

Зоологический журнал, 2020, T. 99, № 6, стр. 656-664
Личинки рода Rainieria Rondani 1843 (Diptera, Micropezidae)
Н. П. Кривошеина a, *, М. Г. Кривошеина a
a Институт проблем экологии и эволюции им. А.Н. Северцова РАН
119071 Москва, Россия
* E-mail: dipteranina@rambler.ru
Поступила в редакцию 11.09.2019
После доработки 29.10.2019
Принята к публикации 19.12.2019
Аннотация
Впервые приводятся сведения об образе жизни и морфологии личинок рода Rainieria: R. hennigi Krivosheina et Krivosheina 1996 и R. latifrons (Loew 1870), обитающих в зоне коры поврежденных и ослабленных древесных стволов. Установлены основные диагностические признаки личинок, различающихся по строению передних и задних дыхалец, а также по строению ротового аппарата.
Micropezidae – довольно крупное семейство двукрылых насекомых, виды которого широко распространены по всему миру. В Палеарктике представлены виды из 3 подсемейств: Calobatinae распространены в Голарктике, Micropezinae известены в Голарктике и Неотропике, Taeniapterinae представлены преимущественно тропическими видами с выходом в Голарктику (Steyskal, 1987).
Имаго часто встречаются на листьях и древесных стволах, а также тяготеют к таким субстратам, как гниющие растения, навоз, гниющие фрукты. Эти двукрылые встречаются преимущественно в лесных районах, хотя некоторые виды являются типичными обитателями лугов и открытых болотистых местностей, а виды рода Micropeza Meigen 1803 обычны на бобовых культурах (Ferrar, 1987).
Сведения по биологии видов очень незначительные и скудные. Но все-таки хорошо заметны различия в образе жизни представителей подсемейств. Так, личинки палеарктических видов подсемейства Calobatinae Compsobata cibaria (Linnaeus 1761) и Calobata petronella (Linnaeus 1761) обитают в скоплениях гниющих трав (Brindle, 1965). Установлены также связи последнего вида с унавоженными переувлажненными субстратами (Лобанов, 1960). Неарктический вид Compsobata univitta (Walker 1849) выведен из скоплений гниющих растений около ручья (Teskey, 1972).
В роде Micropeza Meigen 1803 (Micropezinae), представленном в Палеарктике не менее чем 20 видами, известен образ жизни лишь одного вида M. corrigiolata (Linnaeus 1767). В Европе он выведен из корневых клубеньков некоторых бобовых (Müller, 1957). Имаго откладывают яйца в верхние слои почвы, выходящие из них личинки заселяют только свежие, не поврежденные клубеньки, проделывая в них ходы и формируя внутренние полости. Взрослые личинки уходят на окукливание в почву на глубину до 30 см (Müller, 1957; Лобанов, 1960). Вид не рассматривается в качестве серьезного вредителя, но при высокой численности иногда может наносить заметный вред.
Сведения по экологии видов подсемейства Taeniapterinae не менее скудные. Так, неарктический вид Calobatina geometroides (Cresson 1926) выведен в США из влажных органических остатков дупел древесных пней (Wallace, 1969). Вид Cephalosphen conifrons (Bigot 1886) выведен в Южной Африке из пупария, обнаруженного в гнилой древесине кораллового дерева Erythrina caffra Thunb. (Leguminosae). Вид Mimegrella albimana (Doleschall 1856), известный из Японии, Ориентального региона и Океании (Soós, 1984), выведен на Соломоновых о-вах из-под коры мертвого дерева (Bohart, Gressitt, 1951), а M. coeruleifrons (Macquart 1843) на Филиппинских о-вах – из корней имбиря. Причем последний вид рассматривается в качестве первичного вредителя, его личинки обнаружены в ходах в твердой сочной наружной части корней, а пупарии – в сухой волокнистой центральной части (Steyskal, 1964). Он был выведен также из гниющих растений и фруктов (Hennig, 1936).
