

Зоологический журнал, 2020, T. 99, № 12, стр. 1415-1423
Тестостерон в регуляции различных аспектов поведения и физиологии птиц
М. Я. Горецкая a, *, И. Р. Бёме a
a Биологический факультет, Московский государственный университет имени М.В. Ломоносова
119234 Москва, Россия
* E-mail: m.goretskaia@gmail.com
Поступила в редакцию 02.06.2019
После доработки 07.07.2019
Принята к публикации 13.10.2019
Аннотация
Проанализированы работы, посвященные изучению тестостерона у птиц, за последние 50 лет. Описана роль тестостерона в регуляции различных форм поведения и физиологии птиц. Рассмотрено влияние тестостерона на агрессивное поведение и развитие вторичных половых признаков (окраска многих видов птиц и пение воробьинообразных Passeriformes). Проанализировано воздействие тестостерона на иммунные функции. Обсуждаются функции тестостерона у самок птиц.
Изучение влияния гормонов на поведение птиц ведется давно. Еще в начале 19-го в. была показана корреляция активности пения птиц и размеров семенников (Montagu, 1802 по: Fusani, 2008). Тем не менее, только 50 лет спустя Бертольд (Berthold, 1849 по: Fusani, 2008) осуществил первый эксперимент по поведенческой эндокринологии. Он установил, что у кастрированных молодых петухов не развиваются вторичные половые признаки, такие как поведение, в том числе ухаживание, и красный гребень. Бертольд показал, что имплантация таким петухам семенников от других особей приводит к восстановлению данных признаков, при этом между семенниками донора и организмом реципиента никаких прямых связей не образуется. Таким образом, почти за столетие до того, как были выделены половые гормоны, было показано, что вещества, выделяемые семенниками, – “андрогены” – ответственны за активацию полового поведения у самцов. В дальнейшем количество исследований, посвященных роли тестостерона в жизни птиц, росло, модельными объектами были как одомашненные виды (перепел, куры), так и дикие. В природе большая часть исследований проводилась на воробьинообразных птицах, но имели место исследования и на курообразных, а также на дневных и ночных хищных птицах. Они включали как прямые наблюдения за птицами в природе, так и экспериментальные работы с введением птицам имплантов тестостерона (Fusani, 2008). К настоящему времени собран огромный фактический материал по влиянию тестостерона на поведение и различные аспекты физиологии птиц, который будет по возможности освещен в данной статье.
Тестостерон у птиц вырабатывается из холестерина клетками Лейдига в семенниках, также он может синтезироваться в надпочечниках, яичниках и головном мозге (Hau, Goymann, 2015). Он либо самостоятельно связывается с рецепторами, либо превращается с помощью трех различных энзимов (ферментов) (ароматазы, 5α-редуктазы и 5β-редуктазы) в эстрадиол, 5α-дигидротестостерон и 5β-дигидротестостерон соответственно (Ball et al., 2002; Balthazart, Adkins-Regan, 2002). Дигидротестостерон связывается с рецепторами к тестостерону, причем рецепторы к нему более чувствительны, чем непосредственно к тестостерону, а эстрадиол связывается с рецепторами к эстрогенам.
Тестостерон регулирует различные аспекты поведения птиц. Будучи половым гормоном, он ответствен за формирование агрессивного поведения, поведения ухаживания, развитие вторичных половых признаков, таких как окраска и пение воробьеобразных (DeVoogd, 1991; Cynx, Nottebohm, 1992; Wingfield, 1994; Wingfield, Hahn, 1994; Wingfield et al., 1990, 2000). Кроме того, уровень тестостерона часто положительно связан с физическим состоянием птиц (Duckworth et al. 2004; Chastel et al., 2005; Perez-Rodriguez et al., 2006), социальным статусом (Schoech et al. 1991; Wingfield et al., 1991; Poiani, Fletcher, 1994) и успехом размножения (Raouf et al., 1997; Garamszegi et al., 2005). Также известно, что тестостерон усиливает обмен веществ (Wikelski et al., 1999; Buchanan et al., 2001) и стимулирует предмиграционную гиперфагию (Deviche, 1995; Tonra et al., 2011). Предположительно тестостерон стимулирует гиперфагию не напрямую, а опосредованно, через стимуляцию секреции пролактина (Дьяченко, Дольник, 1984). С другой стороны, тестостерон может снижать степень заботы о потомстве (Wingfield et al., 1990), а также подавлять активность иммунной системы (Folstad, Karter, 1992; Roberts et al., 2004). Исходя из функций тестостерона, многие авторы рассматривают его как гормон, отвечающий за формирование компромисса (trade-offs) в поведении и физиологии птиц, на основании результатов которого самки при выборе полового партнера могут оценить степень приспособленности последнего.
Тестостерон и уровень агрессии
Гипотеза вызова (провокации) (the challenge hypothesis), предложенная Вингфильдом в 1990 г. (Wingfield et al., 1990), предсказывает, что концентрация тестостерона в крови будет увеличиваться в ходе агонистических взаимодействий между самцами. И действительно, у некоторых видов были найдены подтверждения данной гипотезы. У свободноживущих самцов певчей овсянки (Melospiza melodia) уровень тестостерона увеличивался в течение 10–30 мин после имитации территориального вторжения (Wingfield, 1985). Импланты тестостерона повышали интенсивность и настойчивость агонистических взаимодействий у птиц этого вида (Wingfield et al., 1990). Сходные данные были получены на японских перепелах (Coturnix japonica) (Ramenofsky, 1984), красноплечем трупиале (Agelaius phoeniceus) (Harding, Follett, 1979) и на других видах птиц (Vleck, Brown, 1999; Hirschenhauser et al., 2003).
