ее составе, а также замечания о распространении и таксономических признаках некоторых старых таксонов представлены в настоящем (тринадцатом) сообщении. Оно основано на материалах из Зоологического института РАН в С.-Петербурге (ЗИН) и из Национального музея естественной истории (National Museum of Natural History) в Софии (NMNHS).
ТАКСОНОМИЧЕСКИЕ ЗАМЕЧАНИЯ И ОПИСАНИЯ
Триба Phalangopsini Blanchard 1845
Подтриба Phaeophilacridina Gorochov subtrib. n.
Типовой род Phaeophilacris Walker 1870; родовое название женского рода.
Диагноз. Длинноногие сверчки среднего и крупного размера с нормальными прыгательными задними ногами, но без тимпанумов на передних голенях и без стридуляционного аппарата в надкрыльях самца. Эпикраниум с довольно узким и длинным рострумом, который угловидно или округло-угловидно выступает вперед в профиль, не шире скапуса и приблизительно такой же длины, а также с заметно или едва расширенной вершиной, более или менее уплощенной спереди; максиллярные пальпы длинные и тонкие, с последним члеником самым длинным (не меньше расстояние от глаза до вершины рострума) и косо обрубленным на вершине (рис. 1, 1–12). Надкрылья у самки отсутствуют, но у самца обычно в разной степени укороченные и всегда с более или менее параллельными продольными жилками или их следами, а также иногда с поперечным жилкованием (рис. 1, 1–5, 9–11, 13); задние крылья отсутствуют у обоих полов или сильно укороченные, лопастинковидные, склеротизованные и полностью спрятаны под надкрыльями в покое (рис. 1, 6 –8, 10–12). Анальная и генитальная пластинки почти без специализаций у обоих полов; все типичные для семейства склериты в гениталиях самца развиты: эпифаллус с отогнутой вверх или вверх и назад передней частью (либо ее боковыми участками), с раздвоенной задней частью и не слит с рамусами; эктопарамеры развиты, но разнообразные по строению и степени слияния с эпифаллусом; эндопарамеры соединены (слиты) один с другим широкой срединной перемычкой, с задними руками, обычно сочлененными с эктопарамерами, и с довольно крупными передними аподемами; рахис хорошо развит, разнообразный по форме и строению; формула от небольшой, слабо удлиненной, полутрубчатой и полумембранозной структуры до массивного и весьма длинного склерита, снабженного крупной аподемой на вершине (рис. 2, 1–17; 3, 1–3, 6–8, 10–20); яйцеклад типичного строения для трибы Phalangopsini (рис. 3, 4, 5, 9).
Рис. 1.
Подтриба Phaeophilacridina, самец: 1 – Phaephilacris (Arachnacris) troglophila beroni subsp. n.; 2 – Ph. (Speluncacris) ?harzi Kalt.; 3 – Ph. (Cavernacris) microps beshkovi subsp. n.; 4 – Ph. (C.) m. microps (Sjöst.); 5, 6 – Ph. (A.) koilophila sp. n.; 7–9 – Xenadenacris macrorachis sp. n.; 10 – Ph. (Korupacris) korup sp. n.; 11 – Ph. (Djacris) dja sp. n.; 12, 13 – Ph. (Taenacris) anikini sp. n. Голова, переднеспинка и надкрылья в состоянии покоя, вид сверху (1–3, 5, 9); то же, но левое надкрылье отведено в сторону (4, 10, 11); голова, переднеспинка и метанотальная железа сверху, надкрылья отведены в стороны (8, 12) и одно из них оторвано (6); то же, но сбоку и с обоими надкрыльями приподнятыми (7); левое надкрылье сверху-сбоку (13).

Рис. 2.
Род Phaeophilacris Walk., самец: 1–3 – Ph. (Arachnacris) troglophila beroni subsp. n.; 4–6 – Ph. (A.) koilophila sp. n.; 7–9 – Ph. (Cavernacris) microps microps (Sjöst.); 10–12 – Ph. (C.) m. beshkovi subsp. n.; 13–16 – Ph. (Speluncacris) ?harzi Kalt.; 17 – Ph. (S.) malawi Gor. Гениталии сверху (1, 4, 7, 10, 13), снизу (2, 5, 8, 11, 14) и сбоку (3, 6, 9, 12, 15), а также их вершинная часть сверху (16, 17).
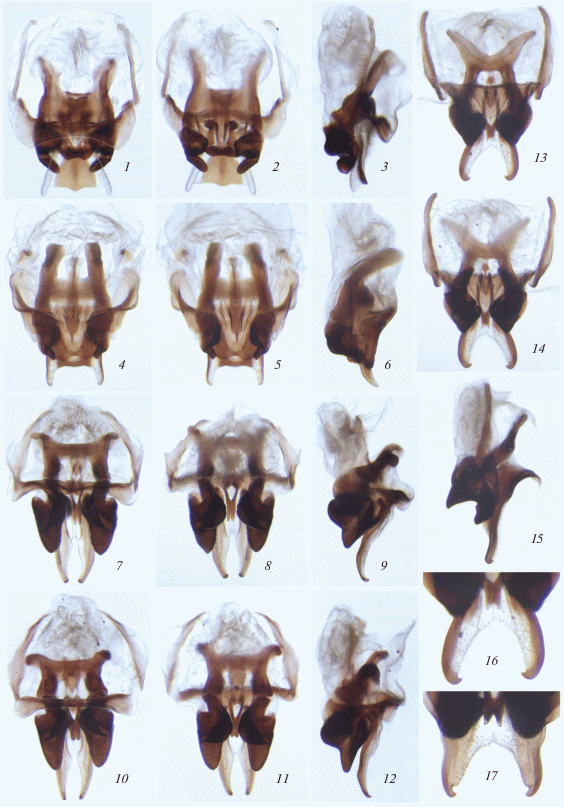
Рис. 3.
Подриба Phaеophilacridina: 1–3 – Phaeophilacris (Bukobacris) bukobiana Rehn; 4 – Ph. (Arachnacris) troglophila beroni subsp. n.; 5 – Ph. (A.) koilophila sp. n.; 6–9 – Ph. (Korupacris) korup sp. n.; 10–12 – Ph. (Djacris) dja sp. n.; 13–15 – Ph. (Taenacris) anikini sp. n.; 16–18 – Xenadenacris macrorachis sp. n.; 19 – Ph. (Hymenacris) townsendi aethiopica Gor.; 20 – Ph. (Guasacris) abyssinica Sauss. Гениталии самца сверху (1, 6, 10, 13, 16), снизу (2, 7, 11, 14, 17) и сбоку (3, 8, 12, 15, 18–20); дистальная часть яйцеклада сбоку (4, 5, 9). [19, 20 – по Горохову (2015а).]
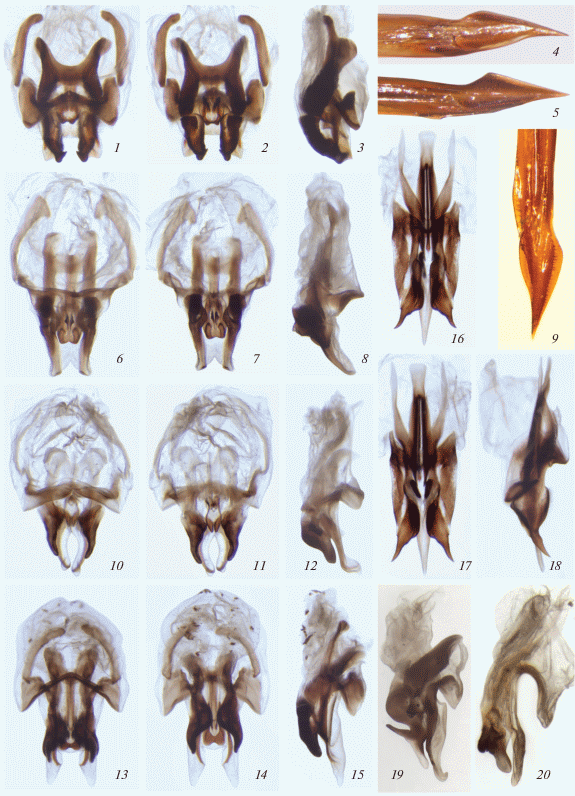
Состав. Два рода: типовой и Xenadenacris gen. n.