Виды рода Rainieria практически не исследованы. Имеются лишь ссылки на топические связи с древесными субстратами (Sabrosky, 1942; Chandler, 1975; Мамаев и др., 1977). Так, неарктический вид R. antennaepes (Say 1823) выведен в США из пупария, найденного в расщепе старого ильмового дерева Ulmus sp. на участке относительно плотной, мертвой, но влажной, частично пораженной грибами древесины (Sabrosky, 1942). В Европе (Великобритания) для Rainieria calceata (Fallén 1820) отмечены связи с упавшими березовыми стволами (Chandler, 1975).
Виды Taeniaptera Macquart 1835: T. annulata (Fabricius 1787) и T. lasciva (Fabricius 1798) в Неотропике развиваются в оставленных в почве черенках бананов (Fischer, 1932). Кроме того, последний вид был выведен в США из-под коры корней инжира (Steyskal, 1964).
На основании имеющихся сведений известных личинок видов Calobatinae следует рассматривать в качестве сапрофагов, питающихся в гниющих травянистых тканях. Личинки Micropeza являются фитофагами, повреждают только живые, здоровые ткани бобовых растений. Разнообразные представители Taeniapterinae обнаружены преимущественно в гниющих древесных субстратах.
В литературе имеются детальные описания личинок и приведены рисунки неарктического вида Compsobata univitta (Calobatinae) (Teskey, 1972), палеарктического вида Micropeza corrigiolata (Micropezinae) (Müller, 1957) и краткое описание пупария Rainieria antennaepes (Taeniapterinae) (Sabrosky, 1942). Для личинок рода Rainieria приведены лишь рисунки передних и задних дыхалец и ротоглоточного аппарата Rainieria latifrons (Loew 1870) как иллюстрации к определительной таблице семейств, без описания (Мамаев и др., 1977).
В данной работе впервые рассмотрены основные морфологические признаки личинок и пупариев видов рода Rainieria – R. hennigi Krivosheina et Krivosheina 1996 и R. latifrons (Loew 1870), а также приводится определительная таблица родов по личинкам.
Материал был собран в виде личинок и куколок, часть которых после ошпаривания кипятком фиксировали в 70% спирте, а часть в лабораторных условиях до выведения имаго помещали в боксы с древесным субстратом. Для выведенных линий личинка–куколка–имаго сохраняли сквозную нумерацию. Определение проводили по вылетевшим имаго.
МОРФО-ЭКОЛОГИЧЕСКАЯ ХАРАКТЕРИСТИКА ЛИЧИНОК
Rainieria hennigi Krivosheina et Krivosheina 1996 (рис. 1, 1–4; 3; 4, 3)
Рис. 1.
Личинки R. hennigi Krivosheina et Krivosheina 1996 (1–4), Compsobata univitta (Walker 1849) (5, 6), Micropeza corrigiolata (Linnaeus 1767) (7, 8), Rainieria antennaepes (Say 1823) (9–10) и R. antennaepes var. brunneipes (Cresson 1938) (11): 1 – ротоглоточный аппарат; 2, 5, 7, 10, 11 – задние дыхальца; 3, 6, 8, 9 – передние дыхальца; 4 – терминальный конец тела сбоку (рис. 5, 6 по: Teskey, 1972; рис. 7, 8 по: Müller, 1957; рис. 9, 10 по: Steyskal, 1964; рис. 11 по: Sabrosky, 1942). a – ротовые крючья, b – вентральный выступ базального склерита ротовых крючьев, c – парастомальный склерит, d – дорсальный отросток фарингеального склерита, f – фарингеальный склерит, g – гипостомальный склерит, v – вентральный отросток фарингеального склерита.

Материал. 2 личинки, 3 пупария, 2 имаго, Россия, Сахалинская обл., Кунашир. Менделеево. 18.IV.1977. № 24, личинки под корой березы; 28.VI.1977. № 498, личинки под корой пихты (А.И. Зайцев, Б.М. Мамаев).
Описание. Личинка. Тело практически белое, с легким желтоватым оттенком. Передний конец сужен, конусовидный. Терминальный сегмент слегка уплощен дорсовентрально, с двумя небольшими конусовидными выступами, несущими задние дыхальца.