Согласно данной гипотезе, высокий уровень тестостерона невыгоден самцам во время выкармливания птенцов, т.к. при повышенном уровне гормона они нацелены преимущественно на демонстрацию территорий и пение (рис. 1), а не на поиск корма для птенцов. В соответствии с этим, уровень тестостерона будет зависеть от степени родительской заботы, свойственной виду (Hegner, Wingfield, 1987; Ketterson et al., 1992; De Ridder et al., 2000; Peters et al., 2002). Данная гипотеза позволяет также предсказать различия в уровне тестостерона в зависимости от системы спаривания (Wingfield et al., 1990). У моногамных видов уровень тестостерона повышается в начале сезона размножения и снижается до базовой концентрации в период после откладки яиц и выведения птенцов (Logan, Wingfield, 1995; Vleck, Brown, 1999). Напротив, у полигамных видов уровень тестостерона также увеличивается в начале сезона размножения, но не снижается во время инкубации яиц самкой. Полигамные самцы пытаются привлечь второго партнера в течение одного и того же репродуктивного цикла и вынуждены конкурировать друг с другом за новую территорию. В соответствии с гипотезой вызова эта конкуренция может отвечать за стабильно высокий уровень тестостерона.
Рис. 1.
Воздействие тестостерона (жирный шрифт) и возможные затраты на поддержание его высокого уровня (светлый шрифт) в сезон размножения и вне сезона размножения. ГГС – гипоталамо-гипофизарная система (по: Soma, 2006, с изменениями).
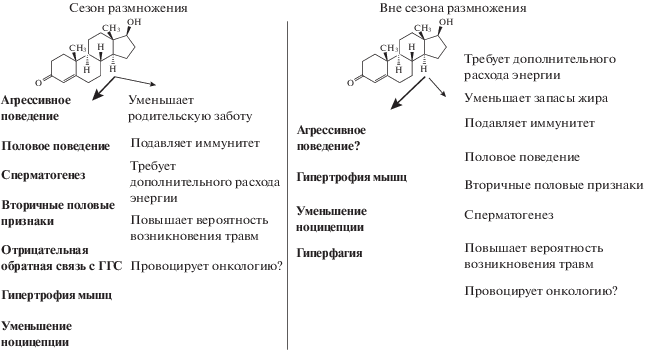
Однако попытки протестировать данную гипотезу на разных видах привели к накоплению противоречивых результатов (Moore et al., 2004). В отличие от певчей овсянки и некоторых других видов, у многих протестированных птиц не выявлено увеличение уровня тестостерона в ответ на симулирование территориального вторжения (Goymann et al., 2007; Goymann, 2009). Кроме того, у видов, продолжающих охранять территорию вне периода размножения, уровень тестостерона повышается при взаимодействиях самцов в период размножения, но остается низким во время взаимодействий вне этого периода (табл. 1) (Soma, 2006; Goymann, 2009). У тропических видов, живущих на одной территории круглогодично, концентрация тестостерона поддерживается на низком уровне в течение всего годового цикла (Hau, 2007).
Таблица 1.
Особенности жизненного цикла четырех видов птиц и влияние тестостерона на некоторые аспекты их поведения
| Показатель | Горихвостка-чернушка | Зарянка | Черноголовый чекан | Певчая овсянка |
|---|---|---|---|---|
| Миграция | Да | Да | Да/Нет | Нет |
| Территориальность | Круглый год | Круглый год | Круглый год | Круглый год |
| Пение | Круглый год | Круглый год | Круглый год | Круглый год |
| Период размножения | 3–4 месяца | 3–4 месяца | 3–4 месяца | 3–4 месяца |
| Рсамец-самец | Нет | Нет | Нет | Да |
| Агрессия в сезон размножения зависит от Т | Нет | Да | Да | Да |
| Агрессия вне сезона размножения зависит от Т | Нет | Нет | Нет | Да |
| Линейная связь между Т и поведением | Нет | Нет | Нет | ? |
Таким неоднозначным результатам были предложены следующие объяснения. Вне сезона размножения, когда гонады редуцированы и концентрация тестостерона в крови низкая, регуляция территориальной агрессии оказывается связанной с выработкой эстрадиола, к которому чувствительны рецепторы в головном мозге птиц (Soma, 2006; Hau, 2007). Эстрадиол синтезируется в головном мозге птиц предположительно из предшественника дегидроэпиандростерона (DHEA, ДГЭА), синтезируемого в надпочечниках. Концентрация дегидроэпиандростерона у певчей овсянки повышена в не репродуктивный период, а введение ДГЭА провоцирует пение во время агрессивных взаимодействий (о пении см. ниже) (Soma, Wingfield, 2001; Soma et al., 2002). Кроме того, эстрадиол может синтезироваться из холестерина непосредственно в головном мозге (Soma 2006). Сходным образом, видимо, регулируется агрессивное поведение у оседлых тропических видов птиц (Hau et al., 2004; Hau, 2007). Таким образом, тестостерон регулирует территориальное поведение круглогодично, но способы его воздействия различаются на разных этапах годового цикла. В период размножения он непосредственно регулирует территориальное поведение, а в осенне-зимний период в головном мозге повышаются число рецепторов и их чувствительность к половым гормонам (в основном эстрадиолу), которые возможно синтезируются из ДГЭА (Hau, 2007).
Тестостерон и иммунокомпетентность
Гипотеза о том, что повышение уровня тестостерона ведет к понижению иммунного статуса (“Handicap hypothesis”) была предложена около 40 лет назад (Folstad, Karter 1992). При этом до сих пор не ясно, как именно связаны половые гормоны с иммунокомпетентностью у птиц (Roberts et al., 2004; Roberts et al., 2007).
