

Зоологический журнал, 2020, T. 99, № 2, стр. 178-188
Гнездование курганника (Buteo rufinus, Accipitriformes, Accipitridae) в искусственных лесных полосах и на одиночных деревьях в глинистой полупустыне Заволжья
А. В. Быков a, О. А. Бухарева a, *
a Институт лесоведения РАН
143030 Московская область, с. Успенское, Россия
* E-mail: buola@yandex.ru
Поступила в редакцию 12.11.2018
После доработки 08.05.2019
Принята к публикации 20.06.2019
Аннотация
Представлены результаты многолетнего мониторинга гнездования курганника в искусственных древесно-кустарниковых сообществах глинистой полупустыни Заволжья. Наблюдения, которые были начаты в 1980-е гг. и которые проводились в Палласовском районе Волгоградской области, а также в Западно-Казахстанской области (Республика Казахстан), показали, что дефицит природных объектов, пригодных для создания гнезд, сделал эту птицу зависимой от деятельности человека. Лесные полосы, созданные в 1950–1970-х гг., кардинально расширили потенциальную гнездовую территорию курганника, выведя его из локальных местообитаний озерных котловин и спирейников на зональную равнину. Здесь, вокруг транспортных путей и в окрестностях поселков на участках, максимально освоенных человеком, уже к началу 21 в. сформировалась стабильная, а затем избыточная по численности популяция этого канюка. В 2010-х гг. наметилось снижение числа гнездящихся птиц, причиной чего является уменьшение численности сусликов и развитие высокого травостоя, затрудняющего птицам добычу зверьков. Деградация искусственных насаждений, лишенных ухода, пока мало влияет на число гнездящихся пар. Птицы упорно выдерживают предпочитаемое расстояние между гнездами в 1.5–2 км. Уменьшение этого расстояния отмечается спорадически и указывает на дефицит гнездовых мест и высокую кормность угодья.
Курганник (Buteo rufinus Cretzschmar 1829), наряду со степным орлом (Aquila nipalensis Hodgson 1833), является самым массовым крупным хищником Прикаспийской низменности. В настоящее время, так как естественная древесно-кустарниковая растительность региона сведена или крайне нарушена (Быков, Бухарева, 2016), курганник гнездится почти исключительно в искусственных лесных насаждениях и на столбах линий электропередач (Линдеман, 1985; Линдеман и др., 2005, Быков, 2010). Сегодня, в условиях меняющегося климата и изменения форм хозяйствования, значительно сократилась протяженность линий электропередач и ускоряется деградация искусственных лесных насаждений. Это делает актуальной оценку различных типов таких насаждений с точки зрения их гнездовой пригодности для рассматриваемого вида, что необходимо для прогнозирования будущего этой птицы, внесенной в Красную книгу России.
МАТЕРИАЛ И МЕТОДИКА
Исследования проводили на базе Джаныбекского стационара Института лесоведения РАН, расположенного на границе России и Казахстана (Палласовский р-н, Волгоградская обл., Россия; Западно-Казахстанская обл., Республика Казахстан) (рис. 1). Район работ занимает равнинную территорию на западе заволжской части Прикаспийской низменности. Климат здесь отличается резкой атмосферной засушливостью и безводностью. Встречающиеся на равнине падины – плоские понижения микрорельефа глубиной до 1 м и площадью от 2–3 до 200 га с лугово-каштановыми почвами, подстилаемыми линзами пресных грунтовых вод, – занимают менее 15% площади и пригодны для создания искусственных насаждений и садов (Доскач, 1979; Роде, Польский, 1961). Большая часть равнины имеет комплексный почвенно-растительный покров, характерный для значительной территории Волго-Уральского междуречья. Преимущественно равнина занята солончаковыми солонцами и без применения специальных агротехнических мероприятий не пригодна для произрастания деревьев и кустарников (Большаков и др., 1983).
Рис. 1.
Район проведения мониторинга гнездования курганника в искусственных лесных насаждениях в 1982–2018 гг.
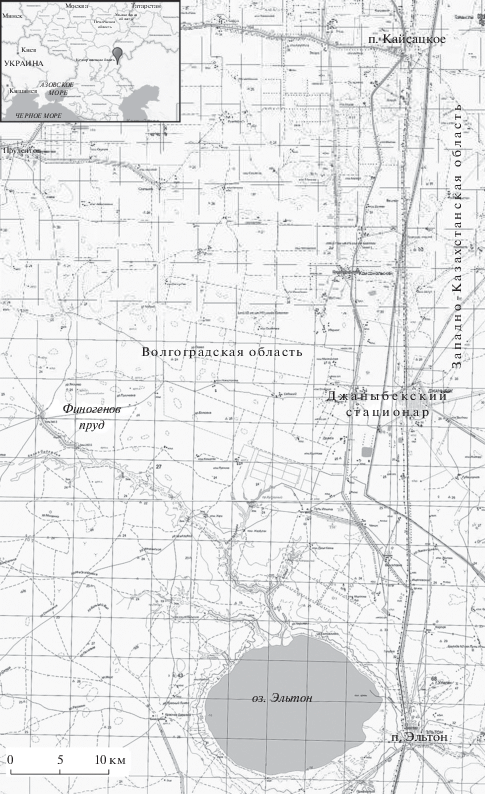
Учеты гнезд курганника начаты в 1980-х гг. и проводились в конце мая–начале июня на автомобильных и пеших маршрутах (табл. 1). Обследовали придорожные и полезащитные лесные полосы, специфические крайне разреженные линейные насаждения, прибрежные посадки около пресного водоема Финогенов пруд, заброшенные сады, одиночные деревья и точечные древесно-кустарниковые биогруппы, разбросанные среди комплексной степи на территории России и Казахстана.
