

Зоологический журнал, 2020, T. 99, № 4, стр. 450-458
Изменчивость пения самцов голосистой пеночки (Phylloscopus schwarzi) в разные дни
Е. М. Шишкина a, *, А. С. Опаев a
a Институт проблем экологии и эволюции им. А.Н. Северцова РАН
119071 Москва, Россия
* E-mail: e.m.shishkina@yandex.ru
Поступила в редакцию 02.05.2019
После доработки 25.08.2019
Принята к публикации 13.10.2019
Аннотация
Структура и организация пения воробьиных птиц подробно описаны для ряда видов, однако недостаточно изученными остаются пределы внутрииндивидуальной изменчивости этих особенностей. Мы сравнили структуру и организацию пения конкретных самцов голосистой пеночки, вокализация которых была записана 2 или 3 раза с интервалом в несколько дней. Проанализировано 49 записей спонтанного пения 19 самцов длительностью в среднем около 5 мин каждая. Для каждой записи мы составили каталог типов песен и рассчитали индекс линейности. Для анализа различий в песенных наборах, исполняемых одной особью в разные дни, использовали коэффициенты Жаккара и Симпсона. Оказалось, что размеры репертуаров (количество разных типов песен) и индексы линейности в целом не различаются на записях одних и тех же особей, сделанных в разные дни. Однако конкретный состав песен может быть заметно различен: медианы коэффициентов Жаккара и Симпсона составили 0.42 и 0.11 соответственно. Мы выделили два дискретных типа изменений в пении: (1) добавление/пропуск одного типа песни (отмечено у всех самцов) и (2) добавление/пропуск целых песенных последовательностей из 5–15 типов песен (12 самцов из 19). Изменчивость в пении самцов в разные дни имеет характер случайных вариаций, а не направленных изменений – например, последовательной смены одних предпочитаемых в данный момент типов песен другими. Кроме того, характер изменчивости пения по дням не зависел от количества соседних самцов и расстояния до них, поэтому такая изменчивость вряд ли является особым механизмом социальной адаптации. Полученные данные мы обсуждаем с точки зрения возможных паттернов развития пения в онтогенезе.
Песня воробьиных птиц имеет много функций. Основные из них – привлечение брачного партнера и регулирование территориальных отношений (Catchpole, Slater, 2008). Пение состоит из стереотипных акустических единиц (например, типов песен или типов звуков), которые обычно чередуются при пении по определенным правилам (Kershenbaum et al., 2014). Поэтому говорят, что пение имеет синтаксис (Иваницкий, 2015). К настоящему времени накопилось уже довольно много примеров сложной синтаксической организации пения. Выявлено два главных типа синтаксиса: линейный и комбинаторный. При линейном синтаксисе очередность исполнения вокальных компонентов более или менее строго фиксирована. При комбинаторном синтаксисе выделяется один кластер компонентов (например, типов песен) или несколько таких кластеров; внутри кластера возможны изменения порядка исполнения типов песен (Okanoya, 2013; Ivanitskii et al., 2017). Каждый из этих типов синтаксиса может проявляться у разных видов птиц по отдельности или может встречаться в пределах одного вида (Иваницкий, Марова, 2010; Opaev, 2016; Опаев, Колесникова, 2019).
На фоне заметного числа работ, посвященных организации пения разных видов (обзор: Иваницкий, 2015), недостаточно изученными остаются пределы внутри-индивидуальной изменчивости этих параметров. В частности, это изменчивость организации пения данной особи на коротких промежутках времени – например, в течение одного сезона. Имеющиеся в литературе материалы говорят о том, что такая изменчивость в организации пения, в принципе, ожидаема. Как показано на примере ряда видов, используемые особью в данный момент времени наборы типов песен и/или звуков могут меняться – постепенно либо от случая к случаю (Kipper, Kiefer, 2010). Подобные изменения могут быть временными – т.е. птица на самом деле “помнит” песни, которые не исполняет в данных момент, но пела раньше, и может вернуться к ним в будущем (Geberzahn, Hultsch, 2003). Причины изменчивости структуры репертуара на коротких промежутках времени не всегда понятны, но в ряде случаев такая изменчивость может зависеть от пения других птиц того же вида. Так, самцы малой воробьиной овсянки (Spizella pusilla) со временем начинают исполнять преимущественно тот тип песни из своего репертуара, который поют большинство соседей. Т.е. в данном случае происходит “сближение” репертуаров разных птиц (Nelson, 1992). А у пеночки-веснички (Phylloscopus trochilus) – вида с варьирующей от случая к случаю структурой песни – длительность песни зависит от плотности популяции (Горецкая, 2013). В силу того, что меняются наборы песен/звуков (структура пения), вполне можно ожидать и соответствующих изменений в организации пения. Именно этому малоизученному вопросу посвящена данная работа.