В части исследований с проведением экспериментальных манипуляций с уровнем тестостерона показано, что он обладает подавляющим иммунитет эффектом (Buchanan et al., 2003; Casto et al., 2001; Duffy et al., 2000 Owen-Ashley et al., 2004; Peters, 2000), однако другие работы этого не подтверждают (Hasselquist et al., 1999). Кроме того, в некоторых случаях выявлено даже позитивное воздействие тестостерона на иммунитет (Evans et al., 2000). Одним из способов воздействия может являться подавление тестостероном иммунитета через повышение концентрации кортикостерона (Buchanan, 2000; Råberg et al., 1998; Sapolsky et al., 2000). Но и данная гипотеза не была полностью подтверждена, и обнаружен обратный эффект. Так, у зебровых амадин (Taeniopygia guttata) и канареек (Serinus canaria) хорошо выраженный иммунный ответ выявлен на фоне высокого уровня обоих гормонов в крови (Roberts et al., 2007; Горецкая и др., 2018).
Уровень тестостерона у самок
У самок уровень тестостерона, как правило, ниже, чем у самцов, но также варьирует в течение годового цикла. Отмечено, что более высокие концентрации гормона наблюдаются непосредственно перед сезоном размножения или во время него (Elekonich, Wingfeld, 2000; Ketterson et al., 2005). Показана положительная корреляция уровня тестостерона у самцов и самок одного вида (Ketterson et al., 2005; Goymann, Wingfield, 2014). По одним данным, уровень тестостерона выше у самок моногамных видов (Ketterson et al., 2005), однако более позднее исследование этого не подтверждает (Goymann, Wingfield, 2014). У некоторых видов птиц уровень тестостерона самок связан с агрессией в период размножения (например, конкуренция за полового партнера или за территорию), что предполагает потенциальную роль тестостерона в качестве модулятора территориального поведения не только у самцов, но и у самок (Staub, de Beer, 1997; Wingfeld et al., 2000; Ketterson et al., 2005; Sandell, 2007; Rosvall et al., 2013).
Тестостерон и окраска птиц
Согласно принципу гандикапа (Zahavi, 1975; Grafen, 1990) только сильные и здоровые самцы могут позволить себе энергетически затратные элементы брачного поведения. В соответствии с этим предполагается, что в случае полового диморфизма яркая окраска может быть одним из тех качеств самца, которое, с одной стороны, выбирается самкой, а, с другой, как раз и реализует компромисс между возможностью демонстрировать признак и состоянием особи. Исходя из этого, окраска самцов может быть связана с уровнем тестостерона (Folstad, Karter, 1992; Jawor, Breitwisch, 2003; Bokony et al., 2008). Положительная корреляция между яркостью оперения и уровнем тестостерона была найдена для зеленушки (Chloris chloris) (Merila, Sheldon, 1999), черноголового дубоносового кардинала (Pheucticus melanocephalus) (Hill, 1994) и атласного шалашника (Ptilonorhynchus violaceus) (Collis, Borgia, 1992), но не была выявлена у обыкновенной чечетки (Acanthis flammea) (Romero et al., 1997).
У серого юнко (Junco hyemalis) размер белой зоны в окраске хвоста, которая, как было показано ранее, отвечает за привлекательность самца для самки, оказался положительно связан с кратковременным повышением уровня тестостерона, вызванного инъекцией гонадотропного гормона (McGlothlin et al., 2008).
У птиц окраска оперения может формироваться по-разному. Выделяют несколько типов окраски: меланиновая (коричневый, черный, красно-коричневый цвет оперения), липохромная (желтый, оранжевый, красный), определяемая каротиноидами, и структурная, определяемая структурой пера (Olson, Owens, 2005). Считается, что меланиновая окраска в большей степени определяется генетически и в меньшей степени зависит от внешних факторов, в отличие от каротиноидной. Хорошо известным примером связи меланиновой окраски с уровнем тестостерона является размер черного пятна у домового воробья (Passer domesticus) (Evans et al., 2000; Buchanan et al., 2001; Gonzalez et al., 2001). Для мексиканской чечевицы (Carpodacus mexicanus) показана связь каротиноидной окраски с уровнем тестостерона (Duckworth et al., 2004), а для обыкновенной лазоревки (Cyanistes caeruleus) связь тестостерона и структурной окраски (степени отражения ультрафиолетового излучения) (Peters et al., 2006). Также на лазоревке показано, что уровень тестостерона во время линьки может оказывать организующий эффект на окраску самцов в последующий год (Roberts et al., 2009). Тем не менее, не всегда понятно, какие именно механизмы отвечают за влияние тестостерона на проявление окраски оперения (Moller, Eritzoe, 1988; Hill, Brawner, 1998; Bennett, Owens, 2002).
У прекрасного расписного малюра (Malurus сyaneus) и красноспинного расписного малюра (Malurus melanocephalus) уровень тестостерона различается у самцов с самцовым типом окраски и у бледных самцов, окрашенных как самки (Peters et al., 2000; Geslin et al., 2004; Lindsay et al., 2009). Причем у самцов обоих фенотипов повышение тестостерона стимулирует линьку в оперение с окраской самцового типа. Сходная тенденция выявлена и для горихвостки-чернушки (Phoenicurus ochruros), вида, имеющего только одну линьку в году; у самцов, линяющих во взрослое оперение, уровень тестостерона выше, чем у молодых самцов, линяющих в субадультное оперение (Schwarzová et al., 2010).
Уровень тестостерона оказался положительно связан с размером белого пятна на голове мухоловки-белошейки (Ficedula albicollis) (Garamszegi et al., 2004).
У самок тропических птиц уровень тестостерона коррелирует с орнаментом оперения (Enbody et al., 2018).
Регуляция пения
У взрослых певчих птиц процесс обучения пению и само пение регулируются сезонными изменениями уровня тестостерона в крови (Tramontin, Brenowitz, 2000; Ball et al., 2002). Так, у видов с открытым типом обучения (учат песню каждый год) сезонная изменчивость песенного поведения самцов (пение в репродуктивный период и отсутствие пения в осенне-зимний период) связана с сезонными изменениями уровня тестостерона (Nottebohm et al., 1987; Ball et al., 2002). Кроме того, песенная активность связана с размером вокальных ядер головного мозга птиц (Saldanha et al., 2000; Schlinger, Brenowitz, 2002).
