

Зоологический журнал, 2020, T. 99, № 4, стр. 373-393
Род Heterocope Sars 1863 (Copepoda, Calanoida) в России: морфология и распространение
Н. Г. Шевелева a, *, В. Н. Подшивалина b, c, **, И. М. Мирабдуллаев d, ***, Е. С. Чертопруд e, f, ****, В. И. Лазарева g, *****, Н. Г. Баянов h, ******
a Лимнологический институт СО РАН
664033 Иркутск, Россия
b Государственный природный заповедник “Присурский”
428034 Чебоксары, Россия
c Чувашский государственный университет им. И.Н. Ульянова
428015 Чебоксары, Россия
d Ташкентский государственный аграрный университет
100140 Ташкент, Узбекистан
e Московский государственный университет им. М.В. Ломоносова
119991 Москва, Россия
f Институт проблем экологии и эволюции им. А.Н. Северцова РАН
119071 Москва, Россия
g Институт биологии внутренних вод им. И.Д. Папанина РАН
152742 пос. Борок, Россия
h Государственный природный заповедник “Керженский”
603001 Нижний Новгород, Россия
* E-mail: shevn@lin.irk.ru
** E-mail: verde@mail.ru
*** E-mail: imirabdullayev@umail.uz
**** E-mail: horsax@yandex.ru
***** E-mail: laz@ibiw.yaroslavl.ru
****** E-mail: bayanovng@mail.ru
Поступила в редакцию 28.04.2019
После доработки 29.08.2019
Принята к публикации 06.11.2019
Аннотация
Представлено сравнительное описание ранее не установленных морфологических особенностей видов рода Heterocope Sars 1863, обитающих на территории России. Благодаря возможностям СЭМ подробно описаны генитальная пластинка, строение и вооружение Р5 самки и самца Heterocope soldatovi Rylov 1922, генитальная пластинка Heterocope saliens (Lilljeborg 1863). Впервые приведены сведения о ротовых придатках исследуемых видов. Выполнен сравнительный анализ морфологических и морфометрических отличий самок и самцов разных видов Heterocope и составлена общая определительная таблица. На основании современных данных уточнены границы ареалов видов.
Род Heterocope на территории России представлен 5 видами: Heterocope appendiculata Sars 1863, H. borealis (Fischer 1851), H. caspia Sars 1897, H. saliens (Lilljeborg 1862) и H. soldatovi Rylov 1922. Все они являются палеарктами, но ареал некоторых из них крайне узок. Так, H. soldatovi обитает только в бассейне р. Амур (Боруцкий и др., 1991). H. caspia – в Каспийском, Азовском и Черном морях и их бассейнах, в том числе в водохранилищах Средней и Нижней Волги, Камы, низовьях рек-притоков Азовского и Черного морей (Боруцкий и др., 1991). Н. appendiculata, H. borealis и H. saliens имеют более широкий ареал, но предпочитают водоемы северной части Палеарктики. Тем не менее, видам рода свойственно естественное стремление к расселению (Ривьер, Дзюбан,1978), что обусловливает необходимость уточнения современных границ их ареалов. Благодаря современным методам исследований выявляются неизвестные ранее особенности строения. Целью настоящей работы был сравнительный морфологический анализ представителей рода Heterocope, населяющих территорию России.
Материалом для изучения морфологии рачков послужили пробы, собранные из водоемов разных точек России в 2016–2018 гг. Ракообразных фиксировали спиртом (96%) или формальдегидом (40%). В работе использовали оптический микроскоп OlympusCX 41 и сканирующий электронный микроскоп Philips525M на базе Приборного центра коллективного пользования физико-химического ультрамикроанализа Лимнологического института СО РАН. Для изучения морфологии из проб выбирали половозрелых самок и самцов. Размер особи измеряли от конца рострума до конца каудальных ветвей. Описание приведены по самкам. Определение ракообразных проводили по морфологическим признакам, используя известные руководства (Боруцкий, 1960; Боруцкий и др., 1991; Kiefer, Fryer, 1978; Dussart, 1967). Особенности распространения охарактеризованы на основе собственных неопубликованных и литературных данных. Сплошной ареал вида на карте обозначен штриховкой. Крайние точки находок вида, упомянутые в тексте, отмечены значками (круг или квадрат).
Отряд Calanoida Sars 1903
Нeterocope appendiculata Sars 1863
Материал. Особи разных полов в массовом количестве из проб зоопланктона Богучанского водохранилища (приплотинная часть: 52°46′ с.ш., 103°41′ в.д) c глубин 5–10 м.
Описание. Самка. Цефалоторакс длинный узкий, его длина не более чем в 3.3 раза больше максимальной ширины, торакальные сомиты одинакового строения.
Генитальный сомит самки прямоугольной формы, его длина больше ширины (табл. 1). Генитальная пластинка несет лентовидные, расщепленные на концах придатки, доходящие до конца заднего края генитального сомита. Латерально от лентовидных придатков расположено по одному раздвоенному на конце шиповидному зубцевидному придатку.
Таблица 1.
Морфологические характеристики видов рода Heterocope фауны России
| Показатель | H. appendiculata | H. soldatovi | H. borеalis | H. saliens | H. caspia |
|---|---|---|---|---|---|
| Самка | |||||
| Длина тела, мм | 2.00–2.63 | 1.55–1.63 | 2.37–3.13 | 2.15–2.18 | 1.50–1.73 |
| Ceph. (L : W) | 2.7–3.3 | Нет данных | 2.3–2.8 | 2.5–2.8 | 3.0–3.2 |
| Генитальный сегмент (L : W) | 1.4–1.7 | 1.2–1.3 | 1.17–1.30 | 1.1–1.4 | 1.16–1.29 |
| Придатки генитальной пластинки | Лентовидные придатки в центре и по 1 двухвершинному латеральному зубцу | 2 латеральных зубца с каждой стороны | 2 закругленных выроста | Придатки отсутствуют | 2 латеральных зубца с каждой стороны |
| Каудальные ветви (L : W) | 1.6–1.7 | 1.4–1.6 | 1.1–1.2 | 1.3–1.6 | 2.9–3.3 |
| Сетулы на внутренней поверхности каудальных ветвей | Имеются на дистальной 1/3 поверхности каудальных ветвей | Имеются на дистальной 1/2–1/3 поверхности каудальных ветвей | Имеются на дистальной 1/2 поверхности каудальных ветвей | Имеются на дистальной 1/2 поверхности каудальных ветвей | Отсутствуют |
| Внешняя щетинка каудальных ветвей | Отсутствует | Отсутствует | Имеется | Имеется | Отсутствует |
| s1:s2 | 0.50–0.70 | Нет данных | 0.80–0.82 | 0.82–0.86 | 0.84–0.94 |
| Внутренние зубцы Р5exp2 | 4 одновершинных зубцевидных выроста | 4 одновершинных зубцевидных выроста | 4 зубцевидных выроста, слабо двувершинных | 4 зубцевидных выроста, ясно двувершинных | 2–3 зубцевидных выроста |
| Самец | |||||
| Р5 правая, exp2 | 1 шип | 3 шипа | 3 шипа | 2 шипа | Нет шипов |
Каудальные ветви самки относительно длинные (длина в 1.6–1.7 раза больше ширины), опушены на 1/3 длины (от дистального конца) редкими тонкими волосками, несут 4 апикальные щетинки. Внутренняя щетинка немного смещена на дорсальную сторону, прозрачная, очень тонкая. Три апикальные щетинки мощные, две крайние почти равной длины, самая длинная – средняя в 1.5–1.7 раз длиннее крайних апикальных.