Таблица 1.
Протяженность маршрутов, годы обследования и число жилых гнезд курганника в различных типах искусственных насаждений
| Типы насаждений | Годы обследований | Число жилых гнезд | |
|---|---|---|---|
| Придорожные лесные полосы на участке Кайсацкое–Эльтон, протяженность 9 км | 1986, 1988, 1990, 1992–1995, 1997–1999 | 40 | |
| Придорожные лесные полосы на участке Кайсацкое–Эльтон, протяженность 19 км | 2001–2018 | 134 | |
| Полезащитные лесные полосы, протяженность 10 км | 1986, 1990, 2001–2018 | 58 | |
| Разреженные линейные насаждения, протяженность 7.4 км | 2016–2018 | 11 | |
| Насаждения вдоль искусственного пруда, протяженность 4.2 км | 2002–2018 | 51 | |
| Одиночные деревья и биогруппы, 108 шт. | 1982–2001, 2004, 2009–2018 | 195 | |
| Заброшенные сады в падинах, 2 сада, суммарный периметр 1.2 км | 1982–2018 | 62 | |
| Насаждения Джаныбекского стационара | |||
| Массивные насаждения в падинах, 5 массивов, периметр 3 км | 1986–2008 | 35 | |
| Одно-двухрядные вязовые полосы протяженностью, км | 27 | 1986–1998 | 78 |
| 7 | 1999–2008 | 78 | |
| 20 | 2009–2018 | 78 | |
| Государственные лесные полосы, протяженностью 12 км | 1986–1998, 2009–2018 | 8 | |
| Всего жилых гнезд | 667 | ||
Как отдельный объект рассматривали насаждения Джаныбекского стационара РАН (табл. 1). Демаркация границы между государствами затруднила или сделала невозможным регулярное обследование некоторых объектов и проведение учетов на маршрутах, намеченных еще в 1980-х гг.
Всего за годы исследований с 1982 по 2018 г. прослежено 667 случаев гнездования курганника, в том числе 174 – в придорожных лесополосах (28 км), 58 – в полезащитных лесополосах (10 км), 11 – в разреженных линейных насаждениях (10 км), 51 – в насаждениях вдоль пруда (4.2 км), 195 – на одиночных деревьях и в биогруппах, 62 – в двух заброшенных садах. На Джаныбекском стационаре зарегистрировано 35 случаев гнездования в 5 лесных массивах среди падин, 78 – в одно-двухрядных вязовых лесополосах (54 км) и 8 – в Государственных лесных полосах общей протяженностью 12 км.
Искусственные лесные насаждения Джаныбекского стационара ИЛАН РАН созданы в исконно безлесной полупустыне и представляют собой комплекс облесенных участков разного типа, пахотных полей (до 1990-х гг.), залежей разного возраста и пространств комплексной степи на площади более 1612 га (рис. 2). Здесь можно выделить следующие типы насаждений:
Рис. 2.
Схема расположения научных объектов на территории Джаныбекского стационара (Научное наследие…, 2012): A – лесные насаждения на почвах солонцового комплекса, B – лесные насаждения на лугово-каштановых почвах больших падин, С – залежные участки, D – заповедный участок целины, E – место расположения усадьбы стационара; 1 – дендрарий на падине, 2 – дендрарий на почвах солонцового комплекса; 3 – “Производственный опыт”, 4 – “Новый опыт”, 5 – “Госфонд” агролесомелиоративные системы разной ширины (1100, 400 и 200 м соответственно), 6 – отрезок Государственной лесной полосы “Чапаевск–Владимировка”, состоящий из четырех полос (в направлении с запада), шириной 60 м, 7 – лесные массивы, 8 – сад, 9 – целинные падины, 10 – магистральный канал, 11 – пруд.
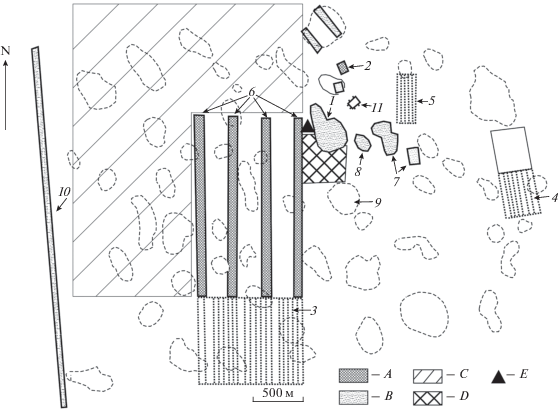
1. Агролесомелиоративные системы представлены тремя участками узкомассивных 1–2-рядных снегонакопительных кулис (на рис. 2 участки 3–5) из вяза мелколистного (Ulmus parvifolia Jacq. 1798) на почвах солонцового комплекса. Суммарная длина этих кулис составляет 3, 4 и 20 км соответственно. Эти системы представляют собой сельскохозяйственные поля, разделенные на межполосные пространства однорядными кулисами. До конца 1980-х гг. эти поля ежегодно распахивались.
