

Зоологический журнал, 2020, T. 99, № 2, стр. 138-145
Ilyocryptus spinifer Herrick 1882 (Crustacea, Branchiopoda, Cladocera): первая находка вида для фауны Европы
В. С. Жихарев a, *, А. Н. Неретина b, **, Т. В. Золотарева a, Д. Е. Гаврилко a, Г. В. Шурганова a
a Национальный исследовательский Нижегородский государственный университет
им. Н.И. Лобачевского
603950 Нижний Новгород, Россия
b Институт проблем экологии и эволюции им. А.Н. Северцова РАН
119071 Москва, Россия
* E-mail: slava.zhiharev@ro.ru
** E-mail: neretina-anna@yandex.ru
Поступила в редакцию 27.01.2019
После доработки 28.03.2019
Принята к публикации 26.05.2019
Аннотация
В 2018 г. в р. Ветлуга (Нижегородская обл., европейская часть России) нами впервые обнаружен ветвистоусый рачок Ilyocryptus spinifer Herrick 1882 (Cladocera, Ilyocryptidae). Этот вид ведет бентосно-фитофильный образ жизни, относится к южному теплолюбивому комплексу кладоцер и имеет циркумтропический ареал. I. spinifer никогда прежде не был отмечен в Европе, единичные находки этого вида известны из немногих регионов в азиатской части России. Проникновение I. spinifer в более высокие широты может быть связано с расширением его естественного ареала в связи с глобальным потеплением климата или переносом с мигрирующими птицами и балластными водами. Эта находка указывает на недостаточную изученность бентических и литоральных ветвистоусых ракообразных рек европейской части России. Дальнейший мониторинг обнаруженных популяций очень важен для контроля состояния водных экосистем России и других стран.
Фауна пресноводных микроракообразных в умеренных регионах Европы инвентаризована лучше, чем в других регионах Земного шара (Błędzki, Rybak, 2016). Наиболее хорошо изучены ветвистоусые ракообразные (Crustacea, Branchiopoda, Cladocera) данного региона (Котов, 2013; Błędzki, Rybak, 2016). История исследований ветвистоусых ракообразных в водоемах Европы насчитывает несколько столетий (Forró et al., 2008; Котов, 2013; Błędzki, Rybak, 2016). За это время накоплен большой массив данных по морфологии, систематике и экологии этих представителей зоопланктона. Сегодня ветвистоусые ракообразные по праву считаются одной из важнейших групп беспозвоночных животных, задействованных в экологическом мониторинге и исследованиях по оценке состояния водных ресурсов (Определитель …, 2010; Błędzki, Rybak, 2016). Однако далеко не все экологические группы ветвистоусых ракообразных учитываются специалистами-гидробиологами.
Как правило, исследователи отбирают пробы только из зоны открытой воды крупных озер, водохранилищ и рек и имеют дело в основном с планктонными видами кладоцер (Korovchinsky, 1997; Котов, 2013; Błędzki, Rybak, 2016). Видовое богатство в таких пробах относительно невелико. В то же время, ветвистоусые ракообразные прибрежных зарослей макрофитов и ила далеко не всегда учитываются в экологических исследованиях, а ведь значительное число видов кладоцер ведет именно бентосно-фитофильный образ жизни (Смирнов, 1975; Котов, 2006, 2013). Такая неравномерность распределения усилий исследователей не позволяет реконструировать целостную картину процессов, происходящих в водных экосистемах. Неудивительно, что среди кладоцер, обитающих в литорали и бентали, скрывается много неучтенных видов (Korovchinsky, 1996). Особенно часто при отборе и обработке речных проб специалисты-гидробиологи просматривают представителей семейства Ilyocryptidae, типичных обитателей зарослей и придонного ила, в том числе, и дна рек (Смирнов, 1976; Kotov, Štifter, 2006; Котов, 2013).
Данное сообщение посвящено описанию популяции Ilyocryptus spinifer Herrick 1882 (Cladocera, Ilyocryptidae), обнаруженной нами в р. Ветлуга (Нижегородская обл., европейская часть России).
Пробы зоопланктона были отобраны на пяти станциях (табл. 1), заложенных на р. Ветлуга в районе природного парка “Воскресенское Поветлужье” в июле 2018 г. Материал отбирали при помощи планктонной сети Джеди (нейлоновое сито с ячеей 70 мкм) и на месте фиксировали 4%-ным раствором формалина.
Таблица 1.