Антеннулы самки длинные, достигают конца генитального сомита.
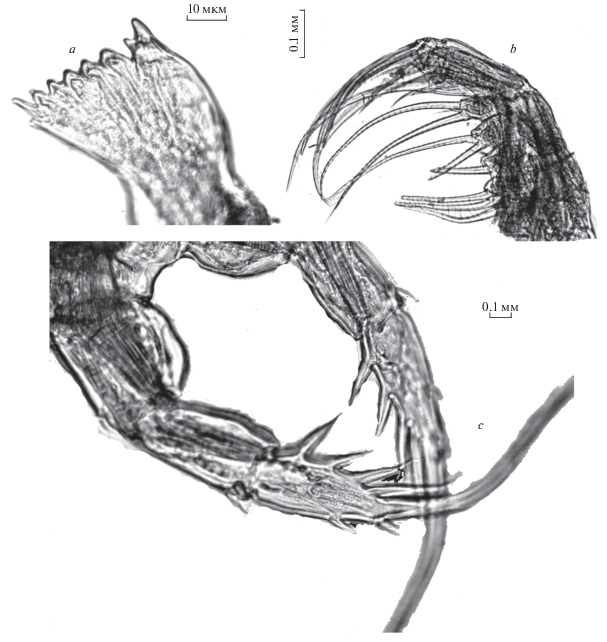
Вентральный зубец мандибулы одновершинный, слегка изогнутый, заостренный, отделен от остальных зубцов глубокой диастемой. Центральные зубцы округлые на конце (рис. 1а, 1b), двухвершинные, с широким основанием, дорсальные – острые одновершинные, длиннее центральных, с узким основанием. У основания центральных зубцов продольные, слегка изогнутые ряды шипиков. Подобные мандибулы типичны для рачков, питающихся животными с непрочным хитином (Монаков, 1998).
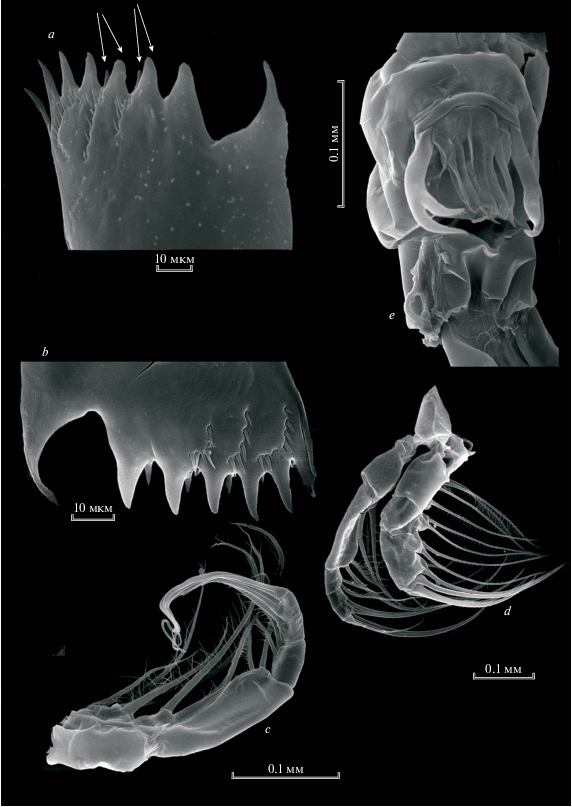
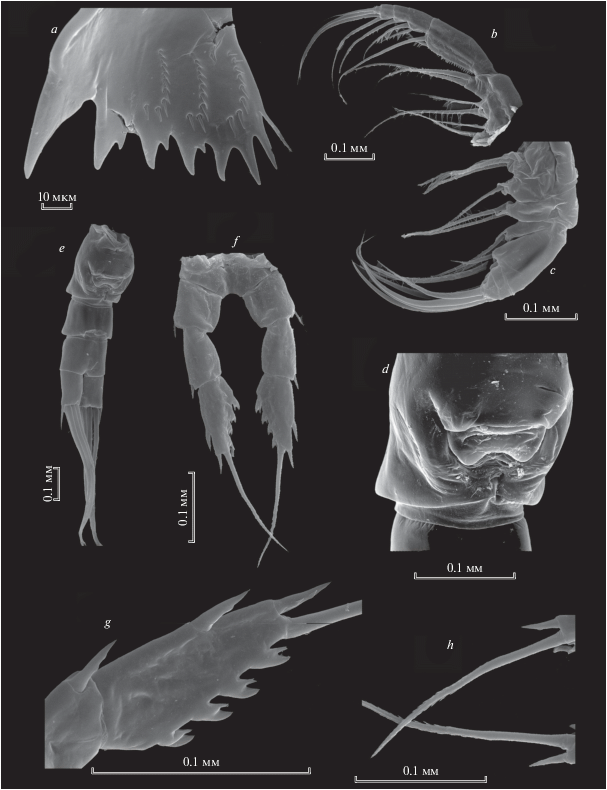
Рис. 1.
Heterocope appendiculata Sars 1863: a, b – жевательная пластинка мандибулы; с –максиллипед; d – максиллипед и максилла; e – генитальный сегмент.
Максилла и максиллипед несут длинные опушенные щетинки (рис. 1с, 1d), что вероятно указывает на способность к фильтрации.
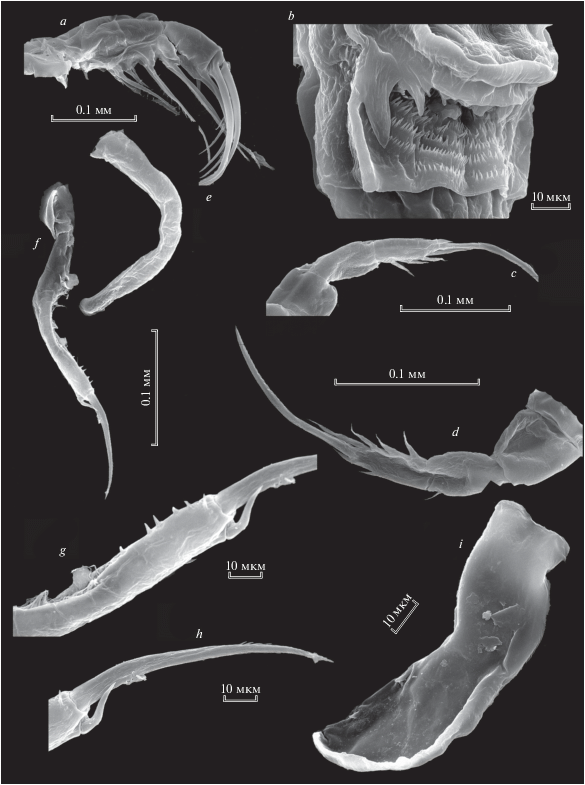
Р5 самки состоит из базиподита и двух члеников экзоподита (рис. 2а, 2b). На первом членике экзоподита с внешней стороны находится 1 шип, на втором членике 2 шипа с внешней стороны и 4 зубцевидных выроста с внутренней стороны. Второй членик экзоподита оканчивется длинным шипом, зазубренным с внутренней стороны (рис. 2c).
