

Зоологический журнал, 2020, T. 99, № 4, стр. 436-449
Об иерархии факторов, определяющих сроки начала прилета воробьеобразных птиц (Passeriformes) в Приобскую лесотундру
В. Н. Рыжановский a, *, А. В. Гилев a, **
a Институт экологии растений и животных УрО РАН
620144 Екатеринбург, Россия
* E-mail: ryzhanovsky@ipae.uran.ru
** E-mail: gilev@ipae.uran.ru
Поступила в редакцию 06.02.2019
После доработки 24.05.2019
Принята к публикации 05.06.2019
Аннотация
Анализируются факторы, определяющие сроки весеннего прилета воробьеобразных птиц в гнездовую область – Нижнее Приобье. Для периода 1971–2015 гг. выявлен достоверный отрицательный тренд смещения прилета птиц на более ранние даты для 7 видов из 16, не достоверный для 6 видов, положительный тренд лишь для 3 видов. С помощью кластерного анализа выделено три группы видов со сходной динамикой прилета. Температура воздуха в районе исследования в начале прилета всех видов повышалась по мере приближения к концу анализируемого периода (1971–2015 гг.), параллельно росту весенних среднемесячных температур. Выявлены значимые корреляции сроков начала прилета птиц с весенними температурами воздуха (преимущественно мая) и датами прохода льда по р. Обь и длиной миграционного пути вида. Связь же даты начала прилета у большинства видов со среднесуточной температурой воздуха непосредственно в день прилета выражена слабо. Таким образом, основными факторами, влияющими на даты прилета, являются среднемесячные весенние температуры воздуха, в первую очередь, мая. Глобальный погодный индекс Северо-Атлантического Колебания (NAO) играет существенно меньшее значение.
Факторы, определяющие сроки прилета воробьеобразных птиц из зимовочной части ареала в гнездовую область на пространстве Северной Евразии анализировались неоднократно. Отметим публикации последних лет, выполненные на основе долговременных наблюдений: Соколов и др., 2001; Пасхальный, 2002; Golovatin, Paskhalny, 2003; Lehikoinen et al., 2004; Соколов, 2006; Соколов, Гордиенко, 2008; Грищенко, 2010; Moller et al., 2010; Венгеров, 2011, 2015; Захаров, 2016; Соколов, Цвей, 2016, Соколов, 2017, Lehikoinen et al., 2019. В этих работах чаще всего обсуждается влияние глобальных или локальных температур воздуха на сроки прилета птиц. В работах, выполненных Л.В. Соколовым (2006) и немецкими исследователями (Haest et al., in press), рассматривается также связь сроков весенней миграции с другими климатическими показателями – глобальным погодным индексом NAO-САК (Северо-Атлантическое Колебание). Причем показано влияние NAO не только на погодную ситуацию в Европе и Северной Африке, но и в Евразии (Sokolov et al., 2003), что должно отражаться на сроках прилета птиц, даже в удаленных от Европы районах исследования. Обзор современных представлений о механизмах контроля сроков весенней миграции у птиц опубликован Соколовым и Цвеем (2016) в Зоологическом журнале.
В Субарктике, по мнению Головатина и Пасхального (Golovatin, Paskhalny, 2003), птицы появляются в районе гнездования независимо от хода весенних процессов, не раньше определенного срока, поскольку этот срок у каждого вида запрограммирован генетически. Конкретная же дата их появления, вероятно, корректируется на месте и зависит от времени наступления благоприятных погодных условий в данное время. Такая программа – программа эндогенного контроля сроков прилета вида в гнездовой ареал – может быть не только видоспецифичной, считают авторы, но и “популяционно-специфичной”, т.е. в запрограммированные сроки прилетает не только вид, но и образующие его популяции (Дольник, 1975; Helm et al., 2009; Носков, Рымкевич, 2010). Влиянием климата и “внутренних часов” на сроки прилета птиц факторы не исчерпываются. Известно влияние светового режима (фотопериода) на сроки отлета птиц из района зимовки и на сроки появления их в районе гнездования (Helm et al., 2009; Носков, Рымкевич, 2010; Соколов, Цвей, 2016). Определенное влияние на сроки прилета птиц оказывает длина миграционного пути, т.е. расположение ареала зимовки относительно района гнездования, а также погодные условия непосредственно на местах их зимовки (Sokolov, Kosarev, 2003; Соколов, Цвей, 2016). Возможно влияние и других факторов – ландшафта, орографии и т.п., а также, конечно, экологии вида, связанной с его питанием – вид прилетает тогда, когда появляется доступная пища и освобождается от снега его биотоп обитания (Лэк, 1957).
Основная задача нашей работы – определение конкретных факторов, влияющих на сроки прилета воробьеобразных (Passeriformes) птиц массовых видов на широту Полярного круга в Ямало-Ненецком автономном округе, а также оценка степени влияния каждого фактора на этот процесс.
МАТЕРИАЛЫ И МЕТОДЫ
Регистрации первой весенней встречи исследуемых видов начались в 1971 г. и не прекращаются по настоящее время. Чаще всего эти наблюдения проводились на территории г. Лабытнанги (66°40′ с.ш., 63°30′ в.д.) и его окрестностях. В 1971–1975 гг. сроки прилета птиц регистрировали на полевом стационаре Харп, в 12 км от г. Лабытнанги; в 1976–1977 в долине р. Собь на ж. д. полустанке Красный камень (66°50′ с.ш., 65°55′ в.д.). Начиная с 1978 г. мы работали в долине Оби, в 5 км от г. Лабытнанги (стационар Октябрьский), где отлавливали птиц большой ловушкой и паутинными сетями. При этом для заполнения таблицы начала прилета мы пользовались данными коллег, зоологов стационара АНИС УрО РАН, если они встретили вид раньше нас. Анализировались также опубликованные данные коллег (Пасхальный, 2002; Пасхальный Головатин, 2007). В последние годы даты начала прилета некоторых видов в г. Лабытнанги нам предоставлял директор АНИС УрО РАН В.Г. Штро. Из Центра кольцевания ИПЭЭ РАН мы получали данные (возвраты) о птицах, ранее окольцованных, преимущественно, нами и коллегами.
Мы не беремся утверждать, что даты начала прилета вида всегда точные. Прилет таких заметных птиц, как серая ворона (Corvus cornix), пуночка (Plectrophenax nivalis), белая трясогузка (Motacilla alba), рябинник (Turdus pilaris), подорожник (Calcarius lapponicus) регистрируется обычно в течение первых суток, но прочие виды, которых труднее наблюдать, мы либо отлавливали, либо регистрировали их песню иногда только с началом массового прилета или с прилетом первых самцов локальной популяции. Однако поскольку общая последовательность прилета видов сохранялась – луговые коньки (Anthus pratensis) наблюдались раньше краснозобых коньков (A. cervinus), тростниковые овсянки (Emberiza schoeniclus) раньше овсянки-крошки (E. pusilla), веснички (Phylloscopus trochilus) и теньковки (Ph. collybita) раньше таловок (Ph. borealis), допущенные неточности в регистрации прилета вида существенно не искажали общую картину весенней миграции.