2. Отрезок Государственной лесной полосы “Чапаевск–Владимировка”, состоящий из четырех полос (в направлении с запада на восток), шириной 60 м каждая. Расположены на почвах солонцового комплекса, посадки преимущественно 1951–1953 гг., сильно преобразованы рубками и реконструкциями. Сомкнутость крон от 0.6 до 1.0.
3. Лесные насаждения на лугово-каштановых почвах больших падин (на рис. 2 участок 6). Здесь сформировался сомкнутый полог, образовался мощный слой лесной подстилки, возник ярус подроста и подлеска из самосева древесных и кустарниковых пород. Сомкнутость крон 0.8–1.0.
Искусственные насаждения вне территории стационара возникли в разные годы и расположены среди комплексной равнины вне озерных депрессий.
Придорожные и полезащитные трехрядные лесные полосы на участке между поселками Кайсацкое и Эльтон, посадки 1960–1970 гг. Они состоят, преимущественно, из вяза мелколистного. Полосы распались на фрагменты, прогалы между которыми составляют десятки и сотни метров. С 1980-х годов в сохранившихся участках число деревьев сократилось почти вдвое и в настоящее время составляет около 160 на 1 км, а средняя высота насаждений снизилась с 8 до 6 м. На месте многих деревьев возникли многоствольные вегетативные биогруппы (Быков, Бухарева, 2017).
Разреженные насаждения линейного типа расположены к западу от дороги Палласовка–Эльтон и приурочены к бровкам заброшенных или недостроенных каналов и к глубоким рвам. Они возникли как побочный результат антропогенной деятельности и представляют собой самосев различных пород и возраста. Число деревьев или биогрупп не превышает 20 на 1 км (Быков, Бухарева, 2017). Насаждения этого типа пока немногочисленны, но их число постепенно растет. Они рассматриваются нами, как аналог насаждений, в которые без надлежащего ухода превратятся лесные полосы через 10–15 лет.
Прибрежные насаждения на берегу пресного Финогенова пруда расположены в верховьях р. Хара, долина которой перекрыта высокой дамбой. Насаждения ленточные, в основном приуроченные к восточному берегу пруда, отдельные самосевные участки есть на всем протяжении береговой линии и на коротких, впадающих в водоем, балочках. Они состоят из старых деревьев ветлы (Salix alba L. 1753) и тополя белого (Populus alba L. 1753) с самосевными и разновозрастными вязами мелколистными, единичными яблонями (Malus praecox (Pall.) Borkh. 1803), подроста перечисленных пород и подлеском из ивы каспийской (Salix caspica Pall. 1789) и других видов. Возраст не менее 70 лет, высота старых деревьев до 17 м. При ширине полосы насаждения от 7 до 20 м ее длина составляла приблизительно 4.2 км. В 2008 г. все насаждение было пройдено сильным низовым пожаром, в результате которого много старых деревьев погибло на следующий год. Это единственное местообитание, которое мы можем рассматривать как своего рода искусственный аналог утраченных байрачных сообществ озерных котловин (Быков, Бухарева, 2017).
Одиночные деревья и биогруппы обследовались к западу от дороги Палласовка–Эльтон и в Джаныбекском р-не Западно-Казахстанской обл. Они разбросаны среди комплексной степи. Чаще всего они растут на месте покинутых хозяйств – базищах. Здесь возникают своеобразные местообитания с рассолонцованными или рассолеными почвами, причем верхние почвенные горизонты удерживают значительное количество воды (Динесман, 1960). Около старых колодцев иногда сохраняются саженые деревья: тополя разных видов, одичавшие груши и яблони, вяз мелколистный, редко другие виды. Обычно это единичные деревья, но иногда сомкнутые биогруппы из 2–4 стволов одного вида или разных видов. Под такими деревьями обычно отдыхает скот, и травяной покров сбит, что спасает такие деревья во время степных пожаров. Чаще всего сгорают тополи, которые всегда имеют много сухих ветвей, и под ними накапливается отпад. На месте сгоревших тополей появляется корневая поросль.
Заброшенные сады обследовались на территориях России и Казахстана. Они приурочены к падинам среди комплексной полупустыни и, в основном, насажены в 1920–конце 1930-х гг. Здесь сохраняются отдельные яблони, груши, заросли вишни, но в целом территория поросла самосевным вязом мелколистным, лохом узколистным (Elaeagnus angustifolia L. 1753) и тополем белым. Высота отдельных деревьев может достигать 7–12 м. Через каждый сад проходит грунтовая дорога, которая разделяет его пополам. Периметр обследованных садов 1.2 км.
