

Зоологический журнал, 2020, T. 99, № 4, стр. 459-466
Влияние экологических факторов на обилие тарбагана (Marmota sibirica, Rodentia, Sciuridae) в юго-западном Забайкалье
Б. Б. Бадмаев *
Институт общей и экспериментальной биологии СО РАН
670047 Улан-Удэ, Россия
* E-mail: bbadm59@mail.ru
Поступила в редакцию 27.12.2018
После доработки 09.03.2019
Принята к публикации 22.08.2019
Аннотация
Тарбаган, или монгольский сурок (Marmota sibirica Radde 1862), – один из характерных представителей степных экосистем юго-западного Забайкалья. Исследование закономерностей, отражающих влияние экологических факторов на обилие тарбагана, позволит разработать модель экологической ниши этого вида и проводить эффективные природоохранные мероприятия по его сохранению. Влияние экологических факторов (топографических, эдафических, растительности, антропогенной нарушенности, совместного обитания с даурской пищухой и длиннохвостым сусликом) на обилие тарбагана изучали в Кяхтинском, Джидинском и Селенгинском административных районах Бурятии (юго-западное Забайкалье). В мае–августе 2015–2018 гг. методом линейных трансект протяженностью 1000 м с заложением четырех пробных площадок размерами 50 × 50 м на каждой из них проложено всего 95 трансект с 380 пробными площадками. Различия по обилию площадок оказались существенными в зависимости от высоты их поверхности над уровнем моря (критерий Пирсона χ2 = 23.422, p < 0.001, df = 2) и каменистости (χ2 = 15.219, p < 0.001, df = 1). Связь обилия с другими параметрами местообитаний (эдафическими, гетерогенности или гомогенности растительности и нарушенности) была не существенной. Присутствие жилых нор и колоний даурской пищухи в большинстве заселенных площадок тарбагана (χ2 = 21.772, p < 0.001, df = 1) указывает на сходные требования этих видов к определенным параметрам местообитаний (например, каменистости местоположений). На заселенных площадках тарбагана жилых нор и поселений длиннохвостого суслика не найдено (χ2 = 0.443, p = 0.506, df = 1), вероятно, семеноядные суслики и сурки имеют разные кормовые предпочтения.
Тарбаган, или монгольский сурок (Marmota sibirica Radde 1862), – один из характерных представителей степных экосистем юго-западного Забайкалья.
Наиболее общая черта рельефа данной территории – чередование обширных котловин и низких и средневысотных хребтов со сглаженными водоразделами, ориентированных в северо-восточном или восточно-северо-восточном направлениях. Равнинные участки межгорных понижений и долин крупных рек расположены на высотах 500–600 м в центральной и северной частях и 700–850 м над ур. м. в западной и восточной частях.
Климат резко-континентальный (Жуков, 1960). Преобладает северо-западный перенос воздушных масс. Среднегодовая температура воздуха отрицательная: от –0.5 (метеостанция Кяхта) до ‒3.0°С (метеостанция Окино-Ключи). Годовое количество осадков невелико и подвержено значительным колебаниям в зависимости от орографии и топографического положения: от 255 (метеостанция Новоселенгинск) до 500–600 мм на наветренных склонах хребта Малого Хамар-Дабана. Для температуры поверхности и температуры внутри почвы характерно увеличение запаздывания при проникновении холода и тепла с увеличением глубины. Промерзание почвы осенью до глубины 1 м (в черте г. Улан-Удэ) происходит в сентябре–второй половине октября, после чего оно медленно распространяется вниз и к марту достигает глубины 3–3.5 м (Жуков, 1960; Сницаренко, 1983). Оттаивание почвы в верхнем метровом слое начинается примерно в середине апреля и к концу апреля–в начале мая температура ее достигает –5°С. Годовые максимумы температуры почвы на глубине 1.6 м наблюдаются в самом конце лета и осенью, а на уровне 3.2 м – поздней осенью. В пределах юго-западного Забайкалья сплошные массивы зоны многолетней мерзлоты отсутствуют, но имеются участки многолетнемерзлых пород зоны островной мерзлоты и перехода от островной зоны к сплошной (Дугаров, Куликов, 1990).
На большей части рассматриваемой территории распространены экспозиционные лесостепные ландшафты, в которых северные склоны горных массивов облесены, а на южных склонах отсутствуют леса, и степные ландшафты, пригодные для обитания сурков.
