

Зоологический журнал, 2020, T. 99, № 3, стр. 326-346
Осенняя авифауна лесных массивов восточного макросклона Тибет-Цинхайского плато (Центральный Китай)
Е. А. Коблик a, *, Ю. Фанг b, Б. И. Шефтель c
a Научно-исследовательский зоологический музей
Московского государственного университета им. М.В. Ломоносова
125009 Москва, Россия
b Институт Зоологии Китайской академии наук
100101 Пекин, Китайская Народная Республика
c Институт проблем экологии и эволюции
им. А.Н. Северцова РАН
119071 Москва, Россия
* E-mail: koblik@zmmu.msu.ru
Поступила в редакцию 24.04.2019
После доработки 28.05.2019
Принята к публикации 22.08.2019
Аннотация
В течение четырех осенних сезонов (сентябрь–октябрь 2011–2012 и 2017–2018 гг.) было проведено орнитологическое обследование восточного макросклона Тибет-Цинхайского плато. Для девяти лесных массивов удалось изучить состав фоновых видов птиц и определить соотношения фаунистических элементов. Сравнение орнитокомплексов позволило выявить их региональную специфику, связанную с ландшафтными и высотными характеристиками, площадью, экологическими параметрами участков. Местную авифауну можно охарактеризовать как палеарктическую с высокой долей эндемиков и субэндемиков. Виды тропического тяготения составили заметную долю лишь на южном макросклоне хребта Циньлин, который очевидно представляет собой северную периферию широкой переходной зоны между Палеарктикой и Ориентальной областью. Выраженный сезонный аспект придают лесным массивам осенние дальние мигранты из северных областей Палеарктики. Особенностью всего региона следует считать широкое проникновение лесных птиц в открытые пространства по интразональным биотопам и проникновение видов открытых пространств на окраины лесных местообитаний в миграционный период. Существует и экологическая группа птиц, связанная преимущественно с экотонами.
В рамках выполнения совместных научных проектов “Сравнительные исследования сообществ позвоночных животных в хвойных лесах Центральной Сибири (Россия) и Тибет-Цинхайского плато (Китай)” и “Структура сообществ позвоночных животных в лесных ландшафтах, расположенных вдоль меридионального трансекта от сибирской тайги до склонов Тибет-Цинхайского плато, и проблемы их охраны”, российско-китайская группа зоологов проводила полевые исследования в Центральном Китае. В состав российской группы исследователей входили сотрудники ИПЭЭ им. А.Н. Северцова РАН, Научно-исследовательского Зоологического музея и кафедры зоологии позвоночных биологического факультета МГУ им. М.В. Ломоносова – специалисты в области териологии, орнитологии, герпетологии и ихтиологии.
В первую очередь нас интересовала фауна позвоночных животных лесных местообитаний восточного макросклона плато и примыкающих к нему с востока хребтов. В зоогеографическом плане эти хребты представляют собой северное продолжение Сино-Тибетских гор в широком понимании (Martens et al., 2011), разграничивающих собственно Тибет и более восточные низкогорные районы Китая, однако иногда их считают восточными отрогами хребта Куньлунь, ограничивающего Тибетское нагорье с севера (Mosbrugger et al., 2018). Ныне большинство лесных массивов представляют собой изолированные “острова” горной тайги среди открытых пространств. Изоляция поддерживается не только за счет вклинивающихся между лесами тундроподобных горных степей, но и за счет антропогенного влияния – вырубки лесов (часто – уже неморального типа) на выположенных нижних участках и практически полной замены исходных лесных биотопов нижних поясов гор на трансформированные сельскохозяйственные и урбанизированные ландшафты. Естественно, фрагментация (как давняя, так и недавняя) лесов привела к определенным трансформациям фауны и населения лесных животных, включая видовой состав, состав доминантов и соотношение видов, наличие фаунистических и экологических замен и лакун, другой специфики. Интерес представляют также историческое и современное проникновение степных фаунистических элементов в соседствующие лесные биотопы, а также обратный процесс – проникновение лесных животных в горную степь. Изучение различных аспектов сообществ позвоночных лесных и мозаичных местообитаний разной степени трансформации стало целью наших работ в регионе.
Одной из главных задач исследований был поиск зоогеографических викариантов и родственных форм, общих для фауны местных хвойных лесов и зональных бореальных лесов севера Палеарктики – в первую очередь сибирской тайги. Существует точка зрения, согласно которой восточные склоны Тибета и Сино-Тибетские горы – это родина таежных экосистем. Именно здесь они возникли, эволюционировали, а потом в несколько видоизмененном виде распространились на территорию Сибири. В частности, на протяжении как минимум последних двух миллионов лет зональная таежная авифауна формировалась путем сменявших друг друга нескольких волн проникновения видов из горно-лесных рефугиумов Южной Палеарктики в меж- и послеледниковые периоды (Назаренко, 1990; Martens et al., 2011; Päckert et al., 2012; 2015). В аридные эпохи лесные “мосты” исчезали, а формы, некогда бывшие единым целым, самостоятельно эволюционировали по обе стороны от великого центральноазиатского пустынно-степного пояса (Назаренко, 1985; Nazarenko, 1990). Более ранние изоляты к настоящему времени приобрели видовой статус и экологическую специфику, а молодые – различаются пока слабо либо неразличимы вовсе.
Мы ставили перед собой также задачу оценки долей палеарктических и ориентальных (индо-малайских) элементов, эндемиков и субэндемиков в разных районах с целью уточнения расположения южной границы Палеарктики. Если в Гималаях эту границу удается четко провести на примере высотного распространения птиц и других групп животных (Коблик и др., 2000; Romanov et al., 2016), то восточнее она более “размыта”, для разных групп животных проводится по-разному (напр. Бобров, 1997, 2005; Norton et al., 2010; Боркин, Литвинчук, 2016).
Некоторые общие и более частные результаты исследований наших комплексных экспедиций в Центральном Китае уже опубликованы (Бобров, 2012; Махров и др., 2013, 2019; Шефтель и др., 2013, 2017; Коблик и др., 2014; Банникова и др., 2017; Бобров и др., 2017; Поплавская и др., 2018). Еще несколько работ, в том числе продолжающих темы этой статьи, готовится к публикации.
РАЙОНЫ И МЕТОДИКИ
В районе исследований хорошо выражена мозаика ландшафтов разного происхождения. Высокогорные холодные степи, типичные для Тибетского нагорья в широком смысле (к нему принадлежит и Тибет-Цинхайское плато), занимают здесь выровненные участки, неглубокие долины рек, а также склоны западной и северной экспозиции. Массивы хвойных лесов таежного типа приурочены к склонам, обращенным на восток и юг, следовательно – увлажненным за счет влияния летнего муссона.
Предварительные сведения по составу авифаунистических комплексов юга провинции Ганьсу и севера провинции Сычуань были собраны нами в 2011 г. (20.09–15.10). Затем целенаправленные орнитологические наблюдения были проведены в том же регионе в 2012 г. (22.09–14.10); 2017 г. (6–25.09) и 2018 г. (5–25.10). В сумме они составили 63 дня за три сезона. Девять пунктов, где исследования лесной авифауны проводили не менее двух дней, показаны на рис. 1, их характеристики даны в разделе результаты. Максимальное расстояние между ближайшими пунктами составляло около 380 км, минимальное – 45 км, средние значения – около 100 км. Самый крупный из обследованных массивов занимал около 11 600 га, большинство были значительно меньше по площади и составляли 2500–1200 га. Для выявления видового состава птиц и оценок их обилия использовали главным образом метод пеших маршрутных учетов (разовых и постоянных) с фиксацией птиц визуально (при помощи 10-кратных биноклей) и по голосу, без фиксированной полосы обнаружения. Общая протяженность маршрутов составила 387 км. Поскольку пересчеты плотности населения по биотопам в осенний период для малознакомой фауны чреваты большими погрешностями, для обычных, регулярно встречающихся видов мы даем здесь лишь экспертные оценки обилия в каждом пункте. Показатели в цифрах для спорадично встречаемых видов приводили, в целях сравнимости с остальными данными, к тем же буквенным символам (табл. 1). В большинстве локалитетов мы проводили также выборочные отловы птиц паутинными сетями – с целью подвидовой (в сложных случаях – и видовой) идентификации, взятия промеров и проб крови для генетического анализа (78 ловушко/суток). Крупных или хорошо заметных птиц учитывали также во время автомобильных маршрутов между пунктами исследований. В те же осенние сезоны мы собрали и сравнительную информацию по фауне птиц других регионов Центрального и Западного Китая – Тибетского автономного района, провинций Цинхай и Шеньси. Специфика исследований, посвященных разным группам позвоночных, а также оптимизация экспедиционной логистики не позволяли нам провести равное количество дней в каждой точке для обеспечения максимальной сравнимости полученных данных. Разные пункты неизбежно оказались обследованы нами с разной степенью полноты. Тем не менее, несмотря на возможный пропуск каких-то спорадично встречающихся видов, для исследованной нами территории мы считаем данные репрезентативными и пригодными для проведения общего сравнения “фоновых” орнитокомплексов лесных “островов”.
Рис. 1.
Основные участки исследований: 1 – резерват Лианхуашань, 2 – окрестности г. Джуни, 3 – резерват Тайзишань, 4 – окрестности монастыря Лангмуси, 5 – окрестности г. Руоэргай, 6 – окрестности пос. Вуджиаоси, 7 – окрестности г. Чуанджуси, 8 – окрестности пос. Сиаобао, 9 – окрестности пос. Шантан.
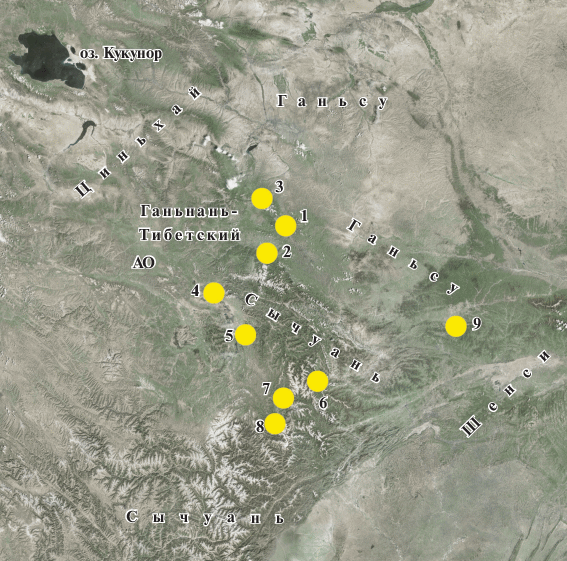
Таблица 1.