Погодный индекс NAO-САК (Северо-Атлантическое Колебание) является показателем метеорологической ситуации в Атлантическом океане в районе Северной Африки и Западной Европы, т.е. в регионе, где зимует значительная часть северных видов воробьеобразных. Ежемесячные индексы САК (NAO) архивируются в National Oceanic and Atmospheric Administration’s Climate Prediction Center website (www.cpс.ncep.noaa.gov/ data/teledoc/nao.html). Вслед за Соколовым (2006) мы использовали показатели NAO-САК за январь–март 1971–2015 гг. для выявления связи между сроками начала прилета и данным индексом. Районы зимовки птиц из Нижнего Приобья определялись по литературным источникам, по отловам птиц с чужими кольцами и по возвратам колец от меченых нами птиц. Источником сведений о температуре воздуха в марте–июне 1971–2015 гг. были данные по метеостанции Салехард (WMO # 233 300, 66°31′ с.ш., 66°36′ в.д., 35 м над ур. м.).
Материал был обработан традиционными для анализа многолетних рядов динамики способами. Сравнение рядов динамики дат прилета и климата проводилось с помощью корреляционного (коэффициенты корреляции Пирсона и Спирмена) и регрессионного анализа. Правомерность применения метода ранговой корреляции Спирмена для выявления влияния температурного фактора на долговременные флуктуации сроков миграции показана Соколовым с соавторами (2001). Для выявления наиболее общих трендов в динамике дат прилета было выполнено сглаживание методом скользящей средней по 7 смежным значениям. Выявление групп птиц со сходной динамикой дат прилета проводилось методами кластерного анализа. Для выявления ведущих факторов, определяющих даты прилета, был использован анализ главных компонент. Все расчеты выполнены в программах Microsoft Excel 2003 и Statistica 6.0 (StatSoft Ink. 1984–2000).
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Динамика климатических условий Нижнего Приобья
Апрель и май, когда начинается прилет большинства видов птиц на широту Полярного круга, характеризуется постепенным повышением температуры воздуха от –20–30°С в первой декаде апреля (min –33.4°С 06.04.2006 г.) до положительных в отдельные дни мая (max 24.5°С 26.05.1991 г.) и постоянно положительных, начиная со второй декады июня. В период наших наблюдений (с 1971 г.) отмечается постепенное повышение среднемесячной температуры воздуха в апреле–июне (рис. 1). И если апрельский тренд повышения температуры выражен незначительно, то в мае и июне тренд становится явным: чем ближе к 2015 г., тем выше среднемесячные температуры. Майские и июньские тренды приобретают положительные, высоко достоверные значения (достоверность коэффициентов корреляции для апреля p = 0.190, а для мая p = 0.001, для июня p = 0.000). Это свидетельствует о том, что устойчивое потепление на севере Западной Сибири действительно имеет место. Рост среднегодовой приземной температуры в конце прошлого века достигал 0.03–0.07°С в год (Павлов, Гравис, 2000), не снижается он и в настоящее время. Существенное повышение весенних температур в последние годы отмечается во многих странах Европы, на Северо-западе России, в Байкальском регионе и на Камчатке (Соколов, 2006, 2017). При этом на Южном Урале в период с 1971 по 2005 гг. в марте и апреле значимых изменений тренда температуры не выявлено (Соколов, Гордиенко, 2008).
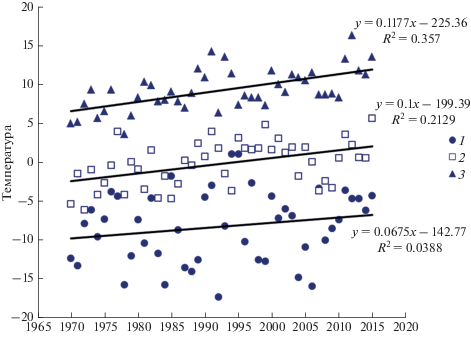
Сроки, последовательность и динамика прилета птиц
Прилет воробьиных птиц в Нижнее Приобье начинают пуночка (24.3–4.05) и серая ворона (30.3–18.4), заканчивают таловка (26.5–15.6), камышевка-барсучок (Acrocephalus schoenobaenus) и береговушка (Riparia riparia) (табл. 1). Можно выделить три волны прилета видов: апрельскую (пуночка и серая ворона); волну первой половины мая и волну конца мая – первой половины июня. Во вторую волну, с появлением воды (заберегов) на Оби и преимущественно положительных температурах воздуха, прилетают первые рогатые жаворонки (Eremophila alpestris) и белые трясогузки. При наличии проталин на полянах, в ранне- и средневесенние годы начинался прилет рябинников, тростниковых овсянок, юрков (Fringilla montifringilla). В годы с поздней весной этой волны не было, она объединялась с последней, основной волной прилета. Начинается эта волна массовым (не ежегодным) прилетом таежных чечеток, а заканчивается, как уже упоминалось выше, таловками, береговушками, камышевками-барсучками. Флуктуация сроков начала прилета сильно выражена, она составляет 20–40 сут (табл. 1) и не меньше, чем в умеренных широтах, например, на Южном Урале (Соколов, Гордиенко, 2008).
Таблица 1.
Сроки начала прилета птиц в Нижнее Приобье и их тренды в период с 1971 по 2015 гг.