Длина первого грудного сегмента значительно превосходит ширину, два других сегмента, как и брюшные, короткие. Брюшные сегменты поперечные, длина первых шести сегментов слегка уступает ширине, длина двух последних сегментов почти в 1.5 раза уступает ширине. Покровы сегментов гладкие, без заметных складок и кутикулярных структур, кроме ползательных валиков, расположенных в переднем отделе брюшных сегментов. Последние с четырьмя–пятью поперечными рядами мелких светлых, слабо заметных шипиков. Короткие ряды таких же шипиков расположены в терминальном отделе предшествующего сегмента.
Анальная пластинка светлая, со слабовыраженными границами, практически овальная, но несколько расширена к заднему краю. Анальное поле спереди и сбоку с единичными рядами белых микроскопических шипиков, расположенных в форме трапеции.
Трахейная система амфипнейстического типа, с двумя парами дыхалец (передними и задними). Передние дыхальца расположены на боковой стороне тела на границе I и II грудных сегментов. Дыхальца с широким коротким атриумом, его длина несколько уступает ширине (рис. 1, 3). Передний край атриума с веерообразно расположенными пальцевидными выростами, несущими на вершине стигмальные камеры. Они расположены на небольших несколько изолированных боковых дуговидных выступах и на вогнутом срединном отделе; по 4 пальцевидных выроста по бокам и с 3 длинными и 3–4 короткими выростами в среднем отделе (всего просматривается 14–15 выростов со стигмальными камерами).
Задние дыхальца расположены на наружной стороне небольших бугорков (рис. 1, 4). Бугорки и стигмальная пластинка дыхалец слегка буроватые, покровы вокруг них также затемнены, буроватые (рис. 1, 2). Стигмальная пластинка светлая, овальная с 3 извилистыми дыхальцевыми щелями, несущими двойной ряд круглых светлых микрокамер. Наружные щели расположены вдоль всей длины пластинки, внутренние более короткие и находятся соответственно в передней и задней половинах стигмальной пластинки. Перитрема, расположенная по периферии стигмальной пластинки, на большем протяжении широкая, хорошо склеротизована, в переднебоковых углах с коротким конусовидным выступом.
Ротоглоточный аппарат типичного для круглошовных двукрылых строения, с хорошо развитыми склеротизованными ротовыми крючьями и отростками фарингеального склерита (рис. 1, 1). Ротовые крючья массивные, темные, серповидные. Дополнительные, акцессорные, склериты отсутствуют. Вентральный выступ базальных склеритов ротовых крючьев более светлый, слегка дуговидно выгнут, его вершина направлена вниз. Гипостомальный и парастомальный склериты хорошо развиты, удлиненные. Дорсальные выросты фарингеального склерита короче и ýже вентральных. Вентральные выросты без заметного бугорка на дорсальной стороне вблизи их основания. Фарингеальная мембрана, соединяющая вентральные отростки фарингеального склерита, хорошо развита. Ее наличие скорее всего говорит о таком типе питания, как сапрофагия.
Длина тела 9 мм, ширина 1 мм.