РЕЗУЛЬТАТЫ
Гнездование курганника в разнообразных искусственных насаждениях Джаныбекского стационара
Искусственные насаждения Джаныбекского стационара состоят более чем из 15 отдельных групп насаждений, разбросанных на значительной площади. Однако число гнездящихся курганников здесь только в 1993 г. превысило 10 пар в год (табл. 2). Это связано с тем, что покидая гнездо, слеток этого вида должен спланировать на открытую поверхность, иначе, попав в густой кустарник, он рискует зависнуть на ветвях, где погибнет. Поэтому курганник не строит гнезда в сомкнутых насаждениях и на участках с развитым подлеском (Быков, Бухарева, 2018). Из жилых 116 гнезд, отмеченных здесь, 95% находилось на внешних границах территории стационара со стороны открытых пространств степи.
Таблица 2.
Абсолютное число жилых гнезд курганника в насаждениях Джаныбекского стационара
| Год | Массивные насаждения в падинах, 5 падин | Узкие вязовые полосы | Госполосы | ||
|---|---|---|---|---|---|
| госфонд, 3 км | новый опыт, 4 км | производственный опыт, 20 км | |||
| 1986 | 2 | 1 | 1 | 2 | 0 |
| 1987 | Нет данных | 1 | 2 | 2 | 1 |
| 1988 | 2 | 1 | 1 | 2 | 0 |
| 1989 | Нет данных | 1 | 1 | 2 | 0 |
| 1990 | 2 | 1 | 0 | 2 | 0 |
| 1991 | Нет данных | 1 | 1 | 1 | 0 |
| 1992 | 2 | 1 | 0 | 1 | 0 |
| 1993 | 3 | 1 | 1 | 2 | 0 |
| 1994 | 2 | 1 | 1 | 2 | 0 |
| 1995 | 2 | 1 | 1 | 2 | 0 |
| 1996 | Нет данных | 1 | 1 | 2 | 0 |
| 1997 | 2 | 1 | 1 | 2 | 0 |
| 1998 | 2 | 1 | 1 | 2 | 0 |
| 1999 | 2 | 1 | 2 | Нет данных | Нет данных |
| 2000 | Нет данных | 1 | 1 | То же | То же |
| 2001 | 2 | 1 | 1 | То же | То же |
| 2002 | 2 | 1 | 1 | То же | То же |
| 2003 | 3 | 1 | 2 | То же | То же |
| 2004 | 2 | 1 | 0 | То же | То же |
| 2005 | 1 | 1 | 1 | То же | То же |
| 2006 | 1 | 1 | 1 | То же | То же |
| 2007 | 1 | 1 | 1 | То же | То же |
| 2008 | 2 | 1 | 1 | То же | То же |
| 2009 | Нет данных | Нет данных | Нет данных | 1 | 0 |
| 2010 | То же | То же | То же | 1 | 0 |
| 2011 | То же | То же | То же | 1 | 0 |
| 2012 | То же | То же | То же | 0 | 1 |
| 2013 | То же | То же | То же | 0 | 1 |
| 2014 | То же | То же | То же | 1 | 0 |
| 2015 | То же | То же | То же | 0 | 1 |
| 2016 | То же | То же | То же | 0 | 1 |
| 2017 | То же | То же | То же | 0 | 1 |
| 2018 | То же | То же | То же | 0 | 1 |
В массивных насаждениях падин курганник занимает не просто опушечные деревья, а те из них, которые заметно выступают за пределы массива, при этом всегда со стороны, обращенной к степи, а не к другим насаждениям. Суммарный периметр массивных насаждений около 4 км. Одно гнездо приходится здесь на 2.6 км опушечной линии.
В длинных и широких госполосах с суммарным периметром 24.5 км курганник практически не гнездится. Лишь одно гнездо в южном конце одной из госполос периодически использовалось как сменное для пары, обычно гнездившейся в наружной вязовой полосе ближайшей агролесомелиоративной системы на расстоянии 250 м. Курганники, внедряющиеся в межполосные пространства этих насаждений, немедленно изгоняются многочисленными здесь черными коршунами (Milvus migrans Boddaert 1783) и сороками (Pica pica L. 1758).
Из 116 гнезд, отмеченных на территории стационара, больше половины (64%) было приурочено к узкомассивным агролесомелиоративным системам. Здесь узкие вязовые полосы сконцентрированы на небольших по площади прямоугольниках. Расстояние между полосами составляет 40–90 м, суммарная протяженность – 27 км. Казалось бы, такие условия максимально соответствуют требованиям курганника. Однако все гнезда располагались исключительно на вязах наружного периметра агролесомелиоративных систем (7.2 км), где одно гнездо приходится на 2 км. Очевидно, что, несмотря на обилие деревьев, пригодных для строительства гнезд, компактная агролесомелиоративная система воспринимается птицами как своего рода массивное насаждение.