Сравнение. От других подтриб Phalangopsini новая подтриба отличается следующим сочетанием признаков: отсутствием тимпанумов у обоих полов и стридуляционного аппарата в надкрыльях самца, сравнительно узким и длинным рострумом головы, наличием хорошо развитого и нормального по строению рахиса в гениталиях самца, а также сравнительно широкой срединной перемычкой, связывающей левый и правый эндопарамеры между собой.
Замечания. В первой половине прошлого века было описано еще несколько “родов” этой подтрибы: Speluncacris Sjöstedt 1910, Opilionacris Sjöstedt 1910, Cavernacris Sjöstedt 1910, Guasacris Sjöstedt 1912, Pholeogryllus Chopard 1923. Но позднее (Chopard, 1968; Kaltenbach, 1983), несмотря на значительное разнообразие в строении гениталий их самцов, все они стали считаться синонимами рода Phaeophilacris. Только недавно было предложено восстановить одно из этих названий (Speluncacris) в качестве подродового (Горохов, 2015а). После изучения дополнительного материала по этой подтрибе стало очевидно, что бывший номинативный подрод этого рода также является очень разнообразным и заслуживающим подразделения на несколько подродов, для которых необходимо восстановить некоторые старые названия и предложить ряд новых. Описания этих подродов и нового рода изучаемой подтрибы даны ниже в определительной таблице родов и подродов Phaeophilacridina (однако гениталии самца для почти половины видов рода Phaeophilacris неизвестны или недостаточно охарактеризованы, поэтому к рассматриваемым подродам здесь отнесены не все 78 описанных видов этого рода).
Определительная таблица родов и подродов Phaeophilacridina
1. Глазки отчетливые; метанотум у самца целиком преобразован в крупную метанотальную железу, состоящую из обширного вздутия и округло-угловидного заднего срединного выступа, между которыми расположена глубокая срединная впадина, захватывающая также большую часть вышеупомянутого выступа (рис. 1, 7, 8); задние крылья у самца присутствуют в виде мелких (но хорошо заметных) склеротизованных лопастинок, полностью спрятанных под надкрыльями в состоянии покоя (рис. 1, 7); передние бедра у самца без вентральных зубчиков. Гениталии самца (рис. 3, 16–18; 4, 13, 14): эпифаллус весьма длинный, глубоко раздвоен сзади (до его передней части), срастается с эктопарамерами в задних (вершинных) частях; рахис очень длинный, выступает позади вершин эпифаллуса и эктопарамеров, полумембранозный; формула также очень длинная (крупная)…………Род Xenadenacris Gorochov gen. n.
[Род включает лишь типовой вид – Xenadenacris macrorachis sp. n.; название рода образовано от латинизированных греческих слов “ξένος” (чужой) и “αδένας” (железа), а также родового названия Phaeophilacris.]
– Глазки неразвиты или неотчетливые; метанотальная железа у самца иного строения или отсутствует (рис. 1, 4, 6, 10–12); задние крылья всегда отсутствуют; передние бедра у самца с заметными вентральными зубчиками. Гениталии самца (рис. 2, 1–17; 3, 1–3, 6–8, 10–15, 19, 20; 4, 1, 3, 4, 6–9, 11, 12): эпифаллус короткий или умеренно длинный, неглубоко раздвоен сзади (до средней части эпифаллуса или еще менее глубоко), не срастается с эктопарамерами или срастается с ними в основании либо по всей длине; рахис короткий или умеренно удлиненный, не выступает позади вершин эпифаллуса и/или эктопарамеров, c более или менее склеротизованными участками; формула короткая или умеренно длинная. [Род Phaeophilacris Walker 1871]………….2
2. Гениталии самца: передняя часть эпифаллуса (или лишь ее боковые участки) отогнута вверх и ясно назад (рис. 2, 1–3, 7–12; 3, 1–3, 10–15, 19, 20), но если почти только вверх, то задняя часть эпифаллуса широко обрублена между довольно короткими, полумембранозными и пальцевидными заднебоковыми лопастями и снабжена срединным склеротизованным участком (рис. 2, 4–6)…3
– Гениталии самца: передняя часть эпифаллуса более или менее отогнута вверх (но не назад); задняя часть эпифаллуса с угловидной или округлой выемкой между его заднебоковыми лопастями либо довольно узко обрублена между ними, а ее срединный участок всегда мембранозный (рис. 2, 13–17; 3, 6–8)…………………………………….11
3. Гениталии самца: задняя часть эпифаллуса более или менее широко обрублена между заднебоковыми лопастями; рахис короткий и широкий, со склеротизованной частью в виде деформированного полукольца, окаймляющего мембрану рахиса сзади и по бокам (рис. 2, 1, 2, 4, 5; 3, 1, 2, 13, 14; 4, 1, 3, 4, 11)……………....……........4
– Гениталии самца: задняя часть эпифаллуса с угловидным или округлым вырезом между заднебоковыми лопастями либо с небольшим округло-угловидным мембранозным выступом между этими лопастями; рахис иной по строению (рис. 2, 7, 8, 10, 11; 3, 10, 11; 4, 6, 12)……………………………....6
4. Гениталии самца: задняя часть эпифаллуса широко обрублена между довольно короткими, полумембранозными и пальцевидными заднебоковыми лопастями и обычно снабжена срединным склеротизованным участком; эктопарамер сравнительно короткий, с небольшой или маленькой вершинной лопастинкой, которая направлена медиально и частично назад и ясно не достигает вершин эпифаллуса (рис. 2, 1, 2, 4, 5; 4, 1, 4)……………………………………Подрод Arachnacris Gorochov subgen. n.
[Типовой вид – Phaeophilacris koilophila sp. n.; подродовое название образовано от латинизированного греческого слова “ ἀράχνη” (паук) и названия рода Phaeophilacris. Состав (кроме типового вида): Ph. troglophila Kaltenbach 1983; возможно, Ph. phalangium Karsch 1892, Ph. leleupi Chopard 1954, Ph. celisi Chopard 1957 и Ph. deheyni Kaltenbach 1986.]
– Гениталии самца: задняя часть эпифаллуса не очень широко обрублена между заднебоковыми лопастями и с мембранозной срединной частью (т.е. без срединного склеротизованного участка); эктопарамер достигает вершин эпифаллуса либо с округлой или постепенно утончающейся дистальной частью (рис. 3, 1, 2, 13, 14; 4, 11)……......5
5. Гениталии самца: эпифаллус с довольно короткими и широкими (почти прямоугольными) заднебоковыми лопастями, лишенными склеротизованных участков; эктопарамер достигает вершин эпифаллуса и с крючковидной вершинной частью (рис. 3, 1–3)……………Подрод Bukobacris Gorochov subgen. n.
[Подрод включает лишь типовой вид – Phaeophilacris bukobiana Rehn 1914; название подрода образовано от видового и родового названий типового вида.]