Объект нашего исследования – голосистая, или толстоклювая пеночка (Phylloscopus schwarzi). Структура и организация пения этого вида неплохо изучена (Иваницкий и др., 2011; Опаев, Колесникова, 2019). Известно, что в пении каждого самца выделяются дискретные (длительностью около 1 с) стереотипные типы песен – до 40 и более в репертуаре одной особи. Характерен линейный синтаксис: большинство самцов исполняют все типы песен в определенной последовательности, а затем начинают все сначала. Цель данного исследования – сравнить структуру и организацию пения на фонограммах одних и тех же особей, но записанных с интервалом в несколько дней. Дополнительно мы посмотрели, зависят ли различия в пении в разные дни от наличия/отсутствия поблизости других самцов – как это выявлено для некоторых других видов (см. выше).
МАТЕРИАЛ И МЕТОДЫ
Материалом для исследования послужили записи пения 19 самцов голосистой пеночки, сделанные на территории Антоновского лесничества Хинганского заповедника (Амурская обл.) в мае–июне 2018 г. 9 из этих самцов были отловлены, помечены для индивидуального опознавания цветными пластиковыми кольцами и окрашены театральными гримом. Мы делали записи спонтанного пения самцов длительностью в среднем 338 с, минимальная длительность составила 157 с, максимальная – 565 с. Всего было сделано 49 записей, по 3 записи спонтанного пения в разные дни для 11 самцов и по 2 записи – для 8. Общая длительность анализируемых записей составила около 4.5 ч (16562 с). Всего было проанализировано 1696 песен, в среднем по 34 песни в каждой записи. Минимальное и максимальное число песен на записях было 13 и 65, соответственно. Записи делались в среднем с интервалом в 4 дня (разброс 1–17 дней), с 8:00 до 18:00. Для записи пеночек использовали диктофон Marantz PMD-660 и микрофон Sennheiser ME66 с предусилителем К6.
Координаты точек, где пел каждый самец, были сняты при помощи GPS-навигатора Garmin и визуализированы в программе SASPlanet для дальнейшей обработки. Локализацию каждого самца отмечали только один раз – в первый день наблюдений за ним. Затем мы посчитали количество особей того же вида в радиусе 400 м от анализируемого самца – по результатам наших наблюдений, это соответствует максимальному радиусу территорий самцов голосистой пеночки данной популяции (см. также: Антонов, Квартальнов, 2014). Кроме этого, мы измеряли расстояние от каждого самца до его ближайшего соседа.
Визуализацию и обработку фонограмм проводили в программе Syrinx. На первом этапе для каждой фонограммы составляли каталог типов песен. Тип песни голосистой пеночки – это стереотипная комбинация звуков длительностью около 1–1.5 с, в практически неизменном виде повторяемая самцом по ходу пения (Опаев, Колесникова, 2019). Количество типов песен на той или иной фонограмме – это наблюдаемый размер репертуара (или разнообразие пения на данном отрезке времени). Затем каждый тип песни был обозначен цифрой, а пение было представлено в виде последовательности цифр. По этой последовательности в программе Past3 мы строили матрицы наблюдаемых переходов. В данном случае последовательность песен рассматривалась как Марковская цепь первого порядка. По матрицам наблюдаемых переходов рассчитывали индекс линейности: SLIN = размер репертуара/количество типов переходов между песнями разных типов (Scharff, Nottebohm, 1991).