Показательно, что на самом узком агролесомелиоративном массиве (№ 5 на рис. 2) максимальное возможное расстояние между гнездами составляло около 0.54 км и за 23 года наблюдений здесь не отмечали более одного гнезда в год. На массиве № 4 (рис. 2) это расстояние составляет 0.64 км и за тот же срок по 2 гнезда в год отмечали 3 раза. На самом крупном массиве № 3 (рис. 2), где возможное расстояние между гнездами 1.4 км, по два гнезда в год отмечали в течение почти половины исследуемого периода.
Гнездование курганника в искусственных насаждениях среди комплексной равнины вне территории стационара
Условия гнездования курганника в заброшенных садах, приуроченных к падинам среди комплексной полупустыни, такие же, как в массивных насаждениях Джаныбекского стационара. Они отличаются лишь тем, что массивные насаждения стационара соседствуют с другими насаждениями, а рассматриваемые сады, разбросанные среди безлесной комплексной равнины, удалены от любых насаждений на десятки километров. Кормность окружающей территории обычно хорошая. В каждом саду почти всегда гнездится одна пара птиц. За 31 год наблюдений по две гнездящиеся пары отмечались в садах лишь четыре раза, причем это событие было приурочено к последовательному ряду лет (1995–1998). Причину этого мы указать не можем, так как кормность окружающей территории в период гнездования заметно не менялась. Однако именно в эти годы на значительных территориях отмечалось атипично позднее залегание сусликов – молодые особи были активны до конца сентября.
Для гнездования курганника оптимальны узкие линейные и особенно точечные насаждения – одинокие деревья и биогруппы, разбросанные среди комплексной степи. Свои гнезда эта птица строит даже на 5–6-летней корневой поросли сгоревших тополей, которая плохо выдерживает тяжесть гнезда. Дважды мы отмечали, что птицы насиживали яйца в сильно наклонившемся гнезде, а затем, кормили выпавших птенцов на земле. В одном из этих случаев успешной была вторая кладка, так как первое гнездо с яйцами было снесено ветром.
Единственным препятствием для гнездования курганника на одиноких деревьях является занятие их степным орлом и могильником (Aquila heliaca Savigny 1809). С начала 1980-х до второй половины 2000-х гг. курганник занимал половину или две трети одиноких деревьев и биогрупп, в остальных случаях здесь гнездились орлы. Затем доля гнезд курганника стала расти и в 2010-х гг. достигла 100%. Известно, что в полидоминантных кустарниковых зарослях озерных депрессий уже с 1980-х гг. курганники заняли постройки степных орлов, гнездившихся в них более 10 лет. Орлы стали строить гнезда на земле, часто недалеко от покинутых гнезд, но вне границ кустарникового насаждения (Быков, Бухарева, 2018). Возможно, это действительно объясняется тем, что в последние десятилетия курганники прилетают раньше орлов, первыми занимают и удерживают гнездовые участки (Пименов, Белик, 2015). К сожалению, в 2000-х гг. среди учетных одиноких деревьев на наших постоянных маршрутах высокие тополя, на которых гнездился могильник, уже отсутствовали, и мы не можем представить новые данные о взаимоотношениях этих видов.
Сравнение показателей средней численности гнезд в придорожных и полезащитных полосах показывает, как эта численность зависит от кормности окружающей территории. Эти полосы близки по породному составу, мозаичности и возрасту. Но полезащитные полосы еще в начале 1990-х гг. были окружены пахотными землями. Суслики (Spermophilus pygmaeus Pall., 1778), основной кормовой объект курганника (Линдеман, 1985; Пименов, Белик, 2015), вблизи полезащитных полос малочисленны и, к тому же, труднодоступны из-за высокого травяного покрова. Придорожные полосы окружены целинными или старопахотными участками, и вокруг них сосредоточены населенные пункты. Выпас и прогон скота у дорог интенсивен, травяной покров разрежен, суслика много, и он доступен для птиц (Пименов, Белик, 2015). В результате в придорожных лесных полосах гнезд курганника больше почти в 1.5 раза.
Разреженные линейные насаждения, в которых расстояние между деревьями или между кустарниковыми биогруппами составляет десятки метров, характеризуются высокой гнездовой плотностью. Количество гнезд курганника здесь ограничивается небольшой протяженностью таких насаждений и в какой-то мере высокой плотностью гнездования степного орла (в среднем 1 пара на 1 км).
Финогенов пруд окружен непаханой комплексной равниной и, частично, старыми залежами, уже заселенными сусликом. Выпас вокруг умеренный, в отдельные годы сильный. В самом насаждении много мест, пригодных для строительства гнезд. Гнездовая плотность на километр насаждения, как и на одиноких деревьях, высокая. В то же время, как и в остальных типах искусственных насаждений, с начала–середины 2010 гг. здесь отмечается тенденция к уменьшению частоты гнездования курганника (рис. 3), но, чтобы подтвердить достоверность этого, необходим бо́льший срок наблюдений.
