

Зоологический журнал, 2020, T. 99, № 2, стр. 189-202
К изучению размножения обыкновенной бурозубки (Sorex araneus) на северной периферии ареала
Э. В. Ивантер *
Петрозаводский государственный университет
185910 Петрозаводск, Россия
* E-mail: Ivanter@petrsu.ru
Поступила в редакцию 26.06.2018
После доработки 28.01.2019
Принята к публикации 15.05.2019
Аннотация
На основе многолетних экспедиционных и стационарных исследований на таежном Северо-Западе России (1965‒2017 гг.) охарактеризовано размножение обыкновенной бурозубки, в том числе особенности, связанные с ее обитанием у северных границ ареала: сроки, половые циклы, ход и интенсивность репродукции, участие в размножении отдельных возрастных групп, вариации величины выводка и др. Согласно полученным данным, в этих условиях у данного вида размножаются в основном перезимовавшие особи с конца апреля по середину сентября, принося за это время 2, реже 3 выводка из 3‒11, в среднем ‒ 6.4 детеныша.
Несмотря на неослабевающий интерес к изучению обыкновенной бурозубки, в том числе и в связи с ее многочисленностью и важным биоценотическим значением, экология и особенно размножение этого вида по-прежнему остаются исследованными совершенно недостаточно. Настоящее сообщение, основанное на многолетних (1965‒2017 гг.) экспедиционных и стационарных исследованиях, охвативших территорию Карелии и прилежащих регионов Восточной Фенноскандии и представляет собой попытку восполнить этот существенный пробел.
МАТЕРИАЛ И МЕТОДИКА
Материалом для настоящего сообщения послужили многолетние (1965‒2017 гг.) сборы зверьков изучаемого вида, отлавливаемых двумя основными методами: ловушко-линиями (плашки Геро) и ловчими 30-метровыми канавками. Первым методом было отработано 243 580 ловушко-суток и добыто 11 570 обыкновенных бурозубок, вторым ‒ 5400 канавко-суток и отловлено 3072 зверька. Таким образом, общее число добытых и проведенных через анализ обыкновенных бурозубок составило 14 650 экз.
Для изучения размножения обыкновенной бурозубки всех отловленных зверьков этого вида подразделяли на две возрастные группы. В качестве основных возрастных критериев использовали форму (конфигурация и состояние швов) черепа и степень стертости зубов, а в качестве второстепенных (вспомогательных) ‒ особенности снашивания волосяного покрова на хвосте, лапах и ушных раковинах. Кроме того, в сомнительных случаях привлекали и некоторые другие показатели, в частности размеры тимуса, который, как известно, полностью инволюирует к осени первого года жизни и у перезимовавших зверьков практически не выражен.
Особи первой возрастной группы (сеголетки) характеризуются высокой сжатой с боков черепной коробкой (череп “вздутый”) и отсутствием гребней. Швы между костями хорошо заметны. Зубы имеют острые вершины и гребни. Эмаль коронок не стерта. Половая система, как правило, недоразвита. Тимус хорошо выражен, его абсолютный вес 15‒250 мг. Хвост равномерно покрыт волосами (“пушистый”). Волосяной покров на лапках и ушных раковинах хорошо развит и почти не имеет следов снашивания. В подавляющем большинстве случаев это неполовозрелые зверьки (subadultus).
Зверьки второй возрастной группы (перезимовавшие) отличаются утолщенным черепом с хорошо развитыми гребнями и слабо заметными швами, последние имеют зигзагообразный рисунок. Вершины промежуточных зубов в результате снашивания округленные, притупленные; коренные зубы уплощены, с тупыми конусами и гребнями. Пигментация на зубах почти не заметна. Волосяной покров хвоста, лап и ушных раковин сильно сношен, а местами совсем отсутствует (“лысины”). Это половозрелые зверьки (adultus).
Различаются возрастные группы и по строению половой системы (табл. 1). У неполовозрелых самцов семенники очень малы и, как правило, округлые форму (диаметр около 2 мм, вес не более 10‒11 мг). Придатки семенника и семенные пузырьки недоразвиты. У половозрелых, перезимовавших бурозубок семенники значительно крупнее (вес двух семенников обычно более 100 мг), имеют продолговатые (длина 5‒11, ширина 4‒7 мм) и хорошо заметны даже у невскрытых зверьков (в виде двух вздутий на нижней стороне тела). Придатки семенника, простатическая и куперова железы и семенные пузырьки очень хорошо выражены; последние наполнены спермой и сильно вздуты. У самцов, участвующих в размножении, кроме того, увеличиваются боковые кожные железы (7 × 3 мм), появление которых принимается в качество симптома полового созревания (Дунаева, 1955; Borowski, 1964; Spilzenberger, 1966).
Таблица 1.
Характеристика половой системы обыкновенной бурозубки
| Показатели | Зимовавшие | Прибылые |
|---|---|---|
| Самцы | ||
| Число исследованных зверьков | 533 | 2401 |
| Длина семенника, мм | 7.5 (1.0‒12.9) | 1.2 (0.8‒8.5) |
| Длина семенных пузырьков, мм | 8.9 (2.5‒16.2) | 8.3 (1.0‒13.9) |
| Вес двух семенников, мг | 244.1 (50‒308) | ‒ |
| Самки | ||
| Число исследованных зверьков | 114 | 563 |
| Суммарная длина рогов матки, мм | 26.8 (4.6‒73.7) | 9.5 (2.8‒25.0) |
| Длина влагалища (тела матки), мм | 14.7 (2.3‒29.3) | 6.6 (1.7‒12.4) |
У молодых неполовозрелых самок матка полупрозрачная, тонкая и короткая; общая длина ее рогов составляет в среднем 9.52 (2.8‒22.0) мм, тела 6.6 (1.7‒22.4) мм. У перезимовавших половозрелых самок матка набухает, утолщается и становится шире, стенки ее гиперемированы, рыхлы. В длину она увеличивается почти втрое: суммарная длина ее рогов 26.8 (4.6‒73.7), тела 14.7 (2.3‒29.3 мм). У взрослых зверьков тело матки составляет в среднем 65% от длины рогов, а у молодых ‒ 75%. При этом соотношение (индекс) тела и длины рогов матки у обыкновенной бурозубки несколько иное, чем у других представителей рода Sorex. Например, у средней (Sorex caecutiens) и малой (S. minutus) бурозубок эти индексы составляют соответственно 40‒50 и 35‒46%. Таким образом, у обыкновенной бурозубки рога матки относительно короче, а тело длиннее, чем у других видов.