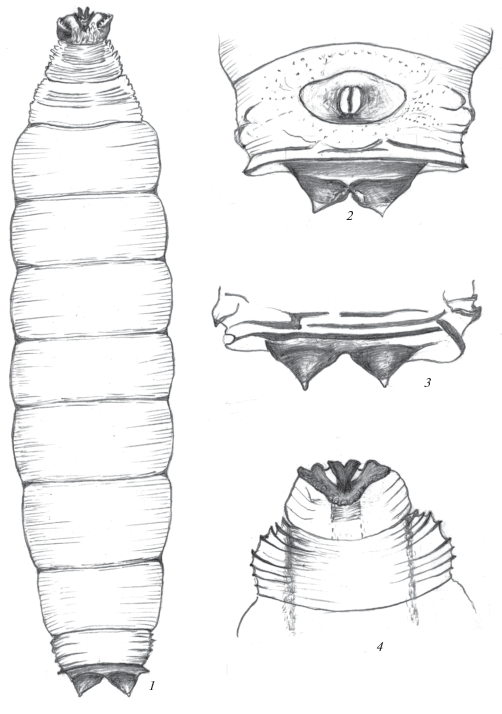
Пупарий. Поверхность покровов пупария светло-коричневая, терминального сегмента – несколько темнее. Кутикулярные структуры на поверхности сегментов не выражены, но все сегменты с четкими поперечными параллельными бороздками, наиболее хорошо заметными на боковых и дорсальной сторонах сегментов (рис. 3, 1). Единичные тупые микробугорки заметны на боковых сторонах сегментов, их число возрастает к концу тела. Первый и второй грудные сегменты в среднем отделе с дорсальной стороны с многочисленными продольными и поперечными короткими бороздками (рис. 3, 3). На боковых сторонах переднегруди расположены передние дыхальца в виде поперечных сероватых пластинок, несущих округлые дыхальцевые камеры. Первый грудной сегмент с вентральной стороны с хорошо склеротизованными широкими бороздками и бугорками характерной формы, срединным вилкообразным, боковыми с массивными поперечными выступами. Остальные сегменты с параллельными поперечными бороздками (рис. 3, 4).
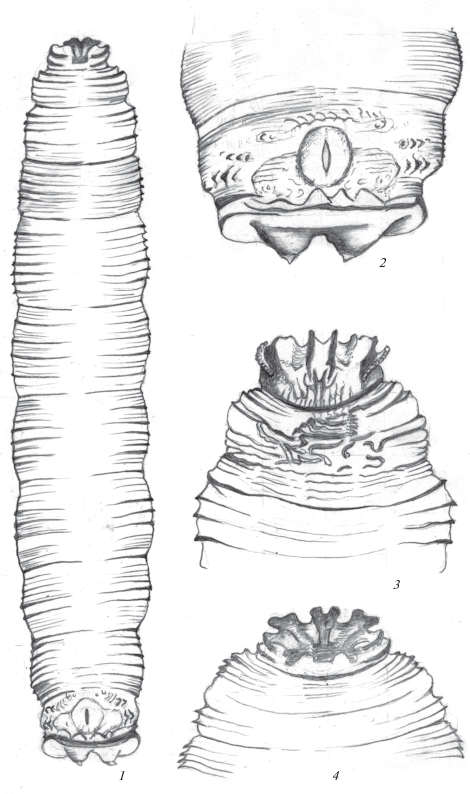
Рис. 2.
Личинка Rainieria latifrons (Loew 1870): 1 – общий вид переднего и терминального отделов тела сбоку, 2 – ротоглоточный аппарат, 3 – терминальный конец тела сбоку, 4 – задние дыхальца, 5 – переднее дыхальце, 6 – анальная пластинка.
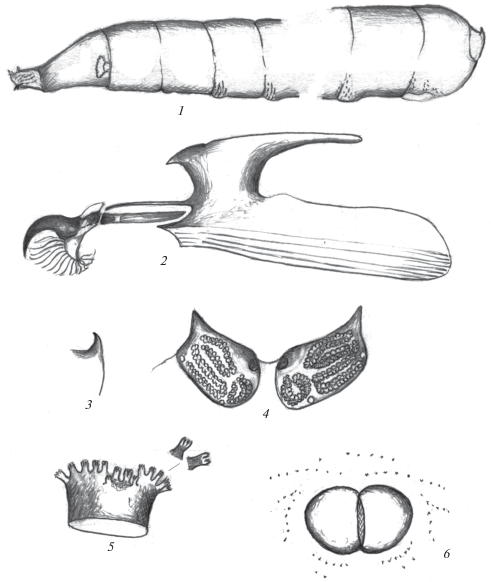
Рис. 3.
Пупарий Rainieria hennigi Krivosheina et Krivosheina 1996: 1 – общий вид с вентральной стороны; 2 – конец тела с вентральной стороны; 3, 4 – передний отдел тела дорсально и вентрально.