Биологические и экологические особенности тарбагана изучались многими авторами и результаты исследований обобщались неоднократно (Огнев, 1947; Некипелов, 1950, 1978; Банников, 1954; Эрегдендагва, 1959; Бибиков, 1967; Швецов, 1978; Сунцов, 1982; Адьяа, 2000 и др.).
Это территориальный вид, роющий систему подземных убежищ, включая зимовальные норы. Тарбаганы объединяются в семейные группы, состоящие, в основном, из близкородственных особей, но существующий обмен особями между семьями приводит к наличию в некоторых семьях неродственных им иммигрантов (Берендяев, Кулькова, 1965; Бибиков, 1967; Сунцов, 1982), которые поддерживают генетическое разнообразие поселений.
Годичный жизненный цикл наземных беличьих (сурков и сусликов) складывается из двух частей: малоактивного состояния вплоть до впадения в глубокую спячку в холодное время года и активного состояния в теплое время года. Такая структура цикла обусловливает упорядоченность основных, тесно сопряженных с сезонной активностью экологических проявлений наземных беличьих: моноцикличного размножения после выхода из спячки, сезонной изменчивости в питании, последовательного в течение года расходования и накопления жировых резервов. Затем цикл замыкается уходом сурков в зимовальные норы в осенний период. Общая синхронизация этих процессов обеспечивается, по-видимому, зимней спячкой, контролируемой большей частью эндогенными факторами (Keyser, 1965, цит. по: Демин и др., 1988; Калабухов, 1985). Экзогенные факторы оказывают модулирующее влияние на выраженность эндогенных ритмов.
Большая часть времени жизни сурков проходит в норах, поэтому микроклиматические условия в них имеют большое значение в биологии этих животных. Тарбаган считается видом, занимающим промежуточное положение между байбаком, зимовальные норы которого располагаются в термически благоприятных условиях, и черношапочным сурком, зимовальные норы которого устраиваются в зоне многолетнемерзлых грунтов (Беловежец, 2015).
На территории юго-западного Забайкалья в отличие от юго-восточной ее части тарбаган не подвергался планомерному уничтожению противочумными организациями, современная его численность позволяет проводить ежегодный промысел.
Состояние популяций тарбагана на территории Тувы и Забайкалья в последнее время оценивалось методом экспресс-обследования на обширной территории с выделением антропогенного фактора как влияющего наиболее отрицательно на его численность по ареалу (Рожнов и др., 2005).
В современных условиях получают интенсивное развитие направления, связанные с ландшафтной экологией млекопитающих. Важным является изучение обилия и качества местообитаний животных в их взаимной связи и отношении с возможно большим количеством других параметров окружающей среды (Пузаченко, 2004). В конечном итоге это позволит разрабатывать модели экологической ниши исследуемых видов, используя быстро развиваемые методы анализа полученных результатов.
Цель работы – исследование закономерностей, отражающих влияние экологических факторов на обилие тарбагана в юго-западном Забайкалье и позволяющих повышать эффективность природоохранных мероприятий по сохранению вида на данной территории.
МАТЕРИАЛ И МЕТОДИКА
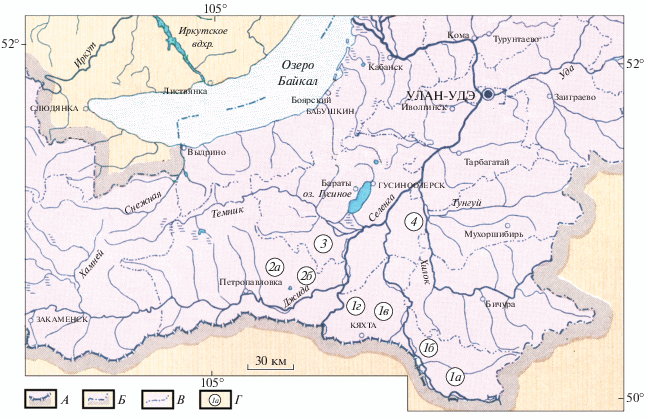
Работа выполнена в Кяхтинском (местообитание “Кяхта”), Джидинском (местообитания “Дапхуры” и “Дырестуй”) и Селенгинском (местообитания “Селендума” и “Зурган-Дэбэ”) административных районах Республики Бурятии (юго-западное Забайкалье) в мае–августе 2015–2018 гг. (рис. 1).
Рис. 1.
