

Зоологический журнал, 2020, T. 99, № 5, стр. 586-598
Динамика видовой структуры сообщества раковинных амеб в ходе сукцессии “водоем–болото” в голоцене на примере болота Мочуля (Калужская область, Россия)
А. Н. Цыганов a, b, *, А. А. Комаров b, **, Н. Г. Мазей a, ***, Т. В. Борисова b, ****, Е. Ю. Новенко a, c, *****, Ю. А. Мазей a, ******
a Московский государственный университет им. М.В. Ломоносова
119991 Москва, Россия
b Пензенский государственный университет
440026 Пенза, Россия
c Институт географии РАН
109017 Москва, Россия
* E-mail: andrey.tsyganov@bk.ru
** E-mail: a1ek.89@yandex.ru
*** E-mail: natashamazei@mail.ru
**** E-mail: toma.borisova.1994@mail.ru
***** E-mail: lenanov@mail.ru
****** E-mail: yurimazei@mail.ru
Поступила в редакцию 02.09.2019
После доработки 10.10.2019
Принята к публикации 20.10.2019
Аннотация
Раковинные амебы широко используются в палеоэкологических исследованиях в качестве индикаторов поверхностной влажности болот и состояния пресноводных экосистем. Цель настоящей работы – выявление закономерностей динамики видовой структуры сообществ раковинных амеб в ходе сукцессии “водоем–болото” по результатам комплексного анализа отложений болота Мочуля (Калужская область, Россия). Для объяснения изменения видового состава раковинных амеб использовали данные по ботаническому составу, степени гумификации и зольности торфа, а также результаты радиоуглеродного и палинологического анализов. Залежь сформирована отложениями водоема, низинного и переходного болот. Возраст отложений в основании составляет 4100 кал. л. н. Динамика видовой структуры сообществ раковинных амеб в значительной мере определяется локальной растительностью, в особенности сфагновыми мхами, которые существенно преобразуют среду. Показано, что превращение водоема в низинное болото, вероятнее всего, произошло в результате действия климатических (аллогенных) факторов: кратковременное иссушение и пожары, приведшие к зарастанию берегов. Трансформация низинного болота в переходное может быть связана как с аутогенными факторами (накопление отложений), так и с аллогенными (снижение активности человека на прилегающей территории).
Трансформация небольших лентических водных экосистем (водоемов) в болотные является закономерным процессом их развития, приводящим к существенному изменению их видового состава и функционирования. При появлении болотной растительности дальнейшая сукцессия обычно направлена на формирование более плотного растительного покрова и торфяной залежи, снижение поверхностной влажности болота, повышение содержания органического вещества и снижение зольности отложений (Ryding, Juglum, 2006). Результат сукцессии зависит от локальных гидрологических и климатических условий и может приводить к формированию низинного болота, заболоченного леса или верхового болота (Волкова, 2018). В целом, генезис водно-болотных экосистем определяется сочетанием как аллогенных (внешних), так и автогенных (внутренних) факторов (Сукачев, 1926), при этом роль этих факторов на разных этапах развития и в разных экосистемах различна. Понимание закономерностей подобных сукцессий необходимо для прогнозирования развития водных и болотных экосистем в нестабильных условиях окружающей среды и возрастающей антропогенной нагрузки.
Для изучения сукцессии водных и болотных экосистем может быть использован палеоэкологический подход, основанный на изучении отложений, в которых содержатся физические, химические и биологические индикаторы условий их формирования. Широкий набор используемых в настоящее время индикаторов обусловлен различиями в их чувствительности к факторам среды, скоростью реагирования в ответ на изменение характеристик и др. (Chambers et al., 2012). Реконструкции генезиса и динамики болот базируются, преимущественно, на результатах анализа ботанического состава торфяных отложений, особенно в сочетании со спорово-пыльцевым анализом для реконструкции динамики прилегающих ландшафтов и климата. Несмотря на важность информации, полученной с использованием этих методов, она может быть существенно дополнена данными о поверхностной влажности болот, которая в значительной степени определяет их гидрологический режим, а также, соответственно, генезис и динамику. В качестве индикаторов поверхностной влажности могут быть использованы зоологические объекты и физико-химические характеристики торфа, такие как степень его гумификации (Mitchell et al., 2008; Chambers et al., 2012). Использование дополнительных индикаторов позволяет получить новую информацию и повысить достоверность палеоэкологических реконструкций.
Раковинные амебы (корненожки или ризоподы) – амебоидные протисты, которые характеризуются наличием внешнего скелетного образования (раковинки) (Мазей, Цыганов, 2006). Основная функция раковинки – защита от иссушения и поедания хищниками. Размер раковинки у большинства видов изменяется в пределах от 20 до 150 мкм. Филогенетически, раковинные амебы относятся как минимум к трем неродственным группам организмов: Euglyphida, Arcellinida и Amphitrematida (Gomaa et al., 2013). Однако для целей биоиндикации и палеоэкологии они рассматриваются совместно, что обусловлено сходством их экологии и методов исследования. Раковинные амебы преимущественно гетеротрофные организмы, питающиеся бактериями, грибами, другими простейшими и водорослями, однако в некоторых случаях способны атаковать объекты намного крупнее них, например, мелких многоклеточных (Gilbert et al., 2000). Некоторые виды содержат в цитоплазме симбиотические водоросли и являются миксотрофами. Раковинные амебы распространены практически повсеместно и являются важным компонентом пресноводных, болотных и почвенных экосистем (Mitchell et al., 2008). Они очень чувствительны к степени увлажнения, кислотности и некоторым другим характеристикам окружающей среды и быстро реагируют на изменения локальных условий (Гельцер и др., 1985). В настоящее время известно около 3000 видов раковинных амеб, описанных преимущественно на основе морфологических критериев раковинок. При этом сами раковинки устойчивы к разложению и формируют обильные скопления в торфяных залежах болот и донных отложениях водоемов. Благодаря сочетанию вышеперечисленных признаков, раковинные амебы являются ценным биоиндикатором для палеоэкологических исследований, на основе чего был разработан метод – ризоподный анализ (Harnisch, 1925; Tolonen, 1966; Warner, Charman, 1994). Анализ степени гумификации торфа часто используется совместно с ризоподным анализом для реконструкции поверхностной влажности болот (Chambers et al., 2010). Данный метод основан на предположении о том, что в теплых и сухих условиях гумификация мертвого органического вещества происходит быстрее, чем в прохладных и влажных.