Состав орнитокомплексов на участках исследований
| № | Виды и их принадлежность к фаунистическим комплексам | Участки исследований | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | ||
| 1. | Фазан (Phasianus colchicus) – P | о | р | р | – | р | р | р | р | м |
| 2. | Темный кундык (Tetraophasis obscurus) – E | р | – | – | – | – | – | – | – | – |
| 3. | Сермун (Itaginis cruentus) – E | о | о | – | – | – | – | – | – | – |
| 4. | Синий ушастый фазан (Crossoptilon auritus) – E | о | о | – | – | – | – | – | – | – |
| 5. | Рябчик Северцова (Tetrastes sewerzowi) – E | о | – | – | – | р | – | – | – | – |
| 6. | Горный гусь (Anser indicus) – P | – | – | – | р | – | – | – | – | – |
| 7. | Огарь (Tadorna ferruginea) – P | – | – | – | – | р | – | – | – | – |
| 8. | Черная кряква (Anas zonorhyncha) – P | – | – | – | – | р | – | – | – | – |
| 9. | Белокрылая цапля (Ardeola bacchus) – P | – | – | – | – | р | – | – | – | – |
| 10. | Серая цапля (Ardea cinerea) – M | р | – | р | – | – | – | – | – | – |
| 11. | Черношейный журавль (Grus nigricollis) – EP | – | – | – | – | р | – | – | – | – |
| 12. | Серый чибис (Vanellus cinereus) – М | – | – | – | – | – | н | – | – | – |
| 13. | Перевозчик (Actitis hypoleucos) – М | – | – | р | – | – | – | – | – | р |
| 14. | Черныш (Tringa ochropus) – М | – | – | – | – | р | – | – | – | – |
| 15. | Лесной дупель (Gallinago megala) – М | – | – | – | – | – | – | – | – | н |
| 16. | Вальдшнеп (Scolopax rusticola) – М | р | – | – | – | – | – | – | – | – |
| 17. | Речная крачка (Sterna hirundo) – P | – | – | н | – | р | – | – | – | – |
| 18. | Сизый голубь (Columba livia) – CP | – | – | о | – | р | р | – | – | – |
| 19. | Большая горлица (Streptopelia orientalis) – CP | – | – | н | – | – | – | – | – | н |
| 20. | Китайская горлица (Spilopelia chinensis) – C | – | – | о | – | – | – | – | – | р |
| 21. | Индийская кукушка (Cuculus micropterus) – C | – | – | – | – | – | – | – | – | р |
| 22. | Бородач (Gypaetus barbatus) – P | р | – | – | р | р | – | – | – | – |
| 23. | Кумай (Gyps himalayensis) – P | р | – | – | о | н | – | – | н | – |
| 24. | Черный гриф (Aegypius monachus) – P | – | – | – | р | р | – | – | – | – |
| 25. | Большой подорлик (Aquila clanga) – М | – | – | – | р | – | – | – | – | – |
| 26. | Беркут (Aquila chrysaetos) – P | – | – | – | – | р | – | – | – | – |
| 27. | Полевой лунь (Circus cyaneus) – М | р | – | – | р | р | – | – | – | – |
| 28. | Ястреб-тетеревятник (Accipiter gentilis) – P | р | – | – | – | – | – | – | – | – |
| 29. | Ястреб-перепелятник (Accipiter nisus) – P | н | н | р | р | р | р | р | р | р |
| 30. | Черноухий коршун (Milvus (migrans) lineatus) – P | р | – | о | р | о | – | – | – | – |
| 31. | Мохноногий курганник (Buteo hemilasius) – P | – | – | – | – | р | – | – | – | – |
| 32. | Восточный канюк (Buteo (buteo) japonicus) – М | о | – | р | р | р | – | – | н | – |
| 33. | Иглоногая сова (Ninox japonica) – М | – | – | – | – | – | – | – | – | р |
| 34. | Домовый сыч (Athene noctua) – P | – | – | – | – | р | – | – | – | – |
| 35. | Удод (Upupa epops) – C | р | – | – | – | р | – | – | – | – |
| 36. | Крапчатый дятелок (Picumnus innominatus) – O | – | – | – | – | – | – | – | – | р |
| 37. | Желна (Dryocopus martius) – P | р | – | – | – | – | – | р | – | – |
| 38. | Китайский пестрый дятел (Dendrocopos (major) cabanisi) – P | р | – | р | – | – | – | р | – | р |
| 39. | Черношапочный дятел (Picus (canus) guerini) – CO | – | – | – | р | р | – | – | – | – |
| 40. | Чеглок (Falco subbuteo) – P | р | – | – | – | – | – | – | – | р |
| 41. | Сапсан (Falco peregrinus) – М | р | – | – | – | – | – | – | – | – |
| 42. | Сорокопутовый рыжекрыл (Pteruthius aeralatus) – O | – | – | – | – | – | – | – | – | р |
| 43. | Длиннохвостый личинкоед (Pericrocotus ethologus) – EO | р | – | – | – | – | – | – | – | н |
| 44. | Буроспинный личинкоед (Pericrocotus cantonensis) – CP | – | – | н | – | – | – | – | – | – |
| 45. | Дальневосточная райская мухоловка (Terpsiphone (paradisi) incei) – P | р | – | – | – | – | – | – | – | – |
| 46. | Лирохвостый дронго (Dicrurus hottentotus) – CO | – | – | – | – | – | – | – | – | р |
| 47. | Сойка (Garrulus glandarius) – P | р | – | – | – | – | – | – | – | – |
| 48. | Красноклювая сорока (Urocissa erythrorhyncha) – CP | – | – | – | – | – | р | – | – | о |
| 49. | Голубая сорока (Cyanopica cyanus) – P | – | р | о | о | н | – | – | – | – |
| 50. | Сорока (Pica pica) – P | – | р | н | о | о | – | о | – | о |
| 51. | Южная кедровка (Nucifraga (caryocatactes) hemispila) – E | м | о | – | – | м | о | – | – | о |
| 52. | Клушица (Pyrrhocorax pyrrhocorax) – P | р | – | – | м | о | о | о | о | – |
| 53. | Даурская галка (Corvus dauuricus) – P | – | – | – | – | н | – | – | – | – |
| 54. | Черная ворона (Corvus (corone) orientalis) – P | – | – | – | о | – | – | – | н | – |
| 55. | Большеклювая ворона (Corvus macrorhynchus) – C | н | р | н | о | н | р | н | о | н |
| 56. | Ворон (Corvus corax) – P | р | – | – | р | н | – | – | – | – |
| 57. | Восточный малый жаворонок (Calandrella (brachydactyla) dukhunensis) – P | – | – | – | – | р | – | – | – | – |
| 58. | Полевой жаворонок (Alauda arvensis) – М | – | – | – | н | м | – | – | – | – |
| 59. | Рыжепоясничная ласточка (Cecropis daurica) – C | – | – | – | – | – | – | – | н | |
| 60. | Скальная ласточка (Ptyonoprogne rupestris) – P | р | – | – | – | – | – | – | – | р |
| 61. | Восточный воронок (Delichon dasypus) – P | – | – | – | – | – | м | – | – | – |
| 62. | Ошейниковый бюльбюль (Spizixos semitorques) – EO | – | – | – | – | – | р | – | – | о |
| 63. | Зеленокрылый бюльбюль (Ixos mcclellandii) – EO | – | – | – | – | – | – | – | – | н |
| 64. | Рододендроновая широкохвостка (Cettia major) – E | р | – | – | – | – | – | – | – | р |
| 65. | Желтоватая широкохвостка (Horornis acanthizoides) – E | р | – | – | – | – | – | – | – | – |
| 66. | Оливковая широкохвостка (Horornis flavolivaceus) – E | р | – | – | – | – | – | – | – | – |
| 67. | Бурая пеночка (Phylloscopus fuscatus) – P | о | – | – | р | – | – | – | – | – |
| 68. | Пеночка Давида (Phylloscopus armandi) – EP | о | – | м | – | р | – | – | – | – |
| 69. | Пеночка Тикелла (Phylloscopus affinis) – E | – | – | о | р | р | – | – | – | – |
| 70. | Толстоклювая пеночка (Phylloscopus schwarzi) – М | о | – | о | – | – | – | – | – | – |
| 71. | Восточная зеленая пеночка (Phylloscopus (trochiloides) plumbeitarsus) – М | р | – | – | – | – | – | – | – | р |
| 72. | Пеночка-таловка (Phylloscopus borealis) – М | р | – | – | – | – | – | – | – | – |
| 73. | Гималайская пеночка (Phylloscopus reguloides) – E | р | – | р | – | – | – | – | – | р |
| 74. | Светлоголовая пеночка (Phylloscopus coronatus) – Р | – | – | – | – | – | р | – | – | – |
| 75. | Желтогрудая пеночка (Phylloscopus ricketti) – EO | – | – | – | – | – | – | – | – | р |
| 76. | Золотополосая пеночка (Phylloscopus pulcher) – EO | р | – | – | – | – | – | – | – | – |
| 77. | Ганьсуйская пеночка (Phylloscopus kansuensis) – E | м | о | о | – | – | – | – | – | – |
| 78. | Сычуаньская пеночка (Phylloscopus forresti) – E | – | – | – | – | р | – | – | р | – |
| 79. | Китайская пеночка (Phylloscopus yunnanensis) – EP | – | – | – | р | н | р | р | р | – |
| 80. | Тусклая зарничка (Phylloscopus humei) – P | р | р | н | н | н | – | н | – | н |
| 81. | Пеночка-зарничка (Phylloscopus inornatus) – М | р | – | – | р | р | р | – | – | – |
| 82. | Очковая пеночка Бианки (Seicercus valentini) – EO | н | – | – | – | – | – | – | – | р |
| 83. | Красноголовая очковая пеночка (Seicercus castaniceps) – O | – | – | – | – | – | – | – | – | р |
| 84. | Малая бесхвостка (Pnoepyga pusilla) – O | – | – | – | – | – | – | – | – | р |
| 85. | Восточная белолицая кустарница (Garrulax cinereiceps) – EO | – | – | – | – | – | – | – | – | о |
| 86. | Белобровая кустарница (Garrulax sannio) – EO | – | – | – | – | – | – | – | – | н |
| 87. | Кустарница Сукачёва (Garrulax sukatschewi) – E | н | н | – | – | – | – | – | – | – |
| 88. | Кустарница Давида (Garrulax davidi) – EP | н | – | – | – | о | – | – | – | – |
| 89. | Большая кустарница (Garrulax maximus) – E | о | о | – | – | – | – | – | н | – |
| 90. | Желтокрылая кустарница (Trochalopteron elliotii) – EP | м | о | р | р | р | – | – | м | – |
| 91. | Пекинский соловей (Leiotrix lutea) – C | – | – | – | – | – | н | – | – | м |
| 92. | Очковая фульветта (Fulvetta ruficapilla) – EO | – | – | – | – | – | – | – | – | о |
| 93. | Пестрогорлая фульветта (Fulvetta striaticollis) – E | – | о | – | – | о | – | н | н | – |
| 94. | Сероголовая фульветта (Fulvetta cinereiceps) – EO | о | о | – | – | – | – | – | – | – |
| 95. | Золотистая фульветта (Lioparus chrysotis) – EO | – | – | – | – | – | – | – | – | р |
| 96. | Красношейная кривоклювая тимелия (Pomatorhinus ruficollis) – O | – | – | – | – | – | – | – | – | р |
| 97. | Усатая кривоклювая тимелия (Erythrogenis gravivox) – EP | – | – | – | – | – | – | – | – | о |
| 98. | Рыжешапочная тимелия (Cyanoderma ruficeps) – O | – | – | – | – | – | – | – | – | р |
| 99. | Диадемовая юхина (Yuhina diademata) – EP | – | – | – | – | – | – | – | – | р |
| 100. | Буробокая белоглазка (Zosterops erythropleurus) – М | – | – | – | – | – | – | – | – | м |