Место проведения исследований в юго-западном Забайкалье: А – государственная граница с Монголией, Б – граница с Забайкальским краем, В – границы административных районов Бурятии, Г – изученные местообитания тарбагана: 1а, 1б, 1в, 1г – в Кяхтинском р-не, 2а – Дапхуры и 2б – Дырестуй в Джидинском р-не, 3 – Селендума и 4 – Зурган-Дэбэ в Селенгинском р-не Бурятии.
В местообитаниях тарбагана прокладывали линейные трансекты протяженностью 1000 м (Allaine et al., 1994; Ricankova et al., 2014). Попеременно в обе стороны от линии каждой трансекты на расстоянии 200 м друг от друга заложены пробные площадки размерами 50 × 50 м. В начальной точке трансекты и угловых точках площадок фиксировали географические координаты и высоту поверхности над уровнем моря с помощью навигатора Garmin-e-trex. В середине площадки определяли географические координаты, высоту над уровнем моря, средний уклон ее в градусах (°) и экспозицию склона. На площадке проводили сбор других топографических параметров местообитания – неровность местообитания, которую разделяли на: (1) низкую, <10° без скальных выступов, (2) среднюю, >10° без скальных выступов и (3) высокую, >10° со скальными выступами, рельеф равнинный и горный, с положениями площадки на склоне: (1) доля в нижней трети холма, (2) доля в средней части и (3) доля в верхней трети холма. На площадке определяли тип почвенного покрова и характер подпочвенных слоев: (1) водопроницаемые и (2) водонепроницаемые. На карте-схеме площадок фиксировали степень каменистости, расположение отдельных деревьев, кустарников и состав растительных сообществ, что позволяло сделать заключение о гомогенности (однородности) или гетерогенности (неоднородности) структуры растительности, антропогенной нарушенности и землепользования (дороги, тропы и выпас животных, следы посещения автомашинами, наличие колышков от петель или капканов и др.). Также фиксировали норы тарбагана, длиннохвостого суслика (Urocitellus undulatus) и колоний даурской пищухи (Ochotona dauurica).
Обилие сурков на площадках подразделяли по балльной шкале: а) отсутствие (нет), б) наличие (да), в) среднее и г) высокое. Отсутствие или наличие хотя бы одной жилой норы позволяло отнести обилие к баллам а) и б) соответственно. Среднее обилие сурков (в) означало не менее трех жилых нор с сурчинами, а высокое обилие (г) – более трех жилых нор, сурчины с выраженными следами жизнедеятельности и диаметром до 2–3 м, тропы, соединяющие норы, ямки со свежими экскрементами сурков (Дмитриев, 2006).
Всего проложены 95 трансект с заложением 380 пробных площадок.
Статистическая обработка результатов проводилась с использованием программ Minitab 13 for Windows и STATISTICA-6.0. При изготовлении графического материала использовали Excel 7.0. Различие площадок по обилию оценивали критерием Пирсона χ2.
РЕЗУЛЬТАТЫ
Обилие тарбагана на пробных площадках в их местообитаниях распределилось следующим образом (табл. 1). Наибольшим обилием отличались группировки сурков в местообитании “Дапхуры” Джидинского р-на. Высокое обилие тарбагана было в местообитании “Селендума” Селенгинского р-на, а низкое – в местообитании “Зурган-Дэбэ” этого же района.
Таблица 1.
Обилие тарбагана в местообитаниях в юго-западном Забайкалье
| Обилие | Местообитания | ||||
|---|---|---|---|---|---|
| Кяхта (N = 80) | Дапхуры (N = 100) | Дырестуй (N = 80) | Селендума (N = 80) | Зурган-Дэбэ (N = 40) | |
| Высокое | 10 (12.5%) | 26 (26.0%) | 5 (6.25%) | 7 (8.75%) | 3 (7.5%) |
| Среднее | 8 (10.0%) | 16 (16.0%) | 10 (12.5%) | 6 (7.5%) | 1 (2.5%) |
| Наличие | 11 (13.75%) | 25 (25.0%) | 21 (26.25%) | 29 (36.25%) | 9 (22.5%) |
| Отсутствие | 51 (63.75%) | 33 (33.0%) | 44 (55.0%) | 38 (47.5%) | 27 (67.5%) |
Поскольку обилие тарбагана определяется качеством их местообитаний, различные свойства параметров их местообитаний сравнивали на площадках, заселенных и незаселенных сурками.