Несмотря на большое число палеоэкологических реконструкций, основных на анализе торфяных залежей, вопросам сукцессионной динамики и закономерностей смены озерно-болотных систем уделяется недостаточно внимания (Бобров, 2003; Elliott et al., 2012; Sonnenburg et al., 2013). Особый интерес представляют болотные экосистемы, расположенные на южной границе своего распространения (в зоне широколиственных лесов и лесостепи), так как именно они наиболее чувствительны к изменению климатических условий. Цель настоящей работы – выявление закономерностей динамики видовой структуры сообществ раковинных амеб в ходе сукцессии “водоем – болото” по результатам комплексного анализа отложений болота Мочуля (Калужская обл., Европейская часть России). Для объяснения изменений видового состава раковинных амеб использовали данные по ботаническому составу, степени гумификации и зольности торфа, а также результаты радиоуглеродного и спорово-пыльцевого анализов, опубликованные ранее (Novenko et al., 2019) и интерпретированные в контексте динамики климата, ландшафтов и активности человека. В данной работе мы приводим новые данные по видовому составу раковинных амеб и степени гумификации торфа, чтобы реконструировать динамику локальной влажности болотной экосистемы и понять закономерности сукцессии “водоем–болото”.
РАЙОН ИССЛЕДОВАНИЙ
Исследования проводили на территории государственного природного заповедника “Калужские засеки” (Калужская область, Европейская часть России). Рассматриваемый район расположен на западной периферии Среднерусской возвышенности, в междуречье рек Вытебети (правый приток р. Жиздры), Нугри и Оки. Он входит в Среднерусскую ландшафтную провинцию широколиственных (Гвоздецкий, Жучкова, 1963) или смешанных (Мильков, Гвоздецкий, 1976) лесов и приурочен к её южной границе. Для рельефа территории характерны пологохолмистые моренные возвышенности с развитой эрозионной сетью и водно-ледниковые (зандровые) равнины.
Климат региона исследования умеренно континентальный (среднегодовая температура + 4.8°С) с холодными зимами (средняя температура января – 10.1°С, средняя температура июля +17.5°С). Годовое количество осадков изменяется в пределах от 360 до 1000 мм со средним значением 630 мм (Калужская метеостанция, http://meteo.ru/ data). Растительность региона представлена преимущественно широколиственными лесами, в которых преобладают Quercus robur L. 1753, Fraxinus excelsior L. 1753, Tilia cordata, Mill. 1768, Ulmus glabra Huds., Acer platanoides L. 1753 и A. campestre L. 1753 с примесью Populus tremula L. 1753 и Betula verrucosa Roth (Smirnova et al., 2017). В кустарничковом ярусе преобладает Corylus avellana (L.) H. Karst. 1881, достаточно часто встречаются Euonymus europaea L. 1753, E. verrucosa Scop. (1771), Lonicera xylosteum L. 1753 и Padus avium L. 1753. Флювиогляциальные равнины и ложбины, сложенные преимущественно песками, заняты еловыми и елово-сосновыми лесами с примесью широколиственных пород деревьев, а также C. avellana, Frangula alnus Mill. 1768 и Sorbus aucuparia в подлеске.
Объект исследования – болотная экосистема Мочуля (53.55247° с.ш., 35.7522° в.д.), расположенная в обширной древней ложбине стока ледниковых вод. Болото ориентировано в направлении с юго-запада на северо-восток и имеет вытянутую форму (длина 400 м, ширина 90–120 м, общая площадь около 4.5 га). Водное питание болота осуществляется за счет небольшого ручья. Растительный покров болота сформирован порослью березы (Betula sp.) высотой до 2 м и осоками Carex spp. в травяном ярусе. На открытых участках развит плотный покров из сфагновых мхов (Sphagnum spp.). Также обильно представлены Calla palustris L., Calamagrostis canescens Roth и Lysimachia thyrsiflora L. Край болота и влажные почвы ложбины покрыты заболоченным лесом, сформированным преимущественно Alnus glutinosa (L.) Gaertn. 1791. Толщина торфяных отложений изменяется от 40 до 70 см в самой глубокой северной части болота. В целом современное состояние болота может быть оценено как мезотрофное. Болото окружено еловым лесом (Picea abies (L.) H. Karst. 1881) c примесью березы, куртинами папоротника орляка обыкновенного (Pteridium aquilinum (L.) Kuhn 1879) в травяном ярусе и разреженным моховым покровом.
МАТЕРИАЛ И МЕТОДЫ
Отложения торфа извлекали с использованием ручного бура (длина пробоотборника 50 см, диаметр 5 см; Eijkelkamp 0409, Нидерланды) в августе 2016 г. Придонные отложения на глубине 90–70 см были сформированы гиттией, а верхние горизонты (70–0 см) – торфом. Остатки растений в гиттии были немногочисленными и представлены преимущественно осоками и рогозом, сфагновыми мхами, а также фрагментами коры сосны, березы и ивы (Novenko et al., 2019). Торфяные отложения были сформированы низинным (эвтрофным) торфом на глубинах 70–37 см и сфагновым мезотрофным торфом в верхних горизонтах на глубинах 37–0 см. Отобранные торфяные колонки разрезаны на последовательные слои толщиной 1–2 см для проведения ризоподного анализа, а также анализа потерь при прокаливании и степени гумификации торфа.
Для определения возраста торфяной залежи отобрали пять образцов торфа (табл. 1). Радиоуглеродное датирование выполнено в Институте геологии и геохимии НАН Беларуси (Минск) с использованием жидкостных сцинтилляционных счетчиков активности углерода. Калибровку радиоуглеродных дат осуществляли с помощью программы clam2.1 (Blaauw, 2010) и калибровочной кривой IntCal13 (Reimer et al., 2013). Новенко с соавторами (Novenko et al., 2019) для построения модели возраст‑глубина была использована простая линейная интерполяция между калиброванными датами. В настоящей работе мы использовали байесовский подход, который обладает большей гибкостью и способен учитывать вероятностный характер датировок с помощью радиоуглеродного метода. Модель возраст–глубина (рис. 1) была построена с помощью пакетного приложения ‘rbacon’ (Blaauw, Christen, 2019) в среде программирования R (R Core Team, 2018). В качестве априорных значений скорости аккумуляции торфа использовали 50 лет см–1, другие параметры модели были использованы с предустановленными значениями (Goring et al., 2012). В тексте статьи используются данные калиброванного возраста (с вероятностью 2σ), выраженные в годах от настоящего времени – лет назад (л. н.).