| 101. | Японская белоглазка (Zosterops japonicus) – P | – | – | – | – | – | – | – | – | н |
| 102. | Бурая сутора (Sinosuthora webbiana) – P | – | – | – | – | – | – | – | – | м |
| 103. | Сероголовая комароловка (Culicicapa ceylonensis) – CO | – | – | – | – | – | – | – | – | м |
| 104. | Желтоголовый королек (Regulus regulus) – P | м | р | – | о | н | – | – | – | – |
| 105. | Расписная синичка (Leptopoecile sophiae) – P | н | о | – | м | р | – | – | – | – |
| 106. | Хохлатая расписная синичка (Leptopoecile elegans) – E | о | – | – | н | н | – | – | – | – |
| 107. | Китайский ополовник (Aegithalos glaucogularis) – P | о | – | – | – | – | – | – | – | – |
| 108. | Сероголовый ополовник (Aegithalus fuliginosus) – E | – | – | – | – | – | – | – | – | н |
| 109. | Черногорлая гаичка (Poecile (palustris) hypermelaenus) – E | – | – | – | – | – | – | – | – | р |
| 110. | Джунгарская гаичка (Poecile (montanus) songarus) – P | м | о | о | о | н | р | н | о | – |
| 111. | Белобровая гаичка (Poecile superciliosus) – E | – | – | – | – | о | – | – | – | – |
| 112. | Желтобрюхая синица (Pardaliparus venustulus) – P | – | – | – | – | – | – | – | – | о |
| 113. | Московка (Periparus ater) – P | р | р | – | – | – | – | – | – | – |
| 114. | Краснобрюхая синица (Periparus rubidiventris) – E | м | р | р | р | р | – | н | – | – |
| 115. | Серохохлая синица (Lophophanes dichrous) – E | м | о | – | н | – | – | – | р | – |
| 116. | Зеленоспинная синица (Parus monticolus) – E | н | – | – | – | – | – | – | – | м |
| 117. | Белобрюхая синица (Parus minor) – CP | р | – | – | – | о | – | – | о | о |
| 118. | Косматый поползень (Sitta villosa) – P | н | – | – | – | – | – | – | – | – |
| 119. | Поползень Пржевальского (Sitta przewalskii) – E | р | н | – | р | р | – | р | – | – |
| 120. | Обыкновенный поползень (Sitta europaea) – P | – | – | – | – | – | р | – | – | р |
| 121. | Стенолаз (Tichodroma muraria) – P | р | – | – | – | – | – | – | – | – |
| 122. | Пищуха (Certhia familiaris) – P | о | р | – | – | – | – | – | – | – |
| 123. | Кашмирская пищуха (Certhia (familiaris) hodgsoni) – E | – | – | – | – | р | – | – | – | – |
| 124. | Крапивник (Troglodytes troglodytes) – P | о | – | р | – | р | – | – | р | – |
| 125. | Оляпка (Cinclus cinclus) – P | р | – | – | – | – | – | – | – | – |
| 126. | Бурая оляпка (Cinclus pallasii) – P | – | – | р | – | – | р | – | – | р |
| 127. | Восточная малая мухоловка (Ficedula (parva) albicilla) – М | – | – | – | – | – | – | – | – | р |
| 128. | Трехцветная мухоловка (Ficedula tricolor) – EO | р | – | – | – | – | – | – | – | р |
| 129. | Синяя мухоловка (Cyanoptila cyanomelana/cumatilis) – М | – | – | – | – | – | – | – | – | р |
| 130. | Синехвостка (Tarsiger cyanurus) – М | р | – | – | – | – | – | – | – | – |
| 131. | Гималайская синехвостка (Tarsiger (cyanurus) rufilatus) – E | м | н | о | р | н | – | н | о | – |
| 132. | Соловей–красношейка (Calliope calliope) – P | р | – | – | – | – | – | – | – | – |
| 133. | Белоножка (Enicurus scouleri) – EP | – | – | р | – | – | р | – | – | – |
| 134. | Белобрюхая горихвостка (Hodgsonius phoenicuroides) – EP | – | – | о | – | – | – | – | – | – |
| 135. | Белогорлая горихвостка (Phoenicurus schisticeps) – E | о | о | – | о | о | – | р | н | – |
| 136. | Алашаньская горихвостка (Phoenicurus alaschanicus) – EP | р | – | – | – | – | – | – | – | – |
| 137. | Полевая горихвостка (Phoenicurus hodgsoni) – E | н | – | – | – | о | – | – | р | – |
| 138. | Синелобая горихвостка (Phoenicurus frontalis) – E | о | о | – | н | р | – | – | о | – |
| 139. | Сибирская горихвостка (Phoenicurus auroreus) – P | н | – | – | – | – | р | – | – | о |
| 140. | Краснобрюхая горихвостка (Phoenicurus erythrogastrus) – P | – | – | – | – | р | – | – | – | – |
| 141. | Водяная горихвостка (Chaimarrornis leucocephalus) – EP | р | р | н | – | – | н | – | р | – |
| 142. | Сизая горихвостка (Rhyacornis fuliginosus) – EP | – | – | р | – | р | н | о | р | р |
| 143. | Серый чекан (Saxicola ferreus) – CO | – | – | – | – | – | – | – | – | р |
| 144. | Азиатский черноголовый чекан (Saxicola (torquatus) maurus) – P | н | – | – | – | – | – | – | – | – |
| 145. | Каменка-плясунья (Oenanthe isabellina) – P | – | – | – | – | р | – | – | – | – |
| 146. | Китайский черный дрозд (Turdus (merula) mandarinus) – CO | – | – | – | – | – | – | – | р | р |
| 147. | Каштановый дрозд (Turdus rubrocanus) – E | м | – | о | – | – | – | н | – | – |
| 148. | Краснозобый дрозд (Turdus ruficollis) – М | о | – | – | – | о | – | – | – | – |
| 149. | Чернозобый дрозд (Turdus atrogularis) – М | н | – | – | р | – | – | – | – | – |
| 150. | Китайский дрозд (Otocichla mupinensis) – P | – | – | – | – | – | – | – | – | р |
| 151. | Длиннохвостый земляной дрозд (Zoothera dixoni) – E | р | – | – | – | – | – | – | – | |
| 152. | Пестрый дрозд (Zoothera varia) – М | р | – | – | – | р | – | – | – | – |
| 153. | Серый скворец (Spodiopsar cineraceus) – P | – | – | н | – | – | – | – | – | – |
| 154. | Рыжегрудая завирушка (Prunella strophiata) – E | о | о | – | о | р | – | – | о | – |
| 155. | Зарянковая завирушка (Prunella rubeculoides) – E | – | – | – | р | р | – | – | – | – |
| 156. | Шиферная завирушка (Prunella immaculata) – E | р | – | – | – | – | – | – | – | – |
| 157. | Острохвостая муния (Lonchura striata) – O | – | – | – | – | – | – | – | – | р |
| 158. | Рыжий воробей (Passer cinnamomeus) – P | – | – | – | – | – | н | – | – | – |
| 159. | Полевой воробей (Passer montanus) – C | – | – | о | н | н | н | о | – | м |
| 160. | Снежный воробей Адамса (Montifringilla adamsi) – EP | – | – | – | – | р | – | – | – | – |
| 161. | Розовый конек (Anthus roseatus) – P | н | – | – | – | р | – | р | – | – |
| 162. | Пятнистый конек (Anthus hodgsoni) – P | о | – | – | – | р | – | – | н | – |
| 163. | Горная трясогузка (Motacilla cinerea) – P | р | – | н | р | р | р | р | – | р |
| 164. | Китайская белая трясогузка (Motacilla (alba) leucopsis) – P | – | – | н | – | н | о | н | – | о |
| 165. | Гималайская белая трясогузка (Motacilla (alba) alboides) – EP | – | – | – | – | р | – | р | – | – |
| 166. | Очковая белая трясогузка (Motacillaalbaocularis) – М | – | – | – | – | – | – | р | – | – |
| 167. | Гималайский вьюрок (Leucosticte nemoricola) – P | н | – | – | н | – | – | – | – | – |
| 168. | Горная коноплянка (Linaria flavirostris) – P | о | – | – | р | н | – | – | – | – |
| 169. | Клест-еловик (Loxia curvirostra) – P | м | н | – | о | о | – | м | – | – |
| 170. | Китайская зеленушка (Chloris sinica) – P | р | – | – | – | – | р | – | – | о |
| 171. | Винно-красная чечевица (Carpodacus vinaceus) – E | – | – | р | – | – | – | – | – | – |
| 172. | Чечевица Бланфорда (Carpodacus rubescens) – E | р | – | – | – | – | – | – | – | – |
| 173. | Серебристая чечевица (Carpodacus pulcherrimus/waltoni) – P | – | – | о | р | м | – | – | м | – |
| 174. | Полосатокрылая чечевица (Carpodacus trifasciatus) – E | – | – | – | – | р | – | – | – | – |
| 175. | Китайская чечевица (Carpodacus (thura) dubius) – E | м | о | – | о | о | о | о | о | – |
| 176. | Урагус (Uragus sibiricus) – P | – | – | – | – | – | – | – | р | – |
| 177. | Зарянковый снегирь (Pyrrhula erythacus) – EP | о | н | о | н | н | – | – | н | – |
| 178. | Арчовый дубонос (Mycerobas carnipes) – P | н | р | – | р | – | – | – | о | – |
| 179. | Овсянка Годлевского (Emberiza godlewskii) – P | – | о | – | – | – | – | – | р | – |
| 180. | Желтогорлая овсянка (Cristemberiza elegans) – P | – | – | – | – | – | – | – | – | р |
| 181. | Овсянка-крошка (Ocyris pusillus) – М | р | р | – | р | – | – | – | р | – |
| Число видов М – 25: | 14 | 1 | 4 | 6 | 6 | 2 | 1 | 2 | 7 | |
| Число видов C – 7: | 2 | 1 | 3 | 2 | 3 | 3 | 2 | 1 | 6 | |
| Число видов P, CP –73: | 38 | 13 | 18 | 23 | 36 | 15 | 12 | 13 | 21 | |
| Число видов E, EP, EO – 64: | 38 | 19 | 14 | 14 | 27 | 7 | 10 | 15 | 19 | |
| Число видов O, CO – 12: | – | – | – | 1 | 1 | – | – | 1 | 11 | |
| ВСЕГО ВИДОВ: | 92 | 34 | 39 | 46 | 73 | 27 | 25 | 32 | 64 | |
Примечания. Номера участков исследований совпадают с номерами пунктов на рис. 1: 1 – резерват Лианхуашань, 2 –окрестности г. Джуни, 3 – резерват Тайзишань, 4 – окрестности монастыря Лангмуси, 5 – окрестности г. Руоэргай, 6 – окрестности пос. Вуджиаоси, 7 – окрестности г. Чуанджуси, 8 – окрестности пос. Сиаобао, 9 – окрестности пос. Шантан. Фаунистические комплексы: М (migrant) – дальние мигранты, палеарктические виды, встречающиеся в регионе только на пролете; P (palearctic) – палеарктические виды; O (oriental) – ориентальные (индо-малайские) виды; C (common, cosmopolitan) – общие для обеих областей виды-космополиты; CP – общие для обеих областей виды, более характерные для Палеарктики; CO – общие для обеих областей виды, более характерные для Ориентальной области; E (endemic) – гималайско-сино-тибетские эндемики; EP – гималайско-сино-тибетские субэндемики с палеарктическим тяготением; EO – гималайско-сино-тибетские субэндемики с ориентальным тяготением. Экспертные оценки обилия: р – редкий или единичный; н – немногочисленный или локально обычный; о – повсеместно обычный; м – многочисленный, фоновый; прочерк – вид не встречен.