Описание станций отбора проб на р. Ветлуга и количество обнаруженных экземпляров Ilyocryptus spinifer Herrick 1882 в июле 2018 г.
| № станции | Географические координаты | n, экз. | Т, °С | рН | Глубина, м | Прозрачность, м |
|---|---|---|---|---|---|---|
| 1 | 56.856974° с.ш., 45.437230° в.д. | – | 21.3 | 9.1 | 1.4 | 0.8 |
| 2 | 56.856010° с.ш., 45.437251° в.д. | – | 21.7 | 9.1 | 1.2 | 1.0 |
| 3 | 56.851307° с.ш., 45.437916° в.д. | – | 22.7 | 8.9 | 1.0 | 0.8 |
| 4 | 56.839760° с.ш., 45.440663° в.д. | 2 | 23.8 | 9.4 | 1.2 | 0.7 |
| 5 | 56.824045° с.ш., 45.447229° в.д. | 4 | 23.0 | 9.1 | 1.2 | 0.7 |
В лаборатории пробы зоопланктона разбирали под бинокулярным микроскопом Carl Zeiss Stemi 2000C (Carl Zeiss Microscopy GmbH, Германия) при малом увеличении. Детальную микроскопическую обработку проводили на микроскопе Meiji Techno MT4200L (Meiji Techno, Япония). Определение особей I. spinifer в пробах из р. Ветлуга проведено согласно общепринятым определителям и специальным статьям (Kotov, Dumont, 2000; Kotov, Williams, 2000; Kotov, Štifter, 2006). Рисунки особей I. spinifer выполнены при помощи рисовального аппарата, установленного на микроскоп Olympus BX41 (Olympus Corporation, Япония). При описании морфологии использована терминология, принятая в отечественных руководствах (Определитель …, 2010; Котов, 2013).
Отряд Anomopoda Sars 1865
Ilyocryptus spinifer Herrick 1882 (рис. 1–3)
Рис. 1.
Ilyocryptus spinifer Herrick 1882, партеногенетическая самка из нижнего течения р. Ветлуга: a – общий вид, b – голова, c – створка, d–g – фрагменты створки при большем увеличении. Масштаб 0.1 мм.

Рис. 2.
Ilyocryptus spinifer Herrick 1882, партеногенетическая самка из нижнего течения р. Ветлуга: a–b – постабдомен, общий вид; c – постабдоминальный коготок; d – антенна I; e–f – фрагменты антенны I при большем увеличении; g – антенна II; h–i – латеральные плавательные щетинки эндоподита антенны II. Масштаб 0.1 мм.
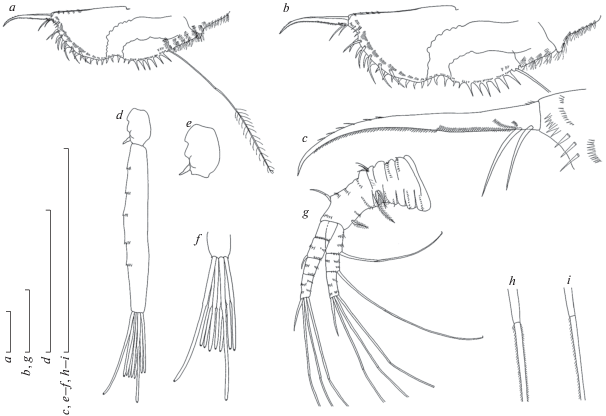
Рис. 3.
Ilyocryptus spinifer Herrick 1882, партеногенетическая самка из нижнего течения р. Ветлуга: a – торакопод I, b – торакопод II, c – торакопод III, d – торакопод IV, e – гнатобаза торакопода IV, f – торакопод V, g – торакопод VI. Масштаб 0.1 мм.

Материал. Шесть партеногенетических самок, территория природного парка “Воскресенское Поветлужье” в нижнем течении р. Ветлуга (табл. 1), литоральная зона реки на песчаном грунте. Найдены на двух из пяти обследованных станций.
Описание. В латеральной проекции тело высокое, широкоовальное (рис. 1a). Максимальная высота тела в задней его половине. Выемка между головой и оставшейся частью тела слабо выражена, спинной край широко-выгнутый (рис. 1a). Задне-спинной угол относительно хорошо выражен, задне-брюшная часть створки широко-округлая (рис. 1a, 1c). Латеральные отростки и волоски на створках отсутствуют. Линька неполная, головной щит и створки многослойные (рис. 1a, 1c).