– Гениталии самца: заднебоковые лопасти эпифаллуса мембранозные, довольно короткие или длинные, но не широкие (пальцевидные) и снабженные узкой склеротизованной ленточкой на каждой лопасти; эктопарамер значительно не достигает вершин эпифаллуса, с округлой или постепенно утончающейся дистальной частью (рис. 3, 13–15; 4, 11)………………Подрод Taenacris Gorochov subgen. n.
[Типовой вид – Phaeophilacris anikini sp. n.; название подрода образовато от латинизированного греческого слова “ταινία [taenia]” (лента) и родового названия Phaeophilacris. Состав (кроме типового вида): Ph. chopardiana Kaltenbach 1983; Ph. tomentosa Kaltenbach 1986; Ph. cycloptera Kaltenbach 1986; возможно, Ph. brevipes Chopard 1957.]
6. Гениталии самца: задняя часть эпифаллуса с небольшой или маленькой округло-угловидной лопастинкой между его заднебоковыми лопастями, а проксимальная часть каждой из последних лопастей с угловидным или округленным медиальным выступом (выпуклостью) около основания (рис. 2, 7, 8, 10, 11; 4, 6)…………………………....7
– Гениталии самца: задняя часть эпифаллуса с угловидной или округлой срединной вырезкой между заднебоковыми эпифаллическими лопастями, а также без заметного медиального выступа (выпуклости) на каждой из последних лопастей около ее основания (рис. 3, 10, 11; 4, 12)……8
7. Гениталии самца: загнутые назад передние части эпифаллуса длинные, равны почти половине эпифаллуса по длине; медиальный выступ, расположенный на каждой из заднебоковых эпифаллических лопастей около ее основания, довольно крупный и отчетливо угловидный (см. Chopard, 1942: fig. 1, 3)……….Подрод Pholeogryllus stat. n.
[Подрод включает лишь типовой вид – Pholeogryllus geertsi Chopard 1923.]
– Гениталии самца: загнутые назад передние части эпифаллуса короткие, значительно короче, чем половина длины эпифаллуса; медиальная выпуклость, расположенная на каждой из заднебоковых эпифаллических лопастей около ее основания, довольно слабая и почти округленная (рис. 2, 7, 8, 10, 11; 4, 6)………Подрод Cavernacris stat. n.
[Подрод включает лишь типовой вид – Cavernacris microps Sjöstedt 1910.]
8. Гениталии самца: дистальная часть эктопарамера не раздвоена или лишь слабо двулопастная (рис. 3, 10–12, 20; 4, 12)…………………………….9
– Гениталии самца: дистальная часть каждого эктопарамера разделена на две сравнительно длинные лопасти (рис. 3, 19; Kaltenbach, 1983: figs 44–47 )………...................................................10
9. Гениталии самца: загнутые назад передние части эпифаллуса короткие, значительно короче, чем половина длины эпифаллуса; дистальная часть эктопарамера не раздвоена (рис. 3, 10–12; 4, 12)…………….Подрод Djacris Gorochov subgen. n.
[Типовой вид – Phaeophilacris dja sp. n.; подродовое название образовано от видового и родового названий его типового вида. Состав (кроме типового вида): возможно, Ph. aroussiensis Kaltenbach 1983.]
– Гениталии самца: загнутые назад передние части эпифаллуса длинные, равны почти половине эпифаллуса по длине; дистальная часть эктопарамера со сравнительно длинной дорсальной и очень короткой вентральной лопастями (рис. 3, 20)……………………Подрод Guasacris stat. n.
[Подрод включает лишь типовой вид – Phaeophyllacris abyssinica Saussure 1878; название Phaeophyllacris является необоснованной поправкой родового названия Phaeophilacris.]
10. Гениталии самца: эктопарамер с глубокой и узкой выемкой, разделяюшей его на две сравнительно длинные и направленные назад лопасти (см. Kaltenbach, 1983: figs 44–47 )…..Подрод Phaeophilacris s. str.
[Типовой вид – Phaeophilacris funesta Walker 1871. Состав (кроме типового вида): Phaeophyllacris martini Bormans 1881; Speluncacris pilipennis Chopard 1934; Phaeophilacris villiersi Chopard 1957; возможно, Ph. minuta Chopard 1957.]
– Гениталии самца: эктопарамер с двумя сравнительно длинными лопастями в дистальной части, направленными в разные стороны: дорсальная лопасть мешковидная, сильно мембранизована и направлена назад; вентральная лопасть сравнительно тонкая, более склеротизованная, отогнута вниз и вперед (рис. 3, 19)…………Подрод Hymenacris Gorochov subgen. n.
[Типовой вид – Phaeophilacris townsendi Kaltenbach 1983; подродовое название образовано от латинизированного греческого слова “ὑμήν [hymen]” (перепонка, мембрана) и родового названия Phaeophilacris. Состав (кроме типового вида): возможно, Ph. abbaica Kaltenbach 1983.]
11. Гениталии самца: заднебоковые лопасти эпифаллуса довольно длинные или умеренно короткие, не более чем в два раза короче срединной части эпифаллуса; эктопарамеры крупные и хорошо склеротизованные, не срастаются с эпифаллусом на всем протяжении, с угловидной или в различной степени раздвоенной вершиной, которая обычно не достигает вершин эпифаллуса; рахис с одной парой удлиненных склеритов, которые в той или иной степени слиты один с другим в дистальной части, и со слегка раздвоенной вершиной (рис. 2, 13–17)……Подрод Speluncacris (=Opilionacris)
[Типовые виды: Speluncacris lucifuga Sjöstedt 1910 и Opilionacris kilimandjarica Sjöstedt 1910 (= Phaeophyllacris spectrum Saussure 1878) соответственно. Состав подрода дан в одном из предыдущих сообщений (Горохов, 2015а: с. 1388), но следует учесть, что Ph. microps, Ph. phalangium and Ph. superba Chopard 1934 теперь удалены из этого подрода, поскольку первые два вида отнесены здесь к подродам Cavernacris и Arachnacris, а третий – недостаточно понятен и представляет, не исключено, еще не описанный подрод.]
– Гениталии самца: заднебоковые лопасти эпифаллуса короткие, примерно в четыре раза короче срединной части эпифаллуса; эктопарамеры более или менее склеротизованные, но на всем протяжении срастаются с боковыми частями эпифаллуса (величина и форма эктопарамеров в связи с этим не очень ясны); рахис весь более или менее склеротизованный, довольно короткий и умеренно широкий, с одной парой вентральных и уплощенных килей у вершины (рис. 3, 6–8; 4, 9)…………………Подрод Korupacris Gorochov subgen. n.
[Подрод включает лишь типовой вид: Phaeophilacris korup sp. n.; название подрода образовано от видового и родового названий типового вида.]
Phaeophilacris (Arachnacris) troglophila beroni Gorochov subsp. n. (рис. 1, 1; 2, 1–3; 3, 4; 4, 1, 2)
Материал. Голотип ♂, Кения, гора “Susua”, пещера № 1, ~2200 м, 18.II.1993, П. Берон и В. Бешков (NMNHS). Паратипы: 1 ♂, 1 ♀, те же данные (ЗИН, NMNHS).