Таблица 1.
Результаты радиоуглеродного датирования торфяной залежи болота Мочуля (Калужская обл.)
| № | Глубина, см | Код образца | Радиоуглеродный возраст (14С л. н.) | Калиброванный возраст (2σ), кал. л. н. |
|---|---|---|---|---|
| 1 | 27–30 | IGSB-1703 | 120 ± 40 | 10–150 (59.2%) |
| 173–179 (1.5%) | ||||
| 182–274 (34.2%) | ||||
| 2 | 40–42 | IGSB-1704 | 810 ± 60 | 663–803 (84.1%) |
| 810–829 (2.7%) | ||||
| 858–904 (8.1%) | ||||
| 3 | 51–55 | IGSB-1705 | 1240 ± 80 | 983–1034 (7.6%) |
| 1045–1295 (87.3%) | ||||
| 4 | 70–73 | IGSB-1706 | 2970 ± 150 | 2787–3451 (95%) |
| 5 | 85–90 | IGSB-1707 | 3780 ± 90 | 3926–3952 (2.2%) |
| 3958–4417 (92.8%) |

Подготовку образцов торфа для ризоподного анализа проводили по методу, основанному на фильтровании и концентрировании водных суспензий (Мазей, Ембулаева, 2009). Видовую идентификацию и учет раковинных амеб проводили методом прямого микроскопирования при увеличении 200× и 400×. При видовой идентификации использовали руководство (Мазей, Цыганов, 2006). В каждой пробе, за исключением проб с очень низким содержанием раковинок, насчитывали не менее 150 амеб. Анализ потерь при прокаливании торфа проводили путем прокаливания 50 г образца при 550°С (Dean, 1974). Степень гумификации торфа определяли с помощью щелочной экстракции гуминовых кислот и последующим измерением оптической плотности раствора (D) спектрофотометрическим методом при длине волны 540 нм (фотометр КФК 3–01 “ЗОМЗ”, Россия) (Chambers et al., 2010). Полученные данные по степени гумификации торфа были преобразованы согласно уравнению D' = D/LOI, где LOI – значение потерь при прокаливании, выраженное в долях от целого. Данное преобразование позволяет ввести поправку на зольность образца, которая снижает значения гумификации за счет более низкой доли органического вещества в образце.
Подготовку образцов для анализа ботанического состава торфа и видовую идентификацию растительных макроостатков проводили согласно общепринятым методикам (Тюремнов, 1959). Образцы для спорово-пыльцевого анализа подготавливали стандартным способом (Moore et al., 1991). Подсчет и определение спор и пыльцевых зерен (Reille, 1992; Beug, 2004) осуществляли методом прямого микроскопирования (увеличение ×400).
Анализ и визуализация данных выполнялись с помощью языка программирования R (R Core Team, 2018) и пакетных приложений “analogue” (Simpson, Oksanen, 2018), “rioja” (Juggins, 2017) и “vegan” (Oksanen et al., 2019). Для выделения однородных зон торфяной залежи по видовой структуре сообществ раковинных амеб использовали последовательный кластерный анализ (Grimm, 1987). Для сопоставления результатов ризоподного анализа с данными по ботаническому составу торфа и спорово-пыльцевому анализу (детально представленными в работе Новенко и др. (Novenko et al., 2019)) проводили неметрическое многомерное шкалирование (НМШ) на основании матрицы несходства сообществ по индексу Брея-Кёртиса (Bray, Curtis, 1957). НМШ – метод ординации, в результате которого объекты (в данном случае палеосообщества) визуализируются в пространстве меньшей размерности, чем пространство их признаков. Полученные координаты проекций сообществ на плоскость двух первых факторов ординации использовали для анализа общих закономерностей динамики в их видовой структуре. Количественную оценку взаимосвязи между координатами осей проводили с помощью корреляционного анализа с использованием непараметрического коэффициента корреляции Спирмена (rho).
РЕЗУЛЬТАТЫ
Слой гиттии (90–70 см, 4100–2970 кал. л. н.) характеризовался низкими значениями потерь при прокаливании (рис. 2А), что указывает на высокое содержание минеральной фракции в этом горизонте. Затем содержание органического вещества резко увеличивается, что связано с началом формирования торфяных отложений в депрессии и остается высоким по всей глубине за исключением слоев 46–30 см (920–280 кал. л. н.), где оно было ниже (74–82%). Это может указывать на активную деятельность человека на прилегающей территории, приведшей к вырубке лесов и увеличению поверхностной эрозии.
Рис. 2.
Физико-химические характеристики торфяной залежи болота Мочуля: A – потери при прокаливании; B – степень гумификации, измеренная как оптическая плотность щелочной вытяжки (сплошная линия – измеренные значения, пунктирная – с поправкой на потери при прокаливании).
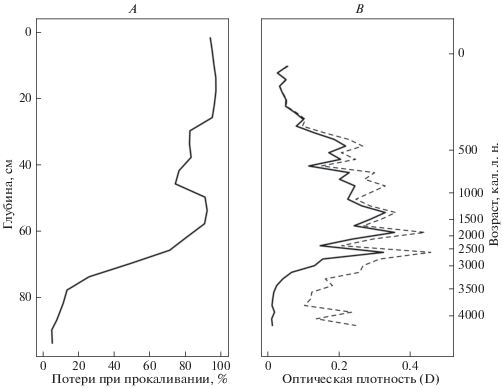
Гумификация торфа (рис. 2B) была низкой в основании торфяной залежи (90–72 см; 4100–3120 кал. л. н.), что вероятно свидетельствует о благоприятных условиях накопления мертвого органического вещества на дне водоема. В вышележащих горизонтах (72–35 см; 3120–500 кал. л. н.) степень гумификации увеличивается, что может быть связано со сменой характера разложения органического вещества в болотной экосистеме, по сравнению с водной (т.е. более высокие температуры и аэрация на поверхности болота, по сравнению с придонными горизонтами водоема). Наиболее высокие значения в начале зоны могут указывать на пересыхание болота или выгорание. В верхних горизонтах (35–0 см, 500 кал. л. н.–настоящее время) степень гумификации существенно снижается, что связано с благоприятными условиями торфонакопления (высокая влажность, низкие температуры, сниженная активность бактерий в результате преобладания сфагнумов в растительном покрове).