Показано, что способность к пению и выучиванию песен, как и повышение активности пения для многих видов, положительно коррелирует с повышением уровня тестостерона в крови (Nottebohm et al., 1987; Canady et al.; 1988, Beletsky et al., 1990; DeVoogd, 1991; Cynx, Nottebohm, 1992; Wingfield, 1994; Wingfield, Hahn, 1994). У мухоловки-пеструшки (Ficedula hypoleuca) с уровнем тестостерона в крови связан также размер репертуара (Lampe, Espmark, 1994). Имплантация тестостерона или эстрадиола может стимулировать самок отвечать более активно на песни самцов и даже петь (Simpson, Vicario, 1991; Vallet et al., 1996). Повышение уровня тестостерона у канарейки и скворца (Sturnus vulgaris) коррелирует с увеличением активности пения и размером вокальных ядер в мозге птиц (Sartor, Ball, 2005; Sartor et al., 2005).
Несмотря на интерес многих исследователей к действию тестостерона на пение птиц, работ, в которых показано его влияние на характеристики песни, гораздо меньше. С уровнем тестостерона в крови у деревенской ласточки (Hirundo rustica) коррелируют длительность и скорость исполнения трещащих элементов песни (Galeotti et al., 1997), а у зебровой амадины – частотно-временные характеристики песни (Cynx et al., 2005). Недавно показано, что введение ингибиторов тестостерона приводит к изменению диапазона частот и максимальной частоты отдельных фраз песни у горихвостки-чернушки (Apfelbeck et al., 2012). Нами на пеночке-трещотке (Phylloscopus sibilatrix) показано увеличение длины песни и первой фразы песни, а также изменение диапазона частот в связи с повышением уровня тестостерона в крови (Горецкая, Москаленко, 2015).
Тем не менее, у некоторых видов (например, деревенской ласточки) индивидуальные вариации активности пения не связаны с уровнем тестостерона в крови (Saino, Moller, 1995). Самцы подвида певчей овсянки Melospisza melodia morphna круглый год остаются акустически активными, тогда как уровень тестостерона осенью и зимой гораздо ниже, чем в весенне-летний период (Wingfield, 1994). Сходная ситуация наблюдается и у синиц: активное пение при высоком уровне тестостерона в крови в весенне-летний период и отсутствие достоверной связи между уровнем тестостерона и активностью пения в осенний период (Rost, 1992). Изучение птиц в природе показало, что базальный уровень тестостерона не всегда связан с уровнем агрессии. У видов, продолжающих петь и охранять территории зимой, песенное поведение может осуществляться на фоне низкого уровня тестостерона (табл. 1) (Ball et al., 2002; Wingfield, Silverin, 2002; Hau, Goymann, 2015).
Последние исследования частично проливают свет на неоднозначность влияния тестостерона на пение, размер вокальных ядер и на процесс обучения пению. Как и в случае с регуляцией агрессивного поведения, на пение, вероятно, влияет эстрадиол, синтезируемый в головном мозге птиц (Soma, 2006). Он превращается в эстроген, как у самцов, так и у самок воробьинообразных птиц и действует на вокальный центр (HVC). Вне сезона размножения положительная связь между уровнем андрогенов и пением может быть прервана за счет понижения чувствительности рецепторов в головном мозге птиц (Ball et al., 2002).
Изменение уровня тестостерона в крови на разных стадиях жизненного цикла довольно хорошо изучено у взрослых особей многих видов воробьинообразных птиц (Wingfield et al., 1990, 2000), в то время как динамика его изменений у молодых птиц практически не исследована. Повышение уровня тестостерона было отмечено у молодых канареек: на 55–75-й день жизни у самцов и на 30–55-й день у самок, это время совпадает с началом исполнения подпесни (Weichel et al., 1986; Weichel et al., 1989). Повышение уровня тестостерона у молодых больших синиц (Parus major) также, предположительно, совпадало с началом песенной продукции (Rost, 1990). Недавнее исследование показало, что введение имплантов тестостерона вызывает раннее формирование песни у зебровой амадины (Leitao, 2013), а также способствует завершению формирования песни у молодых особей певчей овсянки (Templeton et al., 2012). Наши не опубликованные данные также показывают, что введение имплантов тестостерона трехмесячным самцам канареек стимулирует их к исполнению взрослой песни уже на 10-й день эксперимента. Повышение уровня тестостерона у птиц в период формирования пластической песни способствует более точному воспроизведению элементов, характерных для взрослых особей (DeVoogd, Szekely, 1998).
Хотя изучению влияния тестостерона на поведение и физиологию птиц посвящено большое количество работ, до сих пор не сформировалась единая гипотеза о роли этого гормона. Тестостерон, будучи половым гормоном, отвечает не только за размножение, но и за формирование агрессивного поведения, поведения ухаживания и за развитие вторичных половых признаков. Поэтому наибольшей популярностью сейчас пользуются две гипотезы. Согласно одной из них, тестостерон рассматривается как гормон, связанный с качеством песни, окраской, социальным статусом самца и степенью его иммунокомпетентности – признаками, по которым самки при выборе полового партнера могут оценить степень приспособленности последнего. А другая гипотеза Вызова (провокации) говорит о том, что концентрация тестостерона в крови увеличивается в ходе агонистических взаимодействий между самцами. До сих пор не ясно, как именно связаны половые гормоны с иммунокомпетентностью у птиц. Есть сведения, что тестостерон обладает подавляющим иммунитет эффектом, однако другие исследования этого не подтверждают.
Несмотря на интерес многих исследователей к действию тестостерона на пение птиц, до сих пор практически нет работ, в которых показано его влияние на характеристики песни. Практически не изучено изменение уровня тестостерона в крови у молодых птиц. Как воздействует тестостерон на способность формировать видовую песню у птенцов? Получены лишь первые ответы на эти вопросы.
Таким образом, несмотря на почти столетнюю историю изучения тестостерона, до сих пор остаются много загадок и большой простор для дальнейших исследований.