У кормящих или ранее кормивших самок соски обсосаны и волосяной покров вокруг них сильно вытерт (особенно если самка кормит второй выводок), но у первых млечные железы лактируют, а у вторых ‒ нет. У беременных на ранних стадиях хорошо заметны розоватые желтые тела в яичниках, позднее ‒ эмбрионы на рогах матки. Матка сильно увеличена в длину и ширину. Плацентарные пятна образуются, но очень быстро (уже на 2‒3-й день после родов) исчезают (Brambell, 1935; Дунаева, 1955). Иногда удавалось определить и недавно оплодотворенных самок, признаки беременности у которых еще не были выражены. В матке у таких самок бывает заметна мутная беловатая жидкость ‒ сперма.
Сроки и интенсивность размножения обыкновенной бурозубки определялись на основании предпринятого нами специального гистологического исследования полового цикла самцов и самок (Ивантер, 1975). Остальные исследования проводились по общепринятым методикам, неоднократно описанным в наших предыдущих публикациях (Ивантер, Ивантер, 1968; Ивантер и др. 1974; Ивантер, 1975).
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Изучение сперматогенеза самцов показало, что весь цикл превращений, происходящих в гонадах бурозубок, можно подразделить на три периода: покой, пассивный сперматогенез и активный сперматогенез.
Период покоя продолжается с момента рождения (май‒июнь) и по январь включительно. В это время в семенных канальцах присутствуют только клетки Сертоли и спермогонии (табл. 2, рис. 1а). Вес и размеры семенников минимальные (вес 2 testis в среднем 3,5 мг; длина семенника ‒ 1.5 мм), диаметр семенных канальцев 24‒41 мкм.
Таблица 2.
Состояние эпителия семенных канальцев на протяжении жизненного цикла обыкновенной бурозубки
| Клеточные элементы | Месяц | ||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| VI | VII | VIII | IX | X | XI | XII | I | II | III | IV | V | VI | VII | VIII | IX | X | |
| Клетки Сертоли | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + |
| Сперматогонии | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + |
| Сперматоциты I порядка | – | – | – | – | – | – | – | – | + | + | + | + | + | + | + | + | + |
| Сперматоциты II порядка | – | – | – | – | – | – | – | – | – | + | + | + | + | + | + | + | + |
| Сперматиды | – | – | – | – | – | – | – | – | – | + | + | + | + | + | + | + | + |
| Сперматозоиды | – | – | – | – | – | – | – | – | – | – | + | + | + | + | + | + | + |
| Средний диаметр семенных канальцев, мкм | 41 | 41 | 35 | 27 | 28 | 24 | ? | 26 | 31 | 96 | 189 | 178 | 157 | 161 | 169 | ? | ? |
Рис. 1.
Поперечные срезы извитых канальцев семенника обыкновенной бурозубки: а – состояние покоя, б – пассивный сперматогенез, в – активный сперматогенез. Микрофото, ×400.
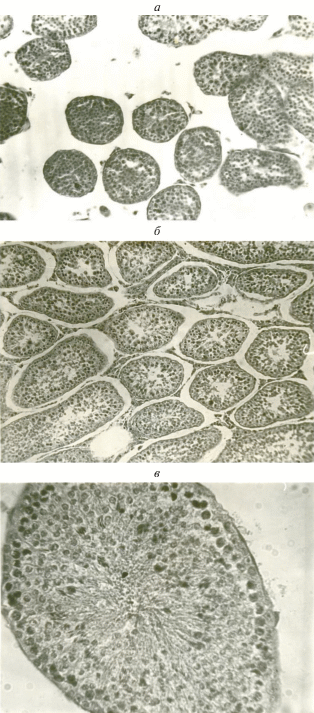
С февраля и до середины апреля у самцов обыкновенной бурозубки проходит пассивный сперматогенез, т.е. наступает стадия подготовки к продуцированию спермы. В этот период наблюдаются начало митотической активности сперматогониев и усложнение зародышевого эпителия семенных канальцев до сперматид. Первый этап пассивного сперматогенеза приходится на февраль и характеризуется появлением отдельных сперматоцитов 1 порядка (табл. 2), которые, однако, еще не включаются в дальнейшее развитие эпителия, а дегенерируют. На втором этапе (с конца февраля по середину апреля) наблюдается уже последовательное развитие эпителия от однорядного до многорядного, содержащего клетки Сертоли, сперматогонии, сперматоциты I и II порядков, сперматиды (рис. 1б) Вес семенников увеличивается в 10‒15 раз, от 1‒2.5 мг в январе до 30‒60 мг в марте; средний диаметр семенных канальцев близок к максимальному (табл. 2).
Последовательность происходящих изменений зародышевого эпителия такова. В феврале он двухрядный. Первый ряд состоит из выстилающих базальную мембрану клеток Сертоли и сперматогониев, второй ‒ из сперматоцитов I порядка, в массе появившихся во всех канальцах. Средний диаметр канальцев семенника ‒ 31 мкм. В середине марта зародышевый эпителий семенных канальцев многорядный. Первый ряд содержит клетки Сертоли и сперматогонии, второй ‒ сперматоциты I порядка, третий ‒ сперматоциты II порядка и сперматиды. Средний диаметр канальцев 96 мкм.
Активный сперматогенез включает в себя две стадии: преобразование сперматид в зрелые сперматозоиды и интенсивное продуцирование и формирование сперматозоидов (рис. 1в). Семенники достигают максимального размера и веса, увеличиваясь в несколько раз, от 30‒60 мг в марте до 150‒300 мг в апреле‒июне. Весь период активного сперматогенеза длится с конца апреля и до конца жизни животного, т.е. до октября, когда были добыты последние взрослые зверьки. В течение всего этого времени в семенных канальцах присутствуют все клеточные элементы от клеток Сертоли и сперматогониев до зрелых сперматозоидов, собирающихся в просветах канальцев, причем никаких признаков затухания сперматогенеза не наблюдается. Средний диаметр канальцев семенника в этот период составляет от 157 до 189 мкм. Вес семенников ‒ от 154 до 308 мг.