В результате ризоподного анализа обнаружено 54 вида и подвида раковинных амеб, относящихся к 21 роду (табл. 2). Всего было учтено 5077 раковинок, при этом в нижних горизонтах 95–75 см раковинные амебы отсутствовали полностью, на глубинах 75–70 см найдены лишь единичные особи (не более двух в образце), а в вышележащих горизонтах (70–0 см) численность раковинных амеб была достаточной для анализа видовой структуры сообщества. Наиболее обильными видами были Trinema lineare (23.2%, здесь и далее от общего числа учтенных особей), Hyalosphenia papilio (7.7%), Cyclopyxis eurystoma (6.7%), Alabasta militaris (6.6%), Centropyxis aculeata (5.1%), Hyalosphenia elegans (5.1%) и Argynnia dentistoma (5.0%). Наиболее встречаемые видами, обнаруженными более чем в 60% всех образцов, были H. papilio, T. lineare, Assulina muscorum, Centropyxis ecornis и A. militaris. Шесть видов (Corythion orbicularis, Euglypha cristata, Euglypha simplex, Heleopera sylvatica, Lesquereusia modesta, Nebela minor) были обнаружены лишь в одном образце с относительным обилием, не превышающим 1.2%. Среднее число видов в пробе составляло 13.6 ± 9.0 (стандартное отклонение) и изменялось в диапазоне от 0 до 31. В целом видовой состав залежи является типичным для водных и болотных экосистем и отражает сукцессионную динамику водно-болотной экосистемы.
Таблица 2.
Список видов раковинных амеб, обнаруженных в торфяной залежи болота Мочуля (Калужская обл.)
| № | Вид | Относительное обилие, % от общего числа обнаруженных особей | Встречаемость, % от общего числа проб | Максимальное относительное обилие, % от общего числа особей в пробе |
|---|---|---|---|---|
| 1 | Alabasta militaris (Penard 1890) Duckert et al. 2018 | 6.6 | 60 | 32.5 |
| 2 | Arcella arenaria Greeff 1866 | 0.2 | 16 | 5.7 |
| 3 | Arcella arenaria sphagnicola Deflandre 1928 | 2.2 | 47 | 22.4 |
| 4 | Arcella catinus Penard 1890 | 1.0 | 31 | 6.7 |
| 5 | Archerella flavum (Archer 1877) Loeblich and Tappan 1961 | 0.4 | 11 | 5.8 |
| 6 | Argynnia dentistoma (Penard 1890) | 5.0 | 47 | 23.7 |
| 7 | Argynnia dentistoma lacustris Wailes 1912 | 0.5 | 9 | 10.5 |
| 8 | Assulina muscorum Greeff 1866 | 3.3 | 64 | 11.6 |
| 9 | Assulina seminulum (Ehrenberg 1848) Leidy 1879 | 0.3 | 22 | 1.3 |
| 10 | Centropyxis aculeata (Ehrenberg 1838) Stein 1857 | 5.1 | 58 | 20.6 |
| 11 | Centropyxis aerophila Deflandre 1929 | 4.0 | 53 | 33.3 |
| 12 | Centropyxis aerophila sphagnicola Deflandre 1929 | 0.8 | 22 | 5.8 |
| 13 | Centropyxis cassis (Wallich 1864) Deflandre 1929 | 0.1 | 9 | 0.7 |
| 14 | Centropyxis constricta (Ehrenberg 1841) Deflandre 1929 | 0.1 | 4 | 1.9 |
| 15 | Centropyxis ecornis (Ehrenberg 1841) Leidy 1879 | 3.2 | 62 | 14.6 |
| 16 | Centropyxis elongata (Penard 1890) Thomas 1959 | 3.9 | 11 | 83.3 |
| 17 | Centropyxis laevigata Penard 1890 | 0.1 | 4 | 1.2 |
| 18 | Centropyxis minuta Deflandre 1929 | 0.6 | 16 | 5.3 |
| 19 | Centropyxis orbicularis Deflandre 1929 | 0.2 | 18 | 16.7 |
| 20 | Corythion dubium Taránek 1881 | 0.2 | 16 | 2.0 |
| 21 | Corythion orbicularis (Penard 1890) Iudina 1996 | 0.1 | 2 | 0.7 |
| 22 | Cyclopyxis arcelloides (Penard 1902) Deflandre 1929 | 3.3 | 53 | 20.7 |
| 23 | Cyclopyxis eurystoma Deflandre 1929 | 6.7 | 38 | 50.0 |
| 24 | Cyclopyxis kahli Deflandre 1929 | 1.1 | 29 | 50.0 |
| 25 | Difflugia globulosa Dujardin 1837 | 0.8 | 20 | 9.0 |
| 26 | Euglypha ciliata glabra (Ehrenberg 1848) Leidy 1878 | 0.6 | 29 | 5.8 |
| 27 | Euglypha cristata Leidy 1879 | 0.1 | 2 | 1.2 |
| 28 | Euglypha cristata major Wailes 1912 | 0.6 | 11 | 9.2 |
| 29 | Euglypha rotunda Wailes 1915 | 0.6 | 33 | 4.6 |
| 30 | Euglypha scutigera Penard 1911 | 0.2 | 13 | 2.7 |
| 31 | Euglypha simplexi Decloitre 1965 | 0.1 | 2 | 0.6 |
| 32 | Euglypha strigosa glabra Wailes 1898 | 0.2 | 16 | 3.2 |
| 33 | Euglypha tuberculata Dujardin 1841 | 0.3 | 16 | 2.7 |
| 34 | Heleopera petricola Leidy 1879 | 0.3 | 22 | 2.9 |
| 35 | Heleopera sphagni Leidy 1874 | 0.2 | 13 | 2.0 |
| 36 | Heleopera sylvatica Penard 1890 | 0.1 | 2 | 0.6 |
| 37 | Hyalosphenia elegans Leidy 1879 | 5.1 | 49 | 33.7 |
| 38 | Hyalosphenia papilio Leidy 1879 | 7.7 | 71 | 34.7 |
| 39 | Lesquereusia modesta Rhumbler 1895 | 0.1 | 2 | 0.7 |
| 40 | Nebela collaris (Ehrenberg 1848) Kosakyan et Gomaa 2013 | 3.1 | 58 | 11.4 |
| 41 | Nebela minor Penard 1902 | 0.1 | 2 | 1.2 |
| 42 | Nebela tincta (Leidy 1879) Kosakyan et Lara 2012 | 1.3 | 40 | 8.0 |
| 43 | Phryganella acropodia Hopkinson 1909 | 0.1 | 4 | 2.0 |
| 44 | Physochilla griseola (Wailes et Penard 1911) Jung 1942 | 0.1 | 9 | 1.2 |
| 45 | Physochilla tenella (Ehrenberg 1871) Jung 1942 | 0.2 | 13 | 1.3 |
| 46 | Plagiopyxis sp. | 0.4 | 16 | 4.5 |
| 47 | Pseudodifflugia gracilis Schlumberger 1845 | 2.3 | 27 | 18.7 |
| 48 | Pseudodifflugia gracilis terricola Bonnet et Thomas 1960 | 0.7 | 24 | 7.2 |
| 49 | Sphenoderia fissirostris Penard 1890 | 0.1 | 4 | 3.3 |
| 50 | Trigonopyxis arcula (Leidy 1879) Penard 1912 | 1.1 | 22 | 11.1 |
| 51 | Trinema complanatum Penard 1890 | 1.2 | 36 | 5.8 |
| 52 | Trinema enchelys (Ehrenberg 1838) Leidy 1878 | 0.1 | 11 | 1.3 |
| 53 | Trinema lineare Penard 1890 | 23.2 | 69 | 100.0 |
| 54 | Trinema penardi Thomas et Chardez 1958 | 0.7 | 16 | 16.9 |
По результатам кластерного анализа видовой структуры сообществ раковинных амеб в исследуемой залежи можно выделить пять зон (рис. 3): 90–70 см (4100–2970 кал. л. н.), 70–61 см (2970–1930 кал. л. н.), 61–42 см (1930–770 кал. л. н.), 42–24 см (770–200 кал. л. н.), 24–0 см (200 кал. л. н.–настоящее время).