Список литературы
Горецкая М.Я., Москаленко В.Н., 2015. Гормональная регуляция вокального поведения птиц // Энергетика и годовые циклы птиц (памяти В.Р. Дольника). Материалы междунар. конф. Москва: Товарищество научных изданий КМК. С. 113–116.
Горецкая М.Я., Царелунга А.А., Сильверстов Н.А., Бёме И.Р., 2018. Влияние тестостерона и кортикостерона на выраженность иммунного ответа на эритроциты барана и активность пения у русской канарейки (Serinus canaria var. domestica) // Орнитология: история, традиции, проблемы и перспективы. Материалы Всероссийской конференции с международным участием, посвященной 120-летию профессора Г.П. Дементьева. Москва: Товарищество научных изданий КМК. C. 129–131.
Дьяченко В.П., Дольник В.Р., 1984. Роль пролактина в регуляции сезонных явлений у птиц // Успехи современной биологии. Т. 97. С. 295–308.
Apfelbeck B., Kiefer S., Mortega K.G., Goymann W., Kipper S., 2012. Testosterone affects song modulation during simulated territorial intrusions in male Black Redstarts (Phoenicurus ochruros) // PLoS ONE. V. 7. P. 1–12.
Ball G.F., Riters L.V., Balthazart J., 2002. Neuroendocrinology of song behavior and avian brain plasticity: Multiple sites of action of sex steroid hormones // Frontiers in Neuroendocrinology. V. 23. P. 137–178.
Balthazart J., Adkins-Regan E., 2002. Sexual differentiation of brain and behavior in birds // Pfaff D.W., Arnold A.P., Etgen A.M., Fahrbach S.E., Rubin R.T. (Eds). Hormones, brain and behavior. Amsterdam: Academic Elsevier. V. 4. P. 223–301.
Beletsky L.D., Orians G.H., Wingfield J.C., 1990. Steroid hormones in relation to territoriality, breeding density, and parental behaviour in male yellow-headed alackbirds // The Auk. V. 107. P. 60–68.
Bennett P.M., Owens I.P., 2002. Evolutionary ecology of birds. Life histories, mating systems and extinction // Oxford Series in Ecology and Evolution. Oxford: Oxford Univ. Press. 278 p.
Bókony V., Garamszegi L., Hirschenhauser K., Liker A., 2008. Testosterone and melanin-based black plumage coloration: a comparative study // Behavioral Ecology and Sociobiology. V. 62. P. 1229–1238.
Buchanan K.L., 2000. Stress and the evolution of condition-dependant signals // Trends in Ecology & Evolution. V. 15. P. 157–160.
Buchanan K.L., Evans M.R., Goldsmith A.R., Bryant D.M., Rowe L.V., 2001. Testosterone influences basal metabolic rate in male house sparrows: a new cost of dominance signalling? // Proceedings of the Royal Society of London. Series B. V. 268. P. 1337–1344.
Buchanan K.L., Evans M.R., Goldsmith A.R., 2003. Testosterone, dominance signaling and immunosuppression in the house sparrow, Passer domesticus // Behavioral Ecology and Sociobiology. V. 55. P. 50–59.
Canady R.A., Burd G.D., DeVoogd T.J., Nottebohm F., 1988. Effect of testosterone on input received by an identified neuron type of the canary song system: a golgielectron microscopy/degeneration study // Journal of Neuroscience. V. 8. № 10. P. 3770–3784.
Casto J.M., Van Nolan Jr., Ketterson E.D., 2001. Steroid hormones and immune function: experimental studies in wild and captive dark-eyed juncos (Junco hyemalis) // The American Naturalist. V. 157. P. 408–420.
Chastel O., Barbraud C., Weimerskirch H., Lormee H., Lacroix A., Tostain O., 2005. High levels of LH and testosterone in a tropical seabird with an elaborate courtship display // General and Comparative Endocrinology. V. 140. P. 33–40.
Collis K., Borgia G., 1992. Age-related effects of testosterone, plumage, and experience on aggression and social dominance in juvenile male satin bowerbirds // Auk. V. 109. P. 442–434.
Cynx J., Bean N.J., Rossman I., 2005. Testosterone implants alter the frequency range of zebra finch songs // Hormones and Behavior. V. 47. P. 446–451.
Cynx J., Nottebohm F., 1992. Testosterone facilitates some conspecific song discriminations in cactrated zebra finches (Taeniopygia guttata) // Neurobiology. V. 89. P. 6232–6234.
De Ridder E., Pinxten R., Eens M., 2000. Experimental evidence of a testosterone-induced shift from paternal to mating behaviour in a facultatively polygynous songbird // Behavioral Ecology and Sociobiology. V. 49. P. 24–30.
Deviche P., 1995. Androgen regulation of avian premigratory hyperphagia and fattening: from eco-physiology to neuroendocrinology // American Zoologist. V. 35. P. 234–245.
DeVoogd T.J., 1991. Endocrine modulation of the development and adult function of the avian song system // Psychoneuroendocrinology. V. 16. P. 41–66.
DeVoogd T.J., Szekely T., 1998. Causes of avian song: using neurobiology to integrate proximate and ultimate level of analysis in animal cognition in Nature // Balda P.P., Pepperberg I.M., Kamil A.C. (Eds). San Diego, London: Academic Press. P. 337–380.
Duckworth R.A., Mendonca M.T., Hill G.E., 2004. Condition dependent sexual traits and social dominance in the house finch // Behavioral Ecology. V. 15. P. 779–784.
Duffy D.L., Bentley G.E., Drazen D.L., Ball G.F., 2000. Effects of testosterone on cell-mediated and humoral immunity in non-breeding adult European starlings // Behavioral Ecology. V. 11. P. 654–662.
Elekonich M.M, Wingfield J.C., 2000. Seasonality and hormonal control of territorial aggression in female song sparrows (Passeriformes: Emberizidae: Melospiza melodia) // Ethology. V. 106. P. 493–510.