Дальнейшую обработку данных проводили в среде программирования R 3.5.2 (R Core Team, 2016). Для визуализации наблюдаемых переходов между песнями разных типов использовали пакет ‘markovchain’ в среде R (Spedicato et al., 2017). Кроме того, мы сравнивали попарно наборы типов песен данного самца голосистой пеночки в разные дни. Для этого мы использовали, во-первых, коэффициент общности Жаккара, который изменяется от 0 до 1 и показывает, насколько схожи (0) или различны (1) сравниваемые наборы песен. Также, во-вторых, мы использовали коэффициент перекрытия Симпсона. Он показывает, в какой степени набор песен, использованный самцом в один из двух сравниваемых дней, полностью повторяется в другой. Чем ближе значение этого показателя к 0, тем больше типов песен, встреченных в один день, использовалось и в другой. Расчеты коэффициентов проводили в пакете ‘betapart’ в среде R (Baselga, Orme, 2012; Baselga et al., 2017). Для выявления связи между всеми анализируемыми параметрами была использована ранговая корреляция Спирмена. В анализе различий между выборками мы применяли критерии Манна-Уитни и Краскела-Уоллиса.
Использованные в работе записи содержали разное количество песен. Чтобы проверить, не повлияло ли это на результаты, мы посмотрели связи (корреляция Спирмена) между (1) индексами Жаккара и Симпсона и (2) количеством песен на сравниваемых индексами записях. Ни в одном тесте мы не обнаружили достоверной корреляции (p > 0.05). Таким образом, структура нашей выборки не влияла на основные результаты исследования.
РЕЗУЛЬТАТЫ
Изменчивость наборов песен
На одной фонограмме (записи) одной особи было выявлено 11–34 типов песен (в среднем 20 типов, n = 49 записей от 19 самцов). Разнообразие пения (количество типов песен) данного самца в разные дни в целом достоверно не различалось (критерий Краскела-Уоллиса = 0.91, p = = 0.63, n = 19) (рис. 1). Однако у некоторых особей различия были заметны (пример: рис. 4).
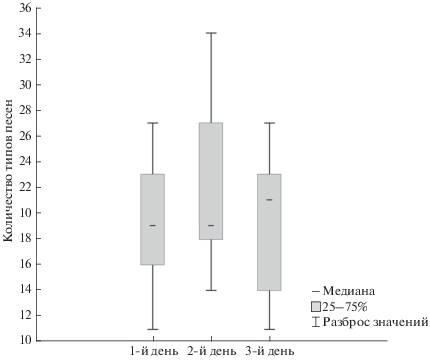
Суммарное количество типов песен на всех записях данного самца (общий размер репертуара) в среднем составило 48 типов песен. Наименьшее количество типов песен в репертуаре – 21 (суммарно на 2 записях), наибольшее – 68 (на 3 записях) (табл. 1). При этом в целом размер общего репертуара самца не зависел от того, записывали ли его в разные дни 2 или 3 раза (Z = 31.5, p = 0.32, n = 19).
Таблица 1.
Размеры репертуаров (количество типов песен) самцов голосистой пеночки в разные дни
| Самец | Репертуар | Число записей | |||
|---|---|---|---|---|---|
| 1-й день | 2-й день | 3-й день | общий | ||
| 24 | 20 | 19 | 21 | 52 | 3 |
| 26 | 17 | 19 | – | 21 | 2 |
| 27 | 20 | 20 | 26 | 68 | 3 |
| 28 | 26 | 27 | – | 40 | 2 |
| 29 | 25 | 19 | – | 34 | 2 |
| 30 | 14 | 29 | 13 | 30 | 3 |
| 31 | 23 | 18 | 21 | 63 | 3 |
| 32 | 13 | 19 | – | 46 | 2 |
| 33 | 15 | 27 | 23 | 61 | 3 |
| 36 | 24 | 14 | 14 | 39 | 3 |
| 37 | 16 | 22 | 23 | 64 | 3 |
| 39 | 22 | 17 | – | 58 | 2 |
| 40 | 18 | 21 | 20 | 41 | 3 |
| 41 | 20 | 18 | 21 | 24 | 3 |
| 42 | 18 | 15 | – | 52 | 2 |
| 43 | 11 | 30 | 11 | 51 | 3 |
| 44 | 18 | 20 | – | 38 | 2 |