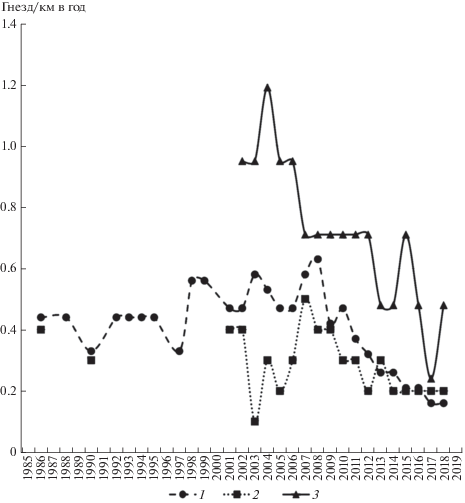
ОБСУЖДЕНИЕ
В рассматриваемом регионе курганники редко строят гнезда на земле (Линдеман, 1985; Пименов, Белик, 2015; Быков, Бухарева, 2018). Они предпочитают гнездиться на деревьях и высоких кустах, а при их отсутствии птицы выбирают для этой цели различные объекты, в той или иной степени приподнятые над поверхностью: столбы и вышки, крыши и остатки стен заброшенных строений, редко стога, кучи хвороста и, даже, расположенные на земле высокие гнезда степных орлов.
На протяжении двух столетий все типы естественной древесно-кустарниковой растительности региона подвергались мощному антропогенному давлению (Динесман, 1960; Быков и др., 2013, 2013а). В результате возник серьезный дефицит объектов, пригодных для гнездования этого канюка. Уже в первой половине 20 в. курганник в глинистой полупустыне был редок, а его гнездование вплоть до конца 1950-х гг. не подтверждено (Линдеман и др., 2005). Однако, по крайней мере с середины 20 в. этот канюк регулярно гнездится по границам региона – в Волго-Уральских песках и в долинах пресных речек (Шевченко и др., 1978; Ходашова, 1960). Во второй половине 20 столетия в Заволжье значительно возросла протяженность линий электропередач, и были созданы многочисленные искусственные лесные насаждения. В 1960–1970-х гг. стала восстанавливаться численность птиц байрачного комплекса, увеличилось число гнездящихся степных орлов, на высокоствольных участках насаждений сформировались летние скопления негнездящихся крупных хищных птиц, численностью от многих десятков до нескольких сотен экземпляров (Линдеман и др., 2005; Линдеман, Быков, 2010, Быков, Бухарева, 2015, 2016). Благодаря появлению многочисленных мест, пригодных для гнездования, с 1970-х годов курганник становится обычной гнездящейся птицей полупустынного Заволжья (Линдеман, 1985).
Лесные полосы и другие искусственные насаждения неравномерно распределены по территории региона. Их больше в западной, российской части Заволжья, но в целом они образуют экологический каркас специфических местообитаний, обеспечивший расселение курганника по всей территории глинистой полупустыни. Появление этого каркаса вывело значительную группу видов из узколокальных местообитаний озерных котловин и спирейников падин на зональную комплексную равнину и, что необходимо подчеркнуть, прежде всего, на те ее участки, которые активно используются человеком (Быков, Бухарева, 2016).
К концу 20 в. искусственные насаждения региона достигли критического возраста, а уход за ними прекратился. Деградация насаждений выражается в разреженности крон, суховершинности, усыхании, как отдельных деревьев, так и протяженных участков лесополос, формировании кустарникоподобных биогрупп порослевого происхождения. Участившиеся пожары уничтожают значительные участки лесных полос, старые сады и одинокие деревья. Часть сгоревших деревьев восстанавливается корневой порослью, на которой менее чем через 10 лет появляются первые гнезда птиц. Естественно, что на этот срок курганники лишаются ряда своих гнездовых угодий. Процесс частичного восстановления древесно-кустарниковой растительности замедляется или останавливается из-за повторных пожаров, интенсивной пастьбы скота, распашки сгоревших насаждений под противопожарные полосы и др. Процесс изреживания насаждений может затянуться на многие годы, но, все же, он конечен. Деревья и кустарники, необходимые для гнездования курганника, рано или поздно исчезнут, что приведет к исчезновению этой птицы. Тенденция такого рода уже проявилась в естественных древесно-кустарниковых сообществах региона (Быков, Бухарева, 2018). Конечно, по различным антропогенным нарушениям (рытвинам, старым колеям, ямам, рвам и т.п.) появляется самосев различных древесных и кустарниковых пород, но его количество минимально и будет уменьшаться по мере исчезновения деревьев, дающих семенной материал.
Пока еще деградация искусственных насаждений затрагивает курганника относительно мало. Мы видим, что крайне разреженные насаждения характеризуются оптимальной гнездовой плотностью птиц, и увеличение мозаичности придорожных и полезащитных лесных полос не критично для вида.