Крупный лесной массив Лианхуашань (пункт 1) стал для нас местом стационарных орнитологических наблюдений (33 дня в течение четырех сезонов), от его авифауны, изученной от верхних до нижних границ леса, мы “отталкивались” при исследованиях прочих локалитетов. В 1983 г. здесь был создан резерват для сохранения лесостепных и лесных экосистем, включая реликтовый участок темнохвойных горных лесов Восточного Тибета. Наши коллеги из Института зоологии Академии наук КНР (Пекин) организовали близ верхней границы леса биостанцию, ведут многолетние исследования экологии и гнездовой биологии рябчика Северцова (Tetrastes sewerzowi), неясыти Давида (Strix (uralensis) davidi), китайского мохноногого сыча (Aegolius funereus beickianus), некоторых видов фазанов (Itaginis, Crossoptilon) и кустарниц (Garrulax, Trochalopteron). Авифауна резервата изучена китайскими орнитологами и включает более 300 видов птиц, более двух третей которых относят к гнездящимся (Sun et al., 2008). Нашими усилиями, за три сезона список птиц Лианхуашаня пополнился еще тремя видами. Литературных источников, характеризующих орнитокомплексы других пунктов наших наблюдений, найти не удалось.
Нельзя не упомянуть, что на восточном склоне Тибетского плато в южных районах Ганьсу и северных районах Сычуани в конце 19 в. проводил орнитологические исследования известный русский зоолог и этнограф М.М. Березовский. Сначала – будучи участником Первой Китайско-Тибетской (Ганьсуйской) экспедиции Г.Н. Потанина – с конца 1884 г до начала 1886 г., в основном в верхнем течении Хуанхе, к югу от Ланьчжоу. Затем – возглавляя отдельный отряд Второй Китайской экспедиции Потанина – в 1892–1893 гг., на хребте Циньлин и прилегающем участке бассейна Янцзы. По материалам, полученным при изучении привезенных в Россию орнитологических сборов, им, в соавторстве с обработавшим коллекции В.Л. Бианки, была написана сводка “Птицы Ганьсуйского путешествия Г.Н. Потанина. 1884–1887. Материалы по орнитологии Китая, главным образом южной части провинции Гань-су” (1891). К сожалению, в связи с нечетко указанными координатами и разнобоем в старых и современных названиях населенных пунктов, нам было трудно сопоставить локалитеты указанных ими экземпляров птиц с пунктами наших работ. Тем не менее, мы использовали информацию из этой книги по экологии отдельных видов. Экспедиции других российских путешественников – Н.М. Пржевальского (1879–1880; 1883–1885) и П.К. Козлова (1899–1901; 1907–1909) – исследовали Центральный Китай несколько севернее и западнее района наших работ.
Принадлежность видов птиц к тем или иным фаунистическим комплексам мы определяли, взяв в качестве основы орнитогеографическое деление Палеарктики Штегмана (1938) и выверив ареалогические данные по постоянно возобновляемому и уточняемому источнику (del Hoyo, J., Elliott, A., Sargatal, J., Christie, D.A., de Juana, E. (eds). Handbook of the Birds of the World Alive. Lynx Edicions, Barcelona (retrieved from https://www.hbw.com/node/55095). Научная номенклатура также дана по указанному выше источнику, русская орнитонимика – по http:// zmmu.msu.ru/spec/publikacii/neserijnye-izdaniya/ pticy-mira-rekomenduemye-russkie-nazvaniya-vidov с некоторыми изменениями. Фаунистические данные обработаны по стандартным методикам – использованы формула Съеренсена и метод средней связи для построения дендрограммы сходства.
РЕЗУЛЬТАТЫ
Общий состав птиц, зарегистрированных нами за все сезоны работы в регионе, насчитывает 236 видов. За сезон 2012 г. нам удалось отметить 139 видов птиц, за сезон 2017 г. – 148 видов, за сезон 2018 г. – 121 вид. В табл. 1 представлены только виды, встреченные в 9 пунктах исследований, имевших лесной покров. Приведены не только лесные и кустарниковые птицы, но также птицы открытых пространств и околоводные виды, встреченные на водотоках, опушках, полянах и других открытых стациях, примыкающих к лесным массивам. Не приводятся птицы, встреченные нами только в населенных пунктах, в полностью открытых степных биотопах, на водоемах с безлесными берегами и т.д. Таковы несколько видов водоплавающих (утки, поганки, бакланы, лысухи, чайки), голубей, дневных хищников, а также травник (Tringa totanus), рогатый жаворонок (Eremophila alpestris), тибетская ложносойка (Pseudopodoces humilis), каменные и земляные воробьи (Petronia, Onychostruthus, Pyrgilauda) и некоторые другие. Таким образом, общий список видов в таблице составляет 181 (включая две видовые пары, неразличимые в полевых условиях). Из этого числа 25 видов мы относим к категории дальних мигрантов из северной Палеарктики, распределение которых в значительной мере случайно, а состав определяется скорее сезоном, чем географическим положением и экологическими характеристиками лесного массива. При сравнении фаунистических характеристик исследованных пунктов целесообразно исключить дальних мигрантов, оставив для сравнительного фаунистического анализа 156 местных оседлых и кочующих (включая ближних мигрантов) видов.
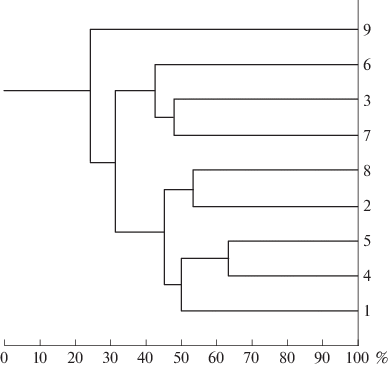
Для выявления видового авифаунистического сходства девяти лесных “островов” восточного макросклона Тибет-Цинхайского плато мы применили формулу Съеренсена: К = 2С/(А + В), где А и В – число видов в сравниваемых районах, а С – число общих видов. Полученные результаты представлены в табл. 2.
Таблица 2.
Сходство фаунистического состава выделенных участков, без учета дальних мигрантов
| Участок | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 |
|---|---|---|---|---|---|---|---|---|---|
| 1 | 78 | 29 | 18 | 29 | 35 | 11 | 17 | 21 | 18 |
| 2 | 0.52 | 33 | 13 | 20 | 21 | 7 | 13 | 17 | 6 |
| 3 | 0.32 | 0.38 | 35 | 15 | 22 | 12 | 14 | 11 | 14 |
| 4 | 0.50 | 0.56 | 0.41 | 39 | 33 | 8 | 15 | 17 | 6 |
| 5 | 0.49 | 0.42 | 0.46 | 0.63 | 66 | 13 | 21 | 22 | 11 |
| 6 | 0.21 | 0.24 | 0.40 | 0.25 | 0.29 | 25 | 11 | 9 | 15 |
| 7 | 0.33 | 0.45 | 0.48 | 0.48 | 0.47 | 0.45 | 24 | 11 | 10 |
| 8 | 0.39 | 0.54 | 0.34 | 0.35 | 0.46 | 0.33 | 0.41 | 30 | 6 |
| 9 | 0.27 | 0.13 | 0.30 | 0.13 | 0.18 | 0.37 | 0.25 | 0.14 | 57 |
Диапазон сходства изученных участков составил 0.13–0.63. В пяти случаях сходство превысило 0.50; в четырех – оказалось меньше 0.20. Построенная по методу средней связи (Жамбю, 1988) дендрограмма наглядно показывает структуру сходства между пунктами, рассчитанного по коэффициенту Съеренсена (рис. 2). Наиболее близкими оказались 4 и 5 пункты. На них наблюдения продолжались сходный период времени и охватывали в основном верхнюю кромку пояса горных хвойных лесов, а кроме того примыкающие участки тундроподобной степи. К обоим близка точка 1, где исследования вели в тех же местообитаниях, но еще и в центре пояса хвойного леса и у его нижней кромки. К этой же кладе относятся 2 и 8 ключевые участки, между собой их объединяет то, что исследования на обоих проводили в долинах рек, хотя степень антропогенной нагрузки в них различалась. Однако то, что реки протекали именно по поясу хвойных лесов, объединяет эти участки с другими участками хвойных лесов (1, 4, 5). Другую кладу составляют локалитеты 7 и 3. Хотя растительность здесь существенно различается, их, возможно, объединяет то, что исследования проводили здесь в короткий период, и в списки попали только наиболее обычные (фоновые) виды. К ним примыкает и участок 6, который, предположительно, можно объединить с участками 3 и 7 по таким показателям, как краткие сроки исследований и наличие неморальных растительных элементов в лесах. И наконец, наименьшее сходство с другими демонстрирует участок 9. Большие фаунистические различия с другими участками, скорее всего, вызваны тем, что он расположен в переходной зоне между Палеарктикой и Ориентальной областями.
Рис. 2.
Рассчитанное по Съеренсену сходство видовых списков птиц, отмеченных на различных ключевых участках. Дендрограмма построена по методу средней связи (Жамбю, 1988).
Краткие характеристики пунктов исследований
1. Резерват Лианхуашáнь (Ганьсу, Ганьнань-Тибетский АО)
34°55′29.3″с.ш., 103°44′43.9″ в.д., высоты 2520– 3250 м над ур. м.