| 46 | 19 | 15 | 27 | 49 | 3 |
| 47 | 27 | 34 | – | 57 | 2 |
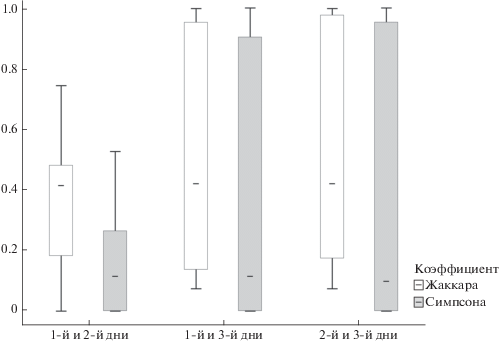
Конкретный самец в разные дни может использовать заметно различные наборы типов песен. Коэффициент Жаккара варьировал в пределах от 0 до 1 (медиана = 0.42). Значение коэффициента Симпсона также изменялось от 0 до 1, однако медиана этого показателя значительно меньше (0.11). Два рассматриваемых коэффициента сильно коррелировали друг с другом (R = 0.95, p < 0.05). При этом коэффициент Симпсона был всегда меньше, т.е. большинство песен, записанных в данный день, встречается и в другой день или другие дни. Коэффициенты Жаккара и Симпсона для песен самцов, записанных в течение трех дней, достоверно не различаются при сравнении каждой из “пар” дней (рис. 2). Таким образом, изменения в структуре репертуара носят скорее случайный, а не направленный характер.
Изменчивость в организации пения
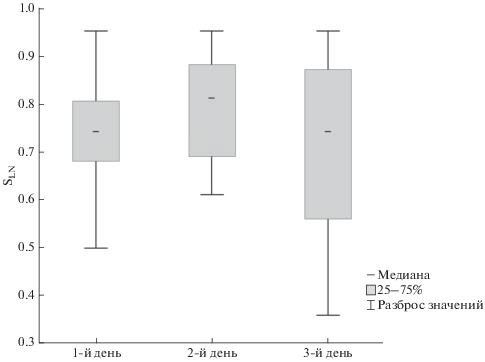
Организация пения большинства изученных самцов соответствовала линейному синтаксису. Значения медиан индексов линейности составили: в первый день записи пения 0.73 (разброс 0.50–0.95, n = 19 записей разных самцов), во второй день 0.80 (0.61–0.95, n=19), в третий 0.71 (0.36–0.95, n = 11) (рис. 3). Достоверных различий по индексам линейности между записями одного и того же самца в разные дни мы не выявили (Н = = 2.99, p = 0.22). Связи значений коэффициента Жаккара с характеристиками линейности пения обнаружено не было (R = –0.11, р > 0.05). Коэффициент Симпсона также не коррелировал с индексом линейности (R = –0.12, р > 0.05).
Таким образом, записи пения одних и тех же самцов голосистой пеночки в разные дни в целом не различаются по разнообразию (размеру репертуара) и организации пения (индексу линейности). Но конкретные наборы используемых песен не одинаковы. Чтобы посмотреть различия между записями в разные дни детальнее, мы построили и сравнили схемы переходов между песнями разных типов для каждой изученной записи. В итоге мы выделили два типа изменений песенных циклов самцов голосистой пеночки. Отметим, что “промежуточных” изменений между этими типами мы не зафиксировали.
В первом случае в уже имеющиеся линейные последовательности песен добавляются и/или исчезают из них отдельные единичные типы песен (рис. 4). Иными словами, несколько видоизменяются одни и те же секвенции песен. Этот тип изменения песенных последовательностей самцов голосистой пеночки мы определили как “добавление/пропуск песни”. Он зафиксирован у всех особей (n = 19).
Пример такого рода показан на рис. 4. Здесь изображены последовательности песен самца № 26 в 1-й и 2-й дни записи. Так, в 1-й день фигурирует последовательность из следующих типов песен: 8–5–7. Она же встречается и во 2-й день, но иногда сюда добавляется “новый” тип 18 (получается, в частности: 8–5–18–7). В другой части цикла в оба дня записи есть последовательность из песен 1–11–13. Однако во 2-й день записи (рис. 4) самец не всегда использовал тип песни 11 (наблюдалось только: 1–13). Стоит отметить, что в обоих случаях организация пения данного самца практически идентична по набору типов песен (рис. 4).