В 1980–1990-х гг. в искусственных насаждениях плотность гнездования курганника колебалась в незначительных пределах и, очевидно, была близка к оптимальной. По наблюдениям Линдемана (1985), в 1960–1970-х гг. в линейных насаждениях расстояние между занятыми гнездами курганника составляло не менее 1 км, а обычно было большим. В 2000-х гг. расстояние между некоторыми гнездами составляло уже 0.5–1 км, но чаще было 3–5 км, что зависело от наличия гнездопригодных мест и кормовой базы. Именно в эти годы учащается встреча гнезд на земле, все чаще отмечается сокращение расстояния между гнездами (Пименов, Белик, 2015). В естественных насаждениях Эльтонской котловины курганники гнездились не менее чем 1.5 км друг от друга (Быков, Бухарева, 2018). Можно заключить, что расстояние между гнездами менее 1 км является критическим, и столь близко друг от друга птицы поселяются в условиях крайнего дефицита пригодных для гнездования мест. Вероятно, расстояние около 2 км является оптимальным, а варьирование этого значения определяется конкретными особенностями насаждения и, прежде всего, наличием открытых пространств непосредственно рядом с местом гнездования. Именно поэтому значительные по облесенной площади, но массивные насаждения Джаныбекского стационара в целом неблагоприятны для гнездования курганника. Они избыточны по соотношению площади, занятой насаждениями, к площади открытых пространств между ними.
Уже в начале 2010-х гг. плотность гнездования вида стала снижаться. Мы считаем, что это объясняется не столько деградацией самих насаждений, сколько изменением форм хозяйственного воздействия на природные системы и климатическими причинами. В последние десятилетия снизилась пастбищная нагрузка, пахотные земли превратились в залежи, увеличилась увлажненность территории и изменилось распределение осадков по сезонам. С 1980-х гг. отмечается мезофитизация травяного покрова и, даже, началось зарастание солонцов (Оловянникова, 2004; Новикова и др., 2010; Сапанов, Сиземская, 2015). Высокий и густой травяной покров негативно сказывается не только на возможности добычи суслика хищными птицами, но и на самом существовании зверька. В последнее десятилетие малый суслик уходит с территории. На ряде участков севернее п. Кайсацкое он уже редок или отсутствует, кроме того, этот грызун практически исчез из озерных депрессий региона. Относительно высокая численность малого суслика сохраняется в комплексной степи, но особенно она высока вблизи поселков и скотоводческих точек, т. е. там, где травяной покров сбит скотом.
Уничтожение естественных древесно-кустарниковых сообществ на территории глинистой полупустыни привело к тому, что курганники региона превратились в своего рода синантропов. Их существование зависит от наличия деревьев, посаженных человеком, размера и структуры искусственных насаждений, их местоположения, частоты пожаров, причиной большинства которых является человек, и интенсивности выпаса скота. Почти все искусственные насаждения находятся в населенных районах, и лишь одинокие деревья или старые сады часто удалены от современных поселений на десятки километров. Разумеется, в период размножения курганники зависят от динамики численности суслика, но и здесь, как говорилось выше, максимальная численность зверька отмечается в местах сильного выпаса, что возвращает нас к вопросу о синантропизации этой птицы.
Для сохранения природного разнообразия региона необходимо вернуться к вопросу о создании системы колочно-западинного ландшафта (Быков и др., 1993). Элементы такого ландшафта определяют сохранение представителей именно зональной фауны – крупных хищных птиц. Необходима и разработка стратегии восстановления уничтоженной и трансформированной человеком естественной древесно-кустарниковой растительности на территории глинистой полупустыни России и Казахстана (Быков, Бухарева, 2016).
ЗАКЛЮЧЕНИЕ
Курганники – птицы открытых пространств. Они не охотятся с присады и сидят на дереве, отдыхая, прячась в тени. Они не нуждаются в лесной обстановке и гнездятся на деревьях лишь потому, что в данном регионе именно деревья обеспечивают птицам безопасность гнезда. В глинистой полупустыне Заволжья уничтожение естественных древесно-кустарниковых сообществ, приуроченных к различного рода депрессиям, привело к тому, что существование популяции курганника стало зависеть от факторов, инициируемых человеком. Появление в 1950–1960-х гг. экологического каркаса искусственных насаждений и многочисленных столбов электропередач расширило потенциальную гнездовую территорию птицы на всю комплексную равнину. В результате использования всех возможных гнездовых ниш к началу 21 в. сформировалась стабильная, а затем избыточная по численности популяция. Флуктуация условий, вызванных антропогенными и климатическими факторами, уже в 2010-х гг. обусловливает снижение численности гнездящихся курганников.
Предпочитаемое курганником расстояние между гнездящимися парами – не менее 1.5 км. Поэтому на территории региона птицы охотно гнездятся на одиночных деревьях или локальных древесно-кустарниковых биогруппах, удаленных друг от друга. Препятствием для их гнездования здесь может служить лишь деятельность человека или занятие местообитания другими видами крупных хищных птиц. Узкие придорожные и полезащитные лесные полосы – наиболее распространенный тип искусственных насаждений региона – для курганника избыточны по количеству деревьев, пригодных для гнездования. Здесь, если позволит длина полосы, он будет гнездиться в 1.5–2 км от соседа. Лишь при дефиците протяженных полос и отсутствии иных мест, пригодных для гнездования, расстояние между гнездами будет уменьшаться. В этих условиях речь идет не о дефиците собственно деревьев, пригодных для строительства гнезда, а о “дефиците” расстояния между деревьями. Именно поэтому деградация искусственных насаждений, усиливающаяся с каждым годом, пока мало влияет на число гнездящихся пар. Сейчас, при уменьшении числа сусликов и исчезновении их на некоторых территориях, возможность добычи корма является главным фактором, регулирующим численность этого канюка. Лишь в условиях дефицита мест, пригодных для постройки гнезда, это расстояние уменьшается.