Конец тела тупо обрублен. Участок с задними дыхальцами ограничен темными дуговидными складками, снаружи от которых расположены более короткие дополнительные складки. Дорсальная сторона терминального сегмента помимо длинной поперечной борозды, отделяющей стигмальное поле, с несколькими поперечными хорошо склеротизованными бороздами перед ней (рис. 4, 3). Вентральная сторона терминального сегмента также, помимо борозды, отделяющей стигмальное поле, с рядом четких борозд перед ней. Анальная пластинка овальная, с четкими границами, к ее боковым сторонам примыкают дополнительные обособленные пластинки с поперечно исчерченными бороздками (рис. 3, 2). Между ними и боковой стороной сегмента расположено несколько коротких склеротизованных бугорков.
Рис. 4.
Пупарий Rainieria latifrons (Loew 1870) (1, 2, 4) и R. hennigi Krivosheina et Krivosheina 1996 (3): 1 – общий вид с дорсальной стороны, 2 – терминальный конец тела с вентральной стороны, 3 – конец тела с дорсальной стороны, 4 – передний конец тела с вентральной стороны.
Длина пупария 7 мм, ширина 0.9 мм.
Биология. Вид описан с о-ва Кунашир. Личинки R. hennigi зарегистрированы в упавших стволах березы, а также в нависающих над землей на высоте 1 м стволах пихты. В стволах березы взрослые личинки и пупарии регистрировались в толще темного влажного луба и в покинутых ходах дровосеков на границе луба и заболони. В стволах пихты, где обитали личинки разных возрастов, они встречались на участках с прочной, плотно прилегающей корой и относительно свежим лубом, заселенным по границе с заболонью личинками жуков-дровосеков Rhagium inquisitor (Linnaeus 1758) (Cerambycidae). Кроме того, личинки R. hennigi встречались в разжиженном лубе, заселенном личинками Mycetobia sp. (Mycetobiidae) и Libnotes kariyana (Alexander 1947) (Limoniidae).
Распространение. Дальний Восток России: Курильские о-ва (Кунашир); Япония (Хоккайдо, Хонсю) (Krivosheina, Krivosheina, 1996).
Rainieria latifrons (Loew 1870) (рис. 2; 4, 1, 2, 4).
Материал. 8 личинок, 6 пупариев, 6 имаго, Россия, Краснодарский край, ст. Северская. 9.VII.1970, личинки в пропитанной водой светлой рыхлой древесине граба Carpinus sp.; Амурская обл., Хинганский заповедник, Кундур, 2.VI.1975, личинки в древесине дуба Quercus sp.; Хабаровский край, Большехехцирский заповедник, Бычиха, 19.IV.1976, под корой маакии Maackia amurensis Rupr et Maxim; там же 23.V.1976, личинки в древесине березы Betula sp.; Приморский край, Лазовский заповедник, Корпадь, 23.VI.1979, личинки в древесине березы; Уссурийский заповедник, 15.IX.1964, личинки в древесине кедровой сосны Pinus koraiensis Siebold et Zucc, и другие сборы (А. Зайцев, Н. Кривошеина).
Описание. Личинка. Тело в спокойном состоянии компактное, с относительно короткими сегментами, при передвижении значительно вытягивается, и передний конец заметно сужается. Первый грудной сегмент удлиненный, его длина превосходит ширину (рис. 2, 1). Последний сегмент относительно крупнее остальных, закруглен на конце. На наружной стороне сегмента расположены небольшие овальные бугорки, несущие задние дыхальца. Стигмальный бугорок плоский, более узкий по сравнению с бугорком у R. hennigi (рис. 2, 3).
Покровы светлые, слегка желтоватые, гладкие, без заметных кутикулярных структур. Небольшие ползательные валики расположены в переднем отделе брюшных сегментов. Ползательные валики с небольшими светлыми слабо заметными шипиками, образующими несколько поперечных рядов. Отсутствие склеротизации и небольшие размеры не позволяют выявить различия в строении полей шипиков у рассматриваемых видов.