Описание. Самец (голотип). Окраска почти полностью однотонная, светло-коричневая с незначительно более светлыми голенями, лапками, вершиной брюшка и остальными вентральными частями тела. Голова довольно высокая и короткая (рис. 1, 1), с едва вогнутым верхним краем в профиль (середина этой вогнутости расположена над основанием рострума) и с умеренно коротким и широким рострумом, который лишь незначительно ýже скапуса; глаза хорошо развиты, причем высота глаза приблизительно в 1.3 раза больше его ширины; глазки утеряны, но на вершине рострума присутствуют слабые следы срединного глазка; максиллярные пальпы умеренно длинные, с последним члеником самым длинным и приблизительно равным по длине расстоянию от основания скапуса до вершины лабрума. Переднеспинка ясно поперечная, с боковыми лопастями в передней половине округленно-угловидными и оттянутыми вниз и немного в стороны (рис. 1, 1); метанотальная железа отсутствует. Надкрылья слабо укорочены и достигают пятого сегмента брюшка, немного расширены в дистальной половине, широко округлены на вершине, с 6–7 несколько неправильными и радиально расходящимися продольными ветвями в дорсальной плоскости, между которыми имеются беспорядочно расположенные и не очень отчетливые поперечные жилки, и практически без жилкования в латеральной плоскости, которая сравнительно узкая и обособлена от дорсальной плоскости ясным сгибом вдоль продольной жилки (рис. 1, 1). Ноги тонкие и умеренно длинные; переднее бедро с многочисленными очень мелкими зубчиками на нижнем внутреннем киле; передние и средние голени с заметно менее многочисленными тонкими и немного более длинными (щетинковидными) шипиками на нижней поверхности; задняя голень с умеренно многочисленными зубчиками на дорсальной поверхности и 3–4 парами более длинных отчлененных дорсальных шипов в дистальной половине, которые в целом не короче пяти вершинных шпор (но шестая из этих шпор – внутренняя дорсальная – наиболее длинная, достигает средней части заднего базитарзуса); задний базитарзус с 3–4 мелкими дорсальными зубчиками и одной парой более длинных вершинных шпор. Анальная пластинка простая, несколько удлиненно-треугольная и с округленной вершиной; генитальная пластинка более длинная, но почти не выступает сзади из-под анальной пластинки, сравнительно широкая (ее ширина немного меньше ее длины) и с округленно-угловидной вершиной; гениталии сходны с таковыми номинативного подвида, но рахис с широким мембранозным срединным полем около вершины и со слабо расходящимися в стороны передними медиальными выступами его склеротизованной части (рис. 2, 1–3; 4, 1).
Вариации. Второй самец с одной парой крупных темноватых (коричневых) точек по бокам от наличника и с чуть более короткими надкрыльями, достигающими задней части четвертого тергита брюшка.
Самка. Очень похожа на самцов, но окраска как у паратипа, надкрылий нет, а переднее бедро сравнительно короче и без зубчиков. Генитальная пластинка умеренно поперечная, но слегка сужается кзади и с задним краем вогнутым, т.е. с широкой и мелкой выемкой сзади (рис. 4, 2); яйцеклад типичного для рода строения (рис. 3, 4), примерно в 1.2 раза короче заднего бедра.
Рис. 4.
Подтриба Phaeophilacridina (схематично): 1, 2 – Ph. (Arachnacris) troglophila beroni subsp. n.; 3 – Ph. (A.) t. troglophila Kalt.; 4, 5 – Ph. (A.) koilophila sp. n.; 6, 7 – Ph. (Cavernacris) microps beshkovi subsp. n.; 8 – Ph. (C.) m. microps (Sjöst.); 9, 10 – Ph. (Korupacris) korup sp. n.; 11 – Ph. (Taenacris) anikini sp. n.; 12 – Ph. (Djacris) dja sp. n.; 13, 14 – Xenadenacris macrorachis sp. n. Гениталии самца (13) и их задняя часть (1, 4, 6, 9, 11, 12, 14) снизу (1, 4, 6, 9, 11, 12, 13) и сверху (14); рахис снизу (3); генитальная пластинка самки снизу (2, 5, 10); правый эктопарамер снизу и немного сбоку (7, 8). [3 – по: Кальтенбах (Kaltenbach, 1983) c изменениями.]
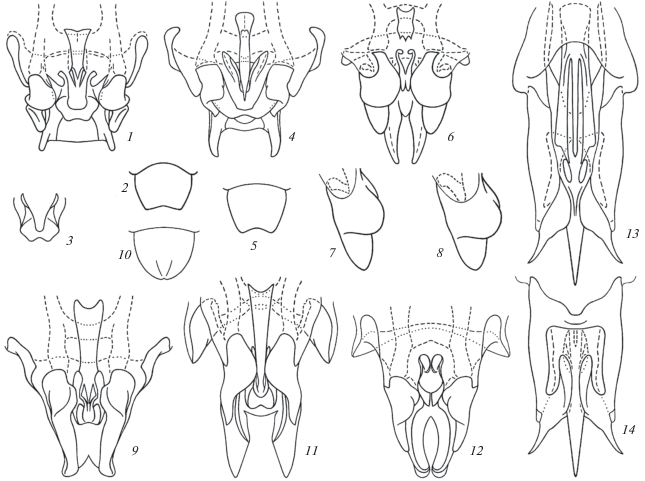
Длина (мм). Тело: самец 12–13, самка 14.5; переднеспинка: самец 2.6–2.7, самка 2.8; надкрылья, самец 6.5–7; передние бедра: самец 9.5–10, самка 8.8; задние бедра: самец 13.8–14.5, самка 14.5; яйцеклад 12.
Сравнение. Новый подвид отличается от Ph. (A.) t. troglophila Kaltenbach 1983 заметно более крупными (более длинными и более широкими) надкрыльями самца с довольно многочисленными продольными ветвями в дорсальной плоскости (6–7 вместо 3–4), а также приведенными выше особенностями строения рахиса в гениталиях самца (сравни рис. 4, 1 и 3). Следует отметить, что типовой местностью номинативного подвида является другой регион Кении (“Nabongo Cave, 12 km WSW Kiminini, Trans-Nzoia Distr., Western Prov.”), но один из его паратипов происходит из пещеры № 14, расположенной на той же горе “Susua” (Kaltenbach, 1983), и, очевидно, принадлежит к новому подвиду. Об этом свидетельствуют промеры, приведенные в этой статье для всех экземпляров Ph. troglophila, – длина накрылий и передних бедер самца сильно варьирует (5.2–7.2 мм и 8.6–12 мм соответственно) по сравнению с длиной задних бедер (13–14 мм), т. е. типовая серия номинативного подвида принадлежит двум разным подвидам или даже двум близким видам.
Этимология. Новый подвид назван в честь одного из сборщиков, болгарского исследователя П. Берона (P. Beron).
Phaeophilacris (Arachnacris) koilophila Gorochov sp. n. (рис. 1, 5, 6; 2, 4–6; 3, 5; 4, 4, 5)
Материал. Голотип ♂, Камерун, Юго-Западный регион недалеко от Нигерии, национальный парк “Korup”, ~300 м, первичный лес, в огромном дупле большого дерева днем, 1–8.II.2016, А. Горохов (ЗИН). Паратипы: 3 ♂♂, 2 ♀♀, те же данные (ЗИН).