Характерен резкий контраст увлажненных лесистых северо-восточных склонов гор с сухими остепненными юго-западными склонами (рис. 3). В древостое преобладают ели (Picea asperata (доминант), P. purpurea, P. wilsonii, 36%), пихты (Abies faxoniana (доминант), A. fargesii, 23%), ивы (Salix, 23%) и березы (Betula utilis, B. albo-sinensis, B. platyphylla, 14%). Сосны (Pinus armandii, P. tabulaeformes) и дубы (Quercus wutaishanica) редки. Развиты средний ярус и подлесок из различных широколиственных пород (Acer, Ulmus, Juglans и др.), рябин (Sorbus, преимущественно белоягодная S. koehneana), барбарисов (Berberis), бамбука (Sinarundinaria nitida и др.). В нижнем ярусе хорошо развиты ягодные кустарнички, папоротники, мхи, травянистые растения. В субальпийском поясе местами доминируют кустарниковые и древовидные рододендроны (Rhododendron capitatum, Rh. przewalskii, Rh. rufum и др.). Зрелые леса занимают около 28% территории (преимущественно по северным и восточным склонам), остальная площадь занята мелколесьем, кустарниками, лугово-степной растительностью, трансформированными мозаичными ландшафтами (Sun et al., 2008). Характерная особенность – наличие обрывистых склонов с выходами скал под пологом леса или вне его, а также обилие мелких горных ручьев с перекатами, густыми куртинами кустарников и трав по берегам. Это обстоятельство создает исключительную мозаичность биотопов, обилие экотонов и возможность перемещения между ними. По степным склонам – выпас яков и овец, что впрочем, не слишком трансформирует ландшафт.

Попутные орнитологические наблюдения проведены здесь 20–30.09.2011, целенаправленные – 30.09–10.10.2012 (отмечено 69 видов); 16–17 и 19–20.09.2017 (56 видов); 06–08 и 20–23.10.2018 (52 вида). Из 78 суммарно учтенных местных видов (без мигрантов) равную представленность (почти по половине) демонстрируют эндемики/субэндемики и палеарктические элементы, лишь два вида – большеклювая ворона и удод – отнесены к космополитам. 21 вид встречен только в этом локалитете, еще 25 – общие только еще с каким-нибудь одним из исследованных нами локалитетов. Наиболее полно представлены такие лесные группы, как куриные (5 видов), кустарницы (4 вида), синицы (6 видов), пеночки (7 видов, не считая мигрантов), мухоловковые (10 видов, не считая мигрантов). Вместе с тем здесь есть труднообъяснимые лакуны – в наших осенних учетах отсутствуют большая и китайская горлицы, сорока, голубая сорока, бурая сутора, белоглазки, некоторые дрозды и овсянки, встреченные в других пунктах. Наибольшее фаунистическое сходство, как и следовало ожидать, выявлено с соседней точкой Джуни. Выраженные фаунистические связи (около 0.50) прослеживаются также с горно-таежными Лангмуси и Руоэргаем, самые слабые – с наиболее далеким Шантаном (что неудивительно, учитывая и ландшафтные различия) и Вуджиаоси (что, возможно, объясняется меньшей исследованностью последнего).
2. Окрестности г. Джунѝ (Ганьсу, Ганьнань-Тибетский АО)
34°34′39.8″ с.ш., 103°27′56.8″ в.д., высоты 2640–2650 м над ур. м.
Лесной массив, который по физиономическим показателям очень напоминает Лианхуашань, находится по другую сторону горного хребта в распадке реки и не полностью изолирован от Лианхуашаня. Занимает значительно меньшую площадь, вытянут вдоль долины реки. Также характерна разница в экспозиции склонов, имеется инверсия – можжевельниковый пояс на южном склоне находится ниже елово-пихтового пояса, вблизи устья распадка развиты зрелые сосняки Pinus armandii, широколиственный средний ярус менее развит, в лесу много участков, трансформированных в результате выпаса коз и овец.
Наблюдения проводили 19–20.10.2018. Из 33 местных видов к эндемикам и субэндемикам относится 58% (наибольший показатель для всех точек), к палеарктическому комплексу – около 40%, присутствует лишь один вид-космополит – большеклювая ворона. Видов, встреченных только в этой точке, нет. Исключительно с Лианхуашанем Джуни сближает наличие двух видов фазанов, кустарницы Сукачёва, московки, пищухи, сероголовой фульветты. Последний вид симпатричен здесь с пестрогорлой фульветтой, предпочитающей не лесные, а опушечно-кустарниковые стации. Пестрогорлая фульветта была и в других точках, но вместе оба вида встречены только здесь. В Джуни отмечены сорока, голубая сорока и овсянка Годлевского, отсутствующие в Лианхуашане. В целом, облик фауны близок к таковому в Лианхуашане, тем не менее, большее (более 0.52) фаунистическое сходство обнаруживается с Лангмуси и Сиаобао (возможно, это следствие неравномерности обследования). Наименьшее сходство (0.13) ожидаемо наблюдается с Шантаном, невысок коэффициент сходства также с Вуджиаоси.
3. Резерват Тайзишáнь (Ганьсу, Ганьнань-Тибетский АО)
35°14′05.8″ с.ш., 103°25′23.9″ в.д., высоты 2300–2490 м над ур. м.
Несмотря на близость к предыдущим пунктам, Тайзишань выглядит иначе – с более выположенными склонами, преобладанием широколиственного и смешанного леса, занимающего большие площади в долине и на склонах, отдельными пятнами зрелых ельников. Иной тип лесов связан с меньшей абсолютной высотой. Большую площадь занимают также кустарниковые заросли в обширной пойме реки и вдоль системы прудов. Антропогенная трансформация выражается в создании элементов парка, наличии одичавших садов, изменении гидрорежима части водотоков (пруды), выпаса нет.
Исследования авифауны проведены 17–18.09.2017, фрагментарно – в октябре 2011. Из 35 местных видов лишь 40% относится к эндемикам и субэндемикам, тогда как к палеарктам – более 51%. Три вида-космополита составляют заметные 9%. Только в этом локалитете отмечены буроспинный личинкоед, белобрюхая горихвостка, серый скворец и винно-красная чечевица. Диапазон сходства/различия с другими точками невелик (0.30–0.48), больше сходства обнаруживается с Чуанджуси и Руоэргаем, меньше – с Лианхуашанем (несмотря на его близость) и Шантаном (несмотря на общее сходство по “неморальному” облику и наличие общих только для этих точек видов – большой и китайской горлиц).
4. Окрестности монастыря Лангмусѝ (Сычуань)
34°04′44.2″ с.ш., 102°38′02.9″ в.д., высоты 3430–3500 м над ур. м.
Сравнительно неширокий пояс зрелого ельника-зеленомошника на северном крутом склоне горы (рис. 4). Ель представлена одним видом, нет лиственного среднего яруса, практически нет подлеска. Опушка у подножия леса представляет собой мозаику полян с высоким кустарником, разделенную лесными перемычками, есть заболоченные низины. Бедность растительных ассоциаций, очевидно, обусловлена значительными высотами и суровостью климата. На опушках – умеренный выпас лошадей и овец.

Исследования проведены 10–12.10.2018, фрагментарные наблюдения – также в октябре 2011. Из 39 местных видов эндемики и субэндемики составляют 36%, виды палеарктического комплекса – 56%, космополиты – 5%. Как ни странно, присутствует вид, связанный с тропиками, это черношапочный (южный седой) дятел. Нигде, кроме Лангмуси, нами не встречен горный гусь (а также пролетный большой подорлик), общие исключительно с Лианхуашанем – гималайский вьюрок и бурая пеночка, общих исключительно с Руоэргаем – 3 вида, не считая мигрантов. Очень высоко фаунистическое сходство с соседним пунктом Руоэргай (0.63 – наивысший показатель сходства для всех точек), несколько меньшее – с Джуни, самое незначительное (0.13) – с Шантаном.
5. Окрестности г. Руоэргáй, р. Баньюхэ (Сычуань)
33°35′05.1″ с.ш., 103°08′43.3″ в.д., высоты 3400–3520 м над ур. м.
Крутой лесистый северный склон распадка, глубоко врезанного в выположенный горно-степной участок (рис. 5). Вдоль быстрой реки развито лиственное мелколесье с преобладанием нескольких видов ив, местами – участки зрелого мелколиственного пойменного леса. Хвойные массивы состоят из нескольких видов елей и пихт, есть очень незначительная примесь сосен и можжевельников. У рек и ручьев развита мозаика луговин с кустарниками барбариса и шиповника (частично антропогенного происхождения – брошенные огороды). Рядом с ущельем, в горной степи в широкой долине крупной реки Баньюхэ со старицами и рукавами развит невысокий (до 6 м) галерейный лес из ив, тополей и карагача с примесью облепихи, других колючих кустарников, физиономически напоминающий западносибирские колки. В степи, мозаичных биотопах, по опушкам ельника – интенсивный выпас яков и овец.

Орнитологические исследования проводили 14–15.09.2017 (отмечено 39 видов) и 13–15.10.2018 (51 вид). Разница в распределении местных видов (66) по сравнению с Лангмуси заключалась в меньшей степени преобладания палеарктического комплекса (53%) над эндемичным (41%), при сходной доле (4.5%) космополитов. Как и в предыдущей точке, присутствует единственный ориентальный элемент – черношапочный дятел. С Лангмуси эту точку сближает и наличие зарянковой завирушки, черного грифа. Общие виды с Лианхуашанем – рябчик Северцова, кустарница Давида и удод, с Тайзишанем – речная крачка. Таким образом не выглядит удивительной фаунистическая близость Руоэргая с Лангмуси и Лианхуашанем. Самое низкое сходство – с Шантаном. Уникальных для локалитета видов, по данным наших учетов, оказалось неожиданно много – 16.
6. Окрестности пос. Вуджиаосѝ (Сычуань)
32°59′31.5″ с.ш., 104°10′30.8″ в.д., высоты 2240–2330 м над ур. м.
Большие площади ельников и пихтачей по горным склонам, есть широколиственные и мелколиственные породы деревьев в подлеске, в поймах лиственное мелколесье и густые заросли колючих кустарников – облепихи, барбариса, дикой яблони и др. Отмечен умеренный выпас мулов, вырубки и огородничество создают мозаичные ландшафты, преимущественно на выположенных участках у водотоков.
Наблюдения за птицами проводили 12–13.09.2017. Несмотря на небольшие высоты и присутствие неморальных элементов растительности, эта точка характеризуется наиболее резким “креном” в сторону палеарктического комплекса – 60% из 25 местных видов. Эндемиков и субэндемиков – 28% (самый низкий показатель из всех локалитетов), космополитов –12% (самый высокий показатель). Фаунистическое сходство с другими пунктами в целом невысоко, не превышает 0.45 с соседним пунктом Чуанджуси. Самые низкие коэффициенты сходства (0.20–0.25) – не с Шантаном, а с Лианхуашанем, Джуни, Лангмуси. Видимо это объясняется наличием только в Вуджиаоси и Шантане красноклювой сороки, ошейникового бюльбюля, пекинского соловья и обыкновенного поползня. С Тайзишанем Вуджиаоси сближает присутствие белоножки. Только в этом локалитете нами отмечены восточный воронок, светлоголовая пеночка и рыжий воробей. Возможно, специфические черты Вуджиаоси объясняются недостаточной полнотой обследованности этого пункта.