Рис. 2.
Heterocope appendiculata Sars 1863: a – Р5 самки (стрелками показаны шипообразные выросты на ехр1 и ехр2); b – Р5exp2 самки; c – апикальный шип Р5exp2 самки; d – левая Р5 самца; e – exp2 левой Р5 самца (стрелками показаны шипики); f – правая Р5 самца; g – exp2 левой Р5 самца, внутренняя сторона.
Самец. Левая Р5 самца состоит из базиподита, серповидного эндоподита (рис. 2d) и двухчленистого экзоподита (рис. 2е, 2g). P5exp2 относительно длинный, с утолщением в первой половине длины, внутренний край несет волоски проксимально и продольный ряд шипиков дистально (рис. 2e, 2g). С внешней стороны второго членика экзоподита находится 3 коротких крепких шипа, членик заканчивается одним апикальным изогнутым шипом (рис. 2e, 2g).
Правая Р5 самца более короткая, изогнутая, 1-членистая (рис. 2f), утолщенная в средней части, на внутренней поверхности дистальной части имеется 2 маленьких шипа на апикальном конце – кривой шип (рис. 2 f).
Распространение. Согласно данным Боруцкого и соавторов (1991), распространение H. appendiculata приурочено к северной части и северу средней части Европейской Палеарктики и Западной Сибири, а также к Восточной Сибири и Дальнему Востоку (рис. 3). Еще в середине 20 в. отмечено распространение этого северного вида на юг, вниз по Волге до Куйбышевского водохранилища (Мордухай-Болтовской, Дзюбан, 1976). В настоящее время вид встречается и ниже по течению, в Саратовском (Попов, 2011) и Волгоградском (Лазарева и др., 2018) водохранилищах. Вид также найден в Шекснинском (Ривьер, Литвинов, 2006) и Уводьском (Семенова и др., 2005) водохранилищах, на прилежащих территориях параллельно течению Волги (не имеющих сообщения с Волгой) до широты г. Казань в малых озерах (Низменное лесное Заволжье (Баянов, 2002; Подшивалина, 2006), Приволжье, г. Казань (Горшкова, 2012), западное побережье Рыбинского водохранилища (Лазарева, 2005) (рис. 3). Вероятно, сплошную часть ареала в Европе можно охарактеризовать как ограниченную арктическими и бореальными широтами, в Азии она включает и умеренные широты (до 51° с.ш. – Байкал (Малое море), р. Зея, (собственные данные Н. Г. Шевелева.). Особо выделяются точки нахождения вида в Киевском водохранилище на Днепре (Семенова, 2009) и в высокогорных озерах Кабардино-Балкарии (Пежева и др., 2016) (рис. 3).

Heterocope borealis (Fischer 1851)
Материал. Особи обоих полов из проб оз. Ильчир (Восточные Саяны, 51°50′ с.ш, 101°00′ в.д июль 2018 г.).
Описание. Самка. Наиболее крупный вид из этого семейства. Цефалоторакс широкий (табл. 1), с наибольшей шириной в средней части тела, торакальные сомиты одинакового строения.
Генитальный сомит самки почти квадратный, его длина немногим больше ширины (табл. 1). Генитальная пластинка с двумя закругленными на заднем крае выростами (рис. 4d).
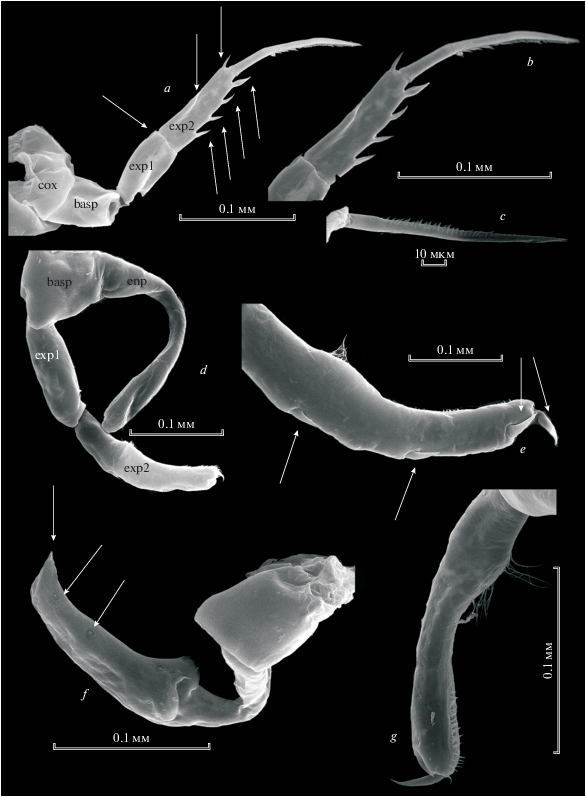
Рис. 4.
Heterocope borealis (Fisher 1851), самка: a – режущий край мандибулярной пластинки; b – максиллипед; c – максилла; d, e – генитальная пластинка; f – Р5; g – Р5exp2; h – апикальный шип Р5exp2.

Антеннула достигает заднего края цефалоторакса.
Режущий край мандибулы (рис. 4а) с вентральным тонким и длинным зубцом, с хорошо заметной коронкой, он отделен диастемой от остальных. Центральные зубцы крупные, двухвершинные, дорсальные – одновершинные, длинные, острые, с узким основанием, последний дорсальный зубец маленький, двухвершинный (рис. 4a). У основания зубцов, начиная со второго центрального, ряды длинных шипиков, расположенные прямым углом.
Максилла и максиллипед несут опушенные щетинки (рис. 4b, 4c).
Каудальные ветви короткие, трапециевидной формы. С внутренней стороны густо опушены с апикального конца до середины. Каудальные ветви вооружены пятью апикальными щетинками. Апикальная внешняя щетинка шипообразная, голая, резко суженная к апикальному концу, равна по длине каудальным ветвям; внутренняя апикальная щетинка – прозрачная, тонкая, смещена на дорсальную сторону, по длине почти равна внешней шипообразной щетинке (табл. 1). Три центральные апикальные щетинки толстые, густо оперенные, почти равной длины.