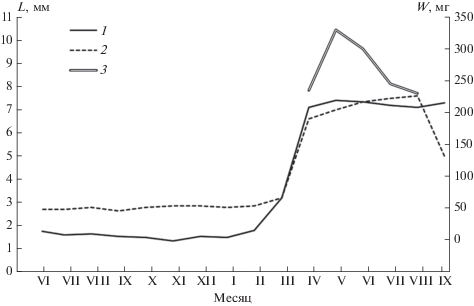
Таким образом, гистологическая картина сперматогенеза обыкновенной бурозубки показывает его стабильную активность в период с конца апреля до октября, отсутствие подъемов и спадов, соответствующих периодам спаривания и паузам между ними. Это подтверждают и данные измерения и взвешивания гонад (рис. 2).
Рис. 2.
Изменение репродуктивных органов у самцов обыкновенной бурозубки: 1 – длина семенника, 2 – вес двух семенников, 3 – высота семенных пузырьков, L – длина, W – вес. По оси абсцисс – месяцы; по оси ординат А – вес двух семенников, мг; по оси ординат Б – длина семенников и высота семенных пузырьков, мм.
Близкая по динамике картина изменений генеративных органов наблюдается у самок. Но если по времени эструсы самок соответствуют активному сперматогенезу самцов, то в начале периода размножения половое созревание самок несколько отстает от такового у самцов.
Судя по гистологическому состоянию яичников, период полового покоя у самок длится с рождения по февраль. Внешне он выражается минимальными размерами рогов и тела матки (табл. 1, рис. 3), составляющими в среднем для этого периода соответственно 7.3 (2.8‒22) и 5.4 (2.4‒12.5) мм. Минимален и средний вес яичников: 1‒3 мг. Матка инфантильна, кровеносные сосуды и маточные железы развиты слабо. Яичник содержит примордиальные фолликулы, некрупные (максимальный диаметр 38 мкм) полостные и атрезирующие фолликулы (рис. 4а).
Рис. 3.
Изменение репродуктивных органов у самок обыкновенной бурозубки: 1 – суммарная длина рогов матки, 2 – длина влагалища на протяжении жизненного цикла.
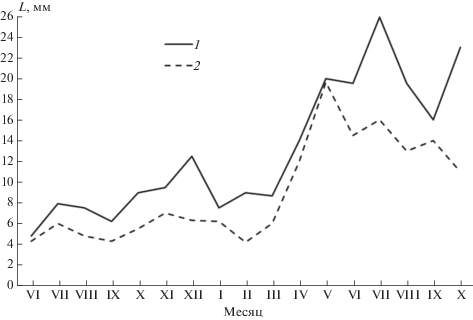
Рис. 4.
Гистологическое строение яичников и слизистой влагалища обыкновенной бурозубки: а – яичник молодой неполовозрелой самки в период полового покоя; б – яичник взрослой половозрелой самки в период интенсивной фолликулярной деятельности; в – яичник взрослой беременной самки с двумя желтыми телами; г – яичник молодой беременной самки с желтым телом и полостными фолликулами; д – влагалище взрослой самки во время эструса. Микрофото, ×400.
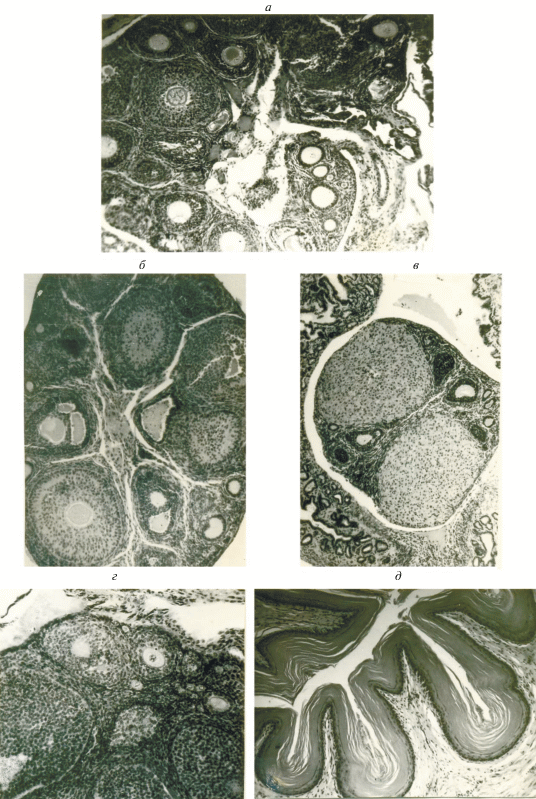
В апреле, т.е. приблизительно через месяц после того как у самцов происходят внешне заметные увеличения размеров гонад, соответствующие изменения половых органов наблюдаются и у самок (рис. 3). Длина рогов достигает в среднем 13.8, тела ‒ 10.8 мм (против 8.3 и 5.0 мм в марте). Увеличивается вес яичников (до 5‒6 мг). В яичниках отмечается много фолликулов на разных стадиях развития, от примордиальных до граафовых пузырьков (рис. 4б). Их диаметр колеблется от 132 до 280 мкм. Ткань яичника хорошо снабжена кровеносными сосудами. Некоторые полостные фолликулы близки к разрыву.
В это же время эпителий слизистой влагалища 8‒10-рядный. В нижних рядах слизистой хорошо заметно ороговение клеток, происходит их усиленная дисквамация; в конце апреля эпителий слизистой 12‒15-рядный, идет активное отслоение ороговевших клеток, которые в большом количестве наблюдаются в просвете влагалища (рис. 4а). Общее состояние характеризуется как эструс.
В дальнейшем, с августа по октябрь можно наблюдать постепенную регрессию гонад, но полной регрессии не происходит вплоть до самой смерти животных в октябре‒ноябре. Здесь также можно видеть некоторый сдвиг во времени по сравнению с самцами, в частности более позднее отмирание самок, сохраняющихся в популяции вплоть до ноября. Самцы, по-видимому, гибнут на месяц, а то и на два, раньше.
В мае встречаются первые беременные самки. Одновременно меняется и гистологическая картина их органов размножения. Яичники содержат желтые тела беременности (их максимальный диаметр 341 мкм), примордиальные и полостные фолликулы на разных стадиях развития (максимальный диаметр 242 мкм) и атрезии (рис. 4в). Ткань яичника хорошо снабжена кровеносными сосудами. Матка гиперемирована, имеет звездчатую форму, в просвете виден секрет маточных желез. Подобную картину можно наблюдать у всех беременных самок в течение сезона размножения.
После родов рост фолликулов замедляется, одновременно увеличивается количество атрезирующих фолликулов. У родивших зверьков наблюдается спад фолликулярной активности. Развитие фолликулов прекращается на стадии образования многослойного зачаткового эпителия. И в сентябре‒октябре рост фолликулов и их атрезия настолько замедлены, что яичники фактически находятся в состоянии покоя.