Голова маленькая, в латеральной проекции треугольная (рис. 1a–1b). Спинной край головы сильно выгнутый, брюшной край практически прямой с небольшой выемкой перед основаниями антеннул. Сложный глаз и глазок расположены у брюшного края головы, глаз значительно крупнее глазка (рис. 1a–1b).
Лабрум вытянутый, с небольшим оттянутым выростом в передней части (рис. 1b).
Створки очень большие, округлые (рис. 1c). Передний, брюшной, и задний края формируют широко-выпуклый свободный край створки (рис. 1a, 1c). На створках хорошо различима гексагональная ретикуляция (рис. 1a, 1c). Вдоль свободного края створки проходит ряд оперенных щетинок (рис. 1a, 1c). Краевые щетинки передней (рис. 1g) и брюшной частей створки оперены длинными сетулами. Краевые щетинки задней и задне-брюшной части створки также оперены длинными сетулами (рис. 1d), но их основания снабжены небольшими одиночными шипами (рис. 1d–1f).
Торакс относительно длинный для Anomopoda, несет шесть пар торакальных конечностей (рис. 1a). Абдомен относительно короткий, значительно короче постабдомена (рис. 1a).
Постабдомен большой, постепенно сужающийся дистально (рис. 2a–2b). Максимальная высота постабдомена в проксимальной части. Брюшной край постабдомена практически прямой, покрыт несколькими рядами сетул. Спинной край постабдомена широко-выпуклый, с небольшим углублением в районе анального отверстия (рис. 2a–2b). Длина постанального края почти в два раза больше длины преанального края (рис. 2b). Вдоль преанального края расположен ряд одиночных зубцов (рис. 2b). Этот ряд переходит в ряд сдвоенных зубцов на анальном и постанальном краях. Боковая поверхность постанальной части несет несколько длинных зубцов. Над ними в постанальной части располагается ряд пучков сетул, плавно переходящий на анальную часть (рис. 2a–2b). Вооружение боковой поверхности преанальной части представлено рядом мелких зубчиков. Группы из нескольких мелких зубчиков также различимы по бокам, у основания постабдоминальных щетинок и коготков (рис. 2b).
Постабдоминальные коготки длинные, постепенно сужающиеся дистально (рис. 2c). Сбоку спинной край каждого коготка покрыт тремя–четырьмя гребнями из тонких сетул. Брюшной край коготков несет несколько относительно длинных щетинок. При основании коготка расположено два длинных тонких базальных шипа (рис. 2c).
Постабдоминальные щетинки превышают по длине постабдомен (рис. 1a, 2a). Дистальный сегмент покрыт тонкими длинными волосками.
Антенна I палочковидная, длинная, состоит из двух сегментов (рис. 2d). Проксимальный сегмент короткий, с небольшим пальцевидным выростом и несколькими бугорками (рис. 2d–2e). Дистальный сегмент длинный, покрыт несколькими поперечными рядами зубчиков, на конце несет девять эстетасков (два длинных и семь коротких) (рис. 2d, 2f).
Антенна II массивная, относительно длинная для представителя рода (рис. 2g). Коксальный сегмент короткий и широкий, со складками, окаймленными мелкими зубчиками и волосками. Дистально на его боковой поверхности расположены две оперенные чувствительные щетинки (рис. 2g). Базальный сегмент длинный, покрыт поперечными рядами зубчиков, несет одну оперенную чувствительную щетинку на внутренней стороне и шип на внешней. Экзоподит четырехчлениковый, эндоподит трехчлениковый. Все сегменты покрыты поперечными рядами мелких зубчиков (рис. 2g). Антеннальная формула: плавательные щетинки 0-0-0-3/1-1-3, шипы 0-1-0-1/0-0-1. Шип апикального сегмента экзоподита чуть короче шипа эндоподита, но почти в два раза длиннее шипа, расположенного на втором сегменте экзоподита. Апикальные плавательные щетинки обеих ветвей очень длинные (рис. 2g). Латеральные плавательные щетинки эндоподита значительно короче апикальных, оперены короткими жесткими сетулами (рис. 2h–2i), их кончики заканчиваются небольшими, характерными крючочками (рис. 2g).
Торакальная конечность I пары с небольшим овальным эпиподитом (рис. 3a). Внешняя дистальная доля конечности покрыта длинными сетулами. Дистально от нее расположены длинная односторонне оперенная щетинка и очень короткая двустороннее оперенная щетинка (рис. 3a). Наружная часть внешней дистальной доли несет относительно длинную двустороннее оперенную щетинку. Два выталкивающих крюка расположены на передне-дистальной поверхности. На внутреннем крае конечности расположено пять двусторонне оперенных щетинок и две сенсиллы (рис. 3a).