Четыре пары плавательных ног самки одинакового строения. Базиподит Р5 самки с внешней стороны несет щетинку (рис. 4f), 1-й членик экзоподита с внешней стороны вооружен одним шипом, 2-й членик экзоподита с внешней стороны вооружен двумя острыми шипами, из которых один смещен к апикальному концу. С внутренней стороны этот членик вооружен четырьмя толстыми двухвершинными зубцевидными выростами (рис. 4f). На апикальном конце имеется длинный слегка искривленный шип, внутренняя поверхность которого с редко посаженными зубчиками (рис. 4h).
Самец. Ноги самца H. borealis (рис. 5а–5g) 2–4 пары асимметричны. Асимметрия проявляется в вооружении и длине 2-го и 3-го члеников экзоподита правой и левой ног (рис. 5а–5c). Левая нога 5-й пары самца: базиподит с серповидно изогнутым эндоподитом (рис. 5е). 1-й членик экзоподита короткий с одним шипом в дистальной части наружного края. 2-й членик экзоподита с двумя тонкими острыми шипами наружного края (рис. 5d). Два шипа на апикальном конце, из которых внешний тонкий, короткий, а внутренний длинный. Внутренняя сторона этого членика с тонкими волосками. Правая нога 5-й пары самца с толстым базиподитом и двумя члениками экзоподита, из которых первый длиннее второго. На 1-м членике экзоподита с наружной стороны один маленький шип, на 2-м членике экзоподита три шипа, два из которых с наружной стороны, один расположен апикально (рис. 5f, 5g).
Рис. 5.
Heterocope borealis (Fisher 1851), самец: a, b, c – плавательные ноги Р2-Р4; d – левая Р5; е – эндоподит левой Р5; f – правая Р5; g – апикальная часть exp2 правой Р5.
Распространение. Основная часть ареала H. borealis приурочена к заполярным широтам Палеарктики, где вид вполне обычен (Боруцкий и др., 1991). Также он встречается в высокогорных водоемах Восточной Сибири (Окинские и Восточные Саяны (Шевелева и др., 2009), Баргузинский и Икатский хребты (Шевелева и др., 2009), Становое и Патомское нагорье (Шевелева и др., 2009), Витимское плоскогорье (Шевелева и др., 2009) и Дальнего Востока (Бассейн р. Анадырь (Стрелецкая, 2010), водоемы Камчатки (Куренков, 2005). Отмечен в крупных глубоководных водоемах Сибири (Хантайское и Красноярское водохранилища) (Шевелева, 1986; Приймаченко и др., 1993), а также в самом Енисее, ниже по течению Красноярской ГЭС, до Игарки (Приймаченко и др., 1993), озера Тоджинской (Попкова, 2004; Зуйкова, Бочкарев, 2009) и Дархатской (Дулмаа, 2009) котловин (рис. 6).
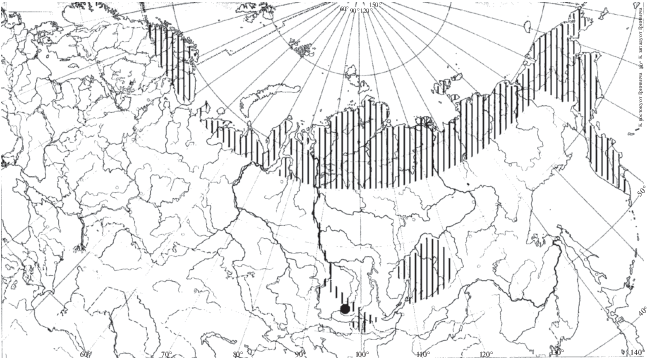
Ранее (Боруцкий и др., 1991) обнаружение вида южнее Полярного круга объясняли случайным заносом. Находки вида в водоемах Украины сомнительны, возможно, являются результатом путаницы с H. caspia (Błędzki, Rybak, 2016). Вероятно, в европейской части ареала распространение ограничено полярными широтами, в азиатской, в условиях резко континентального климата, вид населяет и более южные широты (до 51° с.ш., Окинские озера; данные Н.Г. Шевелевой) в высокогорьях и в глубоководных водоемах. Не исключено также, что такой явно разорванный ареал вида может указывать на возможные таксономические различия между рачками северной и южной частями ареала, что требует дополнительных детальных морфологических исследований.
Heterocope caspia Sars 1897
Материал. Особи разных полов из Нижней Волги (Волгоградское водохранилище, 50°47′ с.ш, 45°59′ в.д и не зарегулированный участок реки ниже г. Волгоград, 47°01′ с.ш и 47°29′ в.д., август, 2018 г.).
Описание. Самка. Тело удлиненное, стройное, хитин очень нежный. Цефалоторакс в 3.0–3.2 раза превышает ширину.
Длина генитального сомита самки в 1.16–1.29 раза превосходит его ширину. Пластинка генитального сомита имеет по два зубца с каждой стороны, из которых внешний зубец крупный с длинным отростком, внутренний – маленький (рис. 8c). Нижняя поверхность генитального сомита кзади от генитального отверстия несет 2 ряда сетул и 1 ряд спинул, расположенных поперечными рядами (рис. 8d).
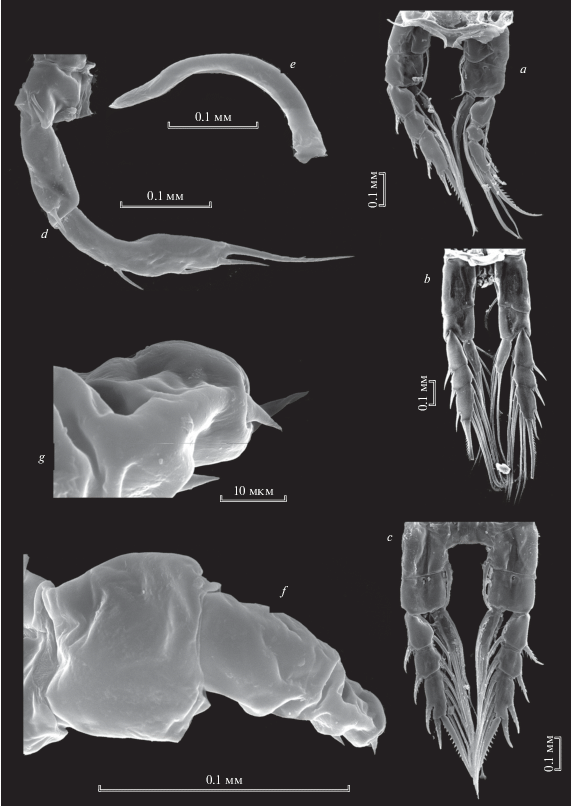
Рис. 7.
Heterocope caspia Sars 1897: a – максиллипед самки; b – режущий край мандибулярной пластинки самки; c – каудальные ветви самца; d – Р5 самца; е – правая Р5 самца.

Рис. 8.