Анальная пластинка светлая, в виде двух овальных бугорков окружающая анальную щель (рис. 2, 6). Единичные ряды мелких шипиков расположены перед ней и вдоль боковых сторон. Два симметричных коротких ряда светлых микрошипиков расположены за анальной пластинкой.
Трахейная система амфипнейстического типа, с 2 парами дыхалец. Передние дыхальца расположены на боковых сторонах переднегруди на границе I и II грудных сегментов. Атриум дыхалец широкий и короткий (рис. 2, 5), на его переднем крае расположено 14 пальцевидных выростов, несущих дыхальцевые камеры. По 5 выростов расположено на дуговидно выступающих боковых сторонах и 4 выроста (два более длинных и два коротких) расположены на вогнутом крае в среднем отделе (рис. 2, 5).
Задние дыхальца распложены на наружной стороне небольших светлых бугорков (рис. 2, 3). Стигмальные пластинки вытянуты в поперечном направлении, их длина в 1.5 раза превосходит ширину (рис. 2, 4). Пластинки дыхалец не соприкасаются, разделены широкой светлой полоской покровов, слегка буроватые, с небольшим более темным конусовидным выступом. На поверхности стигмальной пластинки расположены 3 извилистые дыхальцевые щели, две более длинные занимают большую часть поверхности стигмальной пластинки, третья, более короткая, расположена вдоль ее внутреннего края (рис. 2, 4). Поверхность покровов вокруг стимального бугорка светлая, не буроватая.
Ротоглоточный аппарат с хорошо развитыми ротовыми крючьями и длинными отростками фарингеального склерита (рис. 2, 2). Ротовые крючья массивные, темные, серповидные, заметно утолщены в основании. Вентральные выступы их базальных склеритов дуговидно выгнуты снаружи, а их вершины направлены назад. Вентральные отростки фарингеального склерита заметно длиннее и массивнее дорсальных, с небольшим плоским бугорком с дорсальной стороны вблизи их основания. Фарингеальная мембрана хорошо развита.
Длина тела 7 мм, ширина 0.9.мм.
Пупарий. Поверхность пупария светлая, желтая, со слабо выраженными поперечными бороздками (рис. 4, 1). Морщинистые тонкие продольные складки расположены только в заднем отделе первого грудного сегмента. Передний отдел I грудного сегмента с дорсальной стороны с 2 срединными склеротизованными заостренными бороздками и хорошо развитыми передними дыхальцами, расположенными на боковых сторонах сегмента. Передний отдел I грудного сегмента с вентральной стороны с широкими массивными склеротизованными лопастями (рис. 4, 4). Вершины лопастей вилкообразного срединного склерита не закруглены, а тупо обрублены, плоские. Наиболее четкие темные поперечные бороздки развиты лишь на II–III грудных и терминальном сегментах (рис. 4, 1, 2). Поперечные бороздки на II и III грудных сегментах формируют более темные пояски (рис. 4, 1). Несколько овальных выступов расположено вблизи боковых сторон терминального сегмента (рис. 4, 2). Его конец тупо обрублен. Участок стигмального поля ограничен простой дуговидной складкой с дорсальной и вентральной сторон, дополнительно с вентральной стороны перед складкой — 3 коротких поперечных полоски (рис. 4, 1, 2). Вся вентральная поверхность терминального сегмента с рассеянными точечными микроструктурами.
Анальная пластинка поперечная, овальная, со слегка оттянутыми боковыми сторонами (рис. 4, 2). Длина пупария 6 мм, ширина при не вылетевших имаго – 1.2 мм. Пустые пупарии ýже.