| Вид | Число, лет | Первые даты прилета, lim (средняя дата) |
Показатели линейной регрессии | ||
|---|---|---|---|---|---|
| наклон | r | P | |||
| Eremophila alpestris | 28 | 1.05–30.05 (18.05) |
0.037 | 0.055 | 0.780 |
| Anthus pratensis | 26 | 3.05–14.06 (21.05) |
–0.397 | –0.425 | 0.030* |
| Anthus cervinus | 21 | 17.05–12.06 (26.05) |
–0.278 | –0.332 | 0.142 |
| Motacilla flava | 22 | 19.05–11.06 (31.05) |
–0.354 | –0.472 | 0.023* |
| Motacilla alba | 42 | 4.05–2.06 (12.05) |
–0.055 | –0.091 | 0.566 |
| Corvus cornix | 42 | 30.03–18.04 (7.04) |
–0.124 | –0.325 | 0.035* |
| Phylloscopus trochilus | 29 | 18.05–9.06 (26.05) |
–0.141 | –0.297 | 0.111 |
| Phylloscopus collybita | 17 | 22.05–15.06 (2.06) |
–0.275 | –0.566 | 0.018* |
| Phylloscopus borealis | 27 | 26.05–13.06 (7.06) |
0.023 | 0.072 | 0.720 |
| Luscinia svecica | 27 | 6.05–7.06 (27.05) |
–0.106 | –0.163 | 0.415 |
| Turdus pilaris | 26 | 5.05–1.06 (19.05) |
–0.236 | –0.349 | 0.080 |
| Turdus iliacus | 32 | 11.05–6.06 (21.05) |
–0.248 | –0.360 | 0.043* |
| Emberiza schoeniclus | 21 | 3.05–7.06 (12.05) |
–0.389 | –0.319 | 0.159 |
| Emberiza pusilla | 29 | 14.05–7.06 (26.05) |
–0.255 | –0.418 | 0.024* |
| Calcarius lapponicus | 23 | 18.05–10.06 (26.05) |
–0.186 | –0.355 | 0.097 |
| Plectrophenax nivalis | 39 | 24.03–2.05 (7.04) |
0.444 | 0.593 | 0.0001*** |
Ранее Пасхальный (2002) для серой вороны, лугового конька (Anthus pratensis), белой трясогузки, белобровика (Turdus iliacus), веснички (Phylloscopus trochilus) и юрка показал достоверный отрицательный тренд начала прилета в Нижнее Приобье за период с 1970–2002 гг. Наши данные за несколько больший ряд лет, в целом, подтвердили тенденцию смещения сроков прилета этих видов на более ранние даты (табл. 1). Достоверный отрицательный тренд найден для серой вороны, лугового конька, желтой трясогузки, белобровика, овсянки-крошки и теньковки. Для остальных видов, кроме пуночки, рогатого жаворонка и таловки, характерно недостоверное смещение прилета на ранние даты. У последних (пуночка, рогатый жаворонок, таловка) тренды положительные, причем у пуночки они высоко достоверные, свидетельствующие о смещении сроков начала прилета на более поздние даты. Для рогатого жаворонка, как мы ранее указывали (Рыжановский, 2015), в 1970–1980-е гг. массовый пролет наблюдался до ледохода и интенсивного таяния снега на плакоре, в настоящее же время жаворонки нередко летят значительно позднее, над свободной ото льда рекой и лишенной снега тундрой. Прилет таловки изменился незначительно.
На Южном Урале даты начала прилета воробьиных в период 1971–2005 гг. не изменились (Соколов, Гордиенко, 2008). Авторы отметили явное противоречие между данными по Ильменскому заповеднику и достоверными данными о более раннем прилете многих видов в Западную Европу и Прибалтику во второй половине 20 века (Peintinger, Schuster, 2005; Hüppop, Hüppop, 2005; Соколов, 2006). По мнению авторов, это связано с тем, что на Южном Урале в указанные годы не наблюдалось потепления климата в апреле и мае, когда прилетает большинство видов птиц. На севере Европы также отмечено, что дальние мигранты, зимующие в Африке, в меньшей степени среагировали на потепление климата, чем мигранты из Южной Европы (Saino et al., 2015). Головатин и Пасхальный (Golovatin, Paskhalny, 2003) считают, что у видов, обитающих на севере Субарктики, т.е. настоящих северян, сдвига в сторону более раннего прилета не происходит, это больше относится к видам, проникающим в этот регион с юга. Согласно нашим данным, достоверное смещение сроков прилета на ранние даты, действительно, более характерно для проникающих видов, а не для связанных происхождением с Субарктикой (табл. 1). Причем дальние мигранты (весничка, таловка, варакушка) в меньшей степени среагировали на потепление климата в Субарктике, нежели мигранты из Южной Европы, о чем свидетельствуют недостоверные тренды сроков начала прилета этих видов в наш район исследования (табл. 1).
Наши и литературные данные (Венгеров, 2015; Захаров, 2016) показывают, что практически для всех изученных видов птиц умеренных и высоких широт динамика сроков начала миграции носит сложный нелинейный характер характеризующаяся сменой подъемов и спадов. С целью сглаживания случайных колебаний и выявления наиболее общих тенденций в рядах временнóй динамики традиционно применяется метод скользящей средней. Мы использовали скользящие средние по семи значениям для периода 1975–2005 гг., когда регистрировали начало прилета всех анализируемых видов.
По сглаженным рядам динамики прилета был проведен кластерный анализ (рис. 2). В результате выявлено три группы птиц, сходных по динамике прилета. Первая группа включает серую ворону, лугового конька, желтую трясогузку, весничку, белобровика, овсянку-крошку, тростниковую овсянку и подорожника (рис. 3а, 3б). Она отличается большим сходством динамики прилета. Вначале, в период с 1975 по 1990 гг., наблюдался резкий переход к более ранним срокам прилета, затем наметился переход к более поздним срокам, позже начался повторный переход на более ранние даты, у большинства видов достаточно резкий. У подорожника выявлен опережающий сдвиг на несколько лет в сторону более ранних дат, по сравнению с вороной и остальными видами.
Рис. 2.
Дендрограмма сходства видов по динамике дат прилета (мера сходства – коэффициент корреляции Пирсона, способ объединения – полное присоединение).
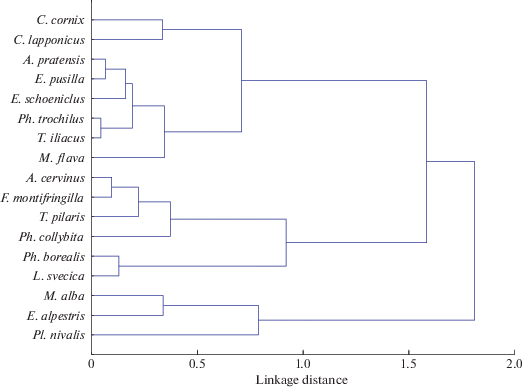
Рис. 3.
Динамика сглаженных дат прилета птиц первой (а, б), второй (в) и третьей (г) групп. 1 – ворона, 2 – луговой конек, 3 – желтая трясогузка, 4 – подорожник, 5 – весничка, 6 – белобровик, 7 – тростниковая овсянка, 8 – овсянка-крошка, 9 – краснозобый конек, 10 – таловка, 11 – теньковка, 12 – варакушка, 13 – рябинник, 14 – юрок, 15 – белая трясогузка, 16 – пуночка, 17 – рогатый жаворонок.