Таким образом, активный половой процесс продолжается у обыкновенной бурозубки с апреля по октябрь. За это время каждая самка дает 2‒3 помета. Сразу же после первых родов она приходит в эструс снова (послеродовой эструс) и может стать беременной во второй раз, несмотря на все еще продолжающееся молочное кормление первого выводка. После рождения второго помета многие самки не беременеют снова и приходят в лактационный анэструс, другие же до или после прекращения кормления молодых второго помета могут опять прийти в эструс.
Сроки размножения обыкновенной бурозубки, выведенные на основе гистологического исследования гонад, подтверждаются и прямыми наблюдениями. Эти данные показывают, что размножение вида в условиях нашего региона начинается в последних числах апреля, когда регулярно попадаются самцы с наиболее крупными семенниками и придаточными железами. К этому времени изменения в генитальной системе, свидетельствующие о приближении течки (набухание и утолщение стенок матки и влагалища), становятся заметны и у самок. Первые признаки наступления гона (резкое увеличение активности зверьков, беготня их друг за другом, “цирканье”, драки и т. д.) отмечаются в конце апрели‒начале мая. Добытые в этот период самцы имели сильно увеличенные семенники в состоянии сперматогенеза, а самки были оплодотворены или готовы к спариванию.
Первые беременные самки отлавливаются с десятых чисел мая (табл. 3). Во второй половине мая все исследованные самки были беременны (вес эмбрионов 39‒310 мг, длина 5‒16 мм). Первая родившая (подсосная) самка была поймана 27 мая 1970 г. Однако беременные первым пометом самки попадались и гораздо позже ‒ до конца июня, а в годы с поздней весной (1966, 1971, 1980, 1996, 2002, 2006, 2011) ‒ до середины июля.
Таблица 3.
Сроки размножения обыкновенной бурозубки в Карелии
| Годы массовых отловов | Дата первой находки | Дата последней находки | |||
|---|---|---|---|---|---|
| беременной самки | кормящей самки | молодых | беременной самки | кормящей самки | |
| 1959 | 12.05 | 2.06 | 6.06 | 4.10 | 13.10 |
| 1961 | 14.06 | 21.06 | 1.07 | 9.09 | ? |
| 1962 | 23.05 | 19.06 | ? | 2.10 | 4.10 |
| 1967 | 18.05 | 18.06 | 16.06 | ? | 11.10 |
| 1968 | ? | 13.06 | 16.06 | 14.08 | 1.10 |
| 1970 | 19.05 | 27.05 | ? | 17.08 | 8.10 |
| 1975‒1976 | 15.06 | 4.06 | 13.06 | 23.08 | 21.08 |
| 1979 | 17.06 | 17.06 | 18.06 | 27.07 | 26.07 |
| 1981‒1983 | 11.06 | 16.06 | 16.06 | 22.08 | 26.07 |
| 1986 | 29.05 | 29.05 | 14.06 | 18.09 | 17.09 |
| 1989 | 28.05 | 29.05 | 8.06 | 7.08 | 22.09 |
| 1992‒2005 | 12.06 | 15.06 | 10.09 | ||
| 2008‒2010 | 20.06 | 1.09 | |||
Данные табл. 3 и 4 свидетельствуют о значительном варьировании сроков размножения в зависимости от хода весны, а также от численности и состояния популяции. В благоприятных условиях (1959, 1977, 1986, 1992, 1998‒1989, 2008, 2013 гг.) размножение начинается раньше и происходит более интенсивно, в неблагоприятных (1966, 1976, 1980, 1996, 2002. 2011 гг.) ‒ позднее и менее активно.
Таблица 4.
Интенсивность размножения взрослых самок обыкновенной бурозубки в Карелии
| Месяц | Общее число исследованных самок | Относительное количество самок (% от общего числа) | ||||
|---|---|---|---|---|---|---|
| беременных | беременных и одновременно кормящих | кормящих | со следами недавнего кормления выводка | без следов размножения | ||
| Заповедник “Кивач”, 1959 г. | ||||||
| Май | 5 | 80.0 | ‒ | ‒ | ‒ | 20.0 |
| Июнь | 11 | 72.7 | 9.1 | 18.2 | ‒ | ‒ |
| Июль | 25 | 60.0 | ‒ | 36.0 | 4.0 | ‒ |
| Август | 37 | 45.9 | ‒ | 43.3 | 10.8 | ‒ |
| Сентябрь | 2 | ‒ | ‒ | 50.0 | 50.0 | ‒ |
| Октябрь | 2 | ‒ | ‒ | ‒ | ‒ | 100.0 |
| Ноябрь | 2 | ‒ | ‒ | ‒ | ‒ | 100.0 |
| Карелия 1958‒2014 гг. | ||||||
| Апрель | 4 | ‒ | ‒ | ‒ | ‒ | 100.0 |
| Май | 30 | 53.3 | 6.7 | 16.7 | ‒ | 23.3 |
| Июнь | 121 | 52.1 | 13.2 | 16.5 | 10.7 | 7.5 |
| Июль | 214 | 50.5 | 8.9 | 25.7 | 10.3 | 4.6 |
| Август | 171 | 46.2 | 3.5 | 25.2 | 18.1 | 7.0 |
| Сентябрь | 41 | 31.7 | 2.5 | 19.5 | 31.7 | 14.6 |
| Октябрь | 11 | 9.1 | ‒ | 9.1 | 27.3 | 54.5 |
| Ноябрь | 2 | ‒ | ‒ | ‒ | ‒ | 100.0 |
Наиболее поздние находки беременных самок ‒ 2.10.1962 и 4.10.1959, при средней многолетней 19.09, последние поимки кормящих ‒ 13.10.1959 и 11.10.2006, средняя многолетняя ‒ 7.10. Следовательно, весь период размножения обыкновенной бурозубки длится в условиях Карелии примерно 5.5 месяцев ‒ с последних чисел апреля по середину октября. Однако такая продолжительность репродуктивного периода бывает характерна лишь для особо благоприятных лет (как и неблагоприятные, они перечислены выше). В неблагоприятных условиях беременные и кормящие самки отлавливались в более сжатые сроки. Общий ход и интенсивность размножения обыкновенной бурозубки по месяцам представлены в табл. 4. Эти данные показывают, что размножение землероек в нашем регионе во все годы наблюдений проходило достаточно интенсивно, в нем, как правило, участвовали все или почти все взрослые самки, причем многие из них, едва закончив кормление первого выводка, оказывались уже беременными вторично.