7. Окрестности г. Чуанджусѝ (Сычуань)
32°45′08.1″ с.ш., 103°38′51.3″ в.д., высоты 3320–3360 м над ур. м.
Зрелый пихтово-еловый лес на гребне горы, ниже переходящий в мозаичный ландшафт луговин с лиственными перелесками и зарослями кустарников. Лес физиономически похож на хвойник с предыдущей точки, но практически лишен лиственного подлеска. Выпаса не обнаружено, на выположенных участках есть вырубки.
Орнитологическое обследование проведено 13–14.09.2017. Из 24 местных видов половина представляет палеарктический комплекс, 42% – эндемики и субэндемики, 8% – космополиты. Фаунистическое сходство с другими пунктами невысоко – от 0,48 с Тайзишанем и Лангмуси (одинаковый коэффициент сходства со столь разными точками труднообъясним) до 0.25 с Шантаном. С Руоэргаем Чуанджуси сближает присутствие гималайской белой трясогузки, с Лианхуашанем – встречи желны. Как и в случае с предыдущим пунктом, странности, скорее всего, объясняются недостаточной полнотой обследованности.
8. Окрестности пос. Сиаобáо (Сычуань)
32°29′59.2″ с.ш., 103°35′24.2″ в.д., высоты 2980–3010 м над ур. м.
Довольно пологая террасированная долина со смешанными лесами по обоим склонам (рис. 6). Леса в нижней части склонов преимущественно молодые, почти без широколиственных пород, часто разреженные, выше ель постепенно начинает преобладать. Пихта и сосны не встречены. Днище долины занято лугами с густыми зарослями облепихи и барбариса вдоль русла реки и на низких террасах. Физиономически немного напоминает Тайзишань. Развит умеренный выпас яков и лошадей.

Наблюдения проводили 16–17.10.2018. Половина из 30 встреченных местных видов относится к эндемикам и субэндемикам, больше 43% приходится на палеарктический комплекс, космополиты и ориентальные виды представлены, соответственно, большеклювой вороной и китайским черным дроздом. Только здесь нам встретился урагус. Общий вид с Шантаном – тот же черный дрозд, с Джуни – овсянка Годлевского, с Руоэргаем – сычуаньская пеночка, с Лангмуси – черная ворона. Наибольшее фаунистическое сходство Сиаобао обнаруживает с Джуни, наименьшее – с Шантаном.
9. Окрестности пос. Шантáн (Ганьсу)
33°40′22.2″ с.ш., 106°15′21.6″ в.д., высоты 1280–1500 м над ур. м.
Этот пункт наших исследований, наиболее удален к востоку, находится ниже всех над уровнем моря и заметно отличается от остальных пунктов в ландшафтном и климатическом (жарко и дождливо) отношении. Развиты неморальные хвойно-широколиственные леса с субтропическими элементами на южном макросклоне хребта Циньлин. Несмотря на небольшие абсолютные высоты, горы очень крутые и скалистые, густые высокие леса далеко не везде могут расти, местами переходят в подобие средиземноморского шибляка (рис. 7). С другой стороны на большинстве выположенных участков лес вырублен и замещен мозаикой огородов, выгонов для скота и плодово-ореховых садов. Доминируют разнообразные широколиственные породы, преимущественно дубы и клены, встречаются густые лиственничники, разреженные сосняки Pinus armandii. Нижний ярус загущен, деревья перевиты жимолостью, диким виноградом, другими лианами, в подлеске доминируют бамбук нескольких видов, аралия (Aralia), по опушкам и краям травянистых луговин – шиповник, ивы, дикая яблоня, ежевика.

Исследования проведены 08–10.09.2017. Местная авифауна по результатам наших наблюдений составила 57 видов, уступив только Лианхуашаню (а, учитывая неравномерность обследования, скорее всего, превосходит фауну последнего). Палеарктических элементов 37% – это наименьшая доля среди всех 9 пунктов. Вместе с тем доля эндемиков и субэндемиков тоже почти минимальна – 33% (превосходит только долю в Вуджиаоси). Зато доля видов-космополитов повысилась до 10.5%, а ориентальные элементы впервые представлены несколькими видами, составляющими 19% от фауны. Специфику авифауне этого пункта придают разнообразие тимелий в широком смысле (включая иной видовой состав кустарниц и фульветт), наличие индийской кукушки, дронго, суторы, комароловки, мунии, белоглазок. Половина видов птиц из этого локалитета больше нигде нами не встречена. Еще четверть – встречена еще только в какой-либо одной из других точек. Из последней группы больше всего видов, общих с Лианхуашанем, – чеглок, длиннохвостый личинкоед, скальная ласточка, рододендроновая широкохвостка, очковая пеночка Бианки, зеленоспинная синица, трехцветная мухоловка. Тем не менее, Лианхуашань демонстрирует с Шантаном наименьшее сходство. Впрочем, значения коэффициента фаунистического сходства у Шантана самые низкие из всех локалитетов – 0.14 и менее для Сиаобао, Лангмуси и Джуни, немного выше еще для трех пунктов. Максимальное сходство (0.37) обнаруживается с наиболее близким географически пунктом – Вуджиаоси, что не выглядит слишком странным, учитывая наличие четырех общих только для этих локалитетов видов.
ОБСУЖДЕНИЕ
Зоогеографические особенности авифауны
Виды палеарктического тяготения – бесспорно палеарктические элементы, совсем не проникающие в тропики, А также общие для Палеарктики и Ориентальной области виды, более характерные для Палеарктики и шире здесь распространенные. Суммарно таковых оказалось 73 вида, или 47%. Их доля в разных пунктах варьировала от 37 до 60%, ожидаемо оказываясь наименьшей в локалитетах с преобладанием неморальных лесов и наибольшей в высоко поднятых над уровнем моря горно-таежных островах среди степей. Много палеарктических элементов представлено среди дневных хищников, врановых, ласточек, трясогузковых, вьюрковых, овсянок, к ним относятся обе оляпки, крапивник, королек и др. Как ни удивидельно, ареалогически в эту категорию попадают и некоторые теплолюбивые виды, характерные для неморальных лесов и даже субтропиков – фазан, буроспинный личинкоед, дальневосточная райская мухоловка, красноклювая сорока, светлоголовая пеночка, японская белоглазка, бурая сутора, желтобрюхая синица, китайский дрозд, рыжий воробей. Если же не исключать из анализа дальних мигрантов (см. ниже), доля видов палеарктического тяготения в осенней авифауне региона составит 54%.
На самом деле облик лесной авифауны региона выглядит еще более палеарктическим, однако недавнее разделение ряда широко распространенных видов на локальные алловиды привело к тому, что некоторые “типичные палеаркты” в широком понимании оказались представлены в регионе эндемичными видами в узком понимании, еще недавно считавшимися лишь подвидовыми группами. Это кедровка (Nucifraga caryocatactes), корольковая пеночка (Phylloscopus proregulus), ополовник (Aegithalos caudatus), черноголовая гаичка (Poecile palustris), синехвостка.
Виды тропического тяготения – всего 12 видов, ареалогические и биотопические связи которых с Ориентальной областью несомненны. В среднем они составили 8% местной фауны, но в 5 из 9 локалитетов не были представлены вовсе. Еще в трех локалитетах присутствовало всего по одному виду (2–3%), при этом черношапочный дятел (Лангмуси, Руоэргай) и китайский черный дрозд (Сиаобао) еще недавно представляли собой ориентальные группы подвидов широко распространенных в Палеарктике седого дятла (Picus canus) и черного дрозда (Turdus merula). Только в Шантане виды тропического тяготения были представлены наиболее полно (отсутствовал лишь черношапочный дятел). Крапчатый дятелок, сорокопутовый рыжекрыл, красноголовая очковая пеночка, малая бесхвостка, рыжешапочная и красношейная кривоклювая тимелии, острохвостая муния – виды, лишь северными краями ареалов проникающие в Палеарктику, – придают неморальному Шантану почти тропический колорит. Лирохвостый дронго, серголовая комароловка, китайский черный дрозд, серый чекан заходят в Палеарктику гораздо севернее, достигая Маньчжурии (залеты первых трех видов зарегистрированы и в Приморье).
Один из вариантов проведения зоогеографической границы между Палеарктикой и Ориентальной областью в Китае – по водоразделу между бассейнами Хуанхе и Янцзы (напр. Norton et al., 2010). Однако, несмотря на авифаунистическое своеобразие Шантана, находящегося на южном макросклоне хребта Циньлин, т.е. уже в бассейне Янцзы, а не Хуанхэ, соотношение здесь палеарктических (37%) и ориентальных (19%) элементов далеко от равновесного. Доля палеарктических элементов превосходит здесь и долю эндемиков/субэндемиков. Таким образом, по видовому составу авифауны, Шантан следует относить к Палеарктической области, а граница с Ориентальной областью (если понимать под ней равную представленность фаунистических элементов) в этом районе, видимо, находится южнее, ближе к Янцзы и ниже по абсолютным высотам. В широком же смысле граница должна представлять собой протяженную переходную зону с тропическими, субтропическими и неморальными анклавами и эксклавами, находящимися на разных высотах, в соответствии со сложным рельефом. Район Шантана вполне может входить в северную периферию этой зоны.
Гималайско-сино-тибетские эндемики и субэндемики – виды, не встречающиеся на гнездовании за пределами Гималаев и примыкающих с востока горных массивов Центрального Китая (собственно эндемики), а также виды, проникающие на палеарктическую (Тибет) и ориентальную (хребты юга Китая и севера Индокитая) окраины этого естественного зоогеографического региона, характеризующегося специфической лесной фауной (субэндемики). Эта группа, по нашим учетам, составила 64 вида птиц, или 41% местной авифауны. Их доля варьировала от 28 до 58%, в целом оказываясь выше в горно-таежных точках и ниже в точках с преобладанием неморальных лесов. Действительно большинство эндемиков оказываются связанными именно с хвойными лесами с примесью мелколиственных пород и представляют собой экологические викариаты видов, характерных для зональных таежных и подтаежных лесов севера Палеарктики. Примеры таких южно-северных пар викариантов: рябчик Северцова – рябчик, южная кедровка – сибирская кедровка, черногорлая гаичка – черноголовая гаичка, хохлатая синица (гренадерка) – серохохлая синица, гималайская синехвостка – синехвостка, каштановый дрозд – рябинник, зарянковый снегирь – снегирь. Викарианты могут различаться по степени экологического сходства и таксономического родства, а также несколько варьируют и параметры занимаемых ими ниш. Например, в Лианхуашане серохохлые синицы были явно привязаны к китайской желтой березе (Betula albo-sinensis), под большими лоскутами отставшей коры которой птицы обычно искали насекомых. Вместе с тем в других точках такой привязанности этот вид не демонстрировал, равно как и его северо-западный викариант – европейская гренадерка. Южная кедровка не встречена нами в локалитетах, где отсутствовали сосны Армана, трофические связи северного члена пары выглядят более разнообразными. В то же время экологическую нищу, занятую в смешанном лесу северной Евразии лишь одним видом (например, ополовником), в Центральном Китае порой эксплуатируют 3–4 родственных и неродственных ему вида. Более подробному разбору темы викариатов и сравнению экологической структуры бореальных авифаун Сибири и Восточного Тибета мы собираемся посвятить специальную публикацию.