Enbody E.D., Boersma J., Schwabl H., Karubian J., 2018. Female ornamentation is associated with elevated aggression and testosterone in a tropical songbird // Behavioral Ecology, International Society for Behavioral Ecology. V. 29(5). P. 1056–1066.
Evans M.R., Goldsmith A.R., Norris S.R.A., 2000. The effects of testosterone on antibody production and plumage colouration in male house sparrows (Passer domesticus) // Behavioral Ecology. V. 47. P. 156–163.
Folstad I., Karter J., 1992. Parasites, bright males, and the immunocompetence handicap // American Naturalist. V. 139. P. 603–622.
Fusani S., 2008. Testosterone control of male courtship in birds // Hormones and Behavior. V. 54. P. 227–233.
Galeotti P., Saino N., Sacchi R., Møller A.P., 1997. Song correlates with social context, testosterone and body condition in male Barn Swallows // Animal Behaviour. V. 53 P. 687–700.
Garamszegi L.Z., Eens M., Hurtrez-Bousses S., Møller A.P., 2005. Testosterone, testes size, and mating success in birds: a comparative study // Hormones and Behavior. V. 47. P. 389–409.
Garamszeg L.Z., Mølle, A.P., Torok J., Michl G., Peczely P., Richard M., 2004. Immune challenge mediates vocal communication in a passerine bird: an experiment // Behavioral Ecology. V. 15. P. 148–157.
Geslin T., Chastel O., Eybert M.C., 2004. Sex-specific patterns in body condition and testosterone level changes in a territorial migratory bird: the Bluethroat Luscinia svecica // Ibis. V. 146. P. 632–641.
Gonzalez G., Sorci G., Smith L.C., de Lope F., 2001. Testosterone and sexual signalling in male house sparrows (Passer domesticus) // Behavioral Ecology and Sociobiology. V. 50. P. 557–562.
Goymann W., 2009. Social modulation of androgens in male birds// General and Comparative Endocrinology. V. 163. P. 149–157.
Goymann W., Landys M.M., Wingfield J.C., 2007. Distinguishing seasonal androgen responses from male–male androgen responsiveness - Revisiting the Challenge Hypothesis // Hormones and Behavior. V. 51. P. 463–476.
Goymann W., Wingfield J.C., 2014. Male-to-female testosterone ratios, dimorphism, and life history what does it really tell us? // Behavioral Ecology. V. 25. P. 685–689.
Grafen A., 1990. Biological signals as handicaps // Journal of Theoretical Biology. V. 102. P. 549–567.
Harding C.F., Follet B.K., 1979. Hormone changes triggered by aggression in a natural population of blackbirds // Science. V. 203. P. 918–920.
Hasselquist D., Marsh J.A., Sherman P.W., Wingfield J.C., 1999. Is avian humoral immunocompetence suppressed by testosterone? // Behavioral Ecology and Sociobiology. V. 45. P. 167–175.
Hau M., 2007. Regulation of male traits by testosterone: implications for the evolution of vertebrate life histories // BioEssays. V. 29(2). P. 133–144.
Hau M., Goymann W., 2015. Endocrine mechanisms, behavioral phenotypes and plasticity: known relationships and open questions // Frontiers in Zoology. V. 12 (Suppl 1). S 7.
Hau M., Stoddard S.T., Soma K.K., 2004. Territorial aggression and hormones during the non-breeding season in a tropical bird // Hormones and Behavior. V. 45. P. 40–49.
Hegner R.E., Wingfield J.C., 1987. Effects of experimental manipulation of testosterone levels on parental investment and breeding success in male house sparrows // Auk. V. 104. P. 462–469.
Hill G.E., 1994. Testis mass and subadult plumage in black- headed grosbeaks // Condor. V. 96. P. 626–630.
Hill G.E., Brawner W.R., 1998. Melanin-based plumage coloration in the house finch is unaffected by coccidial infection // Proceedings of the Royal Society. Ser. B. V. 265. P. 1105–1109.
Hirschenhauser K., Winkler H., Olivera R. F., 2003. Comparative analysis of male androgen re- sponsiveness to social environment in birds: the effects of mating system and paternal incubation // Hormones and Behavior. V. 43. P. 508–519.
Jawor J.M., Breitwisch R., 2003. Melanin ornaments, honesty, and sexual selection // Auk. V. 120. P. 249–256.
Ketterson E.D., Nolan V.Jr., Wolf L., Ziegenfus C., 1992. Testosterone and avian life histories: Effects of experimen- tally elevated testosterone on behavior and correlates of fit- ness in the dark-eyed junco (Junco hyemalis) // The American Naturalist. V. 140. P. 980–999.
Ketterson E.D., Nolan V.Jr., Sandell M., 2005. Testosterone in females: mediator of adaptive traits, constraint on sexual dimorphism, or both? // The American Naturalist. V. 166. P. S85–S98.
Lampe H.M., Espmark Y.O., 1994. Song structure reflects male quality in Pied Flycatchers, Ficedula hypoleuca // Animal Behaviour. V. 47. P. 869–876.
Leitao A., 2013. Testosterone influences the song onset in male Zebra Finch // International Bioacoustics Congress 8-18 of August. Brazil. P. 4.
Lindsay W.R., Webster M.S., Varian C.W., Schwabl H., 2009. Plumage colour acquisition and behaviour are associated with androgens in a phenotypically plastic tropical bird // Animal Behaviour. V. 77. P. 1525–1532.
Logan C.A., Wingfield J.C., 1995. Hormonal correlates of breeding status, nest construction, and parental care in multiple-brooded northern mockingbirds, Mimus polyglottos // Hormones and Behavior. V. 29. P. 12–30.
McGlothlin J.W., Jawor J.M., Greives T.J., Casto J.M., Phillips J.L., Ketterson E. D., 2008. Hormones and honest signals: males with larger ornaments elevate testosterone more when challenged // Journal of Evolutionary Biology. V. 21. P. 39–48.