Тем не менее ход и интенсивность размножения землероек в течение бесснежного периода закономерно изменяются. В мае, особенно в первой половине, размножаются еще не все взрослые самки, часть из них, очевидно, только еще готовится к спариванию. В июне интенсивность размножения наибольшая, и в нем участвуют почти все взрослые самки, из которых половина беременны, а остальные кормят молодых или находятся в течке. В июле и августе количество беременных несколько уменьшается, зато возрастает число кормящих или кормивших самок. В сентябре доля беременных и лактирующих самок еще более сокращается, а в октябре сходит на нет.
Общую картину размножения обыкновенной бурозубки можно проследить по динамике отлова беременных самок и интенсивности расселения молодых зверьков в расчете по пятидневкам. Как видно на графике, приведенном на рис. 5, первый пик выхода и расселения молодых приходится обычно на первую половину или середину июня (11‒25.06). В конце месяца наблюдается спад, а с начала‒середины июля ‒ новый, еще более отчетливый пик выхода и расселения молодых. Третья волна расселения сеголеток отмечается в середине августа (после спада в конце июля ‒ начале августа). Эти подъемы и спады соответствуют аналогичным подъемам и спадам в размножении популяции (Ивантер, Ивантер, 1968) и свидетельствуют о том, что в условиях таежного Северо-Запада России взрослые самки обыкновенной бурозубки приносят за лето два, а иногда и три выводка. Это подтверждается не только двух-трехвершинным характером кривых на графике выхода молодых, но и тем фактом, что в течение пяти месяцев (май‒сентябрь) почти все зимовавшие самки или были беременны, или кормили молодых. Если бы каждая взрослая самка давала в год всего один помет, то весь период беременности и выкармливания молодых занял бы у нее не более 50 дней, а на протяжении остальных 100 дней следов размножения или лактации у нее не было бы заметно. В таком случае “яловые” самки составили бы не менее 2/3 от всех добытых взрослых самок. В действительности же на их долю приходится обычно не более 10%. Наконец, существование по крайней мере двух генераций у одной самки доказывается совмещением беременности с лактацией ‒ поимкой беременных самок со следами кормления выводка (лактирующих).
Рис. 5.
Динамика отлова обыкновенных бурозубок по пятидневкам: 1 – молодые зверьки в заповеднике “Кивач” (1959 г.), 2 – молодые зверьки в окрестностях с. Новинка (1968 г.), 3 – молодые зверьки в районе стационара “Каркку” (1969 г.), 4 – беременные самки по всем пунктам; Nj – число молодых зверьков, Nf – число беременных самок.
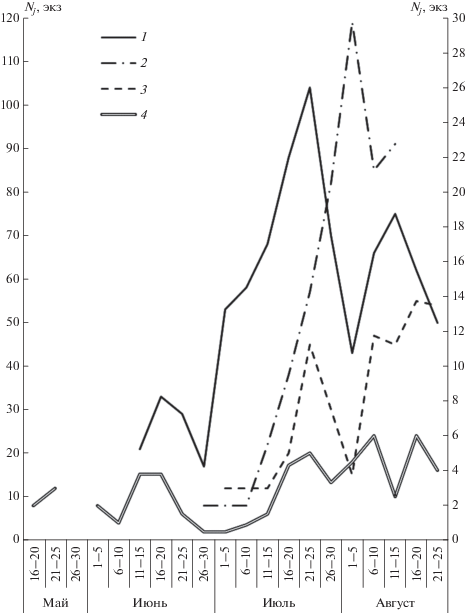
Говоря о числе генерации у обыкновенной бурозубки в течение лета, следует подчеркнуть, что повсюду, в том числе и в Карелии, оно зависит от состояния популяции, т.е. в конечном счете от экологических условий года. В благоприятные годы (1959, 1977, 1985, 1992, 1998‒1999, 2008, 2013) самки приносят два‒три помета, а в неблагоприятные (1966, 1976, 1980, 1996, 2002, 2011) ‒ один‒два. В первом случае все три генерации бывают обычно сильно растянуты и по срокам перекрывают друг друга (отсюда ‒ сближенность волн расселения и неясное их разграничение), во втором ‒ наблюдается более строгая приуроченность появления молодых к определенным срокам (рис. 5). Интенсивность размножения зависит и от плотности популяции. В периоды низкой исходной численности размножение характеризуется большой напряженностью, компенсируя этим малую величину поголовья. Напротив, в годы, следующие за “пиковыми” (например, 1978, 1996, 2002, 2010), размножение происходит вяло и большинство самок приносит только 1‒2 выводка.
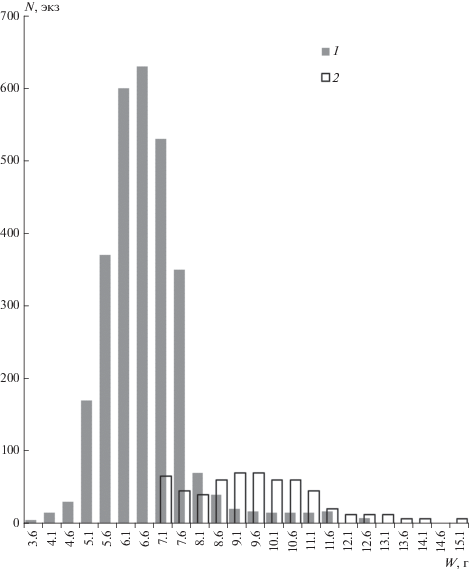
В Карелии и в прилежащих регионах, где проходит северная граница ареала вида, в год своего рождения землеройки обоих полов, как правило, не размножаются. Из 3030 исследованных молодых обыкновенных бурозубок лишь 7 самок (0.2%) имели признаки участия в размножении (рис. 4г). У всех остальных половая система находилась в состоянии полного покоя. При этом, по нашим данным, подавляющее большинство прибылых бурозубок не только не созревает еще в половом отношении, но и не достигает, как правило, веса взрослых половозрелых зверьков, тогда как промеры тела и черепа у обеих возрастных групп близки (рис. 6). Вес тела молодых размножающихся самок колебался от 6.8 до 9.8, составляя в среднем 7.7 г, что гораздо ниже среднего веса взрослых размножающихся бурозубок (10.1 г).
Рис. 6.
Распределение обыкновенных бурозубок по весу тела: 1 –молодые и 2 – взрослые особи, W – вес, верхняя граница классового интервала; N – число зверьков.