Во втором случае, зафиксированном у 12 самцов, добавляются/исчезают уже целые последовательности из пяти или более типов песен: в нашей выборке 5–15 типов, медиана 10 (n = 12). Эти кластеры песен не встречались в пении данного самца на записи в определенный день, но были отмечены на других его записях. Такие кластеры, предположительно, представляли из себя отдельные стереотипные линейные последовательности (рис. 5).
Рис. 4.
Организация спонтанного пения самца голосистой пеночки (№ 26) в разные дни (1-й и 2-й дни записи. Овалами одинакового цвета обозначены участки цикла с одинаковыми типами изменений: красный – пропускание песни, синий – добавление песни).
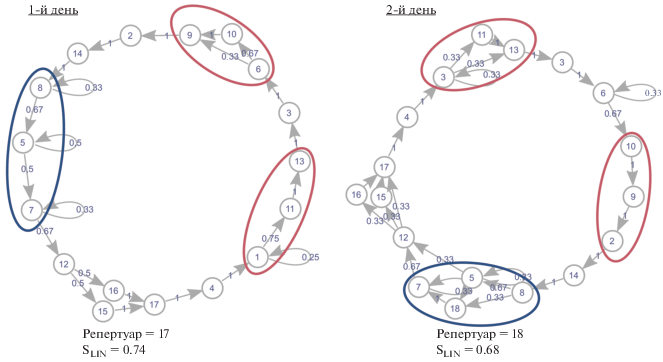
Рис. 5.
Организация спонтанного пения самца голосистой пеночки (№ 36) в разные дни (1-й и 2-й дни записи. Зеленым овалом обведен не повторяющийся участок цикла).
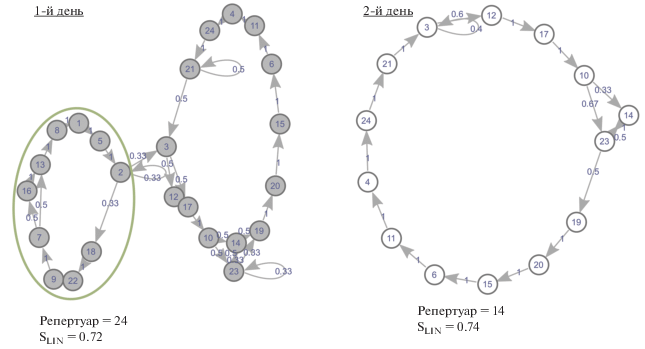
В качестве примера, на рис. 5 приведена организация пения самца № 36 в 1-й и 2-й день записи. Видно, что пение этой особи в разные дни различается добавлением/пропуском значительного по продолжительности фрагмента из 10 типов песен. Кроме того, этот самец, как и все прочие, демонстрировал добавление/пропуск отдельных типов песен в выявленных стереотипных (линейных) последовательностях, о чем сказано выше.
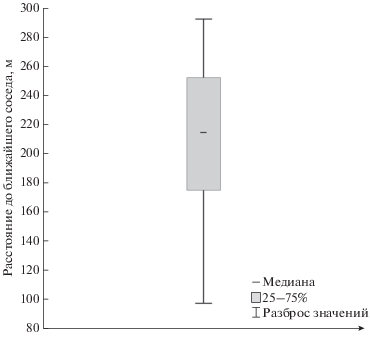
Расстояние до ближайшего соседа и изменчивость пения
Медианное расстояние от анализируемого самца до ближайшего соседа составило 214 м, минимальное значение этого параметра было равно 98 м, максимальное – 292 м (рис. 6). У восьми самцов в радиусе 400 м находилось по две особи того же вида, у 11 самцов мы наблюдали только одного соседа. Расстояние до ближайшего соседа коррелировало с числом соседних особей (R = 0.87, р < 0.05). Мы не выявили связи между (1) количеством самцов в радиусе 400 м от фокального и (2) расстоянием до ближайшего соседа с коэффициентами Жаккара (R = 0.07 и R = 0.15, p > 0.05) и Симпсона (R = 0.19 и R = 0.27, p > 0.05).