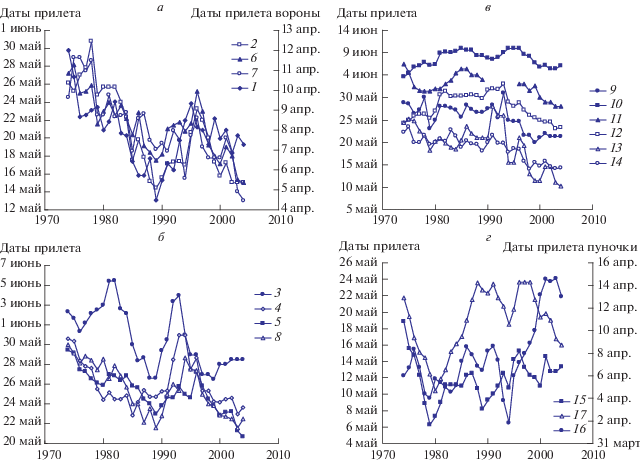
Вторая группа включает краснозобого конька, теньковку, таловку, варакушку, рябинника и юрка (рис. 3в). У всех этих видов до 1990-х гг. сроки прилета колебались около одного уровня, с большим или меньшим размахом, но затем резко сместились к более ранним датам и начали колебаться вблизи нового уровня. Во многих случаях наблюдается поразительное совпадение пиков и спадов и это притом, что на рисунках приведена скользящая средняя, которая нивелирует прямые показатели.
Третья группа объединяет белую трясогузку, пуночку и рогатого жаворонка, она характеризуется вначале резким переходом к более ранним срокам прилета в конце 1970-х гг., затем переходом к более поздним датам (рис. 3г). Дальнейшая динамика у видов этой группы несколько различается, смещение к более поздним датам выражено у них в разной степени. В середине 1990-х у видов этой группы также наблюдался небольшой переход к ранним срокам прилета.
Общеизвестно, что сроки прилета птиц в умеренные и высокие широты зависят от погодных условий весны. Возможно, сходство динамик прилета отдельных видов в Субарктику связано не с температурой воздуха непосредственно в Нижнем Приобье, а с метеорологическими условиями на трассе весенней миграции этих видов, что не позволяет исключить действия других факторов, влияющих на сроки начала прилета птиц в Заполярье.
Влияние глобальных климатических факторов на сроки прилета птиц
При изучении влияния зимне-весеннего климата в Западной Европе и Северной Африке на сроки прилета птиц на Куршскую косу Балтийского моря была выявлена значимая связь сроков миграции некоторых воробьиных видов с погодным индексом NAO (Соколов, 2006), что, учитывая близость косы к Атлантическому океану, не удивительно. Однако Соколов (2010), проанализировав опубликованные Пасхальным (2002) сроки начала прилета птиц в Нижнее Приобье за 1970–2002 гг., выявил значимую корреляционную связь индекса NAO с началом прилета у серой вороны и пеночки-веснички, близкую к достоверности у лугового конька, и отсутствие связи у белой трясогузки, белобровика и юрка. Поскольку мы имеем более длинный ряд лет и большее число видов, все материалы были пересчитаны заново с тем же средним, за январь–март, значением индекс NAO. В нашем случае достоверная связь NAO с датами начала прилета за 1971–2015 гг. по Пирсону и Спирмену найдена для серой вороны, достаточно высокая, но не достоверная по обоим показателям корреляции, для лугового конька, овсянки-крошки, рогатого жаворонка, и не достоверная по одному показателю для желтой и белой трясогузок, краснозобого конька и теньковки (табл. 2). Для остальных рассмотренных видов корреляция отсутствует.
Таблица 2.
Связь сроков прилета птиц (прямые и сглаженные даты) с индексом NAO
| Вид | Коэффициент ранговой корреляции Спирмена | Коэффициент корреляции Пирсона | ||
|---|---|---|---|---|
| прямые даты | сглаженные даты | прямые даты | сглаженные даты | |
| E. alpestris | 0.238 | 0.750*** | 0.278 | 0.766*** |
| A. pratensis | –0.289 | –0.663*** | –0.320 | –0.717*** |
| A. cervinus | –0.042 | –0.158 | –0.228 | –0.128 |
| M. flava | –0.062 | –0.385* | –0.203 | –0.430* |
| M. alba | –0.128 | 0.031 | –0.215 | 0.060 |
| С. соrnix | –0.439** | –0.290 | –0.438** | –0.310* |
| Ph. trochilus | –0.044 | –0.523** | –0.171 | –0.434* |
| Ph. collybita | 0.278 | 0.099 | 0.140 | 0.033 |
| Ph. borealis | –0.079 | 0.410** | –0.194 | 0.443* |
| L. svecica | 0.097 | 0.404* | 0.006 | 0.440* |
| T. pilaris | 0.028 | 0.068 | –0.020 | 0.095 |
| T. iliacus | –0.020 | –0.506*** | –0.056 | –0.417** |
| F. montifringilla | 0.013 | –0.361* | 0.030 | –0.269 |
| E. schoeniclus | –0.074 | –0.719*** | –0.162 | –0.721*** |
| E. pusilla | –0.267 | –0.443* | –0.268 | –0.460** |
| C. lapponicus | –0.093 | 0.127 | –0.149 | 0.070 |
| Pl. nivalis | 0.106 | –0.021 | 0.058 | –0.182 |
В табл. 2 представлены также корреляции индекса NAO и трендов сглаженных дат прилета. В этом случае корреляции достоверны для большинства проанализированных видов, зимующих как в Африке и Южной Европе, так и в Азии. У части видов выявлена отрицательная корреляция, у части положительная. Достоверная отрицательная корреляция характерна для видов (серая ворона, луговой конек, желтая трясогузка, весничка, белобровик, юрок, камышевая овсянка), зимующих к югу и юго-западу от Нижнего Приобья (табл. 2). Для видов, зимующих на юго-востоке (варакушка и таловка), характерна положительная корреляция. Возможно, на юго-востоке, в Монголии, зимуют и рогатые жаворонки, поскольку они входят в одну с таловкой и варакушкой группу. Овсянка-крошка, имеющая отрицательную корреляцию, также зимует на юго-востоке, хотя по результатам кластерного анализа входит в группу мигрантов с юго-запада (рис. 2).
Поскольку индекс NAO отражает зимне-весенний климат в Северной Африке и Западной Европе, никакой связи этого атлантического индекса с динамикой прилета птиц в Урало-Сибирский регион вообще не должно быть, как не найдена такая связь для птиц Южного Урала (Соколов, Гордиенко, 2008). Но для Нижнего Приобья, как оказалось, такая корреляция существует (табл. 2). Возможно, причиной этого является то, что зимой и весной все же имеет место западный перенос теплых воздушных масс с Атлантического океана в этот регион. Соколов с соавторами (Sokolov et al., 2003) показали наличие значимой положительной связи индекса NAO с зимними и весенними (в марте) температурами воздуха на почти всей территории Евразии вплоть до Охотского моря. Кроме этого, климатические условия, характеризуемые индексом NAO, в районе зимовки птиц в Северной Африке и Южной Европе, могут существенно влиять на сроки начала и темпы весенней миграции птиц, что не может не отражаться на времени прилета их в гнездовую область (Sokolov, Kosarev, 2003). Вероятно, глобальные изменения климата в зимне-весеннее время года могут влиять на сроки весеннего прилета воробьиных птиц и в наш район исследования, расположенный в Нижнем Приобье.