Топографические и эдафические параметры местообитаний
Высота над уровнем моря. Все исследованные местообитания располагались на разной высоте над уровнем моря (рис. 2). Самые высокие площадки были расположены в Джидинском р-не, а самые низкие – в Селенгинском р-не. Распределение заселенных и незаселенных площадок в зависимости от высоты над уровнем моря и от крутизны склонов во всех местообитаниях показано в табл. 2. Различие по обилию заселенных и незаселенных площадок в зависимости от уровня их высот оказалось существенным (χ2 = 23.422, p < 0.001, df = 2). В горных ландшафтах тарбаганы обитали на большей высоте.
Рис. 2.
Уровни высот поверхностей площадок в местообитаниях (1–5) тарбагана в юго-западном Забайкалье: 1 – Кяхта, 2 – Дапхуры, 3 – Дырестуй, 4 – Селендума, 5 – Зурган-Дэбэ. Размах обозначает разницу между максимумом и минимумом.
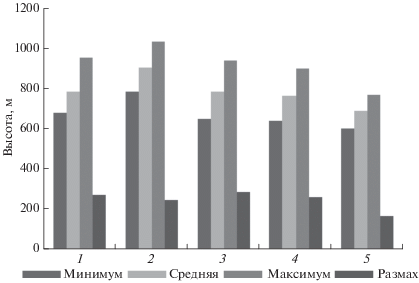
Таблица 2.
Параметры местообитаний и обилие сурка
| Параметры | Обилие на площадке | Критерий Пирсона, χ2 |
df | р | ||
|---|---|---|---|---|---|---|
| Название | Значение | заселенной | незаселенной | |||
| 1 | 2 | 3 | 4 | 5 | 6 | 7 |
| Высота | 600–749 | 48 | 68 | 23.422 | 2 | <0.001 |
| поверхности | 750–899 | 90 | 110 | |||
| над ур. м., м | 900–1048 | 49 | 15 | |||
| Крутизна | 1–7 | 100 | 108 | 0.246 | 2 | 0.884 |
| склона, ° | 8–14 | 67 | 66 | |||
| 15–28 | 20 | 19 | ||||
| Экспозиция | Северная | 21 | 13 | 11.446 | 7 | 0.120 |
| склона* | Северо-восточная | 32 | 20 | |||
| Восточная | 22 | 33 | ||||
| Юго-восточная | 33 | 26 | ||||
| Южная | 26 | 31 | ||||
| Юго-западная | 23 | 28 | ||||
| Западная | 15 | 21 | ||||
| Северо-западная | 12 | 20 | ||||
| Неровность | Высокая | 28 | 28 | 0.031 | 2 | 0.985 |
| поверхности | Средняя | 54 | 55 | |||
| Низкая | 105 | 110 | ||||
| Часть | Верхняя | 34 | 43 | 5.345 | 3 | 0.148 |
| склона | Средняя | 80 | 61 | |||
| Нижняя | 68 | 81 | ||||
| Равнина | 5 | 8 | ||||
| Каменистость | Да | 90 | 108 | 15.219 | 1 | <0.001 |
| Нет | 119 | 63 | ||||
| Типы почв | Каменистые | 35 | 24 | 7.987 | 4 | 0.092 |
| Щебнистые | 17 | 28 | ||||
| Черноземные | 66 | 80 | ||||
| Темно-каштановые | 7 | 16 | ||||
| Каштановые | 46 | 61 | ||||
| Растительность | Гомогенная | 139 | 122 | 1.023 | 1 | 0.312 |
| Гетерогенная | 70 | 49 | ||||
| Нарушенность, | Высокая | 7 | 13 | 2.137 | 2 | 0.344 |
| антропогенная | Средняя | 19 | 23 | |||
| Низкая | 161 | 157 | ||||
| Даурская | Присутствие | 138 | 72 | 21.772 | 1 | <0.001 |
| пищуха | Отсутствие | 71 | 99 | |||
| Длиннохвостый | Присутствие | 69 | 51 | 0.443 | 1 | 0.506 |
| суслик | Отсутствие | 140 | 120 | |||
Крутизна склонов не влияла существенно на обилие сурков. Крутизна склонов площадок в Кяхтинском р-не была от 2° до 25°, в среднем 10.4°. В местообитаниях Джидинского р-на Дапхуры – от 1° до 26°, в среднем 8.0° и Дырестуй от 2° до 21°, в среднем 7.4°. В Селенгинском р-не в местообитании Селендума крутизна склонов была от 2° до 19°, в среднем 7.0°, а в местообитании Зурган-Дэбэ – от 3° до 20°, в среднем 7.1°.