Heterocope caspia Sars 1897, самка: a – каудальные ветви; b, е – Р5 самки; с, d – генитальная пластинка; f – Р5exp2; g – апикальная часть Р5exp2.
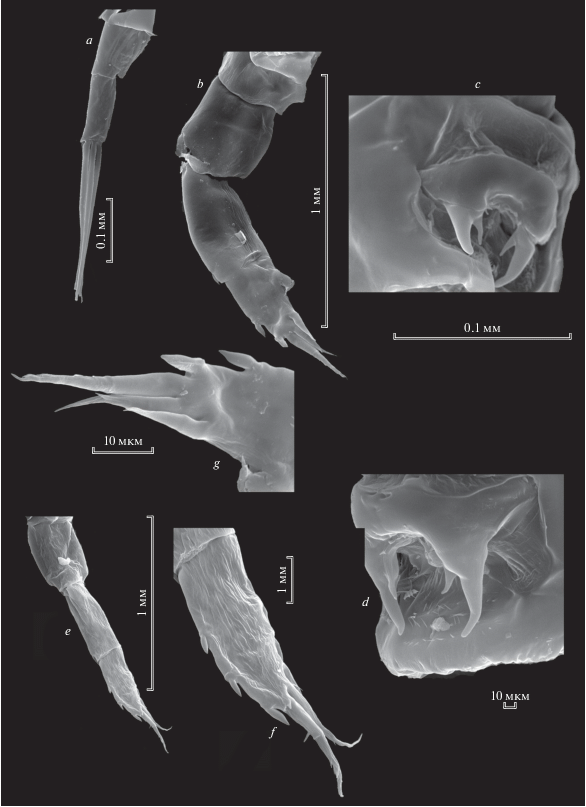
Каудальные ветви длинные (рис. 8a), из 5 исследованных нами видов рода у H. caspia они самые длинные (табл. 1), параллельные, голые. Они несут 4 апикальные щетинки: одну внутреннюю тонкую, которая, как и у всех остальных видов, смещена на дорсальную сторону, она самая короткая среди видов рода (табл. 1). Три апикальные щетинки равномерно сужаются к концу, вооружены относительно короткими, редкими, тонкими сетулами. Внешняя апикальная щетинка, короткая, шипообразная.
Антеннулы достигают заднего края генитального сомита.
Режущий край мандибулы с шестью зубцами (рис. 7b). Вентральный зубец одновершинный, заостренный, с хорошо заметной коронкой, отделен от остальных зубцов глубокой диастемой. Три центральных зубца двухвершинные, с широким основанием, тупые. Следующие два дорсальных зубца с более узким основанием, тоже двувершинные, острые; самый дорсальный зубец шипообразный, тонкий длинный. Дорсальная щетинка длинная, тонкая, зубчатая. В основании зубцов (за исключением двух самых вентральных) поперечный ряд шипиков.
Щетинки максиллипеда опушенные частыми длинными волосками (рис. 7a).
Пятая нога самки одноветвистая, 4-членистая (рис. 8b, 8e). С внешней стороны на ехр1имеется один шип. На ехр2 находятся 2–3 коротких шипа на внешней стороне, 1 длинный апикальный шип. 3–4 зубцевидных выроста на внутренней стороне (рис. 8g). Субапикальный вырост почти такой же длины как апикальный шип.
Самец. Пятая правая нога одноветвистая, 2-членистая, внутренняя сторона гладкая, внешняя – бугорчатая (рис. 7d, 7e). Левая нога двуветвистая с серповидно изогнутым эндоподитом (рис. 7d) и 2-членистым экзоподитом. Второй членик экзоподита в 2 раза длиннее первого, утолщенный посередине, с тупым апикальным концом, на котором два маленьких шипа равные по длине, на внешней стороне имеются маленькие шипики.
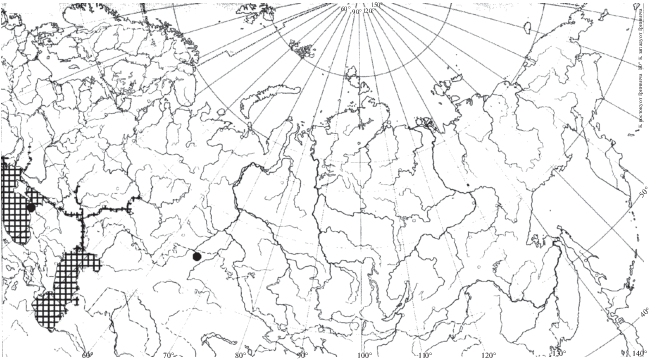
Распространение. Ареал H. caspia приурочен к Азовскому, Каспийскому, Черному морям и их бассейнам (Боруцкий и др., 1991). Из этой основной части ареала рачок расселился вверх по рекам Волге (до Куйбышевского водохранилища включительно, куда вселился еще в 1967 г. (Мордухай-Болтовской, Дзюбан, 1976), Дунаю, Дону, Южному Бугу, где поднимается довольно высоко (Попов, 2007). В 2016 г. отмечен в верховьях Камского водохранилища (Лазарева и др., 2018). Отмечен также в озерах Абрау (Краснодарский край) (Степаньянц и др., 2015), Караколь (Казахстан) (Карапун и др., 2013) и Сапанджа (Турция) (Степаньянц и др., 2015).
Heterocop saliens (Lilljeborg 1862)
Материал. Особи разных полов из проб зоопланктона оз. Пустое (Пустынное) (Нижнегородская обл., 56°28′ с.ш. и 45°18′ в.д., август 2018 г.).
Описание. Самка. Тело удлиненное, стройное, длина цефалоторакса в 2.5–2.8 раза больше ширины.
Длина генитального сомита самки немногим превышает его ширину (табл. 1), его пластинка в проксимальной части с неглубокой выемкой в центре и широкими невысокими латеральными лопастями (рис. 10d).
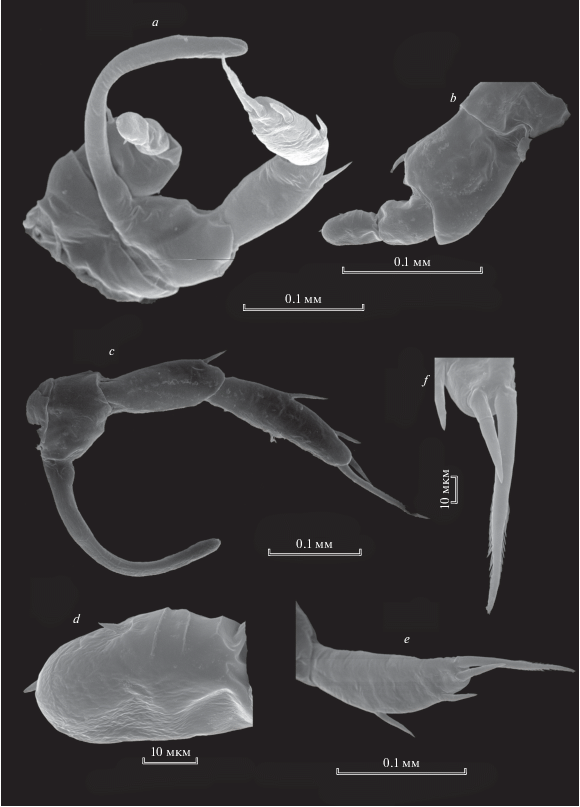
Рис. 10.