Торакальная конечность II пары с небольшим овальным эпиподитом (рис. 3b). Дистальная доля с двумя длинными двусторонне оперенными щетинками и небольшим бугорком (рис. 3b). Вдоль внутреннего края торакопода расположено четыре щетинки и две сенсиллы. Внутренняя часть торакопода представлена овальной опушенной лопастью. При ее основании расположена небольшая сенсилла. Длинная щетинка, оперенная относительно короткими жесткими сетулами, расположена при основании гнатобазы (рис. 3b). Дистальное вооружение гнатобазы представлено тремя сближенными щетинками и толстой сенсиллой с оттянутым концом. Фильтрационная пластинка гнатобазы несет восемь щетинок (рис. 3b).
Торакальная конечность III пары с небольшим овальным эпиподитом (рис. 3c). Экзоподит с пятью терминальными и тремя латеральными щетинками. Дистальный эндит внутренней доли с двумя короткими и тремя длинными, двустороннее оперенными щетинками (рис. 3c). Базальный эндит с четырьмя длинными щетинками, небольшим бугорком и искривленной короткой щетинкой. Гнатобаза состоит из четырех элементов, одна из щетинок гнатобазы очень длинная. Фильтрационная пластинка гнатобазы несет восемь щетинок (рис. 3c).
Торакальная конечность IV пары с вытянутым опушенным преэпиподитом и овальным эпиподитом (рис. 4d ). Экзоподит округлый, несет восемь щетинок. Внутренняя дистальная доля торакопода представлена плоской лопастью, несущей пять щетинок. Около внутреннего края расположен ряд из четырех щетинок (рис. 4d ). Дистальное вооружение гнатобазы представлено четырьмя элементами: очень толстой щетинкой, дистальная часть которой покрыта зубчиками, и тремя тонкими щетинками (рис. 4e ). Фильтрационная пластинка гнатобазы несет восемь щетинок (рис. 4d ).
Торакальная конечность V пары с вытянутым опушенным преэпиподитом и овальным эпиподитом (рис. 4f ). Крупный овальный экзоподит несет семь щетинок. Внутренняя часть торакопода формирует крупную лопасть, ее край покрыт тонкими длинными сетулами. Задняя сторона ноги несет две крупные щетинки. Дистальное вооружение гнатобазы представлено четырьмя элементами: двумя щетинками и двумя мелкими шипами. Фильтрационная пластинка гнатобазы несет четыре щетинки (рис. 4f ).
Торакальная конечность VI пары представлена треугольной лопастью, по краю которой расположено шесть пучков длинных жестких сетул (рис. 4g ).
Размеры: длина тела до 0.75 мм, высота тела до 0.56 мм.
По морфологическим признакам все изученные нами особи (рис. 1–3) идентичны I. spinifer из типового местообитания в Северной Америке (Kotov, Williams, 2000; Kotov, Štifter, 2006). I. spinifer был описан в 1882 г. из Миннесоты (северная часть США) (Herrick, 1882a,b). Для данной территории характерен влажный умеренный континентальный климат со значительными сезонными перепадами температуры. Однако большинство последующих находок I. spinifer были приурочены к тропическим регионам как Восточного, так и Западного полушария (Kotov, Williams, 2000; Kotov, Dumont, 2000; Kotov, Štifter, 2006). На основе этого I. spinifer был отнесен к южному теплолюбивому комплексу кладоцер с циркумтропическими ареалами (Котов, 2016). Тем не менее, и в Восточном полушарии известны единичные находки популяций I. spinifer в регионах с влажным умеренным (Kotov, Dumont, 2000; Tanaka 2001; Kotov, Štifter 2006; Котов и др., 2011; Kotov et al., 2012) или даже континентальным умеренным (Котов, 2016) климатом. Наиболее северными точками обитания этого вида в азиатской части России считаются северные притоки р. Амур (Котов и др., 2011) и верховья р. Обь (Котов, 2016). Находка популяции I. spinifer в р. Ветлуга является первой находкой этого вида не только для европейской части России, но и для Европы в целом (Определитель …, 2010). Он не отмечен даже в самых южных европейских странах: Испании (Alonso, 1996), Италии (Margaritora, 1985), Израиле (Bromley, 1993). Интересно, что изученная нами популяция обнаружена практически на той же широте, что и отмеченные ранее популяции из р. Обь.