Меньшее число эндемиков региона связано не с хвойными, а с лиственными лесами. Таковы длиннохвостый личинкоед, бюльбюли, кустарницы, часть фульветт и тимелий, диадемовая юхина, некоторые синицы и др. Наибольшее количество эндемиков выявлено в таких группах как куриные (4 вида), широкохвостки (3 вида), пеночки (9 видов), тимелии в широком смысле, включая кустарниц и фульветт (11 видов), синицы, ополовники, поползни и пищухи (суммарно 9 видов этой гильдии), горихвостки (7 видов), завирушки (3 вида), вьюрковые (5 видов).
Именно среди гималайско-сино-тибетских эндемиков и субэндемиков наблюдается высокий процент криптических видов, выявляемых в основном в результате отлова сетями и обследования экземпляров “в руках”. Таковы, например, ганьсуйская, сычуаньская и китайская пеночки, входящие в комплекс корольковой пеночки в широком понимании. По крайней мере, для сычуаньской и китайской пеночек нами установлено совместное обитание в Руоэргае и Сиаобао, а хорошо выявляемая по песне ганьсуйская пеночка отмечена нами лишь в трех северных точках – Линьхуашане, Джуни и Тайзишане. Широко распространенную в Палеарктике пищуху, отмеченную нами в Лианхуашане и Джуни, южнее, в Руоэргае сменяет эндемик – кашмирская пищуха, скорее всего в регионе имеет место парапатрия. По крайней мере, для Сиаобао мы предполагаем совместное обитание криптических видов Carpodacus pulcherrimus и С.waltoni, до последнего времени объединяемых под названием серебристая чечевица, но ввиду неопределенности ситуации (“серебристая чечевица” встречена в четырех точках) в таблице эти виды объединены. Первый из этих видов может считаться палеарктическим, второй – эндемичным, их ареалы демонстрируют симпатрию в районе наших исследований. Наконец, два местных подвида гаички, которые в большинстве современных сводок считают принадлежащими к пухляку (Poecile montanus sensu lato), а в отечественной литературе – к джунгарской гаичке, могут оказаться самостоятельным эндемичным алловидом, лишь конвергентно сходным с настоящей джунгарской гаичкой.
Указанные случаи позволяют предположить, что процесс видообразования в регионе весьма интенсивен и следует обращать особое внимание на узкоареальные формы, возможно представляющие собой молодые виды (алловиды, полувиды). Выявление и изучение таких форм актуально, в первую очередь, в связи с тем, что они находятся в угрожаемом состоянии в результате фрагментации исходных биотопов и в связи с необходимостью принять незамедлительные меры для их охраны.
Виды-космополиты (как минимум в масштабах Азии) – виды, современные ареалы и оптимумы распространения которых не демонстрируют отчетливого тяготения к Палеарктической или Ориентальной областям. К космополитам мы относим китайскую горлицу, индийскую кукушку, удода, большеклювую ворону, рыжепоясничную ласточку, пекинского соловья и полевого воробья. Вероятную географическую родину каждого конкретного вида (или рода) мы в дальнем контексте не рассматриваем, представляя, например, что удод происходит из Африки, а пекинский соловей – из субтропиков Восточной Азии. Эти 7 видов составили в среднем примерно 4.5% местной авифауны, при этом их доля в отдельных локалитетах варьировала от 3 (Джуни) до 12% (Вуджиаоси). Тем не менее, по факту их присутствие в большинстве пунктов ограничивается 1–3 видами и лишь в Шантане доходит до 6 (здесь не встречен только удод). Из них по-настоящему повсеместно встречаются в регионе лишь связанные с антропогенными ландшафтами большеклювая ворона (все пункты) и полевой воробей (6 пунктов), остальные встречены лишь в 1–2 пунктах, т.е. имеют локальное распространение.
Экологические особенности авифауны
Сезонный аспект наблюдений и дальние мигранты. При исследовании осенней авифауны Центрального Китая необходимо учитывать сезонный аспект. Временной диапазон осуществляемых в разные годы исследований составил 55 дней – с 6 сентября по 25 октября. В достаточно полной мере сезонные изменения авифауны за эти полтора месяца были прослежены лишь для Лианхуашаня, остальные пункты посещались нами в разные и более краткие отрезки времени. Так, учеты, проведенные в ряде пунктов в начале–середине сентября, включают ласточек, некоторых пеночек и мухоловок, которые гнездятся в регионе, присутствуют здесь ранней осенью, но к октябрю улетают. Отсюда следует неизбежный недоучет этих местных видов для пунктов, где исследования проведены в октябре. В сентябре мы уже учитываем и ранних мигрантов с севера – большинство трясогузок, мухоловок, куликов, некоторых пеночек, буробокую белоглазку, иглоногую сову и др. С октября в исследованных точках выявляется вторая волна северных мигрантов – дневные хищники, темнозобый, краснозобый и пестрый дрозды, овсянка-крошка, полевой жаворонок, серая цапля и др. В большинстве исследованных точек осенние мигранты образуют весьма заметную долю наблюдаемой авифауны, образуют крупные стаи, активно потребляют пищевые ресурсы, конкурируя с местными видами.
Проблема разграничения местной фауны и мигрантов не имеет удовлетворительного решения для видов, представленных как местными, так и северными популяциями. Таковы некоторые водоплавающие и околоводные виды, дневные хищники, ласточки, пеночки, овсянки, соловей-красношейка, сибирская горихвостка, азиатский черноголовый чекан, каменка-плясунья, пятнистый конек, горная трясогузка, гималайский вьюрок, горная коноплянка, китайская зеленушка, желтогорлая овсянка и др. В табл. 1 эти виды были включены в состав местной фауны.
К дальним сезонным мигрантам из северных районов Палеарктики мы безоговорочно отнесли 25 видов (почти 14% от всей фауны), их доля в разных точках составила от 3–4 (овсянка-крошка, Джуни; очковая белая трясогузка, Чуанджуси) до 15% (14 видов, Лианхуашань). Их поведение характерно для мигрантов – они обычно пролетают или кормятся стаями (реже поодиночке), держатся в одном месте непродолжительное время и, как правило, не попадают в повторные учеты. Коэффициенты сходства местных орнитокомплексов мы рассчитывали без учета этой группы.
Локально и широко распространенные в регионе виды. Встреченных более чем в 2/3 обследованных пунктов, т.е. широко распространенных в регионе птиц оказалось 8 видов (более 4% от общего состава фауны и почти 5% от местной фауны). Помимо упомянутого выше космополита – большеклювой вороны – к ним относятся 5 палеарктических элементов – ястреб-перепелятник (все пункты), фазан, джунгарская гаичка (по 8 пунктов), тусклая зарничка, горная трясогузка (по 7 пунктов) – и 2 эндемика – гималайская синехвостка, китайская чечевица (по 7 пунктов).
81 вид (45% от всех встреченных, с разбросом по пунктам от 0 до 50%) отмечен нами только для какого-либо одного из локалитетов. Таковых оказалось довольно много в Шантане, Лианхуашане и Руоэргае, значительно меньше – в прочих пунктах, а в Джуни они не встречены. Почти в два раза меньше оказалось видов, характерных только для двух из девяти точек, – 43 (24% от общей фауны, с разбросом от 8 до 28%). И здесь большие абсолютные цифры (от 25 до 12 видов) характерны для тех же трех пунктов, для остальных же – от 2 до 8. Процентные же соотношения не всегда совпадают с абсолютными значениями, отражая, в том числе, разницу в общем богатстве фауны. Виды, встреченные нами в 3–6 пунктах (49 видов, 27% от общей фауны), заполняют континуум между локально и повсеместно распространенными видами.
Птицы экотонов. Во всех обследованных точках повышенные плотности населения и видовое разнообразие птиц наблюдались в мозаичных стациях и переходных зонах между горно-тундрово-степными и лесными стациями. Особенно показательной была ситуация в Руоэргае. Здесь в долине р. Баньюхе по соседству встречались такие виды открытых пространств, как мохноногий курганник, черношейный журавль, домовый сыч, удод, восточный малый и полевой жаворонки, каменка-плясунья и характерные для бореальных и неморальных лесов голубая сорока, кустарница Давида, желтоголовый королек, хохлатая расписная синичка, белобровая гаичка, белобрюхая синица. Здесь же отмечены виды околоводных биотопов – огарь, черная кряква, белокрылая цапля, черныш, речная крачка, горная трясогузка, сизая горихвостка – и виды, считающиеся ореофилами, – бородач, кумай, черный гриф, беркут, клушица, краснобрюхая горихвостка, розовый конек, снежный воробей Адамса, горная коноплянка. Выход многих лесных (и даже горно-таежных!) видов далеко в открытые пространства по пойменным ивнякам и облепишникам или поливидовым зарослям кустарников отмечен нами практически по всему региону.
Можно выделить несколько видов и целых систематических групп птиц, характерных в регионе преимущественно для колючих кустарниковых зарослей и субальпийского криволесья, встречающегося не только на опушках изолированных лесных массивов (рис. 4, 5), но в еще большей мере – вдоль горных гребней, на границе лесных и остепненных склонов разных экспозиций (рис. 3). В первую очередь, это разнообразные горихвостки (5 видов), чечевицы (5 видов), завирушки (3 вида), а также темный кундык, расписная синичка, рододендроновая широкохвостка, бурая пеночка, тусклая зарничка, китайская зеленушка. Интересно, что расписная синичка, встречающаяся на Тянь-Шане почти исключительно в горных арчевниках, в изученных нами локалитетах придерживалась зарослей колючих лиственных кустарников и никак не была связана с арчой, даже в тех немногих точках, где арча присутствовала.
ЗАКЛЮЧЕНИЕ
Орнитологические исследования, предпринятые в ходе четырех комплексных экспедиций, позволили изучить в общих чертах осенние (сентябрь–октябрь) аспекты авифауны лесных массивов восточного макросклона Тибет-Цинхайского плато, выявить соотношения между разными зоогеографическими группами птиц в девяти изученных нами локалитетах региона и связать авифаунистические параметры с ландшафтными характеристиками изученных точек. Несмотря на значительные фаунистические различия между обследованными пунктами, в регионе в целом преобладают виды, связанные с Палеарктикой, сопоставимую по значению долю составляют эндемики и субэндемики Гималаев и Сино-Тибетских (в широком понимании) гор, а виды ориентального комплекса и космополиты весьма немногочисленны. Таким образом, авифауну региона следует характеризовать как несомненно палеарктическую с обилием специфических эндемичных элементов и незначительной примесью видов тропического тяготения. По данным нашего орнитологического обследования, зоогеграфическая граница между Палеарктикой и Ориентальной областью находится за пределами изученного региона, однако орнитокомплекс южного склона хребта Циньлин можно квалифицировать как маркирующий периферию широкой переходной зоны между этими областями. Заметный сезонный аспект в лесных массивах региона составляют осенние дальние мигранты из северных областей Палеарктики.