Описание. Самец (голотип). Окраска (рис. 1, 5) темно-коричневая с коричневыми передней и боковыми поверхностями эпикраниума, ротовыми частями, ногами, плейритами груди, стернитами и основанием церок, а также с желтоватыми пятнами на месте боковых глазков, одной парой светло-коричневых (почти оранжевых) косых полос на эпикраниуме под глазами и усиками, светло-коричневатой метанотальной железой и затемненными вершинами бедер и основаниями голеней. Тело заметно крупнее, чем у предыдущего вида, рассмотренного здесь. Голова сходная по строению с таковой Ph. (A.) t. beroni, но вершина рострума почти в 1.7 раза ýже скапуса, а относительная длина максиллярных пальп немного больше (их последний членик примерно равен по длине расстоянию между серединой глаза и вершиной лабрума). Переднеспинка также похожа на таковую этого подвида по форме, но сравнительно крупнее (рис. 1, 5); метанотальная железа развита, представляет собой крупное срединное поле на метанотуме, покрытое густыми и сравнительно короткими волосками, но без заметного рельефа (как на рис. 1, 6). Надкрылья достигают середины первого тергита брюшка, овальные, не соприкасающиеся в покое и почти без жилкования (имеются лишь слабые морщинки в дорсальной плоскости и одна продольная жилка в виде низкого киля, отделяющего эту плоскость от узкой латеральной плоскости; рис. 1, 5). Ноги не очень тонкие, но длинные; передняя нога с многочисленными крепкими зубчиками на обоих вентральных килях бедра и столь же многочисленными и крепкими шипиками на вентральной поверхности голени; средние ноги без заметных вентральных зубчиков и шипиков; задние голени с многочисленными крепкими зубчиками на дорсальной поверхности, четырьмя внутренними и четырьмя наружными более крупными дорсальными отчлененными шипами в дистальной половине, а также с немного более длинными в целом вершинными шпорами, из которых дорсальная внутренняя – самая длинная и немного не достигает середины заднего базитарзуса; этот базитарзус с тремя крупными дорсальными зубчиками (два из них предвершинные) и одной парой более крупных шпор. Анальная пластинка удлиненно-трапециевидная, слабо сужается к широкоокругленной (округленно-обрубленной) вершине; генитальная пластинка незначительно длиннее, продолговатая и с более или менее округленной вершиной; гениталии несколько похожи на таковые Ph. troglophila, но заднебоковые выступы эпифаллуса длиннее и более параллельные, задняя срединная часть эпифаллуса значительно менее выступающая позади рахиса, эктопарамеры с более длинными передними частями, рахис более широкий, а формула длиннее и уже (сравни рис. 2, 1–6; 4, 1, 3, 4).
Вариации. Один из паратипов со светло-коричневыми срединной (вертикальной) полоской, тянущейся от вершины рострума до наличника, и передненижними углами боковых лопастей переднеспинки, а также с коричневыми надкрыльями; другой паратип с четырьмя дорсальными зубчиками на заднем базитарзусе (одной парой предвершинных и двумя более проксимальными, но расположенными последовательно).
Самка. Похожа на голотип, но метанотум весь темно-коричневый, надкрылья и метанотальная железа отсутствуют, передние ноги немного короче и лишены зубчиков на бедре, а зубчики на заднем базитарзусе одной из самок – как у последнего из упомянутых самцов-паратипов; генитальная пластинка слабо поперечная (почти квадратная), но заметно сужающаяся кзади и с вогнутым задним краем, т.е. с широкой и мелкой выемкой сзади (короткие выступы по бокам от этой выемки округленные; рис. 4, 5); яйцеклад типичный для рода, приблизительно в 1.4 раза короче заднего бедра, с вершиной – как на рис. 3, 5.
Длина (мм). Тело: самец 21–24, самка 18–21; переднеспинка: самец 4.7–5 , самка 4.1–4.3; надкрылья, самец 6–6.5; передние бедра: самец 16.5–17.5, самка 14–14.5; задние бедра: самец 25–27, самка 23–24; яйцеклад 16–17.
Сравнение. По строению гениталий самца, особенно по форме эктопарамеров, новый вид напоминает Ph. (A.) troglophila, но отличается от него перечисленными выше (в описании) признаками гениталий самца и существенно иным внешним обликом – крупными размерами, темной окраской, относительно короткими надкрыльями самца, а также наличием у самца метанотальной железы. От других видов Arachnacris новый вид отличается крупными размерами, темной окраской, короткими надкрыльями самца, почти лишенными жилкования, и характерными гениталиями самца, особенно формой их эктопарамеров. Два вида (Ph. phalangium Karsch 1892 и Ph. matilei Chopard 1969), известные из Камеруна, внешне похожи на Ph. koilophila, но подродовая принадлежность этих видов не очень ясна или неизвестна, поскольку гениталии их самцов недостаточно изучены; первый из них отличается от нового вида более светлой окраской головы и более короткими надкрыльями (4 мм длиной), а второй – более ясными продольными жилками в дорсальной плоскости надкрылий и более длинным (относительно рамусов) эпифаллусом в гениталиях самца.
Этимология. Название нового вида состоит из двух латинизированных греческих слов “κοίλος” (дупло) и “φίλος” (любитель) в связи с местообитанием этого вида.
Phaeophilacris (Cavernacris) microps microps (Sjöstedt 1910) (рис. 1, 4; 2, 7–9; 4, 8)
Материал. Танзания: 3 ♂♂, “Tanga”, пещера “Amboni”, 18.VIII.2016, Н. Клюге (ЗИН).
Замечания. Этот подвид был описан как вид из “Usambara: Tanga, … Mkulumuzi-Höhlen” (Sjöstedt, 1910: Cavernacris microps), т.е. из местности расположенной недалеко от того места, где были собраны указанные выше самцы. Внешний облик этих самцов в целом соответствует оригинальномуописанию Ph. microps – окраска желтоватая с коричневыми рострумом и дистальными частями скапусов, со светло-коричневыми полосами на переднеспинке, с частично светло-коричневыми или коричневыми надкрыльями, с коричневатыми пятнами на ногах, с коричневыми или почти темно-коричневыми поперечными полосами на задних частях тергитов брюшка (кроме двух–трех самых задних тергитов) и с более или менее сходной окраской заднеспинки (рис. 1, 4). Строение их гениталий также очень сходно с таковым самца из типовой местности, изученного Кальтенбахом (Kaltenbach, 1983), включая глубину переднебоковой вырезки эктопарамера и длину его верхнезаднего выступа (рис. 2, 7–9; 4, 8).
Phaeophilacris (Cavernacris) microps beshkovi Gorochov subsp. n. (рис. 1, 3; 2, 10–12; 4, 6, 7)
Материал. Голотип ♂, Танзания, о-в Занзибар, пещера “Pango Managole”, у дер. “Macunduchi”, 12.IX.1983, П. Берон и В. Бешков (NMNHS).
Описание. Самец. Окраска почти однотонная, светло-коричневая с немного более темными рострумом головы, линией по переднему краю переднеспинки и одной парой расплывчатых пятен вдоль ее заднего края, а также с желтоватыми пятнами на месте боковых глазков. Голова и грудь примерно как у Ph. (A.) t. beroni; однако верхний край головы над основанием рострума почти прямой в профиль, глаза вертикальные (высота глаза примерно вдвое больше его ширины), а максиллярные пальпы заметно более длинные, имеющие последний членик приблизительно равный по длине расстоянию от верхнего края глаза до вершины лабрума. Надкрылья немного более короткие и значительно более узкие (не расширенные в дистальной половине), чем у этого вида, но со сходными по строению вершинной частью и латеральной плоскостью, а также с 5–6 продольными ветвями в дорсальной плоскости, почти лишенной поперечных жилок (рис. 1, 3). Ноги тоже примерно как у этого вида, но более длинные, с редкими щетинками на передней и средней голенях, с многочисленными зубчиками и четырьмя парами дорсальных отчлененных шипов на задней голени, с длиннейшей шпорой этой голени, значительно не достигающей середины заднего базитарзуса, и с последним базитарзусом, имеющим одну пару зубчиков, расположенных около шпор этого базитарзуса. Анальная и генитальная пластинки также похожи на таковые Ph. t. beroni, но гениталии другие – наиболее сходны с гениталиями номинативного подвида и отличаются от них эктопарамерами с менее глубокими переднебоковыми вырезками и с более длинными верхнезадними выступами (сравни рис. 2, 7–12; 4, 6–8).