Влияние местных погодных факторов на прилет птиц
В качестве основного фактора, влияющего на сроки прилета птиц в Субарктику и Арктику, признаются климатические изменения, связанные, прежде всего, с колебаниями температуры воздуха (Успенский, 1969). В табл. 3 приведены температуры воздуха (температурные пороги) в районе нашего исследования, при которых начинается прилет птиц. Из всех исследованных видов только желтая трясогузка и таловка всегда прилетали в Нижнее Приобье при положительной температуре воздуха. Насекомоядные виды, прилетающие преимущественно во второй половине мая, появляются в нашем районе исследования обычно при положительных температурах, но в годы с поздней весной (при потеплении в конце мая – начале июня) – при отрицательных температурах. Наблюдения показывают, что при среднесуточной температуре выше –3°С, в ясную маловетреную погоду, к середине дня воздух прогревается до положительных величин, что может способствовать началу прилета ряда видов.
Таблица 3.
Минимальная, максимальная и средняя температура воздуха (°С) в первый день прилета птиц
| Вид | Температура (°С) прилета: мин; макс (средняя) |
Показатели линейной регрессии | ||
|---|---|---|---|---|
| наклон | r | P | ||
| E. alpestris | –4.0; 15.1 (1.1) | 0.123 | 0.297 | 0.132 |
| A. pratensis | –10.7; 17.2 (2.5) | 0.186 | 0.377 | 0.070 |
| A. cervinus | –2.8; 5.4 (2.5) | 0.070 | 0.273 | 0.258 |
| M. flava | 0.8; 6.8 (2.5) | 0.091 | 0.611 | 0.003** |
| M. alba | –8.8; 12.2 (0.3) | 0.124 | 0.404 | 0.010** |
| C. cornix | –23.6; 5.6 (–8.9) | 0.116 | 0.183 | 0.255 |
| Ph. trochilus | –3.6; 12.2 (3.4) | 0.118 | 0.396 | 0.025* |
| Ph. collybita | –0.6; 7.8 (3.6) | 0.061 | 0.333 | 0.163 |
| Ph. borealis | 0.8; 21.5 (7.4) | 0.112 | 0.290 | 0.127 |
| L. svecica | –1.8; 14.3 (3.1) | 0.077 | 0.207 | 0.272 |
| T. pilaris | –8.5; 11.9 (1.3) | 0.141 | 0.374 | 0.046* |
| T. iliacus | –1.6; 11.9 (2.5) | 0.041 | 0.171 | 0.341 |
| F. montifringilla | –2.8; 10.3 (2.4) | 0.034 | 0.014 | 0.542 |
| E. schoeniclus | –3.6; 14.1 (3.1) | 0.123 | 0.209 | 0.375 |
| E. pusilla | –5.9; 11.7 (2.5) | 0.147 | 0.392 | 0.026* |
| C. lapponicus | 10.9; 8.1 (1.0) | 0.090 | –0.029 | 0.896 |
| Pl. nivalis | –25.9; 2.9 (–9.7) | 0.199 | 0.336 | 0.049* |
Температура воздуха в период начала прилета всех видов повышалась по мере приближения к концу анализируемого временнóго ряда (1971–2015 гг.), параллельно росту среднемесячных температур (рис. 1). Многолетние тренды повышения температуры воздуха в день прилета оказались достоверны для 6 видов (табл. 3). Связь же среднесуточной температуры воздуха непосредственно в день прилета с датой появления большинства видов выражена слабо или вообще отсутствует. Только у желтой трясогузки обнаружена достоверная корреляция дат прилета с температурой воздуха в день прилета (табл. 4). У серой вороны, лугового конька, варакушки, белобровика, рябинника, веснички, овсянки-крошки и подорожника выявлены достоверные корреляции дат начала прилета с температурой в предыдущие два или три года. Чем это объясняется, сказать трудно. Вряд ли взрослые птицы, которые летят обычно первыми (Дольник, 1975), руководствуются погодной ситуацией прошлых лет. Возможно, это характерно для серых ворон, которые прилетают для участия в гнездовании на второй год жизни.
Таблица 4.
Связь даты начала прилета птиц с температурой воздуха (Т, °С) в день прилета и среднемесячными температурами
| Вид | Т °С в день прилета | Коэффициент корреляции Спирмена | |||
|---|---|---|---|---|---|
| март | апрель | май | июнь | ||
| E. alpestris | 0.231 | 0.115 | 0.084 | –0.241 | 0.092 |
| A. pratensis | –0.047 | –0.079 | –0.248 | –0.747*** | –0.332 |
| A. cervinus | 0.088 | –0.080 | –0.323 | –0.733*** | –0.379 |
| M. flava | –0.553* | –0.579** | –0.070 | –0.324 | –0.279 |
| M. alba | –0.040 | 0.061 | –0.290 | –0.551*** | –0.317* |
| С. сornix | –0.266 | –0.377* | –0.248 | –0.106 | –0.104 |
| Ph. trochilus | –0.322 | –0.195 | –0.110 | –0.775*** | –0.079 |
| Ph. borealis | –0.006 | –0.081 | –0.187 | –0.219 | –0.370 |
| Ph. collybita | –0.099 | 0.304 | 0.024 | –0.499* | –0.419 |
| L. svecica | –0.149 | –0.032 | –0.182 | –0.548** | –0.216 |
| T. pilaris | –0.213 | –0.008 | –0.624*** | –0.776*** | –0.427* |
| T. iliacus | 0.075 | –0.034 | –0.283 | –0.882*** | –0.229 |
| F. montifringilla | –0.089 | 0.033 | –0.131 | –0.656*** | –0.122 |
| E. schoeniclus | –0.305 | 0.041 | –0.367 | –0.722*** | –0.261 |
| E. pusilla | 0.088 | –0.224 | –0.066 | –0.485** | –0.221 |
| C. lapponicus | 0.111 | –0.177 | –0.190 | –0.589** | –0.364 |
| Pl. nivalis | 0.148 | –0.001 | –0.006 | 0.079 | 0.392* |
Температура воздуха в день прилета исследуемых видов – очень изменчивая величина в условиях Субарктики. Кратковременные подъемы и спады температуры могут и не оказывать существенного влияния на прилет птиц. Большее значение имеет общий ход весны, который можно охарактеризовать средними температурами за определенный период. В табл. 4 представлены коэффициенты корреляции Спирмена дат прилета птиц и среднемесячных температур с марта по июнь. Из таблицы видно, что для большинства видов птиц имеют место значимые корреляции со средними температурами мая – месяца, когда у подавляющего большинства птиц наблюдается прилет. У двух видов, вороны и желтой трясогузки, выявлены корреляции со средними температурами марта. Практически все эти корреляции – отрицательные, т.е. чем выше средние температуры, тем раньше наблюдается прилет. Для трех видов, пуночки, таловки и рогатого жаворонка, таких корреляций не отмечено. Для пуночки отмечена положительная корреляция со средними температурами июня: чем выше температуры, тем позже прилетают пуночки. Но пуночки прилетают в апреле, и остается непонятным, как на них могут влиять температуры июня. Скорее всего, это ложная корреляция. Также не понятна связь прилетающей в мае желтой трясогузки с температурой марта. Аналогично были рассчитаны корреляции для сглаженных рядов динамики дат прилета и температурных данных. Число достоверных корреляций в этом случае существенно возросло, однако вновь такие корреляции были отмечены преимущественно с температурами мая – месяца прилета подавляющего числа видов птиц. Для пуночки отмечены положительные корреляции с температурами апреля, мая и июня; для таловки – положительные корреляции с температурами марта, что также не понятно, как и в случае с желтой трясогузкой; для рогатого жаворонка найдены положительные корреляции со всеми месяцами весны. Коэффициенты корреляции для сглаженных рядов были в большинстве случаев существенно ниже, чем для исходных данных. Таким образом, наиболее важное значение имеют непосредственно температуры в месяц прилета птиц, нежели их многолетний тренд.