Heterocope saliens (Lilljeborg 1862), самка: а – режущий край мандибулярной пластинки; b – максиллипед; c – максилла; d – генитальный сегмент; e – генитальный сегмент и абдомен; f – Р5; g – Р5exp2; h – апикальный шип Р5exp2.
Каудальные ветви относительно короткие, их длина в 1.3–1.6 раз превышает ширину (табл. 1), немного расширены к апикальному концу, с внутренней стороны опушены грубыми частыми волосками, от апикального конца к середине. Каудальные ветви несут три перистые относительно длинные щетинки, одну шипообразную внешнюю и одну тонкую внутреннюю щетинку. Внешняя апикальная голая шипообразная щетинка в 2 раза короче, чем у H. borealis (табл. 1), тонкая апикальная внутренняя, как и у всех видов этого рода, смещена на дорсальную сторону. Она также короче, чем у H. borealis.
Антеннулы достигают заднего конца цефалоторакса.
Режущий край мандибулярной пластинки с острыми зубцами, характерными для питания животной пищей. Вентральный зубец острый 1-вершинный, отделен от остальных (рис. 10а). Центральных 3 зубца 2-вершинные, с широким основанием, три более дорсальных зубца 1-вершинные, с узким основанием, дорсальная щетинка острая, тонкая. В основании четырех центральных зубцов есть три продольных ряда шипиков (рис. 10a).
Щетинки максиллы и максиллипеда покрыты длинными волосками, максиллипеды крупные (рис. 10b, 10c).
Пятая нога имеет сходные черты с H. borealis (рис. 10f), за исключением того, что P5exp2 более короткий и утолщенный в базальной части (рис. 4f). Зубцевидные выросты внутреннего края в количестве 4 этого членика на вершине четко раздвоены (рис. 10g). На апикальном конце членика длинный шип (рис. 10h).
Самец. Плавательные ноги Р1–Р4 симметричные. Базиподит правой ноги 5-й пары самца крупный, с шипом на внешнем крае (рис. 11b). Его 2-членистый экзоподит четко отделен, два членика экзоподита почти равной длины, на его втором членике имеются 2 маленьких шипа и поперечные складки с наружной стороны (рис. 11d). Левая нога 5-й пары самца имеет тонкий серповидный эндоподит, который четко отделен от базиподита и двух экзоподитов, из которых 1-й экзоподит короче второго (рис. 11c). Второй членик экзоподита с выпуклым внешним краем, несет 2 латеральных шипа. Апикально расположены 2 шипа, из них короткий шип достигает середины длины второго шипа (рис. 11e). Длинный шип на конце зазубрен (рис. 11 f).
Рис. 11.
Heterocope saliens ( Lilljeborg 1862), самец: a – Р5; b – правая Р5; c – левая Р5; d – апикальная часть exp2 правой Р5; e – exp2 левой Р5; f – апикальная часть exp2 левой Р5.
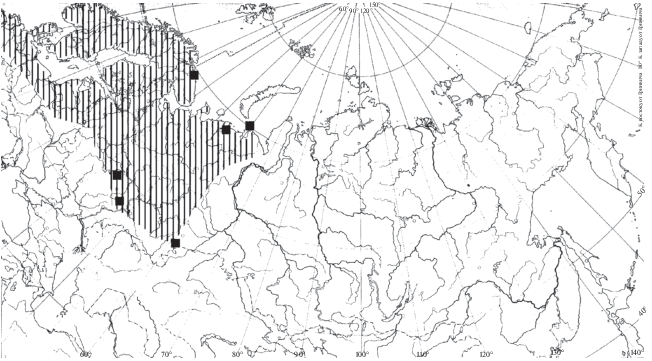
Распространение. H. saliens заселяет водоемы разного типа в различных ландшафтах в полярных, бореальных и, частично, в умеренных широтах Европейской части Палеарктики (Боруцкий и др., 1991; Błędzki, Rybak, 2016) (с полярных широт до 56ос.ш. – оз. Пустое (Пустынное), Нижегородская обл.) (рис. 13). В отличие от H. appendiculata и H. borealis, в Европе встречается часто (Błędzki, Rybak, 2016).
Heterocope soldatovi Rylov 1922
Материал. Особи обоих полов из пробы зоопланктона оз. Болонь (Хабаровский край, 49°84′ с.ш, 36°27′ в.д.).
Описание. Самка. Тело удлиненное, равномерно суживается кзади.
Генитальный сомит почти квадратный, в базальной части немного расширен. Внутренняя поверхность генитального сомита устлана 4 рядами поперечно расположенных шипиков (рис. 14b). Половое отверстие имеет генитальную пластинку с двумя зубцами с каждой стороны (рис. 14b).
Рис. 14.
Heterocope soldatovi Rylov 1922: а – максилла самки; b – генитальный сегмент; с, d – Р5 самки; е – эндоподит левой Р5 самца; f – левая Р5 самца; g – exp2 левой Р5 самца; h – апикальные шипы exp2 левой Р5 самца; i – правая Р5 самца.
Каудальные ветви удлиненные, к апикальному концу слегка расширены, опушенные сетулами на 1/2–1/3 своей длины. Тонкая апикальная внутренняя щетинка смещена на дорсальную сторону.
Антеннулы длинные, достигают последнего абдоминального сомита.
Режущий край мандибулярной пластинки представлен семью зубцами, из которых 6 – одновершинные (рис. 13a).
Щетинки максиллы длинные оперенные (рис. 13b, 14a).
Пятая нога самки: базиподит широкий, с закругленным внутренним краем (рис. 13c, 14d). Первый членик экзоподита удлиненный, на внешней стороне маленький шипик. P5exp2 примерно такой же длины, как P5exp1. На его наружной стороне два маленьких шипа, один из которых вблизи апикального шипа; на внутренней стороне 4 удлиненных, тонких и заостренных зубцевидных выроста, из них зубцевидный вырост около апикального шипа длиннее остальных трех; этот членик заканчивается длинным загнутым внутрь сильным шипом, на ½ своей длины зазубренным (рис. 14c, 14d).
Самец. Базиподит правой ноги самца толстый, экзоподит неправильной формы, 1-членистый, широкий и закругленный на конце (рис. 14i), с внутренней стороны имеет три маленьких шипа. Эндоподит левой ноги серповидно изогнут, закругленный на конце, в базальной и дистальной частях утолщен (рис. 14e). 1-й членик экзоподита короткий, расширен в дистальной части. 2-й членик экзоподита вытянутый, на апикальном конце вооружен длинным толстым острым шипом (рис. 14 f). Возле апикального шипа прикреплен изогнутый внутрь относительно короткий острый шип (рис. 14h). На внутреннем крае 2-го членика экзоподита находится 6–8 конических шипиков, из которых первый расположен примерно на середине края, а последний в основании апикального шипа. Эти шипики заходят на поверхность членика (рис. 14g).