Рис. 3.
Диаграмма видового состава сообществ раковинных амеб в торфяной залежи болота Мочуля и зонирование по результатам последовательно кластерного анализа. Показаны только наиболее обильные виды, обнаруженные как минимум в трех образцах с относительным обилием 3% и более. РА1–РА5 – зоны с однородным видовым составом раковинных амеб.

В отложениях, сформированных на стадии водоема (Зона РА1, 90–70 см, 4100–2970 кал. л. н.), раковинные амебы отсутствовали полностью либо были представлены единичными раковинами, относящимися к видам T. lineare, Cyclopyxis kahli, C. eurystoma. Эти виды характеризуются различными экологическими предпочтениями и являются эврибионтными или почвенными, что может указывать на неглубокий зарастающий водоем, возможно с сильной эрозией на водосборе или часто пересыхающий.
Зона РА2 (70–61 см, 2970–1930 кал. л. н.) характеризовалась преобладанием пресноводных (Centropyxis elongata 43%, здесь и далее от общего числа особой, обнаруженных в данной зоне) и эврибионтных (C. eurystoma 19%, Centropyxis aerophila 15%) видов, характерных для зарастающих водоемов или сильно обводненных низинных болот.
Зона РА3 (61–42 см, 1930–770 кал. л. н.) характеризуется более высоким видовым разнообразием раковинных амеб по сравнению с предыдущей. В сообществах преобладают эврибионтные виды (C. eurystoma 19%, T. lineare 15%, Pseudodifflugia gracilis 8%), а также присутствуют водные (C. aculeata 7%) и сфагнобионтные (H. papilio 7%, C. aerophila 6%, A. muscorum 5%) представители. Подобный видовой состав может свидетельствовать о высокой пространственной неоднородности болота (наличие низинных участков наряду с биотопами с преобладанием сфагнума различной степени увлажненности) либо о нестабильном гидрологическом режиме.
В зоне РА4 (42–24 см, 770–200 кал. л. н.) видовая структура сообществ изменяется и существенную долю занимает эврибионтный вид T. lineare (60%). В сообществах также присутствуют водные (Cyclopyxis arcelloides 7%, Centropyxis ecornis 4%), а также сфагнобионтные (Arcella arenaria sphagnicola 5%) виды. Существенная доля эврибионтного вида, вероятнее всего, характеризует засушливые периоды или периоды нарушения экосистемы.
Нижняя граница зоны РА5 (24–0 см, 200 кал. л. н.– настоящее время) маркирует существенные изменения в видовой структуре доминантного комплекса. В сообществе преобладают сфагнобионтные виды (A. militaris 16%, H. papilio 14%, Hyalosphenia elegans 13%, A. dentistoma 12%, Nebela collaris 7%), при сохранении незначительной доли водных (C. aculeata 8%) и эврибионтных (T. lineare 6%) видов. Такая видовая структура сообществ раковинных амеб характерна для сфагновых болот с влажной поверхностью.
На основании результатов анализа ботанического состава (БС) и спорово-пыльцевого (СП) анализа в торфяной залежи болота выделено шесть зон, подробное описание которых приводится в работе Новенко с соавторами (Novenko et al., 2019). Для сопоставления этих данных с динамикой сообществ раковинных амеб на рис. 4 представлено их краткое описание и результаты неметрического многомерного шкалирования. Первая ось ординации данных по ботаническому составу торфа (рис. 4A) отображает четкие различия между эвтрофным (зоны БС1–БС3) и мезотрофным болотом с преобладанием сфагнума (БС5–БС6). Вторая ось ординации отражает изменения в субдоминантном комплексе с низкими значениями в зоне БС1, в которой отмечено значительное присутствие остатков древесных растений, в то время как положительные значения маркируют изменчивость видового состава субдоминантных видов сфагновых мхов в покрове (зона БС6).
Рис. 4.
Координаты образцов по результатам неметрического многомерного шкалирования (НМШ) матриц несходства Брея-Кёртиса: A – ботанический состав торфа, B – раковинные амебы, С – пыльца наземных растений. Координаты по оси OX показаны сплошной линией, ось OY – пунктирной. Зонирование залежи по данным ботанического состава торфа (БС1–БС6) и спорово-пыльцевого анализа (СП1–СП6) приведено согласно Новенко и др. (Novenko et al., 2019). РА1–РА5 – зоны с однородным видовым составом раковинных амеб.