ОБСУЖДЕНИЕ
Полученные в данной работе материалы по структуре и организации спонтанного пения голосистой пеночки в целом соответствуют опубликованным данным (Иваницкий и др., 2011; Опаев, Колесникова, 2019). Мы выявили также, что такие количественные параметры, как наблюдаемый размер репертуара и индекс линейности пения достоверно не различаются у конкретной особи в разные дни. Это значит, что по однократной записи пения мы можем вполне корректно оценить эти особенности, которые, вероятно, индивидуальны в том смысле, что характеризуют самца в течение, по крайней мере, нескольких недель.
Между тем, конкретный состав используемых типов песен в разные дни различался, иногда довольно заметно. Состав песен, использованных данным самцом в разные дни, различался в среднем на 42% (т.к. медианное значение коэффициента Жаккара равно 0.42). Мы предполагаем, что изменчивость в пении самцов в разные дни обусловлена скорее случайными вариациями, а не направленными изменениями (к примеру, постепенной сменой одних предпочитаемых типов песен другими). В пользу этого говорит то, что коэффициенты Жаккара и Симпсона между разными “парами” дней не различались (рис. 2). Об этом говорит также то, что коэффициент перекрытия Симпсона в основном стремится к нулю. Следовательно, изменчивость репертуара чаще достигается путем добавления новых типов песен в одну песенную последовательность, по сравнению с другой (или, по-другому, пропуском песен в одной из двух песенных последовательностей) (рис. 4), но не заменой одних типов на другие.
Анализ последовательностей песен самцов в разные дни также указывает на наличие здесь случайных вариаций. Действительно, у всех изученных самцов мы выявили модификации стереотипных линейных последовательностей песен, обусловленные добавлением/пропуском какой-то одной песни. Например, в одних случаях самец может исполнять последовательность типов песен 1–2–3–4–5, в других: 1–2–3–5 и т.п.
Однако мы выявили и другой тип изменений в организации пения. Речь идет о добавлениях/пропусках целых песенных последовательностей из 5–15 типов песен. Такой тип изменений был выявлен хотя и не у всех, но у большинства самцов. Мы предполагаем следующий механизм таких модификаций. Известно, что для развития нормальной дефинитивной песни певчим птицам требуется вокальное обучение: молодые копируют песни своих “учителей”, т.е. более старших самцов (Catchpole, Slater, 2008). Бывает, что неопытные птицы запоминают не отдельные песни, а целые их последовательности и/или наборы. Так, молодые южные соловьи (Luscinia megarhynchos) заучивают несколько (2–4) типов песен в ассоциативной связи друг с другом: такие ассоциированные песни формируют в репертуаре особи так называемые “упаковки” (packages: Todt, Hultsch, 1998). Нечто подобное может иметь место и у голосистой пеночки. Можно предположить, что все типы песен из репертуаров самцов этого вида распадаются на “упаковки” из нескольких (5–15) типов песен. При пении песни каждой “упаковки” исполняются в линейной последовательности. А последовательность из нескольких “упаковок”, формирующих репертуар особи, в свою очередь тоже линейна (этим и достигается линейный синтаксис всего пения). При этом в пении возможны случайные (предположительно) модификации, связанные с добавлением/пропуском одной или другой “упаковки” или “упаковок” .
В заключение отметим, что мы не выявили связей между изменчивостью пения самцов в разные дни и числом их ближайших соседей и/или расстоянием до ближайшего соседа. Таким образом, характер выявленной изменчивости не зависел от других особей. На наш взгляд, это дополнительно свидетельствует, что описанная изменчивость носит скорее характер случайных модификаций, а не является каким-то особым механизмом социальной адаптации.
Список литературы
Антонов А.И., Квартальнов П.В., 2014. Уточнение списка гнездящихся птиц Хинганского заповедника // Амурский зоологический журнал. Т. VI. С. 85–87.
Горецкая М.Я., 2013. Изменчивость тонкой структуры песни у воробьиных птиц: случайные вариации или направленные изменения, несущие информацию // Зоологический журнал. Т. 92. № 6. С. 718–730.