Средний размер выводка у обыкновенных бурозубок Карелии на основании подсчета эмбрионов у 235 беременных самок равен 6.7 ± 0.1 с колебаниями от 1 до 11, но чаще всего 5‒8 (табл. 5). В сравнении с соответствующими данными для других популяций вида (табл. 6) такой показатель выглядит невысоким. Пучковский (1970) пишет об увеличении разовой плодовитости обыкновенной бурозубки с продвижением на север и восток. Однако данные, приведенные в табл. 6, опровергают это заключение и свидетельствуя об отсутствии закономерных географических изменений в плодовитости.
Таблица 5.
Вариации величины выводка у зимовавших самок обыкновенной бурозубки по месяцам (сводные данные по всему периоду исследований)
| Месяц | Общее число беременных самок | Количество самок (экз.) с числом эмбрионов | Среднее число эмбрионов на 1 самку | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | |||
| Май | 15 | 1 | 1 | 3 | 3 | 6 | 1 | 7.9 (4.2 + 3.7) | |||||
| Июнь | 51 | 4 | 4 | 9 | 17 | 13 | 3 | 1 | 7.9 (3.8 + 4.1) | ||||
| Июль | 97 | 5 | 5 | 12 | 22 | 32 | 16 | 5 | 6.4 (3.3 + 3.1) | ||||
| Август | 59 | 1 | 1 | 1 | 6 | 9 | 17 | 15 | 7 | 2 | 6.3 (3.3 + 3.0) | ||
| Сентябрь | 11 | 1 | 1 | 1 | 1 | 1 | 4 | 2 | 5.8 (3.1 + 2.7) | ||||
| Октябрь | 2 | 2 | 6.0 (3.0 + 3.0) | ||||||||||
| Всего | 235 | 1 | 2 | 7 | 13 | 26 | 47 | 63 | 45 | 26 | 4 | 1 | 6.7 (3.5 + 3.2) |
Таблица 6.
Средние и экстремальные значения плодовитости обыкновенной бурозубки в различных частях ареала
| Страна и район исследований | n | Число эмбрионов на 1 самку | Источник |
|---|---|---|---|
| Россия | |||
| Карелия | 250 | 6.4 (1‒11) | Наши данные |
| Коми | 109 | 6.4 (1‒11) | Куприянова, 1994 |
| Печеро-Илычский заповедник | 182 | 7.1 (1–13) | Бобрецов и др., 2004 |
| Ленинградская обл. | ‒ | 7.1 (2‒10) | Айрапетьянц, 1970 |
| Московская обл. | ‒ | 6.6 (1‒11) | Дунаева, 1965 |
| Юго-восточная часть Черноземного центра | ‒ | 6.4 (4‒9) | Барабаш-Никифиров, 1957 |
| Волжско-Камский край | 58 | 6.5 (3‒12) | Попов, 1960 |
| Башкирия | ‒ | 7.1 (2‒12) | Снигиревская, 1947 |
| Томская и Новосибирская обл. | 23 | 7.7 (5‒10) | Юдин, 1962 |
| Барабинская лесостепь | 7 | 7.6 (6‒10) | Юдин, 1962 |
| « | ‒ | 7.0 (5‒10) | Юрлов и др., 1965 |
| « | 13 | 8.0 (5‒10) | Строганов, 1957 |
| « | 117 | 6.4 (2‒10) | Глотов и др., 1978 |
| Кулундинская степь | 19 | 9.0 (8‒11) | Юдин, 1962 |
| Горный Алтай | 15 | 5.6 (1‒8) | Юдин, Барсова, 1967 |
| Приобские боры (Алтай) | 15 | 8.0 (6‒10) | Строганов, 1957 |
| Западная Сибирь | ‒ | 8.0 (4‒10) | Строганов, 1957 |
| Южная тайга Средней Сибири | ‒ | 7.2 (1‒12) | Реймерс, 1966 |
| Верхняя Лена | ‒ | 6.7 (1‒11) | Реймерс, Воронов, 1963 |
| Англия | ‒ | 6.5 (1‒10) | Brambell, 1935 |
| Болгария | ‒ | 5.2 (4‒10) | Марков, 1957 |
| Чехословакия | ‒ | 5.7 и 7.0 | Pelikan, 1955 |
| Польша | 104 | 5.2 (1‒10) | Borowski, Dehnel, 1952 |
| Латвия | ‒ | 5.6 (4‒8) | Лапинь, 1963 |
Указанный выше средний показатель плодовитости подвержен значительным колебаниям по годам, месяцам, а также в разных регионах, как в связи с изменениями экологических условий, так и с состоянием и численностью популяции.. Наибольшее число эмбрионов, как это видно из табл. 5, наблюдается в мае‒июне, т.е. в оптимальных экологических условиях. Ближе к осени среднее число эмбрионов на самку закономерно снижается. При этом главной причиной снижения плодовитости самок к осени является, скорее всего, то, что в это время они дают уже не первый, как в мае‒июне, а второй и даже третий помет. Естественно, что они истощены и не могут выносить и выкормить такое же количество детенышей, как весной, когда они находятся в расцвете сил. Плодовитость самок, как и рассмотренные выше показатели интенсивности репродукции, также зависит от плотности популяции (табл. 7). В местах и в годы высокой численности (например, 1959, 1968, 1975, 1981, 1986, 1992‒1993, 1998‒1999, 2001, 2008‒2010, 2013) величина выводка была ниже, а при депрессии (1961, 1966‒1967, 1976, 1978, 1980, 1996, 2002‒2003, 2006, 2011 и 2014 гг.) ‒ достоверно выше.
Таблица 7.