Самка неизвестна.
Длина (мм). Тело 13; переднеспинка 2.4; надкрылья 6; передние бедра 13; задние бедра 16.5.
Сравнение. От Ph. (C.) m. microps новый подвид отличается заметно более однотонной окраской, особенно на брюшке, и вышеупомянутыми особенностями гениталий самца.
Этимология. Новый подвид назван в честь одного из сборщиков, болгарского исследователя В. Бешкова (V. Beshkov).
Phaeophilacris (Speluncacris) ?harzi Kaltenbach 1983 (рис. 1, 2; 2, 13–16)
Материал. Кения: 1 ♂, пещера “Panga Yambo” между “Qede” и “Watomo”, 13.III.1993, П. Берон и В. Бешков (NMNHS).
Замечания. Определение этого вида, описанного из другой точки в Кении (“Shimoni, Südost-Kenya”), проблематично, поскольку внешне и по гениталиям самца этот вид похож на целую группу видов, самцы которых имеют широкие надкрылья. Однако гениталии самца Ph. harzi недостаточно изучены и не очень понятно изображены автором (Kaltenbach, 1983: fig. 35 ). Более того, гениталии самца у этих видов наиболее хорошо различаются при рассматривании сбоку, но в таком ракурсе они были изображены лишь для некоторых из них (Горохов, 2015а). По форме надкрылий и наличию заметного заднебокового выреза в эктопарамерах изученный самец напоминает Ph. (S.) spectrum Saussure 1878, Ph. (S.) malawi Gorochov 2015 и Ph. (S.) boldyrevi Gorochov 2015. Первый и последний виды характеризуются заметно менее глубоким заднебоковым вырезом эктопарамеров, причем у Ph. spectrum эктопарамер с нижнезадним выступом более выступающим назад, чем его верхнезадний выступ, а Ph. boldyrevi известен только из Малави; Ph. malawi имеет такой же глубокий заднебоковой вырез эктопарамера, как у рассматриваемого самца, но этот вид также известен лишь из Малави, его нижнезадний эктопарамеральный выступ – как у Ph. spectrum, а форма вершинных частей заднебоковых лопастей эпифаллуса совсем иная (сравни рис. 2, 16–17).
Phaeophilacris (Speluncacris) spectrum Saussure 1878
Материал. Танзания: 2 ♂♂, 4 ♀♀, о-в Занзибар, пещера “Pango Managole” у дер. “Macunduchi”, 12.IX.1983, П. Берон и В. Бешков (NMNHS, ЗИН).
Замечания. Этот вид, описанный из Занзибара, вероятно, широко распространен от Мозамбика и Замбии до Танзании и Кении, и, возможно, представлен более чем одним подвидом. Но данные экземпляры почти ничем не отличаются от изученного мной ранее материала из Кении (Горохов, 2015а).
Phaeophilacris (Bukobacris) bukobiana Rehn 1914 (рис. 3, 1–3)
Материал. Уганда: 1 ♂, Центральный регион, район “Mpigi”, национальный парк “Mpanga”, 0°12′38″ с.ш., 32°18′15″ в.д., 1197 м, лес, 25–29.X.2014, В. Аникин (ЗИН).
Замечания. Этот очень короткокрылый вид, описанный из Танзании (Bukoba), был указан также из Заира (Congo Belge) и Уганды (Chopard, 1934; Kaltenbach, 1986). Рассмотренный здесь самец может относиться к новому подвиду, поскольку по сравнению с самцом из типовой местности, изученному мной ранее (Горохов, 2015а), он окрашен немного более пестро, а дорсальная плоскость его надкрылий несколько короче и шире; однако, по гениталиям он почти идентичен самцу из типовой местности.
Phaeophilacris (Korupacris) korup Gorochov sp. n. (рис. 1, 10; 3, 6–9; 4, 9, 10)
Материал. Голотип ♂, Камерун, Юго-Западный регион недалеко от Нигерии, национальный парк “Korup”, ~300 м, первичный лес, среди лесной подстилки ночью, 1–8.II.2016, А. Горохов (ЗИН). Паратипы: 1 ♂, 1 ♀, те же данные (ЗИН).
Описание. Самец (голотип). По внешнему виду похож на Ph. (S.) koilophila, но заметно мельче и слегка светлее: голова светло-коричневая с рыжеватым оттенком, коричневой верхней поверхностью эпикраниума (но дистальная половина рострума темно-коричневая сверху и с желтоватым треугольным пятном на вершине), двумя парами рыже-коричневых полос на передней поверхности эпикраниума (под медиальными половинами усиковых впадин и под глазами), желтоватыми участками на ротовых частях (включая пальпы) и серовато-коричневыми усиками, имеющими почти светло-коричневые основание и нижнюю поверхность скапуса; переднеспинка темно-коричневая, однотонная, но со светловатым маленьким участком в передненижнем углу каждой боковой лопасти; надкрылья интенсивно коричневые (почти темно-коричневые; рис. 1, 10); ноги светло-коричневые с более темными (почти коричневыми) дистальными частями бедер и большей частью задней голени, а также с желтоватыми коксами; среднеспинка светло-коричневая; заднеспинка и тергиты брюшка почти темно-коричневые, но со светлым срединным участком на заднеспинке; стерниты от желтоватых до светло-коричневых; анальная и генитальная пластинки, а также церки серовато-коричневые, но со светлыми основаниями церок. Наружное строение тела очень похоже на таковое Ph. koilophila, но вершинa рострума почти в 1.6 раза ýже скапуса, метанотальной железы нет, надкрылья незначительно менее сужены в дистальной части и имеют 3–4 более заметные продольные жилки в дорсальной плоскости (одна или две их этих жилок двуветвистые; рис. 1, 10), а ноги немного более тонкие и с двумя парами дорсальных зубчиков на заднем базитарзусе (кроме одной пары вершинных шпор). Гениталии (рис. 3, 6–8; 4, 9) существенно отличаются от таковых всех других видов подрода Speluncacris с изученными гениталиями самца тем, что их эктопарамеры полностью слиты с эпифаллусом и выглядят как более склеротизованные нижнебоковые участки эпифаллуса, в профиль заметно расширяющиеся к основанию; эпифаллус удлиненный, с угловидной и не очень глубокой вырезкой на вершине и с передней (непарной) лопастью сравнительно короткой, широкой и изогнутой вверх; рахис короткий и умеренно широкий, со слабой округло-угловидной выемкой на вершине и с парой слегка дуговидных нижних килей, тянущихся от вершины рахиса вперед и имеющих уплощенную нижнюю поверхность с заметным выступом (зубцом) в основании.
Вариации. Другой самец с менее развитыми косыми полосами на передней поверхности эпикраниума, незначительно более короткими надкрыльями, в которых продольное жилкование почти светло-коричневое, а также немного более светлыми (коричневыми) тергитами брюшка и незатемненными дистальными частями бедер.
Самка. Очень похожа на самца-паратипа, но средне- и заднеспинка темные, надкрылий и метанотальной железы нет, а передние ноги относительно короче и без зубчиков на бедрах; генитальная пластинка и яйцеклад также сходны с таковыми самки Ph. koilophila, но первая из них с более узкой дистальной частью и менее заметной вершинной выемкой (рис. 4, 10), а яйцеклад (рис. 3, 9) почти в 1.3 раза короче заднего бедра.