В долине Оби, как и других долинах крупных северных рек, текущих в меридиональном направлении, интегральным климатическим показателем являются сроки прохода льда. Головатин и Пасхальный (Golovatin, Paskhalny, 2003) отмечали высокие значения коэффициентов корреляции дат прилета и ледохода на притоке Нижней Оби, р. Войкар, водно-болотных птиц, связанных с водой, и совпадение сроков прилета и ледохода у других видов. Открытая вода приносит тепло, температура воздуха в долине Оби повышается, после чего, как правило, начинается массовый прилет воробьеобразных птиц, мигрирующих по берегам. Корреляции начала прилета первых представителей обсуждаемых видов не всегда высокие, но они есть (табл. 5). Сравнение значений коэффициентов корреляций табл. 4 и 5 показывает, что влияние среднемесячных температур воздуха на сроки начала прилета птиц в наш район исследования больше, чем влияние сроков ледохода.
Таблица 5.
Связь сроков начала прилета птиц в долину Нижней Оби с датой ледохода на р. Обь на створе г. Салехард
| Вид | Коэффициент корреляции Спирмена | Вид | Коэффициент корреляции Спирмена |
|---|---|---|---|
| E. alpestris | 0.083 | L. svecica | 0.430* |
| A. pratensis | 0.493* | T. pilaris | 0.705*** |
| A. cervinus | 0.527* | T. iliacus | 0.671*** |
| M. flava | 0.264 | F. montifringilla | 0.371 |
| M. alba | 0.611*** | E. schoeniclus | 0.605** |
| С. сornix | 0.275 | E. pusilla | 0.259 |
| Ph. trochilus | 0.500** | C. lapponicus | 0.399 |
| Ph. collybita | 0.386 | Pl. nivalis | –0.040 |
| Ph. borealis | 0.317 |
Фотопериодические интервалы сроков прилета птиц
Весенняя миграция птиц приходится на ту часть годового цикла, которая контролируется фотопериодической реакцией на увеличение длины дня. Согласно данным Носкова и Рымкевич (2010), эффект увеличивающейся продолжительности дня проявляется в характерном не только для каждого вида, но даже для разных подвидов и популяций, фотопериодическом интервале миграционной активности. Существование нижних и верхних порогов фотопериодического интервала, по мнению этих авторов, может влиять на время окончания весенней миграции у видов, гнездящихся в умеренных широтах. Особенностью широты Полярного круга, где находится район нашего исследования, является быстро растущий день, от 0 ч солнечного сияния в период зимнего солнцестояния до 24 ч в период летнего солнцестояния. Наиболее ранний мигрант, пуночка, встречен в г. Лабытнанги (24.03) при минимальном дне – 12 ч 40 мин солнечного сияния, первая регистрация серой вороны пришлась на 30 марта, при 13 ч 24 мин. В годы с наиболее поздним началом прилета пуночки появились при 17 ч 34 мин (2.05), вороны – при 15 ч 45 мин (18.04) солнечного сияния. Прилет других средних и дальних мигрантов начинается при белых ночах в первой половине мая и при солнечных ночах во второй половине мая–начале июня. Массовый прилет практически всех северных воробьиных, за исключением вороны и пуночки, имеет место во второй половине мая – начале июня при 24-часовом дне. Таким образом, верхнего порога фотопериодического интервала окончания весенней миграции у северных птиц не существует. Нижние фотопериодические пороги у исследованных видов разные (табл. 6). Следует обратить внимание на отсутствие регистраций прилета первых птиц, за исключением вороны и пуночки, раньше начала мая и продолжительности дня меньше 17 ч 30 мин. При этом во второй половине апреля в 1977, 1982, 1990, 1991 и 2007 гг. преобладали достаточно теплые дни (температура поднималась до 11.7°С), в третьей декаде апреля снег в населенных пунктах и на тундровых участках плакора сошел полностью, среднедекадная температура в эти годы была близка к нулю (от –2 до 1.7°С), однако прилета большинства видов не наблюдали. По-видимому, для прилета на широту Полярного круга основной массе видов помимо температурного порога следует преодолеть и свой фотопериодический порог. Т.е. северные популяции, видимо, имеют фотопериодические пороги для окончания весеннего миграционного состояния. В северную тайгу эти птицы прилетают раньше, при более коротком дне, что дает основание предполагать определенную ступенчатость фотопериодических интервалов весенней миграции, скоррелированную с широтой. Отметим, что к выводу о существовании у видов и популяций фотопериодических интервалов, в пределах которых функционируют элементы годового цикла, к настоящему времени пришли и другие исследователи (Гвиннер, 1984; Helm et al., 2009; Носков, Рымкевич, 2010; Соколов, Цвей, 2016).
Таблица 6.
Нижние пороги продолжительности дня (ч: мин) в период начала прилета птиц на широту Полярного круга
| Вид | Длина дня | Вид | Длина дня |
|---|---|---|---|
| С. сornix | 13:24 | Ph. collybita | 19:31 |
| A. pratensis | 18:06 | L. svecica | 18:06 |
| A. cervinus | 19:31 | T. pilaris | 17:58 |
| M. flava | 20:08 | T. iliacus | 18:47 |
| M. alba | 17:50 | E. schoeniclus | 17:42 |
| Pl. nivalis | 12:40 | F. montifringilla | 17:42 |
| C. lapponicus | 20:00 | E. pusilla | 18:14 |
| Ph. trochilus | 20:00 | E. alpestris | 17:26 |
| Ph. borealis | 21:43 |
Разумеется, для птиц важно не само солнечное сияние, а световой день определенной продолжительности, в середине мая в лесотундре за счет рефракции световой день становится “полярным”. В теплые весны птицы летят достаточно активно при длине солнечного дня, превышающей 18 ч света. В поздние весны в лесотундре прилет большинства видов начинается и протекает при солнечных ночах, в тундрах Среднего Ямала (68°–70° с.ш.) прилет всех птиц, кроме пуночек, независимо от климатических условий происходит в условиях не заходящего солнца.