Иваницкий В.В., Марова И.М., Малых И.М., 2011. Контрасты синтаксиса в рекламном пении близких видов пеночек (Phylloscopus, Sylviidae) // Доклады Академии наук. Федеральное государственное унитарное предприятие Академический научно-издательский, производственно-полиграфический и книгораспространительский центр “Наука”. Т. 438. № 4. С. 570–573.
Иваницкий В.В., 2015. Песня птиц как микрокосм современной науки: между биоакустикой и лингвистикой // XIV орнитологическая конференция Северной Евразии. II. Доклады. Алматы. С. 418–435.
Иваницкий В.В., Марова И.М., 2010. К проблеме вокальной памяти у певчих птиц: сверхсложные акустические конструкции в песне корольковой пеночки (Phylloscopus proregulus) // Доклады Академии наук. Федеральное государственное унитарное предприятие Академический научно-издательский, производственно-полиграфический и книгораспространительский центр “Наука”. Т. 432. № 3. С. 426–429.
Опаев А.С., Колесникова Ю.А., 2019. Роль частоты исполнения и сложности пения в территориальном поведении голосистой пеночки (Рhylloscopus schwarzi) // Зоологический журнал. Т. 98. № 3. С. 319–331.
Baselga A., Orme D., 2012. betapart: an R package for the study of beta diversity // Methods in Ecology and Evolution. V. 3. P. 808–812.
Baselga A., Orme D., Villeger S., De Bortoli J., Leprieur F., 2017. Partitioning beta diversity into turnover and nestedness. https://cran.r-project.org/web/packages/betapart
Catchpole C.K., Slater P.J.B., 2008. Bird song: biological themes and variations. Cambridge: Cambridge University Press. 335 p.
Geberzahn N., Hultsch H., 2003. Long–time storage of song types in birds: evidence from interactive playbacks // Proceedings of the Royal Society of London. Series B: Biological Sciences. V. 270. № 1519. P. 1085–1090.
Ivanitskii V.V., Ivlieva A., Gashkov S., Marova I., 2017. The song structure of the Siberian Blue Robin Luscinia [Larvivora] cyane and a comparison with related species // Ornithological Science. V. 16. P. 71–77.
Kershenbaum A., Blumstein D.T., Roch M.A., Akcay C., Backus G. et al., 2014. Acoustic sequences in non-human animals: a tutorial review and prospectus // Biological Reviews of Cambridge Philosophical Society. V. 91. P. 13–52.
Kipper S., Kiefer S., 2010. Age-related changes in birds’ singing styles // Advances in the Study of Behavior. V. 41. P. 77–118.
Nelson D.A., 1992 Song overproduction and selective attrition lead to song sharing in the field sparrow (Spizella pusilla) // Behavioral Ecology and Sociobiology. V. 30. № 6. P. 415–424.
Okanoya K., 2013. Finite–state song syntax in Bengalese finches: sensorimotor evidence, developmental processes, and formal procedures for syntax extraction // Bolhuis J.J., Everaert M., eds. Birdsong, speech, and language. Exploring the evolution of mind and brain. London. P. 229–242.
Opaev A., 2016. Relationships between repertoire size and organization of song bouts in the Grey-crowned Warbler (Seicercus tephrocephalus) // Journal of Ornithology. V. 157. P. 949–960.
R Core Team, 2016. R: A language and environment for statistical computing. Vienna: R Foundation for Statistical Computing. https://www.r-project.org
Scharff C., Nottebohm F., 1991. A comparative study of the behavioral deficits following lesions of various parts of the zebra finch song system: implications for vocal learning // Journal of Neuroscience. V. 11. № 9. P. 2896–2913.
Spedicato G.A., Kang T.S., Yalamanchi S.B., Thoralf M., Yadav D. et al., 2017. Easy handling discrete time Markov chains. https://cran.r-project.org/web/packages/markovchain
Todt D., Hultsch H., 1998. Hierarchical learning, development and representation of song // Animal Cognition in Nature. Academic Press. P. 275–303.
Дополнительные материалы отсутствуют.
Инструменты
Зоологический журнал