Биология. Личинки в Амурской обл. найдены в светлой влажной древесине лежащих на земле стволов липы Tilia amurensis Rupr., в толще желтой влажной древесины дуба Quercus mongolica Fisch., в буром влажном лубе в зоне коры маакии Maackia amurensis Rupr. и пихты Abies sp. В Хабаровском крае личинки зарегистрированы в темной трухе среди волокон луба и непосредственно в рыхлой влажной древесине липы. В Лазовском заповеднике личинки обнаружены в светлой влажной древесине упавших стволов березы Betula sp. Перед окукливанием личинки перемещаются в черную влажную гниль луба непосредственно под поверхностный слой корки. В Уссурийском заповеднике личинки обитают в мягкой светлой древесине упавших стволов кедра корейского (Pinus koraiensis), где зарегистрированы совместно с личинками двукрылых Clusia unita Mamaev 1974 (Clusiidae). На территории Кедровой Пади личинки R. latifrons были обнаружены в красно-бурой древесине лежащих на земле стволов чозении Salix arbutifolia (Pall.) A.K. Skvortsov в прибрежной зоне р. Кедровка. Взрослые личинки зарегистрированы непосредственно под корой, где встречались совместно с личинками Sciaridae. Личинки на Дальнем Востоке и в Бурятии неоднократно регистрировались в лежащих на земле стволах Populus tremula L., где обитали под корой на поверхности заболони. Пупарии, как правило, регистрировались под легко отслаивающейся коркой, на поверхности луба. В осине личинки встречались совместно с личинками нескольких видов рода Homalocephala Zetterstedt 1838 (Otitidae).
Распространение. Вид распространен на юге Европы, включая юг европейской части России, в Западной и Восточной Сибири, на Дальнем Востоке, включая Хабаровский край и Южное Приморье. На Курильских о-вах (Кунашир) собраны единичные экземпляры имаго. Распространение вида в Японии требует подтверждения (Krivosheina, Krivosheina, 1996).
На основании имеющихся в литературе сведений и собственных сборов составлена определительная таблица родов Micropezidae.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА РОДОВ И ВИДОВ MICROPEZIDAE ПО ЛИЧИНКАМ
1. Задние дыхальца с коротким конусовидным выступом, его длина в несколько раз меньше ширины стигмальной пластинки (рис. 1, 2, 10; 2, 4). Извилистые дыхальцевые щели занимают практически всю поверхность стигмальной пластинки. Передние дыхальца не менее чем с 14 дыхальцевыми камерами. Taeniapterinae. Rainieria Rondani 1843 ………………………….………………............. 2.
— Задние дыхальца с массивным конусовидным выступом. Его длина лишь на 1/5 меньше ширины стигмальной пластинки (рис. 1, 5, 7) ...3.
2. Стигмальная пластинка задних дыхалец с 1 длинной и 2 короткими извилистыми дыхальцевыми щелями (рис. 1, 2). Длинная щель расположена вдоль наружной стороны стигмальной пластинки, а короткие – симметрично в ее переднем и заднем отделах с внутренней стороны. Дыхальца практически соприкасаются и окружены коричневатыми склеротизованными покровами. Вентральный отросток фарингеального склерита без выраженного дорсального выступа (рис. 1, 1). Вершина вентрального выступа ротовых крючьев направлена вертикально вниз. ……………R. hennigi Kriv. et Kriv.
— Стигмальная пластинка задних дыхалец с 2 длинными и 1 короткой извилистой дыхальцевой щелью (рис. 2, 4). Две длинные дыхальцевые щели расположены симметрично в переднем отделе стигмальной пластинки. Дыхальца не соприкасаются и окружены светлыми покровами. Вентральный отросток фарингеального склерита с дорсальным овальным выступом в основании (рис. 2, 2). Вершина вентрального выступа ротовых крючьев направлена назад ………… R. latifrons Loew
3. Передние дыхальца вееровидные, с 4 пальцевидными выростами (рис. 1, 8), задние дыхальца темные, удлиненные, их длина у взрослых личинок не менее, чем в 3 раза превосходит их ширину (рис. 1, 7). Дыхальца с 3 широкими прямыми дыхальцевыми щелями, несущими многочисленные светлые микроотверстия. Micropezinae ………………....... Micropeza Meigen 1803 [M. corrigiolata (Linnaeus 1767)]
— Передние дыхальца с коротким атриумом, несущим 8 коротких закругленных дыхальцевых камер, расположенных в один ряд (рис. 1, 6). Задние дыхальца с 3–4 узкими, лентовидными дыхальцевыми щелями различной длины. Calobatinae ……………………………………………...............4.