Влияние дальности миграции на сроки прилета птиц
На север Западной Сибири для участия в размножении прилетают птицы, зимовавшие в разных регионах, от Юго-Восточной Азии до Западной Европы и Африки, от тропиков до тайги. Вероятные районы зимовки рассматривались в предыдущих разделах. Длину миграционного пути мы определяли прямой линией на глобусе. В природе миграционный путь, естественно, больше, поскольку птицы вынуждены огибать разного рода физические преграды в виде морей, горных хребтов и т.п. В результате анализа нами была выявлена достоверная положительная корреляция средней даты начала прилета вида в наш район исследования с длиной его миграционного пути (r = 0.71, p = 0.001): чем длиннее была трасса миграции, тем позже появлялись птицы.
Иерархия факторов, влияющих на сроки прилета птиц
Среди факторов, рассмотренных нами, можно выделить как относительно постоянные факторы (район зимовки и длина миграционного пути), так и переменные (индекс NAO, среднемесячные температуры, даты ледохода). Именно последняя группа факторов, как показал анализ, существенным образом влияет на даты прилета птиц, определяя их разброс. Поскольку для многих видов выявлены корреляции дат прилета с несколькими факторами, встает закономерный вопрос – какие из них являются наиболее значимыми? Ответ на него можно найти, просто сравнивая коэффициенты корреляции между собой. Наиболее высоко достоверные из них – корреляции со средними температурами мая (табл. 4). Но, поскольку климатические факторы часто связаны между собой, для оценки влияния каждого из них мы использовали многомерные методы обработки данных (метод главных компонент), позволяющие получить как общую картину, так и определить отдельный вклад в нее каждого фактора (рис. 4). В качестве основных переменных, формирующих факторную структуру, были взяты даты прилета птиц. Климатические факторы использовались в качестве вспомогательных переменных для интерпретации полученной картины.
Рис. 4.
Характер взаимосвязи начала прилета птиц с климатическими факторами (метод главных компонент): обозначения ГК 1 и ГК 2 – главные компоненты 1 и 2; Март, Апрель, Май – среднемесячные температуры воздуха; NAO – глобальный погодный индекс; Ледоход – время начала ледохода на р. Обь; E.a. – рогатый жаворонок, A.pr. –конек луговой, A.c. – конек краснозобый, M.f. – желтая трясогузка, M.a. – белая трясогузка, C.c. – серая ворона, Ph.t. – весничка, Ph.b.–таловка, L.s. – варакушка, T.i. – белобровик, E.p. – овсянка-крошка, C.l. – подорожник, Pl.n. – пуночка.
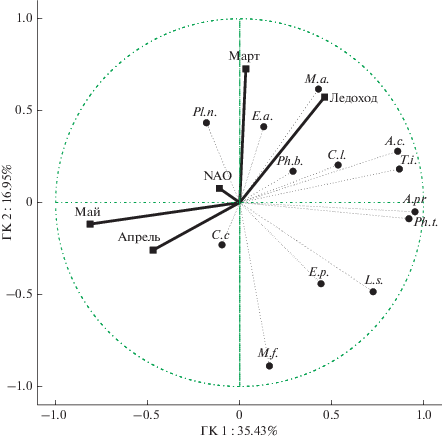
Расчеты показали, что первые две главные компоненты (ГК) объясняют вместе примерно 52% наблюдаемой дисперсии дат прилета. С первой компонентой теснее всего связаны средние температуры апреля и мая (35% наблюдаемой дисперсии), со второй – средние температуры марта и дата начала ледохода (17% дисперсии). Прочие учтенные нами климатические показатели не вносят существенного вклада в наблюдаемую картину. Оставшаяся дисперсия (48%) остается необъясненной и, следовательно, связана с еще какими-то невыясненными факторами. Особо следует подчеркнуть, что их влияние, по крайней мере, не меньше влияния тех факторов, которые мы рассмотрели.
Направления векторов на рис. 4 позволяет выявить наиболее тесные связи основных и вспомогательных переменных. Сроки прилета большинства видов птиц, как это было показано выше, тесно связаны со средними температурами мая, месяца, когда в нашем районе исследования появляется большинство видов. Соответствующие векторы на рис. 4 направлены в противоположные стороны. Даты прилета белой трясогузки и рогатого жаворонка, а также отчасти пуночки, положительно связаны с началом ледохода и температурами марта. С температурами марта отрицательно связаны даты прилета серой вороны и желтой трясогузки.
Таким образом, основными факторами, влияющими на даты прилета птиц в Приобскую лесотундру, являются локальные среднемесячные, но не среднесуточные, весенние температуры воздуха и время начала ледохода. Глобальный погодный индекс NAO играет существенно меньшую роль в нашем регионе исследования, поскольку он влияет на зимне-весенний климат на огромных пространствах – от Атлантического до Тихого океана и может не быть тесно связан с локальной погодной ситуацией в том или ином отдельном районе Евразии. В недавней работе немецкие исследователи (Haest et al., in press), которые провели мета-анализ сроков прилета 23 видов птиц на о-в Гельголанд (Германия) в период с 1960 г. по настоящее время, показали наличие значимого влияния NAO на весеннюю фенологию исследуемых видов, однако вклад этого глобального погодного фактора в межгодовые флуктуации сроков прилета птиц не превышал 6%.
ЗАКЛЮЧЕНИЕ
Исходя из полученных результатов, можно утверждать, что глобальные климатические изменения, происходящие в настоящее время, существенно влияют на сроки прилета птиц на Ямале, поскольку существует тесная связь средних весенних температур воздуха и сроков прилета большинства воробьеобразных видов птиц. Чем выше среднемесячные, но не среднесуточные, температуры в весенний период, тем раньше появляются птицы. Однако температурные тренды неустойчивы и характеризуются чередованием длительных периодов повышения и понижения средних температур воздуха, что определяет и соответствующую динамику сроков прилета большинства видов птиц. Именно это объясняет кажущееся противоречие между данными разных авторов, анализирующими разные периоды времени, подтверждающими или опровергающими факт смещения дат прилета птиц в Приобскую лесотундру на все более ранние сроки. Наше исследование показало, что четко выраженная тенденция к смещению сроков прилета у большинства видов на более ранние календарные даты, наблюдавшаяся в период с 1970 по 1990 гг., сменилась обратной тенденцией в 1990–1995 гг., а затем вновь продолжилась с 1995 г. по настоящее время, в результате чего общая тенденция за все исследованные годы стала менее выраженной.