Список литературы
Большаков А.Ф., Эрперт С.Д., Шейнин Л.Б., 1983. Пути сельскохозяйственного освоения полупустыни. М.: Наука. 72 с.
Быков А.В., 2010. Значение древесно-кустарниковой растительности для позвоночных животных глинистой полупустыни Заволжья // Аридные экосистемы. Т. 16. № 5 (45). С. 90–97.
Быков А.В., Бухарева О.А., 2015. Изменения в населении лесных и дендрофильных птиц глинистой полупустыни Волго-Уральского междуречья за 60 лет // Поволжский экологический журнал. № 2. С. 148–158.
Быков А.В., Бухарева О.А., 2016. Современное состояние кустарниковой растительности байрачного типа в окрестностях оз. Эльтон // Аридные экосистемы. Т. 22. № 1 (66). С. 70–76.
Быков А.В., Бухарева О.А., 2017. Гнездящиеся птицы байрачного комплекса в лесных полосах глинистой полупустыни Заволжья // Лесоведение. № 3. С. 221–227.
Быков А.В., Бухарева О.А., 2018. Гнездование курганника (Buteo rufinus, Accipitriformes, Accipitridae) в естественных древесно-кустарниковых сообществах глинистой полупустыни Заволжья // Зоологический журнал. Т. 97. № 5. С. 582 – 590.
Быков А.В., Бухарева О.А., Колесников А.В., 2013. Воздействие пожаров на естественные терновники озерных депрессий Северо-Запада Прикаспийской низменности // Лесоведение. № 2. С. 31–37.
Быков А.В., Колесников А.В., Шадрина М.Б., Бухарева О.А., Шабанова Н.П., 2013а. Воздействие пожаров на естественные тамарисковые сообщества озера Булухта (Северный Прикаспий) // Лесоведение. № 6. С. 3–9.
Быков А.В., Оловянникова И.Н., Сапанов М.К., 1993 Роль зоогенных факторов при создании колочно-западинного ландшафта в глинистой полупустыне Заволжья // Лесоведение. № 6. С. 27–33.
Динесман Л.Г., 1960. Изменение природы северо-запада Прикаспийской низменности. М.: Изд. АН СССР. 160 с.
Доскач А.Г., 1979. Природное районирование Прикаспийской полупустыни. М.: Наука. 142 с.
Линдеман Г.В., 1985. Курганник (Buteo rufinus Cretzschm.) в междуречье Волги и Урала // Бюллетень Московского общества испытателей природы. Отдел биологический. Т. 90. Вып. 6. С. 27–37.
Линдеман Г.В., Абатуров Б.Д., Быков А.В., Лопушков В.А., 2005. Динамика населения позвоночных животных Заволжской полупустыни. М.: Наука. 252 с.
Линдеман Г.В., Быков А.В., 2010. Летние скопления негнездящихся орланов-белохвостов Haliaeetus albicilla (L.) в заволжской глинистой полупустыне // Бюллетень Московского общества испытателей природы. Отдел биологический. Т. 115. Вып. 5. С. 28–32.
Научное наследие Джаныбекского стационара. 2012. Отв. ред. Сапанов М.К. М.: Товарищество научных изданий КМК. 97 с.
Новикова Н.М., Вышивкин А.А., Шадрина, М.Б., Бухарева О.А., 2010. Изменения растительности солонцового комплекса Джаныбекского стационара и его окрестностей при разной интенсивности выпаса // Аридные экосистемы. Т. 16. № 5. С. 98–110.
Оловянникова И.Н., 2004. Динамика продуктивности растительного покрова в Заволжской глинистой полупустыне // Ботанический журнал. Т. 89. С. 1122–1137.
Пименов В.Н., Белик В.П., 2015. Курганник в Волгоградском Заволжье // Стрепет. Т. 13. Вып. 1. С. 120–144.
Роде А.А., Польский М.Н., 1961. Почвы Джаныбекского стационара, их морфологическое строение, механический и химический состав и физические свойства // Труды Почвенного института. М.: Изд-во АН СССР. Т. 56. С. 3–214.
Сапанов М.К., Сиземская М.Л., 2015. Изменение климата и динамика целинной растительности в Северном Прикаспии // Поволжский экологический журнал. № 3. С. 307–320.
Ходашова К.С., 1960. Природная среда и животный мир глинистой полупустыни Заволжья. М.: Изд-во АН СССР. 131 с.
Шевченко В.Л., Гаврилов Э.И., Наглов В.А., Федосенко А.К., Татаринова О.М., 1978. Об орнитофауне Волжско-Уральского междуречья // Труды Института зоологии АН КазССР. Т. 38. Алма-Ата. С. 99–114.
Дополнительные материалы отсутствуют.
Инструменты
Зоологический журнал