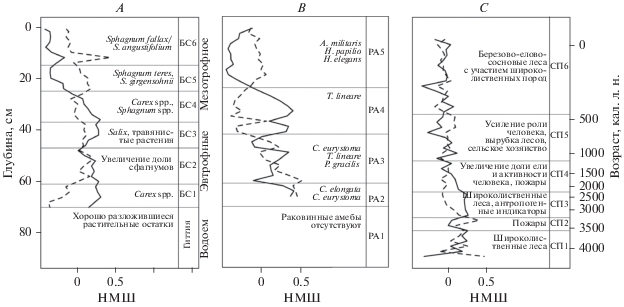
По результатам анализа видовой структуры сообществ раковинных амеб (рис. 4B) изменчивость вдоль первой оси ординации соответствует изменениям в составе доминантного комплекса видов. Зона с преобладанием T. lineare (РА4) четко выделяется по высоким значениям координат. Низкие значения координат отмечают зоны с преобладанием сфагнобионтных видов. Четкая смена структуры сообществ отмечена на нижней границе зоны РА5, где изменяются и доминантный, и субдоминантный комплексы.
Первая ось ординации по данным спорово-пыльцевого анализа (рис. 4C) отображает нарушения в составе преобладающих видов растений на региональном уровне (положительные значения соответствуют естественному растительному покрову, а отрицательные – нарушенному). Вторая ось отражает нарушения в естественном покрове, вызванные пожарами (положительные значения).
Результаты корреляционного анализа свидетельствуют о наличии сильной достоверной положительной взаимосвязи между видовой структурой локальной растительности и видовой структурой сообществ раковинных амеб (по первым осям ординации) (табл. 3). Это объясняется тем, что локальная растительность во многом определяет характер сообществ раковинных амеб, населяющих биотоп, при этом сфагновые биотопы характеризуются специфическим набором видов. Отмечается четкое совпадение границ по ботаническому составу торфа и видовому составу сообществ раковинных амеб при переходе от водоема к низинному болоту и от низинного болота к мезотрофному с преобладанием сфагнума. Достоверные положительные значения корреляции отмечаются между второй осью спорово-пыльцевого анализа и первыми осями ботанического состава торфа и видового состава раковинных амеб, а также между первой осью споро-пыльцевого анализа и второй осью видового состава раковинных амеб. Это может быть связано с тем, что пожары и региональные нарушения на водосборе (в том числе вызванные антропогенными причинами) влияют на болотную экосистему практически на всех уровнях – локальной растительности и микробного населения.
Таблица 3.
Коэффициент корреляции Спирмена (rho) между координатами сообществ в первых двух осях ординации (НМШ1 и НМШ2) по результатам неметрического многомерного шкалирования данных по ботаническому составу торфа, ризоподного и спорово-пыльцевого анализа
| Показатель | Ось ордина-ции | Ботанический состав | Раковинные амебы | Спорово-пыльцевой анализ | |||
|---|---|---|---|---|---|---|---|
| НМШ1 | НМШ2 | НМШ1 | НМШ2 | НМШ1 | НМШ2 | ||
| Ботанический состав | НМШ1 | – | ns | 0.74** | ns | ns | 0.56** |
| НМШ2 | – | ns | ns | ns | ns | ||
| Раковинные амебы | НМШ1 | – | ns | ns | 0.64** | ||
| НМШ2 | – | 0.41* | ns | ||||
| Спорово-пыльцевой анализ | НМШ1 | – | ns | ||||
| НМШ2 | – | ||||||
ОБСУЖДЕНИЕ
Полученные данные свидетельствую о том, что болото Мочуля в ходе своего развития прошло через классическую сукцессионную серию от мелководного водоема через низинное обводненное эвтрофное болото к мезотрофному болоту с преобладанием сфагнумов. Накопление органических отложений на дне небольшого мелкого водоема началось в середине суббореальной стадии голоцена (4100 кал. л. н.). В то время исследуемый водоем располагался в зоне широколиственных лесов (Novenko et al., 2019), широко простиравшейся в Европе. Незначительное присутствие остатков Carex spp., Typha latifolia, Sphagnum spp. совместно с остатками коры сосны, березы и ивы свидетельствует о том, что водоем был окружен типичной прибрежной растительностью и располагался в лесном массиве. К сожалению, раковинки амеб в отложениях, сформированных на стадии мелкого водоема, также немногочисленны, что может быть связано с неблагоприятными условиями для их захоронения (высокие температуры, аэрация, периодические пересыхания и т.д.) либо с преобладанием видов с неустойчивыми раковинками (Payne, 2010, 2011). Относительно низкие показатели степени гумификации отложений могут быть связаны с высокой долей остатков в торфе древесных растений, устойчивых к разложению.
Результаты спорово-пыльцевого анализа свидетельствуют о том, что на начальной стадии формирования отложений (3700–3300 кал. л. н.) в растительном покрове произошли существенные нарушения, вызванные с пожарами. Наиболее вероятной причиной этих преобразований стали климатические изменения, т.е. летний дефицит увлажненности, что привело к снижению поверхностного стока и скорости отложения донных осадков в водоеме (Novenko et al., 2019). Эти аллогенные факторы могли привести к заболачиванию водоема в конце суббореального периода (3000 кал. л. н.). В работах (Ireland, Booth, 2011; Booth et al., 2016) было показано, что снижение уровня водоемов может приводить к их резкому заболачиванию после возвращения к исходной обводненности, за счет затопления разросшейся прибрежной растительности (периферийное заболачивание). В целом, для водоемов, расположенных в лесных массивах, характерны интенсивные процессы заболачивания, так как поступление органического вещества в водоем вместе с опадающими листьями и ветками окружающих деревьев способствует формированию сплавины на его поверхности (Волкова, 2010). Однако в данном случае наиболее вероятно преобладание периферийного заболачивания с накоплением осадков на дне водоема.
Скорость аккумуляции торфа на начальной стадии формирования низинного болота (3000–2600 кал. л. н.) была низкой, что совместно с высокой концентрацией углей (Novenko et al., 2019) в этих слоях может указывать на перерыв (хиатус) в осадконакоплении и утрате части нижних горизонтов в результате выгорания. Максимальные значения степени гумификации торфа в начале зоны могут быть связаны с локальными пожарами. Сообщества раковинных амеб были сформированы видами C. elongatа, C.eurystoma и C.aerophila, которые широко представлены в зарастающих водоемах и сильно обводненных низинных болотах. Так, например, вид C. aerophila толерантен к низким значениям pH до 5.5 (Patterson et al., 2002). В целом данные виды являются оппортунистическими и способны переносить относительно низкие уровни содержания питательных элементов (Schönborn, 1984).