Общее число всех площадок градуально снижалось с увеличением крутизны склона (табл. 2). Тем не менее, между заселенными и незаселенными площадками в зависимости от крутизны склона различий по обилию не выявлено (χ2 = = 0.246, p = 0.884, df = 2).
Экспозиция склона. Различие в распределении площадок в зависимости от экспозиции раздельно по местообитаниям были значительными. В совокупности по всем местообитаниям эти различия несколько выравниваются, однако заметных экспозиционных различий между обитаемыми и необитаемыми площадками не выявлено (табл. 2, χ2 = 11.446, p = 0.120, df = 7).
Неровность поверхности площадок и положение их на склоне не оказывает влияния на их заселенность (табл. 2, χ2 = 0.031, p = 0.985, df = 2).
В дополнение к изучению влияния неровности поверхности в горных условиях площадки сравнивали по положению на склоне, подразделив склон на три части: верхнюю треть, средний уровень и нижнюю треть.
Общее количество всех площадок постепенно снижается от нижней к верхней части склона. Однако заселенность и незаселенность этих площадок в зависимости от того, на какой части склона они находятся, не различались заметно (табл. 2, χ2 = 5.345, p = 0.148, df = 3).
Каменистость местообитаний оказывает существенное влияние на заселенность их сурками (табл. 2, χ2 = 15.219, p < 0.001, df = 1).
Типы почв в местообитаниях. На площадках всех местообитаний тарбагана были выделены 5 типов почв: каменистые, щебнистые, черноземные, темно-каштановые и каштановые. Среди них преобладали черноземные почвы, которым незначительно уступали по частоте каштановые почвы.
Распределение заселенных и незаселенных площадок в зависимости от наличия этих типов почв представлено в табл. 2: на заселенных площадках выявлено сравнительно меньшее разнообразие типов почв, хотя значительного различия между площадками не наблюдалось (χ2 = 7.987, p = 0.092, df = 4).
Гетерогенность и гомогенность растительности
Большая часть фитоценозов исследованных площадок (97% и выше) была представлена травянистой растительностью, лишь оставшаяся часть включала кустарниковую и древесную растительность. Высота травянистой растительности в подавляющем большинстве случаев относилась к категории низкотравья.
Как следует из данных табл. 2, площадки с гетерогенной и гомогенной растительностью не различались по показателю заселенности и не заселенности (χ2 = 1.023, p = 0.312, df = 1).
Антропогенная нарушенность
В исследованных местообитаниях в результате в целом неумеренной пастьбы домашнего скота нарушенными в основном оказались травостой и растительный покров. Менее нарушен был почвенный покров, хотя из-за постоянной езды на автомобилях или других транспортных средствах на поверхности грунта возникают колеи. Кроме того, постоянное или периодическое вытаптывание грунта крупным рогатым скотом привело к обнажению почвенного покрова.
Распределение заселенных и незаселенных тарбаганом площадок в зависимости от нарушенности местообитаний представлено в табл. 2. Как следует из этих данных, хотя количество площадок в зависимости от степени нарушенности местообитаний снижается, заметных различий между заселенными и незаселенными площадками по этому параметру не обнаруживается (χ2 = 2.137, p = 0.344, df = 2).
Отношения с другими видами млекопитающих: даурской пищухой и длиннохвостым сусликом
Из травоядных млекопитающих, конкурирующих с тарбаганом, в их местообитаниях в юго-западном Забайкалье наиболее часто встречаются даурская пищуха и длиннохвостый суслик. Исходя из этого, были рассмотрены случаи распределения этих видов на заселенных и незаселенных тарбаганами площадках.
Как показывают данные табл. 2, распределение даурских пищух не является случайным – между обитанием сурков и пищух имеется существенная связь (χ2 = 21.772, p < 0.001, df = 1).
Длиннохвостый суслик визуально и акустически часто обнаруживается на поселениях тарбагана, в то же время норы сусликов в отличие от обитаемых нор даурской пищухи труднее обнаружить среди травянистой растительности. Как следует из данных табл. 2, суслики в отличие от пищух не выявляют заметной связи с сурками (χ2 = 0.443, p = 0.506, df = 1).