Поскольку I. spinifer очень широко распространен на нескольких континентах, то можно предположить, что проникновение в более высокие широты обусловлено значительной экологической пластичностью данного вида. При этом достаточно часто находки теплолюбивых организмов в более северных регионах связывают с расширением их естественных ареалов в связи с глобальным потеплением климата (Семенов, 2009; Перова и др., 2018). Однако такие заключения должны быть основаны на достоверных данных о том, что ранее вид отсутствовал на данной территории. Такой сценарий не исключен и для I. spinifer, но подтвердить это можно только филогеографическими методами.
Немногочисленность находок I. spinifer за пределами тропических и субтропических регионов косвенно подтверждает гипотезу о недавнем расширении его ареала на север. Стоит отметить, что в центральной части Европы были сделаны единичные находки популяций и других теплолюбивых тропических видов ветвистоусых ракообразных, например Disparalona hamata s. l. (Illyová, Hudec, 2004; Louette et al., 2007; Neretina et al., 2018). Популяции I. spinifer явно способны существовать в регионах с морозными зимами. В частности, данный вид широко распространен в Южной Корее, где формирует стабильные популяции в водоемах, замерзающих в зимнее время (Kotov et al., 2012).
С другой стороны, проникновение I. spinifer в более высокие широты может быть объяснено разовыми заносами этого вида на мигрирующих водоплавающих птицах или при участии человека, например с балластными водами судов. Действительно, перенос птицами рассматривается ныне как основной естественный способ распространения пресноводных кладоцер (Figuerola, Green, 2002; Reynolds et al., 2015). В последнее время зарегистрированы и случаи расширения ареалов кладоцер за счет деятельности человека (Hebert, Cristescu, 2002; Havel, Medley, 2006).
При этом остается открытым вопрос о том, насколько популяция из Нижегородской обл. стабильна во времени. Это может быть выяснено только в ходе дальнейших работ по мониторингу кладоцер р. Ветлуга. Именно долгосрочные исследования должны быть основой контроля состояния водных экосистем России и других стран, в частности, именно они позволят уверенно говорить о расширении ареала того или иного вида.
Список литературы
Котов А.А., 2006. Адаптации ветвистоусых ракообразных отряда Anomopoda (Cladocera) к бентосному образу жизни // Зоологический журнал. Т. 85. № 9. С. 1043–1059.
Котов А.А., 2013. Морфология и филогения Anomopoda (Crustacea: Cladocera). М.: Товарищество научных изданий КМК. 638 с.
Котов А.А., 2016. Фаунистические комплексы Cladocera (Crustacea, Branchiopoda) Восточной Сибири и Дальнего Востока России // Зоологический журнал. Т. 95. № 7. С. 748–768.
Котов А.А., Синёв А.Ю., Коровчинский Н.М., Смирнов Н.Н., Беккер Е.И., Шевелева Н.Г., 2011. Cladocera (Crustacea, Branchiopoda) бассейна реки Зеи (Амурская область, Российская Федерация). 1. Новые таксоны для фауны России // Зоологический журнал. Т. 90. № 2. С. 131–142.
Определитель зоопланктона и зообентоса пресных вод Европейской России, 2010. Зоопланктон. Т. 1. Под ред. Алексеев В.Р., Цалолихин С.Я. М.: Товарищество научных изданий КМК. 495 с.
Перова С.Н., Пряничникова Е.Г., Жгарева Н.Н., 2018. Появление и распределение вселенцев в макрозообентосе водохранилищ верхней Волги // Российский журнал биологических инвазий. № 4. С. 41–52.
Семенов Д.В., 2009. Красноухая черепаха, Trachemys scripta elegans, как инвазивная угроза (Reptilia; Testudines) // Российский журнал биологических инвазий. № 1. С. 36–44.
Смирнов Н.Н., 1975. Биология ветвистоусых ракообразных // “Зоология беспозвоночных”. 3. Итоги науки и техники. М.: ВИНИТИ АН СССР. 116 с.
Смирнов Н.Н., 1976. Macrothricidae и Moinidae фауны мира // Фауна СССР, новая серия. Ракообразные 1 (3). Л.: Наука. 237 с.
Alonso M., 1996. Crustacea, Branchiopoda. Fauna Iberica 7. Crustacea Branchiopoda. Madrid: Museo Nacional de Ciencias Naturales. Consejo Superior de Investigaciones Científicas. 486 p.