4. Задние дыхальца с относительно светлой стигмальной пластинкой и 4 узкими, прямыми, лентовидными дыхальцевыми щелями различной длины (рис. 1, 5) ……………………………Compsobata Czerny 1930 [С. univitta (Walker 1849)
— Задние дыхальца с темной стигмальной пластинкой и с 3 лентовидными, дуговидно изогнутыми стигмальными щелями …………Calobata Meigen 1803 [C. petronella (Linnaeus 1761)]
Список литературы
Лобанов А.М., 1960. Материалы к биологии и экологии Trepidaria petronella L. (Diptera, Tylidae) // Зоологический журнал. Т. 39. № 6. С. 888–891.
Мамаев Б.М., Кривошеина Н.П., Потоцкая В.А., 1977. Определитель личинок хищных насекомых – энтомофагов стволовых вредителей. М.: Наука. 392 с.
Bohart G.E., Gressitt J.L., 1951. Filth-inhabiting flies of Guam // Bulletin Bernice P. Bishop Museum. V. 204. P. 1–152.
Brindle A., 1965. Taxonomic notes on the larvae of British Diptera. No. 19. The Micropezidae (Tylidae) // Entomologist. V. 98. P. 83–86.
Chandler P.J., 1975. Notes on the British status of three unusual acalypterate flies (Diptera) // Proceeding of the British Entomological (Natural History) Society. V. 8. P. 66–72.
Ferrar P., 1987. A Guide to the breeding habits and immature stages of Diptera Cyclorrhapha. Entomonograph V. 8. Part 1: text. Leiden – Copenhagen: E.J. Brill/ Scandinavian Science Press. 478 p.
Fischer C.R., 1932. Contribuicao para o conhecimento da metamorphose e posicao systematic da familia Tylidae (Micropezidae, Dipt.). Revista de Entomologia. Rio de Janeiro. V. 2. P. 15–24.
Hennig W., 1936. Revision der Tyliden (Dipt., Acalypt.). II. Teil // Konowia. V. 15. S. 201–239.
Krivosheina M.G., Krivosheina N.P., 1996. The species from the genus Rainieria Rondani (Diptera, Micropezidae) of Russia including the description of a new species // Studia Dipterologica. Bd. 3. H. 1. S. 97–100.
Müller H., 1957. Legumunosenknöllchen als Nahrungsquelle heimischer Micropezidae- (Tylidae-) Larven Diptera. Zur Morphologie und Biologie der bisher unbekannten Larve von Micropeza corrigiolata L.) // Beiträge zur Entomologie. Bd. 7. S. 247–262.
Sabrosky C.W., 1942. An unusual rearing of Rainieria brunneipes (Cresson) (Diptera: Micropezidae) // Entomological News. V. 53. P. 283–285.
Soós A., 1984. Family Micropezidae (Tylidae). P. 19–24. In: Soós A., Papp L. (eds). Catalogue of palaearctic Diptera. V. 9. Micropezidae – Agromyzidae. Budapest: Akadémiai Kiadó. 460 p.
Steyskal G.C., 1964. Larvae of Micropezidae (Diptera), including two species that bore in ginger roots // Annals of the entomological Society of America. V. 57. P. 292–296.
Steyskal G.C., 1987. Micropezidae. P. 761−767. In: McAlpine J.F. (ed.). Manual of Nearctic Diptera. V. 2. Monograph № 28. Agriculture Canada. P. 675−1332.
Teskey H.J., 1972. The mature larva and pupa of Compsobata univitta (Diptera: Micropezidae) // Canadian Entomologist. V. 104. P. 295–298.
Wallace J.B., 1969. The mature larva and pupa of Calobatina geometroides (Cresson) (Diptera: Micropezidae) // Entomological News. V. 80 P. 317–321.
Дополнительные материалы отсутствуют.
Инструменты
Зоологический журнал