Сроки весенней миграции птиц тесно связаны не только с погодой на местах их зимовки и на пролете, но и с продолжительностью их остановки на трассе, зависящей от наличия доступного корма и скорости накопления ими энергетических ресурсов, необходимых для дальнейшего полета. Сроки прилета птиц в гнездовую область также зависят от протяженности миграционного пути вида и популяции.
Судя по глобальным климатическим прогнозам, современная тенденция к потеплению климата в ближайшее десятилетие сохранится, а значит, и продолжится тенденция к смещению дат прилета птиц в Северном полушарии на более ранние сроки. Последствия этих существенных изменений в сроках миграции птиц для северных экосистем еще предстоит оценить.
Список литературы
Венгеров П.Д., 2011. Влияние изменений климата на сроки прилета и размножения певчего дрозда (Turdus philomelos) и зяблика (Fringilla coelebs) в Воронежском заповеднике // Успехи современной биологии. Т. 131. № 4. С. 416–424.
Венгеров П.Д., 2015. Сроки весеннего прилета птиц в Воронежском заповеднике на фоне длительных климатических изменений // Научные ведомости БелГУ. Серия: Естественные науки. Т. 30. № 3. (200). С. 82–92.
Гвиннер Э.,1984. Цирканнуальные системы // Биологические ритмы. М.: Мир. Т.1. С. 55–80.
Грищенко В.Н., 2010. Изменение сроков миграции птиц в Каневском заповеднике // Бранта: Сборник научных трудов Азово-Черноморской орнитологической станции. № 13. С. 33–39.
Дольник В.Р., 1975. Миграционное состояние птиц. М.: Наука. 398 с.
Захаров В.Д., 2016. Влияние изменений климата на сроки прилета птиц в Ильменский заповедник (Южный Урал) // Экология. № 6. С. 455–460.
Лэк Д., 1957. Численность животных и ее регуляция в природе. М.: Иностранная литература. 404 с.
Носков Г.А., Рымкевич Т.А., 2010. Регуляция параметров годового цикла и ее роль в микроэволюционном процессе у птиц // Успехи современной биологии. Т. 130. № 4. С. 346–359.
Павлов А.В., Гравис Г.Ф., 2000. Вечная мерзлота и современный климат // Природа. № 4. C. 10–17.
Пасхальный С.П., 2002. Сроки прилета некоторых видов птиц в низовья Оби в 1970–2002 гг. // Многолетняя динамика численности птиц и млекопитающих в связи с глобальным изменением климата. Казань: ЗАО “Новое знание”. С. 151–156.
Пасхальный С.П., Головатин М.Г., 2007. Весенний пролет птиц в низовьях Оби // Научный вестник ЯНАО. Экосистемы Субарктики: структура, динамика, проблемы охраны. Вып. 6 (50). Ч. 2. С. 23–57.
Рыжановский В.Н., 2005. Сроки и продолжительность сезонных явлений годового цикла жизни воробьиных Субарктики на примере птиц Нижнего Приобья // Сибирский экологический журнал. № 3. С. 475–487.
Рыжановский В.Н., 2015. Экология рюма (Eremophila alpestris flava Gm.) и пуночки (Plectrophenax nivalis L.) в Субарктике и Арктике – сравнительный аспект // Сибирский экологический журнал. № 3. С. 379–387.
Соколов Л.В., 2006. Влияние глобального потепления климата на сроки миграции и гнездования воробьиных птиц в XX веке // Зоологический журнал. Т. 85. № 3. С. 317–341.
Соколов Л.В., 2010. Климат в жизни растений и животных. СПб.: Тесса. 343 с.
Соколов Л.В., 2017. Связь сроков прилета птиц в разных регионах России с температурным режимом // Труды Мензбировского общества. Т. 3. С. 157–180.
Соколов Л.В., Тропп Э.А, Морозов Ю.Г., Ефремов В.Д., 2001. Влияние температурного фактора на долговременные флуктуации сроков миграции, гнездования и расселения у воробьиных птиц // Доклады РАН. Т. 379. № 2. С. 282–285.
Соколов Л.В., Гордиенко Н.С., 2008. Повлияло ли современное потепление климата на сроки прилета птиц в Ильменский заповедник на Южном Урале // Экология. № 1. С. 58–64.
Соколов Л.В., Цвей А.Л., 2016. Механизмы контроля сроков весенней миграции у птиц // Зоологический журнал. Т. 95. № 11. С. 1362–1376.
Успенский С.М., 1969. Жизнь в высоких широтах. На примере птиц. М.: Мысль. 455 с.
Golovatin M.G., Paskhalny S.P., 2003. Timing of arrival and breeding of binds in the Nest of Western Siberia: relationship with the weather // Avian Ecology Behavior. V. 11. P. 47–69.
Haest B., Hüppop O., Bairlein F., (in press). Challenging a 15-year-old claim: The North Atlantic Oscillation index as a predictor of spring migration phenology of birds // Global Change Biology. P. 1–15. https://doi.org/10.1111/gcb.14023
Helm B., Schabl I., Gwinner E., 2009. Circannual basis of geographically distinct birds schedules // Journal of Experimental Biology. V. 212. P. 1259–1269.
Hüppop K, Hüppop, O., 2005. Atlas zur Vogelberingung auf Helgoland. T. 3. Veranderungen von Heim-und Verzugzeiten von 1960 bis 2001 // Vogelwarte. Bd. 43. S. 217–258.
Lehikoinen E., Sparks T.H., Zalakevicius M., 2004. Arrival and departure dates // The effect of climate change on birds. Advances in ecol. res. London: Academic Press. V. 35. P. 1–31.
Lehikoinen A., Linden A., Karlsson M., Andersson A., et al., 2019. Phenology of the avian spring migratory passage in Europe and North America: Asymmetric advancement in time and increase in duration // Ecological Indicators. V. 101. P. 985–991.
Moller A.R., Fiedler W., Berthold P., 2010. Effects of Climate Change on Birds. Oxford: University Pres. 318 p.
Peintinger M, Schuster S., 2005. Veränderungtn den Erstankünfte bei häufigen Zugvogelarten in Südwestdeutland // Vogelwarte. Bd. 43. S. 161–169.
Saino N., Bazzi G., Gatti E., Cecere J.G., et al., 2015. Polymorphism at the Clock gene predicts phenology of long-distance migration in birds // Molecular Ecology. V. 23. № 8. P. 1758–1773.
Sokolov L.V., Kosarev V.V., 2003. Relationship between timing of arrival of passerines to the Courish Spit and North Atlantic Oscillations index (NAOI) and precipitation in Africa // Proceedings Zoological Institute of Russian Academy of Science. V. 299. P. 141–154.
Sokolov L.V., Kosarev V.V., Fedoseeva N.V., et al., 2003. Relationship between autumn numbers of the Coal Tit Parus ater, air temperatures and North Atlantic Oscillation index // Avian Ecology and Behaviour. V. 11. P. 71–88.
Дополнительные материалы отсутствуют.
Инструменты
Зоологический журнал