Плодовитость обыкновенной бурозубки в Карелии по годам и регионам
| Место и год отлова | Общее число беременных самок | Количество самок (экз.) с числом эмбрионов | Среднее число эмбрионов на 1 самку | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | |||
| Заповедник “Кивач” (1959 г.) | 31 | ‒ | 1 | 3 | 1 | 6 | 5 | 9 | 6 | ‒ | ‒ | ‒ | 6.0(3.1 + 3.0) |
| Кивач и Пудожский р-н | |||||||||||||
| 1961 г. | 9 | ‒ | ‒ | ‒ | ‒ | ‒ | 1 | 3 | 5 | ‒ | ‒ | ‒ | 7.4(3.1 + 3.6) |
| 1962 г. | 10 | 1 | 1 | 1 | 1 | 1 | 2 | 2 | 1 | ‒ | ‒ | ‒ | 4.9(2.5 + 2.5) |
| Кивач, Каркку | |||||||||||||
| Агробаза (1967 г.) | 7 | ‒ | ‒ | 1 | ‒ | 1 | ‒ | 4 | ‒ | 1 | ‒ | ‒ | 6.4(3.2 + 3.2) |
| Новинка (1968 г.) | 26 | ‒ | ‒ | 1 | 3 | ‒ | 11 | 7 | 2 | 1 | 1 | ‒ | 6.3(3.2 + 3.1) |
| Каркку | |||||||||||||
| 1969‒1979 гг. | 43 | ‒ | ‒ | 1 | 3 | 9 | 8 | 5 | 9 | 7 | ‒ | 1 | 6.7 (3.1 + 3.6) |
| 1980‒1990 гг. | 112 | ‒ | 1 | 2 | 4 | 6 | 23 | 3 | 2 | 1 | 4 | ‒ | 7.0 (3.3 + 3.7) |
| 2 | 2 | 8 | |||||||||||
| 1991‒2000 гг. | 17 | ‒ | ‒ | ‒ | 1 | 3 | 1 | 5 | 5 | 2 | ‒ | ‒ | 6.9 (3.3 + 3.8) |
| 2001‒2007 гг. | 13 | ‒ | ‒ | 1 | 1 | 3 | ‒ | 4 | 3 | 1 | ‒ | ‒ | 6.3 (3.2 + 3.1) |
| 2008‒2014 гг. | 11 | 4 | 2 | 2 | 3 | 5.3 (2.8 + 2.6) | |||||||
Изучение асимметричности распределения количества эмбрионов у самок с вычислением соответствующего показателя “косости” (рис. 7) проводилось нами по методике, заимствованной из нескольких вариационно-статистических изданий (Урбах, 1964; Плохинский, 1970; Зайцев, 1981), а сама идея – из известной статьи Шварца (1966). Полученные этим методом результаты убедительно показали, что в исследуемой популяции обыкновенной бурозубки не только еще не произошла стабилизация средней величины выводка, но даже имеется заметная тенденция к снижению плодовитости (статистический показатель асимметрии, или “косости”, А равен ‒0.54 + 0.22). Этот факт вместе с относительно низкой величиной выводка (0.38 + 0.15) говорит о закономерном процессе снижения плодовитости, намечающемся в популяции. Скорее всего, это связано с тем, что основная масса беременных самок была добыта нами в годы высокой численности популяции и, следовательно, их плодовитость отражает главным образом именно эту ситуацию. А при высокой численности, естественно, действуют регуляторные механизмы, ведущие, в частности, к снижению плодовитости самок. В справедливости такого представления легко убедиться, сравнивая показатели “косости”, вычисленные отдельно для периодов высокой и низкой численности землероек. Как и следовало ожидать, за годы депрессии при высокой средней плодовитости самок показатель “косости” минимальный и с положительным знаком, а при высокой численности ‒ максимальный и с отрицательными знаком (А1 = +0.05 ± 0.52, t1 = 09, χ2 = = 7.6 ± 0.29; А2 = –0.70 ± 0.28, t2 = 2.5, χ2 = 6.0 ± 0.19).
Рис. 7.
Распределение беременных самок обыкновенной бурозубки по числу эмбрионов: 1 – все годы исследований; 2 – годы со средней величиной выводка 4.9–6.4; 3 – годы со средней величиной выводка 7.4–7.8.
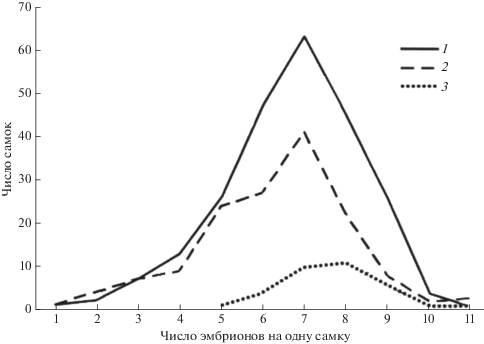
Это говорит о том, что тенденция к снижению плодовитости и вообще механизмы регуляции проявляются в основном только при высокой плотности популяции.
Значительных биотопических различий в плодовитости, о которых пишут Попов (1960) и Штейн (Stein, 1961), наши исследования не выявили (табл. 8). Тем не менее у самок из хвойных и смешанных лесов величина помета оказалась несколько меньше, чем у зверьков из лиственных н окультуренных участков. Это можно связать с повышенной эмбриональной смертностью, наблюдающейся у самок из хвойных и смешанных насаждений (Tarkowski, 1957).
Таблица 8.
Плодовитость самок обыкновенной бурозубки, добытых в разных биотопах
| Биотоп | Общее число беременных самок | Количество самок (экз.) с числом эмбрионов | Среднее число эмбрионов на 1 самку | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | |||
| Сосновые леса | 53 | 1 | ‒ | 2 | 1 | 6 | 13 | 17 | 8 | 5 | ‒ | 6.8 |
| Еловые леса | 68 | ‒ | 2 | 3 | 5 | 8 | 16 | 17 | 11 | 5 | 1 | 6.5 |
| Смешанные леса | 26 | ‒ | 1 | 1 | 2 | 2 | 10 | 4 | 5 | 1 | ‒ | 6.6 |
| Лиственные леса | 66 | ‒ | ‒ | 2 | 2 | 5 | 16 | 20 | 10 | 13 | 4 | 7.5 |
| Участки культурного ландшафта | 24 | ‒ | ‒ | 1 | ‒ | 4 | 2 | 7 | 7 | 3 | ‒ | 7.3 |
В условиях Карелии по нашим данным рассасывающиеся эмбрионы у бурозубок отмечаются редко. Мы обнаружили их лишь у 15 из 122 беременных самок (12.3%), причем общая доля резорбирующихся эмбрионов составила всего 2.1% (из 769 зародышей явно отставало в росте 16).