После трансформации водоема в низинное болото основными факторами его развития стали аутогенные процессы, связанные с накоплением торфяных отложений и возрастающей изоляцией от грунтовых вод. В составе локальной растительности это приводит к увеличению доли гипновых и сфагновых мхов (Sphagnum fallax/angustifolium) типичных для низинных эвтрофных и мезо-эвтрофных болот, а также закономерному снижению степени гумификации торфа. В составе сообществ раковинных амеб отмечается увеличение видового разнообразия, однако преобладающими видами остаются эврибионты C. eurystoma и T. lineare, которые являются типичными для низинных болот (Payne, 2011). Обычно в низинных болотах раковинные амебы плохо сохраняются по причине хрупкости раковин у видов, населяющих их, и более высокой степени разложения (Payne, 2011). Однако в исследованной колонке видовое разнообразие раковинных амеб на стадии низинного болота было достаточно высоким, что может объясняться присутствием сфагновых мхов в растительном покрове и, соответственно, сфагнобионтных раковинных амеб, которые более устойчивы к разложению.
Аутогенное развитие низинного болота было прервано внешним воздействием, связанным с повышением активности человека на прилегающей к болоту территории в период с 1160 по 450 кал. л. н. На региональном уровне это выразилось в сведении лесов и распашке территорий для сельского хозяйства (Novenko et al., 2019), что привело к повышенной эрозии на водосборе и увеличенному поступлению минеральных веществ в болото. В результате этого отмечено повышение зольности отложений (в период с 920 по 300 кал. л. н.), а также снижение доли сфагнума и проникновение ивы, березы и трав на болото (зона БС3). В видовой структуре сообществ раковинных амеб это выразилось в сверхдоминировании одного эврибионтного вида T. lineare и упрощении субдоминантного комплекса видов (зона РА4). Данный вид в контексте анализа сукцессионной динамики водно-болотных экосистем можно рассматривать как оппортунистический.
Еще одна важная смена в характере функционирования изучаемой болотной экосистемы отмечена 550 кал. л. н., когда антропогенная активность в регионе снизилась, в результате отнесения исследуемой территории к Засечной черте, а впоследствии заповедования. В локальной растительности болота начинает увеличиваться доля сфагнума, что указывает на активные процессы олиготрофизации. Основным фактором этого процесса могут быть как аутогенные (накопление торфяных отложений, что приводит к изменению микрорельефа и изоляции от грунтовых вод) и аллогенные факторы (снижение антропогенного воздействия и восстановление лесного покрова). Изменения в характере локальной растительности послужили толчком к изменению в видовой структуре сообществ раковинных амеб, а именно снижению доли T. lineare и увеличению доли сфагнобионтных видов. Подобные трансформации в видовом составе сообществ раковинных амеб были отмечены ранее при изучении торфяной залежи болота Старосельский мох в зоне средней тайги (Payne et al., 2015). Практически синхронные изменения в характере локального растительного покрова (преобладание сфагновых мхов) и в видовой структуре сообществ раковинных амеб (преобладание сфагнобионтов) происходят 200 кал. л. н. и характеризуют формирование современного мезотрофного сфагнового болота.
ЗАКЛЮЧЕНИЕ
Полученные результаты свидетельствуют о том, что динамика структуры сообществ раковинных амеб в ходе сукцессии “водоем–болото” в значительной степени соотносится с характером локальной растительности. Совместно они определяются либо аутогенными, либо аллогенными факторами сукцессии, сменяющими друг друга во времени. В частности, трансформация водоема в низинное болото вероятнее всего произошла в результате действия климатических (аллогенных) факторов: кратковременное иссушение и пожары, приведшие к зарастанию берегов. Трансформация низинного болото в мезотрофное сфагновое может быть связана как с аутогенными факторами (накопление отложений), так и с аллогенными (снижение активности человека на прилегающей территории).
Список литературы
Бобров А.А., 2003. Историческая динамика озерно-болотных экосистем и сукцессии раковинных амеб (Testacea) // Зоологический журнал. Т. 82. № 2. С. 215–223.
Волкова Е.М., 2010. Заболачивание карстовых и карстово-суффозионнных депрессий на территории Тульской области // Направления исследований в современном болотоведении России (под ред. Юрковской Т.К.). Санкт-Петербург – Тула. С. 146–163.
Волкова Е.М., 2018. Болота Среднерусской возвышенности: генезис, структурно-функциональные особенности и природоохранное значение. Дис. … докт. биол. наук. Тула: Тульский государственный университет. 453 с.
Гвоздецкий Н.А., Жучкова В.К. (ред.), 1963. Физико-географическое районирование Нечерноземного центра. М.: Изд-во Московского ун-та. 451 с.
Гельцер Ю.Г., Корганова Г.А., Алексеев Д.А., 1985. Практическое руководство по идентификации почвенных тестаций. М.: Изд-во Московского университета. 84 с.
Мазей Ю.А., Ембулаева Е.А., 2009. Изменение сообществ почвообитающих раковинных амеб вдоль лесостепного градиента в Среднем Поволжье // Аридные экосистемы. Т. 15. С. 13–23.
Мазей Ю.А., Цыганов А.Н., 2006. Пресноводные раковинные амебы. М.: Товарищество научных изданий КМК. 300 с.
Мильков Ф.Н., Гвоздецкий Н.А., 1976. Физическая география СССР. Общий обзор. Европейская часть СССР. Кавказ. М.: Мысль. 448 с.
Сукачев В.Н., 1926. Болота, их образование, развитие и свойства. Л.: Изд-во Лесного института. 162 с.
Тюремнов С.Н., 1959. Атлас растительных остатков, встречаемых в торфе. М.-Л.: Госэнергоиздат. 227 с.
Beug H.J., 2004. Leitfaden der Pollenbestimmung für Mitteleuropa und angrenzende Gebiete. Verlag Friedrich Pfeil, Munich. 542 s.
Blaauw M., 2010. Methods and code for ‘classical’ age-modelling of radiocarbon sequences // Quaternary Geochronology. V. 5. № 5. P. 512–518.
Blaauw M., Christen J.A., 2019. rbacon: Age-depth modelling using Bayesian statistics. R package version 2.3.9.1. https://CRAN.R-project.org/package=rbacon.
Booth R.K., Ireland A.W., LeBoeuf K., Hessl A., 2016. Late Holocene climate-induced forest transformation and peatland establishment in the central Appalachians // Quaternary Research. V. 85. P. 204–210.