Błędzki L.A., Rybak J.I., 2016. Freshwater Crustacean Zooplankton of Europe. Cladocera & Copepoda (Calanoida, Cyclopoida). Key to species identification, with notes on ecology, distribution, methods and introduction to data analysis. Switzerland: Springer International Publishing. 918 p.
Bromley H.J., 1993. A checklist of the Cladocera of Israel and Eastern Sinai // Hydrobiologia. V. 257. P. 21–28.
Figuerola J., Green A.J., 2002. Dispersal of aquatic organisms by waterbirds: a review of past research and priorities for future studies // Freshwater biology. V. 47. № 3. P. 483–494.
Forró L., Korovchinsky N.M., Kotov A.A., Petrusek A., 2008. Global diversity of cladocerans (Cladocera; Crustacea) in freshwater // Hydrobiologia. V. 595. P. 177–184.
Havel J.E., Medley K.A., 2006. Biological invasions across spatial scales: Intercontinental, regional, and local dispersal of cladoceran zooplankton // Biological Invasions. V. 8. № 3. P. 459–473.
Hebert P.D.N., Cristescu M.E.A., 2002. Genetic perspectives on invasions: the case of the Cladocera // Canadian Journal of Fisheries and Aquatic Sciences. V. 59. № 7. P. 1229–1234.
Herrick C.L., 1882. Notes on some Minnesota Cladocera // The Geology of Central and Western Minnesota, 10th annual report. P. 235–252.
Herrick C.L., 1882a. Mud-inhabiting Crustacea // Bulletin of the scientific laboratories of Denison University. P. 37–42.
Illyová M., Hudec I., 2004. Disparalona hamata (Birge, 1879) (Crustacea, Anomopoda) – the second record from Europe // Biologia, Bratislava. V. 59. № 2. P. 287–288.
Korovchinsky N.M., 1996. How many species of Cladocera are there? // Hydrobiologia. V. 321. № 3. P. 191–204.
Korovchinsky N.M., 1997. On the history of studies on cladoceran taxonomy and morphology, with emphasis on early work and causes of insufficient knowledge of the diversity of the group // Hydrobiologia. V. 360. P. 1–11.
Kotov A.A., Dumont H.J., 2000. Analysis of the Ilyocryptus spinifer s. lat. species group (Anomopoda, Branchiopoda), with description of a new species // Hydrobiologia. V. 428. P. 85–113.
Kotov A.A., Jeong H., Lee W., 2012. Cladocera (Crustacea: Branchiopoda) of the south-east of the Korean Peninsula, with twenty new records for Korea // Zootaxa. V. 3368. P. 50–90.
Kotov A.A., Štifter P., 2006. Cladocera: family Ilyocryptidae (Branchiopoda: Cladocera: Anomopoda) // Guides to the identification of the microinvertebrates of the Continental Waters of the world. V. 22. Leiden: Kenobi Productions, Ghent & Backhuys Publishers. 172 p.
Kotov A.A., Williams J.L., 2000. Ilyocryptus spinifer Herrick 1882 (Anomopoda, Branchiopoda): a redescription based on North American material and designation of a neotype from Minnesota // Hydrobiologia. V. 428. P. 67–84.
Louette G., De Bie T., Vandekerkhove J., Declerck S., De Meester L., 2007. Analysis of the inland cladocerans of Flanders (Belgium)-Inferring changes over the past 70 years // Belgian Journal of Zoology. V. 137. № 1. P. 117–123.
Margaritora F.G., 1985. Cladocera. Fauna d’ltalia 23. Bologna: Edizioni Calderini. 399 p.
Neretina A.N., Garibian P.G., Sinev A.Y., Kotov A.A., 2018. Diversity of the subgenus Disparalona (Mixopleuroxus) Hudec, 2010 (Crustacea: Cladocera) in the New and Old World // Journal of Natural History. V. 52. № (3–4). P. 155–205.
Reynolds C., Miranda N.A., Cumming G.S., 2015. The role of waterbirds in the dispersal of aquatic alien and invasive species // Diversity and Distributions. V. 21. № 7. P. 744–754.
Tanaka S., 2001. Three species of the genus Ilyocryptus (Anomopoda, Branchiopoda) occurring in Japan // Limnology. № 2. P. 219–222.
Дополнительные материалы отсутствуют.
Инструменты
Зоологический журнал