Длина (мм). Тело: самец 15–16, самка 16; переднеспинка: самец 3–3.5, самка 3.3; надкрылья, самец 4–4.6; передние бедра: самец 9.3–11, самка 9.5; задние бедра: самец 18, самка 17; яйцеклад 13.
Сравнение. Новый вид отличается от всех видов подродов Speluncacris и Phaeophilacris, гениталии самца которых изучены, эктопарамерами, полностью слитыми (сросшимися) с эпифаллусом. От похожих видов из Камеруна с недостаточно изученными гениталиями самца, он отличается следующими признаками: от Ph. phalangium – заметно более мелкими размерами тела и наличием заметных продольных жилок в дорсальной плоскости надкрылий; от Ph. matilei – заметно менее длинным (относительно рамусов) эпифаллусом в гениталиях самца.
Этимология. Новый вид назван по названию национального парка (Korup).
Phaeophilacris (Djacris) dja Gorochov sp. n. (рис. 1, 11; 3, 10–12; 4, 12)
Материал. Голотип ♂, Камерун, заповедник “Dja” между Южным и Северным регионами, первичный лес у реки “Dja”, на лесной тропе ночью, 15–22.II. 2016, А. Горохов (ЗИН).
Описание. Самец (голотип). Окраска, размеры и наружное строение тела похожи на таковые Ph. (K.) korup, но имеются следующие отличия: эпикраниум интенсивно коричневый с темно-коричневыми вершиной рострума и дорсальной частью позади основания рострума; скапус однотонно коричневый; надкрылья темно-коричневые; ноги незначительно темнее, чем у этого вида, с крупным темно-коричневым участком в средней трети дорсальной половины заднего бедра и со светло-коричневыми задними голенями; тергиты брюшка серо-коричневые (почти светло-коричневые), но со слабыми затемнениями вдоль задних краев и срединной линии тела; вершина рострума между усиковыми впадинами приблизительно вдвое уже скапуса; надкрылья с тремя заметными продольными жилками в дорсальной плоскости (одна из них двуветвистая; рис. 1, 11); задний базитарзус с шестью дорсальными зубчиками (двумя парами дистальных зубчиков, а также двумя более проксимальными наружными зубчиками). Гениталии ясно отличаются от других видов рода с известными гениталиями самца следующими признаками: эпифаллус с передними частями, отогнутыми вверх и назад (но не только вверх), с заднебоковыми лопастями довольно длинными, пальцевидными, частично мембранозными и снабженными узкой склеротизованной лентой, тянущейся вдоль всей длины этой лопасти по ее вентральной поверхности, а также с угловидным вырезом между этими лопастями; эктопарамеры удлиненные, постепенно сужающиеся к вершине, большей частью не слитые с эпифаллусом и без заметных бугорков на медиальных поверхностях; рахис короткий и не широкий, состоит из пары тонких склеротизованных ленточек, которые формируют одну пару продолговатых утолщений в дистальной части, прижатых одно к другому (но не слитых вместе) и образующих почти заостренную вершину рахиса (рис. 3, 10–12; 4, 12).
Самка неизвестна.
Длина (мм). Тело 16; переднеспинка 3.2; надкрылья 4.3; передние бедра 8.7; задние бедра 14.2.
Сравнение. Новый вид наиболее близок по строению гениталий самца к другому виду того же подрода – Ph. (D.) aroussiensis, но он отчетливо отличается от последнего вида не бугорчатыми медиальными частями эктопарамеров, менее узкой (не почти шиповидной) вершиной рахиса и немного более узкими склеротизованными лентами на заднебоковых лопастях эпифаллуса. От похожих видов из Камеруна новый вид отличается теми же признаками, что и Ph. korup.
Этимология. Новый вид назван по названию заповедника (Dja).
Phaeophilacris (Taenacris) anikini Gorochov sp. n. (рис. 1, 12, 13; 3, 13–15; 4, 11)
Материал. Голотип ♂, Уганда, Центральный регион, район “Mpigi”, национальный парк “Mpanga”, 0°12ʹ38″ с.ш., 32°18ʹ15″ в.д., 1197 м, лес, 25–29.X.2014, В. Аникин (ЗИН).
Описание. Самец (голотип). Размеры тела промежуточные между таковыми Ph. (A.) koilophila и Ph. (D.) dja. Окраска коричневая с немного более светлой рыжевато-коричневой лицевой частью головы, желтоватым пятном на наличнике, светло-коричневатыми остальными участками ротовых частей, темно-коричневыми дорсальной частью эпикраниума (включая рострум) и переднеспинкой (но последняя со светловатым пятнышком в передненижнем углу каждой боковой лопасти), интенсивно коричневыми надкрыльями, имеющими рыжевато-коричневые (незначительно более светлые) жилки, и со светло-коричневыми следующими участками: средне- и заднеспинкой, плейритами, ногами, стернитами и основаниями церок (однако, бедра, голени и лапки с рыжеватым оттенком, а также с легкими затемнениями в вершинных частях всех бедер и с темноватыми косыми штрихами на наружной и верхней поверхностях остальной части заднего бедра). Голова сходна по строению с таковой Ph. dja, но рострум между усиковыми впадинами примерно в полтора раза уже скапуса; грудь также похожа на таковую этого вида, но метанотальная железа хорошо развита, крупная, представляет собой обширную вогнутость, окаймленную довольно высоким килем сзади и по бокам, перегороженную посередине еще одним килем (срединным) и снабженную не многочисленными и довольно короткими волосками в боковых частях этой вогнутости (рис. 1, 12); надкрылья кожистые, значительно укороченные (но длиннее переднеспинки), продолговато-овальные, с 4–5 продольными жилками в дорсальной плоскости (в левом надкрылье одна из этих жилок отчетливо двуветвистая) и беспорядочными короткими жилочками у их дистальных частей, с несколькими почти беспорядочно расположенными жилочками в боковой плоскости, отделенной от дорсальной плоскости заметной килевидной жилкой (рис. 1, 13); ноги по строению напоминают таковые Ph. koilophila, но задний базитарзус с 3–4 заметными дорсальными зубчиками (кроме одной пары вершинных шпор); анальная пластинка треугольная, но с отчетливо обрубленной вершиной; генитальная пластинка немного длиннее анальной, примерно такая же по ширине и с узкозакругленной вершинной частью, самый кончик которой загнут вперед; гениталии с загнутыми назад и довольно длинными передними частями эпифаллуса, с длинными и частично мембранозными заднебоковыми эпифаллическими лопастями (эти лопасти снабжены вентральными склеротизованными лентами, достигающими лишь середины длины лопасти), с эктопарамерами, которые несколько похожи на таковые Ph. dja, и с относительно широким рахисом, имеющим одну пару коротких склеротизованных лопастей (слитых одна с другой) на вершине (рис. 3, 13–15; 4, 11).
Самка неизвестна.
Длина (мм). Тело 20; переднеспинка 4.7; надкрылья 8.8; передние бедра 15; задние бедра 23.
Сравнение. Новый вид отличается от всех других явных видов этого подрода заметно более короткими склеротизованными вентральными лентами заднебоковых эпифаллических лопастей, а также от Ph. chopardiana и Ph. tomentosa – более узким вырезом эпифаллуса между этими лопастями, и от Ph. cycloptera – более длинными эктопарамерами. От Ph. brevipes, который может принадлежать к этому же подроду, новый вид отличается более длинными заднебоковыми эпифаллическими лопастями и изогнутыми (при рассматривании снизу) дистальными половинами эктопарамеров.