Bray J.R., Curtis J.T., 1957. An ordination of the upland forest communities of southern Wisconsin // Ecological Monographs. V. 27. № 4. P. 325–349.
Chambers F.M., Beilman D.W., YuZ., 2010. Methods for determining peat humification and for quantifying peat bulk density, organic matter and carbon content for palaeostudies of climate and peatland // Mires and Peat. V. 7. P. 1–10.
Chambers F.M., Booth R.K., De Vleeschouwer F., Lamentowicz M., Le Roux G.G. et al., 2012. Development and refinement of proxy-climate indicators from peats // Quaternary International. V. 268. P. 21–33.
Dean W.Jr., 1974. Determination of carbonate and organic matter in calcareous sediments and sedimentary rocks by loss on ignition: comparison with other methods // Journal of Sedimentary Research. V. 44. P. 242–248.
Elliott S.M., Roe H.M., Patterson R.T., 2012. Testate amoebae as indicators of hydroserial change: An 8500 year record from Mer Bleue Bog, eastern Ontario, Canada // Quaternary Interantional. V. 268. P. 128–144.
Gilbert D., Amblard C., Bourdier G., Francez A.J., Mitchell E.A.D., 2000. Le régime alimentaire des thécamoebiens // L’Année Biologique. T. 39. P. 57–68.
Gomaa F., Mitchell E.A., Lara E., 2013. Amphitremida (Poche, 1913) is a new major, ubiquitous labyrinthulomycete clade // PLoS ONE. V. 8. e53046.
Goring S., Williams J.W., Blois J.L., Jackson S.T., Paciorek C.J. et al., 2012. Deposition times in the northeastern United States during the Holocene: establishing valid priors for Bayesian age models // Quaternary Science Reviews. V. 48. P. 54-60.
Grimm E.C., 1987. CONISS: A FORTRAN 77 program for stratigraphically constrained cluster analysis by the method of incremental sum of squares // Computers & Geosciences. V. 13. P. 13–35.
Harnish O., 1925. Studien zur Okologie und Tiergeographie der Moore // Zoologische Jahrbücher. (Abt. Syst.). Bd. 51. S. 1–166.
Ireland A.W., Booth R.K., 2011. Hydroclimatic variability drives episodic expansion of a floating peat mat in a North American kettlehole basin // Ecology. V. 92. № 1. P. 11–18.
Juggins S., 2017. rioja: Analysis of quaternary science data, R package version (0.9-15.1). (http://cran.r-project.org/ package=rioja).
Mitchell E.A.D., Charman D.J., Warner B.G., 2008. Testate amoebae analysis in ecological and paleoecological studies of wetlands: past, present and future // Biodiversity and Conservation. V. 17. P. 2115–2137.
Moore P.D., Webb J.A., Collinson M.E., 1991. Pollen Analysis. Oxford: Blackwell. 216 p.
Novenko E.Y., Tsyganov A.N., Mazei N.G., Kupriyanov D.A., Rudenko O.V. et al., 2019. Palaeoecological evidence for climatic and human impacts on vegetation in the temperate deciduous forest zone of European Russia during the last 4200 years: A case study from the Kaluzhskiye Zaseki nature reserve // Quaternary International. V. 516. P. 58–69.
Oksanen J., Blanchet F.G., Friendly M., Kindt R., Legendre P. et al., 2019. vegan: Community Ecology Package. R package version 2.5-4. https://CRAN.R-project.org/ package=vegan
Patterson R.T., Dalby A., Kumar A., Henderson L.A., Boudreau R.E.A., 2002. Arcellaceans (thecamoebians) as indicators of land-use change: settlement history of the Swan Lake area, Ontario as a case study // Journal of Paleolimnology. V. 28. P. 297e–316.
Payne R.J., 2010. Testate amoeba response to acid deposition in a Scottish peatland // Aquatic Ecology. V. 44. № 2. P. 373–385.
Payne R.J., 2011. Can testate amoeba-based palaeohydrology be extended to fens? // Journal of Quaternary Science. V. 26. № 1. P. 15–27.
Payne R.J., Malysheva E.A., Tsyganov A.N., Pampura T.V., Novenko E.Y.et al., 2016. A multi-proxy record of Holocene environmental change, peatland development and carbon accumulation from Staroselsky Moch peatland, Russia // The Holocene. V. 26. № 2. P. 314–326.
Reille M., 1992. Pollen et spores d’Europe et d’Afrique du Nord. Laboratoire de Botanique Historique et Palynologie. Marseille. 543 p.
Reimer P.J., Bard E., Bayliss A., Beck J.W., Blackwell P.G. et al., 2013. IntCal13 and Marine13 radiocarbon age calibration curves 0–50,000 years cal BP // Radiocarbon. V. 55. № 4. P. 1869–1887.
R Core Team, 2018. R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. URL https://www.R-project.org/
Ryding H., Juglum J., 2006. The biology of peatlands. Oxford: Oxford University Press. 343 p.
Schönborn W.E., 1984. Studies on remains of testacea in cores of the Great Woryty Lake (NE Poland) // Limnologica. V. 16. P. 185–190.
Simpson G.L., Oksanen J., 2018. analogue: Analogue matching and Modern Analogue Technique transfer function models. (R package version 0.17-1). https:// cran.r-project.org/package=analogue
Smirnova O.V., Bobrovsky M.V., Khanina L.G., Braslavskaya T.Yu., Starodubtseva E.A. et al., 2017. Nemoral forests // Smirnova O.V., Bobrovsky M.V., Khanina L.G. (Eds), European Russian Forests. Their Current State and Features of Their History. Springer Netherlands. P. 337–480.
Sonnenburg E.P., Boyce J.I., Reinhardt E.G., 2013. Multi-proxy lake sediment record of prehistoric (Paleoindian–Archaic) archaeological paleoenvironments at Rice Lake, Ontario, Canada // Quaternary Science Reviews. V. 73. P. 77–92.
Tolonen K., 1966. Stratigraphic and rhizopod analyses on an old raised bog, Varrassuo, in Hollola, South Finland // Annales Botanici Fennici. V. 3. P. 147–166.
Warner B.G., Charman D.J., 1994. Holocene changes on a peatland in Northwestern Ontario interpreted from testate amoebae (Protozoa) analysis // Boreas. № 23. P. 270–279.
Дополнительные материалы отсутствуют.
Инструменты
Зоологический журнал