ОБСУЖДЕНИЕ
Исследованы параметры местообитаний тарбагана в юго-западном Забайкалье. Выявлена существенная связь обилия тарбагана с высотой, на которой расположена поверхность площадки над уровнем моря и каменистостью этих площадок. Джидинский и Кяхтинский районы по отношению к Селенгинскому расположены выше по р. Селенге, которая протекает по территории юго-западного Забайкалья субмеридионально в северном направлении. Вероятно, поэтому площадки, которые были заселены, расположены на большую высоту над уровнем моря в этой части местообитаний тарбагана. Верхние уровни горных склонов местообитаний тарбагана в юго-западном Забайкалье лучше обеспечены влагой в условиях сухостепных и степных экосистем.
Высокая степень каменистости снижает качество местообитаний сурка, поскольку обеспечивает высокую теплообеспеченность и низкую влагообеспеченность. В летний период в дневное время камни сильно нагреваются и после захода солнца постепенно излучают тепло, верхний слой почвы получает дополнительное количество тепла, что может приводить к большей сухости таких местообитаний. При увеличении каменистости субстрата увеличивается теплоемкость этого слоя (Куликов и др., 1997). Каменистый субстрат в условиях Западного Забайкалья усиливает действие основного климатического фактора – температурного, что приводит к распространению криоксерофильной растительности (Рещиков, 1961).
Из травоядных млекопитающих норы и обитаемые колонии даурской пищухи довольно часто встречаются на заселенных площадках тарбагана. Это свидетельствует о том, что у этих видов экологические требования к среде обитания имеют много общего. Одним из факторов, одинаково отрицательно влияющих на распределение даурских пищух и тарбагана, оказалась каменистость местоположений. Это было хорошо видно на примере местообитания тарбагана в Дырестуйских горах Джидинского р-на, степные участки которых отличаются большой степенью каменистости. Здесь из 80 площадок 53 площадки, или 66.2% оказались каменистыми и в то же время практически незаселенными пищухами, т.к. лишь на одной площадке нашли их обитаемые норы. В местообитании Дырестуй и на других площадках, с большей степенью каменистости обилие сурков в сравнении с этим показателем в других местообитаниях и на других площадках, было меньшим (табл. 1).
Длиннохвостый суслик часто посещает участки обитания тарбагана, однако его жилые норы, по-видимому, устраиваются вне этих участков. С одной стороны, при индивидуальных поведенческих взаимодействиях тарбаган агрессивно преследует сусликов, оказавшихся на их сурчинах. Суслики обычно обживают брошенные норы сурков. С другой стороны, высокое обилие зерноядных сусликов и их норы наблюдали в Кяхтинском р-не вдоль полей с сельскохозяйственными злаками, которые не были заселены сурками.
Обилие животных определяется в основном качеством местообитаний: доступностью полноценной пищи, меньшим прессом хищников, меньшим уровнем межвидовых конкурентных отношений (Batzli et al., 1999; Хански, 2015). От качества местообитаний зависят выбор определенным видом определенных местообитаний, демографические показатели популяции вида, уровень рождаемости и смертности, иммиграции и эмиграции (Хански, 2015).
В наших исследованиях среди условий окружающей среды большое внимание уделяется топографическим параметрам, связанным с особенностями рельефа конкретных местоположений. Влияние этих местоположений на популяционные характеристики изучаемых видов может быть опосредовано через более близкое влияние на формирование определенных типов почвенного и растительного покрова. Поскольку цель исследований состоит в выявлении закономерностей выбора местообитания конкретными видами для построения в перспективе моделей экологических ниш, важно выявить реальные существующие отношения между ними и различными компонентами природной среды. В этом направлении наиболее значимыми оказались факторы окружающей среды, например, высота над уровнем моря, определяющая общее распределение тепла и влаги, и каменистость определенных местоположений.
ЗАКЛЮЧЕНИЕ
В проведенном исследовании получены данные по обилию тарбагана и параметрам его местообитаний в Кяхтинском, Джидинском (местообитания Дапхуры и Дырестуй) и Селенгинском (местообитания Селендума и Зурган-Дэбэ) районах Бурятии (юго-западное Забайкалье), что позволило оценить связь между этими экологическими показателями.
Высота местообитаний, каменистость поверхности, конкурентный травоядный вид даурская пищуха выделены нами как существенные факторы, связанные с обилием тарбагана.
Список литературы
Банников А.Г., 1954. Млекопитающие Монгольской Народной Республики. М.: Изд-во АН СССР. 669 с.