ЗАКЛЮЧЕНИЕ
Отмеченные выше экологические особенности размножения обыкновенной бурозубки, проявляющиеся в специфических условиях северной периферии ареала, такие как значительная лабильность и разнообразие адаптивных реакций репродуктивной сферы, их трансформация и смена во времени и пространстве, и наконец, выработка широких проявляющихся на популяционном уровне адаптивных комплексов, отличающихся динамичностью и высокой скоростью компенсаторной перестройки, ‒ все это вплотную приближает нас к разработке общей концепции периферических популяций мегаареальных политипических видов животных. Анализ закономерных географических различий, характеризующих популяционную специфику репродукции землероек-бурозубок, обитающих в разных регионах, подтвердило положение о том, что в экологическом центре (оптимуме) видового ареала плотность популяций не только выше, но и устойчивее, тогда как у северных границ распространения она изменяется в широком диапазоне и с большей амплитудой. Соответственно, более выражены и гораздо рельефнее проявляются в условиях северной периферии ареала и специфические структурно-популяционные адаптации, направленные на максимально эффективное воспроизводство видового населения, способное обеспечить видам и их популяциям необходимое преодоление экстремальных условий экологического пессимума.
Список литературы
Айрапетьянц А.Э., 1970. Насекомоядные и грызуны // Звери Ленинградской области. Л.: Изд-во ЛГУ. С. 7‒165
Барабаш-Никифоров И.И., 1957. Звери юго-восточной части черноземного центра. Воронеж, 370 с.
Бобрецов А.В., 2004. Насекомоядные. Мышевидные грызуны // Млекопитающие Печеро-Илычского заповедника. Сыктывкар: Коми книжн. изд-во. С. 38‒109, 206‒301.
Глотов И.Н., Ердаков Л.Н., Кузякин В.А., 1978. Сообщества млекопитающих Барабы. Новосибирск: Наука. 231 с.
Дунаева Т.Н., 1955. К изучению биологии размножения обыкновенной бурозубки (Sorex araneus) // Бюллетень Московского общества испытателей природы. Отд. биол. Т. 60. Вып. 6. С. 27‒43.
Зайцев Г.Н., 1981. Математический анализ биологических данных. М.: Наука. 483 с.
Ивантер Т.В., Ивантер Э.В., Терноушко Е.И., 1974. Биология размножения и структура популяции землероек (Soricidae) Карелии // Вопросы экологии животных. Петрозаводск. С. 95‒143.
Ивантер Э.В., 1975. Популяционная экология мелких млекопитающих таежного Северо-Запада ССС. Л.: Наука. 246 с.
Ивантер Э.В., Ивантер Т.В., 1968. Обыкновенная бурозубка в заповеднике “Кивач” // Труды гос. заповедника “Кивач”. Вып. 1. Петрозаводск. С. 115‒135.
Куприянова И.Ф. 1994. Сем. Soricidae ‒ землеройковые // Фауна европейского Северо-Востока России. Млекопитающие. СПб.: Наука. Т. 2. Ч. 1 С. 11‒67.
Лапинь И.М., 1963. Биология и паразитофауна мелких лесных млекопитающих Латвийской ССР. Рига. 135 с.
Марков Г., 1957. Насекомоядни бозайници в България. София. 387 с.
Плохинский Н.А., 1970. Биометрия. М.: Изд. МГУ. 650 с.
Попов В.А., 1960. Млекопитающие Волжско-Камского края. Казань. 466 с.
Пучковский С.В., 1970. Сравнительно-экологические исследования бурозубок Онежского полуострова. Автореф. дис. … канд. биол. наук. М. 21 с.
Реймерс Н.Ф., 1966. Птицы и млекопитающие южной тайги Средней Сибири. М.– Л.: Наука. 419 с.
Реймерс Н.Ф., Воронов Г.А., 1963. Насекомоядные и грызуны Верхней Лены. Иркутск. 190 с.
Снигиревская Е.М., 1947. Материалы по биологии размножения и колебания численности землероек в Башкирском заповеднике // Труды Башкирского гос. заповедника. Вып. 1. С. 12‒18.
Строганов С.У., 1957. Звери Сибири. Насекомоядные. М.: Изд. АН СССР. 267 с.
Урбах В.Ю., 1964. Биометрические методы. М.: Наука.
Шварц С.С., 1966. Биология землероек лесостепного Зауралья // Зоологический журнал. Т. 34. Вып. 3. С. 915‒927.
Шварц С.С., 1966. Опыт изучения направления изменчивости в природных популяциях животных // ДАН СССР. Т. 166. Вып. 6. С. 60‒69.
Юдин Б.С., 1962. Экология бурозубок (род Sorex) Западной Сибири // Вопросы экологии, зоогеографии и систематики животных. Труды биологического ин-та СО АН СССР. Новосибирск. Вып. 8. С. 33‒134.
Юдин Б.С., Барсова Л.И., 1967. Землеройки кедровых лесов Прителецкого очага клещевого энцефалита // Природа очагов клещевого энцефалита на Алтае. Новосибирск: Наука СО. С. 51‒59.
Юрлов К.Т., Юдин Б.С., Потапкина А.Ф., 1965. К характеристике фауны мелких млекопитающих северной лесостепи Барабинской низменности // Животный мир Барабы. Новосибирск: Наука. С. 184‒207.
Borowski S., 1964. Moult of shrews (Sorex L.) under laboratory conditions. Acta theriol. V. 8. № 8. P. 125‒135.
Borowski S., Dehnel A., 1952. Materialy do biologii Soricidae // Ann. Univ. M. Curie-Sklodowska. № 7. S. 13‒21.
Brambell R., 1935. Reproduction of the lesser shrew (S. araneus L.). 1. The oestrus cycle of the female // Phil. Trans. Soc. B. V. 518. № 225. P. 236‒23.
Pelikan Z., 1955. A contribution to the biology of the common shrew, Sorex araneus L. // Acta theriol. V. 7. № 9. P. 23‒31.
Pucek Z., 1960. Sexual maturation and variability of the reproductive system in young shrews (Sorex L.) in the first calendar year of life // Acta theriol. T. 3. № 12. P. 75‒84.
Stein G.H., 1961. Beziehungcn zwischen Bestandsdichte und Vermehrung bei der Waldspitzmaus Sorex araneus und weiteren Rotzahnspitzmauzen // Z. Saugetierkunde. 26. 1. S. 143‒156.
Spitzenberger F.H., 1966. Uber die makroskopishe Ausbildung der eitendrusen bei der Waldspitzmaus (Sorex araneus L.) // Zool. Anz. T. 177. № 5‒6. S. 87‒98.
Tarkowski A.K., 1957.Badania nad rozrodemi smierteinoscia zarodkowa u ryjowski aksamitnej (Sorex araneus L.). Crese II. Rozrod w warunkach naturalnych // Ann. Univ. M. Curie-Sklodowska. V. 10. № 8. P. 177‒223.
Дополнительные материалы отсутствуют.
Инструменты
Зоологический журнал