Этимология. Новый вид назван в честь сборщика, российского исследователя В. Аникина.
Xenadenacris macrorachis Gorochov sp. n. (рис. 1, 7–9; 3, 16–18; 4, 13, 14)
Материал. Голотип ♂, Камерун, Юго-Западный регион, гора Камерун недалеко от города “Buea”, ~1500 m, первичный лес, среди крупных камней ночью, 9–12.II.2016, А. Горохов (ЗИН). Паратипы: 4 ♂♂, те же данные (ЗИН).
Описание. Самец (голотип). Размеры тела мелкие для подтрибы. Окраска пестрая: голова коричневая с желтыми пятью продольными линиями на задней части темени (эти линии местами соединены одна с другой желтыми участками) и вертикальной полоской на передней части головы (от срединного глазка до середины лабрума), с темно-коричневыми извилистой узкой полоской, тянущейся вдоль верхних краев усиковых впадин и соединяющей все глазки на роструме, и пятном на мембране каждой из этих впадин (над скапусом), с желтоватыми щеками позади глаз и ниже (вдоль заднего края эпикраниума), со светло-коричневыми пятном между глазками, остальными участками ротовых частей (но пальпы и мандибулы чуть темнее, почти коричневые), основанием скапуса и мелкими умеренно редкими пятнышками на жгуте усиков (жгут в дистальной половине между этими пятнышками почти темно-коричневый); переднеспинка темно-коричневая с желтыми пятнами на диске, но передняя часть диска темная и лишь с одной срединной светлой полоской; средне- и заднеспинка коричневые, но последняя с черноватой полоской вдоль заднего края метанотальной железы и с желтоватой впадиной перед ней, снабженной желтыми волосками по бокам (рис. 1, 7–9); надкрылья темно-коричневые, однотонные, блестящие; ноги отчетливо пятнистые, т. е. с коричневыми, светло-коричневыми и желтоватыми участками; плевриты и тергиты брюшка светло-коричневые, но первые с розоватым оттенком и почти однотонные, а вторые с многочисленными темно-коричневыми и коричневыми пятнами; стерниты от светло-коричневых до почти желтоватых, без пятен; анальная пластинка коричневая с тремя светловатыми отметинами; генитальная пластинка и церки серо-коричневые с немного более светлыми основаниями церок. Голова похожа на таковую Phaeophilacris s. l., но глазки мелкие и отчетливые (боковые – по бокам основания рострума, срединный – на его вершине), верхний край головы над основанием рострума прямой в профиль, глаза крупные (заметно больше скапусов и усиковых впадин), рострум между этими впадинами примерно в 1.3 раза ýже скапуса, последний членик максиллярных пальп равен по длине расстоянию от нижнего края глаза до вершины лабрума; переднеспинка умеренно поперечная, с почти прямыми передним и задним краями диска, а также с приподнятой кверху задней половиной нижнего края боковых лопастей (рис. 1, 7–9); метанотальная железа – как на рис. 1, 7 и 8; надкрылья кожистые (но относительно твердые), умеренно укороченные (ясно длиннее переднеспинки), удлиненно-овальные, с медиальным и дистальным краями слегка утолщенными и в положении покоя слабо отогнутыми вверх (но проксимальный участок медиального края без такой специализации), а также с четырьмя немного беспорядочными и килевидными продольными жилками (самая латеральная из них наиболее развита и отделяет дорсальную и латеральную плоскости надкрылья одну от другой; рис. 1, 7, 9); задние крылья в виде узких склеротизованных лопастинок, слегка расширяющихся к вершине и достигающих примерно середины заднеспинки (рис. 1, 7, 8); ноги длинные и тонкие, без вентральных зубчиков на переднем бедре, но с умеренно многочисленными дорсальными зубчиками на задней голени, четырьмя парами не очень длинных отчлененных шипов среди этих зубчиков, шестью также не очень длинными вершинными шпорами этой голени, длиннейшая из которых (средняя внутренняя) достигает середины заднего базитарзуса, и с 11 крохотными дорсальными зубчиками (шипиками) на этом базитарзусе (вдобавок к одной паре небольших вершинных шпор); анальная пластинка похожа на таковую Ph. (T.) anikini, но генитальная пластинка почти вдвое длиннее, а ее вершина узкозакругленная и слегка складчатая; гениталии – как на рис. 3, 16–18 и 4, 13, 14.
Вариации. Один самец немного темнее – с диском переднеспинки почти полностью темным, а у другого самца тергиты брюшка серо-коричневые и почти однотонные; количество зубчиков на заднем базитарзуса также незначительно варьирует.
Самка неизвестна.
Длина (мм). Тело 11–12.5; переднеспинка 2–2.2; надкрылья 3.6–3.9; передние бедра 6.5–7; задние бедра 12–13.
Сравнение. Род содержит лишь один вид, отличия которого от всех других видов Phaeophilacridina даны при описании рода Xenadenacris в определительной таблице надвидовых таксонов этой подтрибы.
Этимология. Название нового вида происходит от морфологического термина рахис, основанного на латинизированном греческом слове “ῥάχις [rachis]” (стержень, хребет), и связано с особо крупной величиной этой структуры в гениталиях самца.
Список литературы
Горохов А.В., 2015. Новые и малоизвестные сверчки подсемейства Phalangopsinae (Orthoptera, Gryllidae). 8. Род Paragryllodes из Африки и ближайших островов // Зоологический журнал. Т. 94. № 8. С. 905–916.
Горохов А.В., 2015а. Новые и малоизвестные сверчки подсемейства Phalangopsinae (Orthoptera, Gryllidae). 9. Род Phaeophilacris // Зоологический журнал. Т. 94. № 12. С. 1387–1399.
Chintauan-Marquier I.C., Legendre F., Hugel S., Robillard T., Grandcolas Ph., Nel A., Zuccon D., Desutter-Grandcolas L., 2016. Laying the foundations of evolutionary and systematic studies in crickets (Insecta, Orthoptera): a multilocus phylogenetic analysis // Cladistics. V. 32. P. 54–81. https://doi.org/10.1111/cla.12114
Chopard L., 1934. Catalogues raisonnés de la Faune Entomologique du Congo Belge. Orthoptères – Gryllides // Annales du Musée du Congo Belge (Zoologie. Série 3. Section 2). T. 4. F. 1. P. 1–88.
Chopard L., 1942. Trois gryllides cavernicoles nouveaux du Congo Belge (Orthoptera) // Revue française d’entomologie. T. 9. P. 51–54.
Chopard L., 1968. Gryllides // Orthopterorum Catalogus. Pars 12. P. 213–500.
Kaltenbach A., 1983. Vorarbeiten für eine Revision der Phalangopsidae der äthiopischen Faunenregion (Saltatoria – Grylloidea). 2. Revision der ostafrikanischen und nordostafrikanischen Art der Gattung Phaeophilacris Walker // Sitzungsberichten der Österreichische Akademie der Wissenschaften Mathematisch-naturwissenschaftlichen Klasse. Abteilung 1. B. 192. H. 5–10. S. 267–306.
Kaltenbach A., 1986. Vorarbeiten für eine Revision der Phalangopsidae der äthiopischen Faunenregion (Saltatoria – Grylloidea). 3. Die zentralafrikanischen Arten der Gattung Phaeophilacris Walker // Sitzungsberichten der Österreichische Akademie der Wissenschaften Mathematisch-naturwissenschaftlichen Klasse. Abteilung 1. B. 195. H. 6–10. S. 201–215.
Дополнительные материалы отсутствуют.
Инструменты
Зоологический журнал