Беловежец К.И., 2015. Теория температурного режима спячки сурков // Прошлое, настоящее и будущее сурков Евразии. Сб. науч. тр. М.: АБФ Медиа. С. 42–51.
Берендяев С.А., Кулькова Н.А., 1965. О внутривидовых отношениях серых сурков // Зоологический журнал. Т. 44. Вып. 4. С. 110–116.
Бибиков Д.И., 1967. Горные сурки Средней Азии и Казахстана. М.: Наука. 200 с.
Демин Н.Н., Шортанова Т.Х., Эмирбеков Э.З., 1988. Нейрохимия зимней спячки млекопитающих. Л.: Наука. 136 с.
Дмитриев П.П., 2006. Млекопитающие в степных экосистемах Внутренней Азии // Труды совместной российско-монгольской комплексной биологической экспедиции. Т. 48. М. 214 c.
Дугаров В.И., Куликов А.И., 1990. Агрофизические свойства мерзлотных почв. Новосибирск: Наука. 255 с.
Жуков В.М., 1960. Климат Бурятской АССР. Улан-Удэ: Бурятское книжное изд-во. 188 с.
Калабухов Н.И., 1985. Спячка млекопитающих. М.: Наука. 260 с.
Куликов А.И., Дугаров В.И., Корсунов В.М., 1997. Мерзлотные почвы: экология, теплоэнергетика и прогноз продуктивности. Улан-Удэ. 312 с.
Некипелов В.Н., 1950. Очерк биологии тарбагана // Изв. Иркутск. науч.-исслед. противочумн. ин-та Сибири и Дальнего Востока. Т. 8. С. 27 - 44.
Некипелов В.Н., 1978. Тарбаган. Юго-восточное Забайкалье // Сурки. Распространение и экология. М.: Наука. С. 164–177.
Огнев С.И., 1947. Звери СССР и прилежащих стран. Т. 5. Грызуны. М.-Л.: Изд-во АН СССР. 809 с.
Пузаченко Ю.Г., 2004. Математические методы в экологических и географических исследованиях. М.: Изд-во ACADEMIA. 408 с.
Сницаренко Н.И., 1983. Термический режим // Климат Улан-Удэ. Л.: Гидрометеоиздат. С. 59–82.
Рещиков М.А., 1961. Степи Западного Забайкалья. М.: Изд-во АН СССР. 174 с.
Рожнов В.В., Поярков А.Д., Брандлер О.В., Громов В.С., Бадмаев Б.Б., 2005. Состояние монгольского сурка (тарбагана) (Marmota sibirica Radde, 1862) на территории России в начале XXI в. // Бюл. МОИП. Отд. биол. Т. 110. Вып. 4. С. 21–32.
Сунцов В.В., 1982. Экология тарбагана в Туве. Автореф. дис. … канд. биол. наук. М. 163 с.
Хански И., 2015. Ускользающий мир: экологические последствия утраты местообитаний. 2-е изд. М.: Товарищество научных изданий КМК. 344 с.
Швецов Ю.Г., 1978. Тарбаган. Юго-западное Забайкалье // Сурки. Распространение и экология. М.: Наука. С. 154–164.
Эрегдендагва Д., 1959. Тарбаган Монгольской Народной Республики и его хозяйственное использование. Автореф. дис. … канд. биол. наук. Иркутск. 27 с.
Allaine D., Rodrigue I., Le Berre M., Ramousse R., 1994. Habitat preferences of alpine marmots, Marmota marmota // Canadian Journal of Zoology. V. 72. P. 2193–2198.
Batzli G.O., Harper S.J., Lin Y-T.K., Desy E.A., 1999. Experimental analyses of population dynamics: scaling up to the landscape // Landscape ecology of small mammals. New-York: Springer. P. 107–127.
Keyser Ch., 1965. Hibernation // Physiological mammalogy. New York, London: Acad. Press. V. 2. P. 179–211.
Ricankova V.P., Riegert J., Semancikova E., Hais M., Cejkova A., Prach K., 2014. Habitat preferences in grey marmots (Marmota baibacina) // Acta Theriol. V. 59. P. 317– 324.
Адъяа Я., 2000. Монгол тарвага: биологи, экологи, хамгаалал, аж ахуйн холбогдол. Улаанбаатар: Аdmon. 199 х. (на монг.).
Дополнительные материалы отсутствуют.
Инструменты
Зоологический журнал