Merila J., Sheldon B.C., 1999. Testis size variation in the green- finch Carduelis chloris: relevance for some recent models of sexual selection // Behavioral Ecology and Sociobiology. V. 45. P. 115–123.
Moller A.P., Eritzoe J., 1988. Badge, body and testes size in House Sparrows Passer domesticus // Ornis Scandinavica. V. 19. P. 72–73.
Moore I.T., Wada H., Perfito N., Buch D.S., Hahn T.P., Wingfield J.C., 2004. Territoriality and testosterone in an equatorial population of Rufous-collared Sparrows, Zonotrichia capensis // Animal Behaviour. V. 67. P. 411–420.
Nottebohm F., Nottebohm M.E., Crane L.A., Wingfield J.C., 1987. Seasonal changes in gonadal hormone levels of adult male Canaries and their relation to song // Behavioral and neural biology. V. 47. P. 197–211.
Olson V.A., Owens I.P.F., 2005. Interspecific variation in the use of carotenoid-based coloration in birds: diet, life history and phylogeny // Journal of Evolutionary Biology. V. 18. P. 1534–1546.
Owen-Ashley N.T., Hasselquist D., Wingfield J.C., 2004. Androgens and the Immunocompetence Handicap Hypothesis: unravelling direct and indirect pathways of immunosuppression in song sparrows // The American Naturalist. V. 164. P. 490–505.
Perez-Rodriguez L., Blas J., Vinuela J., Marchan, T.A., Bortolotti, G.R., 2006. Condition and androgen levels: are condition-dependent and testosterone- mediated traits two sides of the same coin? // Animal Behaviour. V. 72. P. 97–103.
Peters A., 2000. Testosterone treatment is immunosuppressive in superb fairy- wrens, yet free-living males with high testosterone are more immunocompetent // Proceedings of the Royal Society of London. V. 267. P. 883–889.
Peters A., Astheimer L.B., Boland C.R.J., Cockburn A., 2000. Testosterone is involved in acquisition and maintenance of sexually selected male plumage in superb Fairy-Wrens, Malurus cyaneus // Behavioral Ecology and Sociobiology. V. 47. P. 438–445.
Peters A., Cockburn A., Cunningham R., 2002. Testosterone treatment suppresses paternal care in superb fairy-wrens, Malurus cyaneus, despite their concurrent investment in courtship // Behavioral Ecology and Sociobiology. V. 51. P. 538–547.
Peters A., Delhey K., Goymann W., Kempenaers B., 2006. Age-dependent association between testosterone and crown UV coloration in male blue tits (Parus caeruleus) // Behavioral Ecology and Sociobiology. V. 59. P. 666–673.
Poiani A., Fletcher T., 1994. Plasma levels of androgens and gonadal development of breeders and helpers in the bell miner (Manorina melanophrys) // Behavioral Ecology and Sociobiology. V. 34. P. 31–41.
Råberg L., Grahn M., Hasselquist D., Svensson E., 1998. On the adaptive significance of stress-induced immunosuppression // Proceedings of the Royal Society of London. V. 265. P. 1637–1641.
Raouf S.A., Parker P.G., Ketterson E.D., Nolan V.Jr., Ziegenfus C., 1997. Testosterone affects reproductive success by influencing extra-pair fertilizations in male dark-eyed juncos (Aves: Junco hyemalis) // Proceedings of the Royal Society of London. Series B. V. 264. P. 1599–1603.
Ramenofsky M., 1984. Endogenous plasma hormones and agonistic behavior in male Japanese quail, Coturnix coturnix // Animal Behaviour. V. 32. P. 698–708.
Roberts M.L., Buchanan K.L., Evans M.R., 2004. Testing the immunocompetence handicap hypothesis: a review of the evidence // Animal Behaviour. V. 68. P. 227–239.
Roberts M.L., Buchanan K.L., Hasselquist D., Evans M.R., 2007. Effects of testosterone and corticosterone on immunocompetence in the zebra finch // Hormones and Behavior. V. 51. P. 126–134.
Roberts M. L., Ras E., Peters A., 2009. Testosterone increases UV reflectance of sexually selected crown plumage in male blue tits // Behavioral Ecology. V. 20. P. 535–541.
Romero L.M., Soma K.K., O´ Reilly K.M., Suydam R., Wingfield J.C., 1997. Territorial behaviour, hormonal changes, and body condition in an arctic-breeding song bird, the Redpoll (Carduelis flammea) // Behaviour. V. 134. P. 727–747.
Rost R., 1990. Hormones and behaviour: A joint examination of studies on seasonal variation in song production and plasma levels of testosterone in the Great Tit Parus major// Journal of Ornithology. V. 131. P. 403–411.
Rosvall K.A., 2013. Proximate perspectives on the evolution of female aggression: good for the gander, good for the goose? // Philosophical Transactions of the Royal Society Biological Sciences. V. 368 P. 2013–2083.
Saino N., Møller A.P., 1995. Testosterone correlates of mate guarding, singing and aggressive behaviour in male Barn Swallows, Hirundo rustica // Animal Behaviour. V. 49. № 2. P. 465–472.
Saldanha C.J., Tuerk M.J., Kim Y.H., Fernandes A.O., Arnold A.P., Schlinger B.A., 2000. Distribution and regulation of telencephalic aromatase expression in the Zebra Finch revealed with a specific antibody // Journal of Neuroendocrinology. V. 423. P. 619–630.
Sandell M.I., 2007. Exogenous testosterone increases female aggression in the European starling (Sturnus vulgaris) // Behavioral Ecology and Sociobiology. V. 62. P. 255–262.
Sapolsky R.M., Romero M., Munck A.U., 2000. How do glucocorticoids influence stress responses? Integrating permissive, suppressive, stimulatory, and preparative actions // Endocrine Reviews. V. 21. P. 55–89.