Тема викариата северных (палеарктических) и южных (преимущественно эндемичных) видов птиц, связанных с лесными биотопами, будет подробно рассмотрена нами в другой статье, здесь же мы высказали лишь самые общие соображения. Особенностью региона следует считать широкое проникновение лесных видов птиц в открытые пространства по интразональным биотопам и (в меньшей степени) проникновение видов открытых пространств в экотоны между лесом и степью. Существуют и виды, а также целые систематические группы птиц, характерные именно для экотонов региона.
Дальнейшего специального исследования требует фауногенетический аспект. Характерное для региона богатство и разнообразие видов пеночек (Phylloscopus, Seicercus), горихвосток (Phoenicurus, Chaimarrornis, Rhyacornis) вьюрковых (Carpodacus, Uragus, Pyrrhula, Mycerobas, Chloris), синиц (Parus, Periparus, Lophophanes, Poecile) контрастирует с обедненным составом представителей этих же групп в Сибири и на Дальнем Востоке (Price, 2005; Коблик и др., 2017). Это соотношение подтверждает мнение, что исторически превалировал вектор расселения многих таксонов птиц из этого горно-лесного региона на север – в области зональной тайги Евразии (Päckert et al., 2012). При этом обилие гималайско-сино-тибетских эндемиков и очень небольшое число общих с Сибирью местных видов говорит о том, что большинство связей между лесными регионами оборвались давно. Судя по родственным связям, представители лишь немногих групп демонстрируют обратный тренд, т.е. вселились в горы Центрального Китая с севера – из хвойных лесов Сибири или даже Северной Америки. Из них уже стали эндемиками видового уровня рябчик Северцова, а также не встреченные нами сычуаньская кукша (Perisoreus internigrans), китайский трехпалый дятел (Picoides funebris), возможно – неясыть Давида, а вот родового уровня никто пока не достиг. Сложнее с более поздними вселенцами – северное происхождение логично предположить для местного мохноногого сыча, а из млекопитающих – для бурундука (Eutamias sibiricus). Определить, какие географические расы и популяции пришли с севера, продолжается ли фаунистический обмен между лесными птицами Сибири и восточных окраин Тибета, помогут планируемые филогеографические исследования.
Список литературы
Банникова А.А., Абрамов А.В., Лебедев В.С., Шефтель Б.И., 2017. Неожиданное генетическое разнообразие азиатских короткохвостых землероек рода Blarinella (Mammalia, Lipotyphla, Soricidae) // Доклады АН. Т. 474. № 1. С. 132–136.
Березовский М.М., Бианки В.Л., 1891. Птицы Ганьсуйского путешествия Г.Н. Потанина. 1884–1887. Материалы по орнитологии Китая, главным образом южной части провинции Гань-су. СПб. 270 с.
Бобров В.В., 1997. О границе между Палеарктическим и Индо-Малайским фаунистическими царствами в материковой части Азии (по данным о распространении ящериц (Reptilia, Sauria)) // Известия АН. Серия биол. № 5. С. 580–591.
Бобров В.В., 2005. О самостоятельности Центральноазиатской горной фаунистической области (по данным о распространении ящериц (Reptilia, Sauria)) // Известия АН, Серия биол. № 6. С. 694–709.
Бобров В.В., 2012. Герпетологические исследования в Центральном Китае (провинции Ганьсу и Сычуань) // Ананьева Н.Б., Новицкий Р.В. и др. (ред.). Вопросы герпетологии. Материалы Пятого съезда Герпетологического общества им. А. М. Никольского, Минск, 24–27 сентября 2012 г. Минск: ИООО “Право и экономика”. С. 37–41.
Бобров В.В., Шефтель Б.И., Сун Ю.-Х., Фанг Ю., Александров Д.Ю., Артамонова В.С., Банникова А.А., Демидова Т.Б., Коблик Е.А., Махров А.А., 2017. Исследования позвоночных животных в юго-западном Китае // Российские гималайские исследования: вчера, сегодня, завтра. Сборник научных статей. Под ред. Боркина Л.Я.. СПб. С. 152–157.
Боркин Л.Я., Литвинчук С.Н., 2016. О фауне амфибий Тибета (предварительный анализ) // Вестник СПбГУ. Серия 3, биология. Вып. 3. С. 25–30.
Волков С.В., Коблик Е.А. Птицы Мира: рекомендуемые русские названия видов; http://zmmu.msu.ru/spec/ publikacii/neserijnye-izdaniya/pticy-mira-rekomenduemye-russkie-nazvaniya-vidov
Жамбю М., 1988. Иерархический кластер-анализ и соответствия. М.: Финансы и статистика. 342 с. (Перевод с французского Б.Г. Миркина).
Коблик Е.А., Архипов В.Ю., Волков С.В., Мосалов А.А., Редькин Я.А., 2017. Гималаи – “ключ” к пониманию таксономического разнообразия азиатских пеночек (Phylloscopidae, Aves) // Российские гималайские исследования: вчера, сегодня, завтра. Сборник научных статей. Под ред. Боркина Л.Я. СПб. С. 173–178.
Коблик Е.А., Банникова А.А., Махров А.А., Шефтель Б.И., Бобров В.В., Артамонова В.С., Александров Д.Ю., 2014. Тибет: последняя пастораль Земли // Природа. № 5. С. 78–88.
Коблик Е.А., Черняховский М.Е., Волцит О.В., Васильева А.Б., Формозов Н.А., 2000. Некоторые характеристики первостепенного фаунистического рубежа в Непальских Гималаях // Бюлетень Московского общества испытателей природы, Отд. биол. Т. 105. № 4. С. 3–21.
Махров А.А., Артамонова В.С., Бобров В.В., Коблик Е.А., Лебедев В.С., Павлова С.В., Шефтель Б.И., 2019. От субтропиков до тайги и тундростепей: путешествие по горам Цинлин и восточному краю Тибета // Природа. № 3. С. 70–83.
Махров А.А., Артамонова В.С., Карабанов Д.П., 2013. Обнаружение амурского чебачка Pseudorasbora parva (Temminck et Schlegel) (Actinopterygii: Cyprinidae) в бассейне реки Брахмапутра (Тибетское плато, Китай) // Российский журнал биологических инвазий. Т. 6. № 1. С. 66–74.
Назаренко А.А., 1985. Некоторые историко-биогеографические проблемы, связанные с Гималаями (на примере дендрофильной орнитофауны) // Журнал общей биологии. Т. 46. № 1. С. 41–54.
Назаренко А.А., 1990. Орнитофаунистический обмен между Южной и Северной Азией на восточной периферии континента: последний ледниково-межледниковый цикл // Журнал общей биологии. Т. 51. № 1. С. 89–106.
Поплавская Н.С., Банникова А.А., Фанг Ю., Шефтель Б.И., Ушакова М.В., Суров А.В., Лебедев В.С., 2018. Центр происхождения длиннохвостого хомячка, Cricetulus longicaudatus Milne-Edwards, 1867 (Rodentia, Cricetidae) – Тибетское плато? // Доклады АН. Т. 479. № 6. С. 716–719.
Шефтель Б.И., Банникова А.А., Фанг Ю., Демидова Т.Б., Александров Д.Ю., Лебедев В.С., Сун Ю.-Х., 2017. Заметки по фауне, систематике и экологии мелких млекопитающих юга провинции Ганьсу (Китайская Народная Республика) // Зоологический журнал. Т. 96. № 2. С. 232–248.
Шефтель Б.И., Махров А.А., Бобров В.В., Артамонова В.С., Александров Д.Ю., Коблик Е.А., Банникова А.А., 2013. Затерянный мир “Лианхуашаня” // Природа. № 7. С. 56–65.
Штегман Б.К., 1938. Основы орнитографического деления Палеарктики. Т. 1. Вып. 2. Фауна СССР. Нов. сер. № 19 : Птицы. М.-Л.: Изд-во АН СССР. 156 с.
del Hoyo J., Elliott A., Sargatal J., Christie D.A. & de Juana E. (eds). Handbook of the Birds of the World Alive. Lynx Edicions, Barcelona; https://www.hbw.com/node/55095
Martens J., Tietze D.T., Päckert M., 2011. Phylogeny, biodiversity, and species limits of passerine birds in the Sino-Himalayan region – a critical review // Ornithological Monographs. 70. P. 64–94.
Mosbrugger V., Favre A., Muellner-Riehl A.N., Päckert M., Mulch A., 2018. Cenozoic Evolution of Geobiodiversity in the Tibeto-Himalayan Region // Mountains, Climate and Biodiversity. Hoorn C., Perrigo A., Antonelli A., eds. Oxford. P. 429–448.
Nazarenko A.A., 1990. Recent history of the East Palearctic avifauna: transzonal interchange of the forest elements between South and North Asia since the last 35 000 years // Proceeding Intel 100 DO-G Meeting. Bonn. P. 81–87.
Norton C.J., Jin C., Wang Yu., Zhang Y., 2010. Rethinking the Palearctic-Oriental Biogeographic Boundary in Quaternary China // Asian Paleoantropology. From Africa to China and Beyond. Norton C.J., Braun D.R., eds. Dordrecht, Heidelberg, London, New York. Springer. P. 81–100.
Päckert M., Martens J., Sun Yue-Hua, Severinghaus L.L., Nazarenko A.A., Ji T. Topfer T., Tietze D.T., 2012. Horizontal and elevational phylogeographic patterns of Himalayan and Southeast Asian forest passerines (Aves: Passeriformes).] Publication online 20.10.2011 // Journal of Biogeography. V. 39 (3). P. 556–573.
Päckert M., Martens J., Sun Yue-Hua, Tietze D.T., 2015. Evolutionary history of passerine birds (Aves: Passeriformes) from the Qinghai-Tibetan plateau: from a pre-Quarternary perspective to an integrative biodiversity assessment. Publication online 12.03.2015 // J. Ornithol. https://doi.org/10.1007/s10336-015-1185-6
Price T.D., 2005. Correlated evolution of ecological differences among the Old World Leaf Warblers in the breeding and nonbreeding seasons // Birds of two worlds: the ecology and evolution of migration (ed. by R. Greenberg, P.P. Marra)/ Baltimore, MD. Johns Hopkins University Press. P. 359–370.
Romanov A.A., Koblik E.A., Melikhova E.V., Arkhipov V.Yu., Golubev S.V., Volchenkova M.V., Astakhova M.A., 2016. Richness in Bird Species of the Eastern Himalayas in Early Spring. // Contemporary Problems of Ecology. V. 9(5). P. 529–534. https://doi.org/10.1134/S1995425516050127
Sun Yue-Hua, Fang Yun, Klaus S., Martens J., Scherzinger W., Swenson J.E., 2008. Nature of the Lianhuashan Natural Reserve. Liaoling science and technology publishing house. 100 p.
Дополнительные материалы отсутствуют.
Инструменты
Зоологический журнал