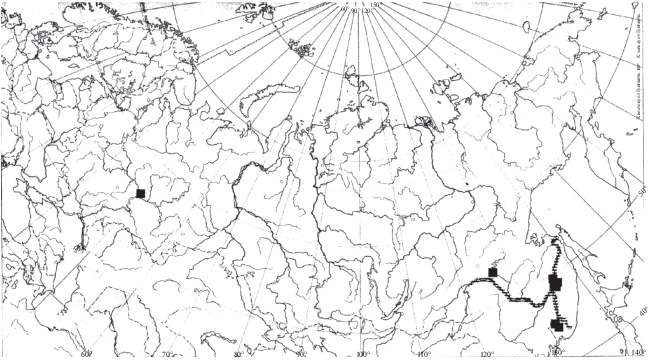
Распространение. Вид характеризуется крайне ограниченным распространением – нижним течением р. Амур, а также реками Уссури (Рылов, 1922; 1932), Сунгача (Барабанщиков, 2001) и озерами Хасан (собственные данные Н.Г. Шевелевой.), Болонь, Лебехэ, Ханка, Петропавловское, Кабар, Гасси (Боруцкий, 1952) его бассейна (рис. 15). Находка этого вида в европейской части Палеарктики, в одном из водоемов г. Казань (Мингазова и др., 2008) сомнительна. Возможно, H. caspia (населяющая Куйбышевское водохранилище, с которым сообщаются водоемы г. Казань) была ошибочно принята за H. soldatovi.
Для практических целей по общим и отличительным морфологическим признакам составлена определительная таблица для самок (табл. 2). Ранее подобные, но отличные таблицы были предложены Рыловым (1930) и Боруцким и соавт. (1991). Наиболее схожие черты морфологии (например, наличие внешней шипообразной щетинки на каудальных ветвях) имеют H. borealis и H. saliens, но по самкам легко отличаются. Так, у H. borealis: 1) относительно короткие каудальные ветви, которые имеют трапециевидную форму; 2) большая длина шипообразной щетинки; 3) генитальная пластинка, имеет по одному короткому закругленному выросту с каждой стороны; 4) есть четыре слабо двухвершинных зубцевидных выроста у Р5ехр2 с внутренней стороны. У самца вооружение плавательных ног Р2–Р4 асимметричное. H. saliens у самца вооружение плавательных ног – симметричное. У самки генитальная пластинка без придатков; Р5ехр2 внутренние зубцевидные выросты четко двувершиные. В живом и фиксированном состоянии особи разных полов H. borealis окрашены в голубой цвет.
Таблица 2.
Ключ для определения видов рода Heterocope фауны России
| 1 | L : W каудальных ветвей более 2.5; их внутренние поверхности голые…………………………… | H. caspia |
| – | L : W каудальных ветвей менее 2.0; их внутренние поверхности несут волоски……………… | 2 |
| 2 | Внешняя щетинка каудальных ветвей имеется…………………………… | 3 |
| – | Внешняя щетинка каудальных ветвей отсутствует……………………… | 4 |
| 3 | Генитальная пластинка с двумя закругленными выростами…………… | H. borеalis |
| – | Генитальная пластинка без выростов, с двумя невысокими, широкими лопастями……… | H. saliens |
| 4 | Генитальная пластинка несет лентовидные придатки в центре и по 1 двухвершинному латеральному зубцу с каждой стороны……………….. | H. appendiculata |
| – | Генитальная пластинка несет 2 латеральных зубца с каждой стороны…………………………… | H. soldatovi |
Самки H. appendiculata имеют лентообразную генитальную пластинку, самец этого вида на Р5 левой ноги на втором экзоподите имеет два маленьких шипа на внешней стороне и два шипа на его апикальном конце разной длины. Правая нога Р5 самца неявно расчлененная, утолщенная в средней части, заканчивается маленьким шипом.
Самки H. caspia и H. soldatovi имеют генитальную пластинку с двумя латеральными зубцами с каждой стороны, но генитальная поверхность H. caspia внутри устлана тонкими сетулами, в то время как у H. soldatovi эти сетулы более крупные и явно заметные. У самки H. caspia 2-й членик экзоподита Р5 с внутренней стороны вооружен 2–3 одновершинными зубцевидными выростами; апикально расположен один относительно длинный шип. Левая нога Р5 самца H. caspia на 2-м членике экзоподита имеет очень маленькие шипики как на внешней стороне, так и на апикальном конце, правая нога без вооружения, имеет волнистую поверхность. Cамки и самцы H. soldatovi имеют очень характерные морфологические признаки, присущие только этому виду. Так, 2-й членик экзоподита Р5 самки с внутренней стороны несет 4 и с внешней 2 тонких, относительно длинных искривленных зубцевидных выроста, на апикальном конце 1 длинный шип (табл. 1). У самца Р5 левая и правая ноги по строению и вооружению отличаются от всех других видов. Так, экзоподит Р5 правой ноги широкий, плоский, закругленный с маленькими шипиками по внутреннему краю. 2-й членик экзоподита левой ноги прямоугольной формы, вытянутый, почти с параллельными антериорной и постериорной сторонами, на конце несет два шипа, из которых один длинный, второй короткий изогнутый. На внутренней стороне экзоподита находится 6–8 конических хитиновых выростов.
Таким образом, все пять видов Heterocope, обитающих на территории России, четко отличаются друг от друга как по самкам, так и по самцам. Подробный сравнительный анализ морфологических различий и составленная на основании его определительная таблица видов послужат для однозначной идентификации представителей рода. Это особенно актуально для анализа фауны регионов, где совместно обитают разные виды Heterocope, что типично для севера Палеарктики.
Список литературы
Барабанщиков Е.И., 2001.Характеристика планктонного сообщества нижней части бассейна реки Сунгача // Рыбохозяйственная наука на пути в XXI век: Тез. докл. Всерос. конф. молодых ученых. Владивосток: Изд-во ТИНРО-Центра. С. 80–82.
Баянов Н.Г., 2002. Зоопланктон заволжских озер Нижегородской области // Труды Государственного природного заповедника “Керженский”. Н. Новгород. С. 87–103.
Боруцкий Е.В., 1952. Сестон бассейна Амура и его роль в питании амурских рыб // Труды Амурской ихтиологической экспедиции 1945–1949. (Издание московского общества испытателей природы). Т. III. С. 141–229.
Боруцкий Е.В., 1960. Определитель свободноживущих пресноводных веслоногих раков СССР и сопредельных стран по фрагментам в кишечниках рыб. М.: АН СССР. 216 с.