Sartor J.J., Ball G.F., 2005. Social suppression of song is associated with a reduction in volume of a song-control nucleus in European Starlings (Sturnus vulgaris) // Behavioral Neuroscience. V. 119. P. 233–244.
Sartor J.J., Diekamp B., Fortune E.S., Ball G.F., 2005. Neuroplasticity in the song nucleus HVC is modulated by testosterone and production of the bird’s own song in songbirds // Hormones and Behavior. V. 48. P. 126.
Schlinger B., Brenowitz E.A., 2002. Neural and Hormonal Control of Birdsong // Pfaff D.W. Hormones, Brain and Behavior. V. 2. Elsevier. P. 799–838.
Schoech S.J., Mumme R.L., Moore M.C., 1991. Reproductive endocrinology and mechanisms of breeding inhibition in cooperatively breeding Florida scrub jays (Aphelocoma c. coerulescens) // Condor. V. 93. P. 354–364.
Schwarzová L., Fuch, R., Frynta D., 2010. Delayed Plumage Maturation Correlates with Testosterone Levels in Black Redstart Phoenicurus ochruros Males // Acta Ornithologica. V. 45. P. 91–97.
Simpson H.B., Vicario D.S., 1991. Early estrogen treatment of female zebra finches masculinizes the brain pathway for learned vocalizations // Journal of Neurobiology. V. 22. P. 777–793.
Soma K.K., 2006. Testosterone and aggression: Berthold, birds and beyond // Journal of Neuroendocrinology. V. 8. P. 543–551.
Soma K.K., Wingfield J.C., 2001. Dehydroepiandrosterone in songbird plasma: Seasonal regulation and relationship to territorial aggression // General and Comparative Endocrinology. V. 123. P. 144–155.
Soma K.K., Wissman A.M., Brenowitz E.A., Wingfield J.C., 2002. Dehydroepiandrosterone (DHEA) increases territorial song and the size of an associated brain region in a male songbird // Hormones and Behavior. V. 41. P. 203–212.
Staub N.L., de Beer M., 1997. The role of androgens in female vertebrates // General and Comparative Endocrinology. V. 108. P. 1–24.
Templeton C.N., Burt J.M., Campbell S.E., Lent K., Brenowitz E.A., Beecher M.D., 2012. Immediate and long-term effects of testosterone on song plasticity and learning in juvenile song sparrows // Behavioral Processes. V. 90. P. 254–260.
Tonra C.M., Marra P.P., Holberton R.L., 2011. Migration phenology and winter habitat quality are related to circulating androgen in a long-distance migratory bird // Journal of Avian Biology. V. 42. P. 397–404.
Tramontin A.D., Brenowitz E.A., 2000. Seasonal plasticity in the adult brain // Trends of Neuroscience. V. 23. P. 251–258.
Vallet E., Kreutzer M., Gahr M., 1996. Testosterone induces sexual release quality in the song of female canaries // Ethology. V. 102. P. 617–628.
Vleck C.M., Brown J.L., 1999. Testosterone and social and reproductive behaviour in Aphelocoma jays // Animal Behaviour. V. 58. P. 943–951.
Weichel K., Schwager G., Heid P., Güttinger H.R., 1989. 17 β-Estradiolbenzoate-dependent song induction in juvenile female canaries (Serinus canaria) // Long-term Measurements of Song Activity. V. 80. P. 55–70.
Weichel K., Schwager G., Heid P., Güttinger H.R., Pesch A., 1986. Sex differences in plasma steroid concentrations and singing behavior during ontogeny in Canaries (Serinus canaria) // Ethology. V. 73. P. 281–294.
Wingfield J.C., 1985. Short-term changes in plasma levels of hormones during establishment and defense of a breeding territory in male song sparrows, Melospiza melodia // Hormones and Behavior. V. 19. P. 174–187.
Wingfield J.C., 1994. Regulation of territorial behaviour in the Sedentary Song Sparrow, Melospiza melodia morphna // Hormones and Behavior. V. 28. P. 1–15.
Wingfield J.C., 2005. A continuing saga: The role of testosterone in aggression // Hormones and Behavior. V. 48. P. 253–255.
Wingfield J.C., Hahn T.P., 1994. Testosterone and territorial behaviour in sedentary and migratory sparrows // Animal Behaviour. V. 47. P. 77–89.
Wingfield J.C., Hegner R.E., Dufty Jr.A.M., Ball G.F., 1990. The “challenge hypothesis”: theoretical implications for patterns of testosterone secretion, mating systems, and breeding strategies // American Naturalist. V. 136. P. 829–846.
Wingfield J.C., Hegn R.E., Lewis D.M., 1991. Circulating levels of Luteinizing- hormone and steroid-hormones in relation to social-status in the cooperatively breeding white-browed sparrow weaver, Plocepasser mahali // Journal of Zoology. V. 225. P. 43–58.
Wingfield J.C., Jacobs J.D., Tramontin A.D., Perfito N., Meddle S., Maney D.L., Soma K., 2000. Toward an ecological basis of hormone–behavior interactions in reproduction of birds // Reproduction in Context: Social and Environmental Influences on Reproduction. Wallen K., Schneider J.E. (Eds) Cambridge, MA: MIT Press. P. 85–128.
Wingfield J.C., Silverin B., 2002. Ecophysiological studies of hormone–behavior relations in birds // Pfaff D.W. (Ed.) Hormones, Brain and Behavior. Amsterdam: Academic Press. P. 587–647.
Wikelski M., Lynn S., Breuner C., Wingfield J.C., Kenagy G.J., 1999. Energy metabolism, testosterone and corticosterone in white-crowned sparrows // Journal of Comparative Physiology. V. 185. P. 463–470.
Zahavi A., 1975. Mate selection – a selection for a handicap // The Journal of Theoretical Biology. V. 67. P. 603–605.
Дополнительные материалы отсутствуют.
Инструменты
Зоологический журнал