Боруцкий Е.В., Степанова Л.А., Кос М.С., 1991. Определитель Calanoida пресных вод СССР. СПб.: Наука. 502 с.
Горшкова А.Т., 2012. Оценка уровня самоочищения озер Кабан по анализу зоопланктонного комплекса // Георесурсы. № 7 (49). С. 29–32.
Дулмаа А., 2009.Ракообразные (Crustacea) водоемов Дархатской котловины // Аннотированный список фауны озера Байкал и его водосборного бассейна. Т. II. Водоемы и водотоки юга Восточной Сибири и Северной Монголии. Книга I. Новосибирск: Наука. С. 628–642.
Зуйкова Е.И., Бочкарев Н.А., 2009. Характеристика пелагического зоопланктона крупных озер Тоджинской котловины (бассейн реки Большой Енисей, Тува) // Биология внутренних вод. № 1. С. 53–61.
Карапун М.Ю., Юрченко В.В., Бирюкова М.Г., Айтимова А.М., 2013. Качественный состав планктонных организмов озера Караколь под влиянием техногенного фактора // Вестник АГТУ. Сер. Рыбное хозяйство. № 2. С. 42–49.
Куренков И.И., 2005. Зоопланктон озер Камчатки. Петропавловск-Камчатский: КамчатНИРО. 178 с.
Лазарева В.И., 2005. Цикл развития и динамика численности хищного рачка Heterocope appendiculata Sars (Copepoda, Temoridae) в небольшом лесном пруду // Биология внутренних вод. № 4. С. 40–46.
Лазарева В.И., Сабитова Р.З., Быкова С.В., Жданова С.М., Соколова Е.А., 2018. Распределение летнего зоопланктона в каскаде водохранилищ Волги и Камы // Труды ИБВВ РАН. Вып. 83 (86). С. 62–83.
Монаков А.В., 1998. Питание пресноводных беспозвоночных. М: ИППЭ. 318 с.
Мингазова Н.М., Деревенская О.Ю., Палагушкина О.В., Павлова Л.Р., Набеева Э.Г., Зарипова Н.Р., Замалетдинов Р.И., Кондратьева Т.А., Павлов Ю.И., Унковская Е.Н., Борисович М.Г., Халиуллина Л.Ю., 2008. Биоразнообразие водных объектов г. Казани // Ученые записки Казанского государственного университета. Серия: Естественные науки. Т. 150. № 4. С. 252–260.
Мордухай-Болтовской Ф.Д., Дзюбан Н.А., 1976. Изменение в составе и распределении фауны Волги в результате антропогенных воздействий // Биологические продукционные процессы в бассейне Волги. Л.: Наука. С. 67–81.
Пежева М.Х., Казанчев С.Ч., Авалишвили Е.Т., 2016. Типологическая классификация высокогорных карстовых озер Кабардино-Балкарской Республики на основе данных по зоопланктону // Вестник Красноярского государственного аграрного университета. № 6 (117). С. 50–56.
Попкова Л.А., 2004. Фауна зоопланктона республики Тыва // Проблемы вида и видообразования: Доклады III Междунар. конф. С. 96–101.
Попов А.И., 2007. Некоторые данные по видовому составу и структуре зоопланктона Саратовского водохранилища // Водные экосистемы. Т. 9. № 4. С. 1013–1019.
Подшивалина В.Н., 2006. Элементы северной фауны в зоопланктоне разнотипных озер Низменного Заволжья (на примере водоемов Чувашской Республики) // Современные экологические проблемы Севера: материалы Междунар. конф.: в 2 ч. Ч. 1. Апатиты: Изд-во Кольского научного центра РАН. С. 201–202.
Приймаченко А.Д., Шевелева Н.Г., Покатилова Т.Н. и др., 1993. Продукционно-гидробиологические исследования Енисея. Новосибирск: Наука. 194 с.
Ривьер И.К., Дзюбан Н.А., 1978. Зоопланктон // Волга и ее жизнь. Л.: Наука. С. 153–178.
Ривьер И.К., Литвинов А.С., 2006. Сравнительный анализ зоопланктона Шекснинского водохранилища в 1987 и 2001 гг. // Водные ресурсы. Т. 33. № 5. С. 615–629.
Рылов В.М., 1922. Heterocope soldatovi n. sp., новый вид пресноводного ракообразного (Copepoda, Calanoida) // Ежегодник Зоол. музея Рос. Акад. Наук. Т. 23. С. 164–178.
Рылов В.М., 1930. Пресноводные CALANOIDA С.С.С.Р. // Определители организмов пресных вод С.С.С.Р. А. Пресноводная фауна. Вып. 1. Ленинград: ВАСХНИЛ. 288 с.
Семенова Л.М., 2009. Состояние зоопланктона в водоеме-охладителе Чернобыльской АЭС и в верховьях Киевского водохранилища // Биология внутренних вод. № 3. С. 79–86.
Семенова Л.М., Маркевич Г.И., Гребенюк Л.П., 2005. Видовой состав и сезонная динамика Уводьского водохранилища // Биология внутренних вод. № 2. С. 75–81.
Степаньянц С.Д., Хлебович В.В., Алексеев В.Р., Данелия М.Е., Петрошев В.В., 2015. Определитель рыб и беспозвоночных Каспийского моря. Т. 2. Стрекающие, гребневики, черви, веслоногие ракообразные и мизиды. СПб. М.: Товарищество научных изданий КМК. 244 с.
Стрелецкая Э.А., 2010. Обзор фауны коловраток (Rotatoria), ветвистоусых (Cladocera) и веслоногих ракообразных (Copepoda) бассейна реки Анадырь // Сибирский экологический журнал. Т. 17. № 4. С. 533–542.
Шевелева Н.Г., 1986. Зоопланктон // Гидрохимические и гидробиологические исследования Хантайского водохранилища. Новосибирск: Наука. С. 81–100.
Шевелева Н.Г., Аров И.В., Шабурова Н.И., Евстигнеева Т.Д., Итигилова М.Ц., 2009. Биоразнообразие коловраток (Rotifera) и низших ракообразных (Cladocera, Calanoida, Cyclopoida, Harpactocoida) горных озер юга Восточной Сибири // Биота водоемов байкальской рифтовой зоны. Иркутск. С. 83–94.
Błędzki L.A., Rybak J.I., 2016. Freshwater Crustacean Zooplankton of Europe. Cladocera and Copepoda (Calanoida, Cyclopoida). Key to species identification, with notes on ecology, distribution, methods and introduction to data analysis. Switzerland: Springer. 923 p.
Dussart B.H., 1967. Les Copepodes des eauxcontinentales d’Europeoccidentale. T. I: Calanoideset Harpacticoides. Paris. 500 p.
Kiefer F., Fryer G., 1978. Das Zooplankton der Binnengewässer. Bd. 26. Teil. 2. Stuttgart. 342 p.
Дополнительные материалы отсутствуют.
Инструменты
Зоологический журнал