

Зоологический журнал, 2020, T. 99, № 6, стр. 612-615
Новые данные о репродуктивной системе Syrnolopsis minuta Bourguignat 1885 (Gastropoda, Cerithioidea) из озера Танганьика (Африка)
Т. Я. Ситникова *
Лимнологический институт СО РАН
664033 Иркутск, Россия
* E-mail: sit@lin.irk.ru
Поступила в редакцию 14.02.2019
После доработки 18.03.2019
Принята к публикации 15.05.2019
Аннотация
На тотальных препаратах и гистологических срезах исследована половая система брюхоногого моллюска Syrnolopsis minuta Bourguignat 1885 из оз. Танганьика. Показано, что паллиальный женский гонодукт исследованных улиток сходен по общему плану строения с гонодуктом, описанным ранее для других представителей трибы Syrnolopsini, но имеет гермафродитную гонаду. Выявленный парадокс свидетельствует о необходимости продолжения исследований.
В настоящее время 4 рода (Syrnolopsis Smith 1880, Anceya Bourguignat 1885, Martelia Dautzenberg 1908 и Tanganyicia Crosse 1880) эндемичных брюхоногих моллюсков оз. Танганьика на основании морфологических и молекулярно-генетических данных (Wilson et al., 2004; Strong, Glaubrecht, 2007) принято рассматривать в качестве монофилетичной трибы Syrnolopsinni Bourguignat 1890 в рамках подсемейства Hauttecoeuriinae Bourguignat 1885 семейства Paludominae Stoliczka 1868 (Bouchet at al., 2017). Род Syrnolopsis включает 3 вида S. lacustris E.A. Smith 1880, S. minuta Bourguignat 1885 и S. gracilis Pilsbry et Beecher 1929, населяющих мягкие грунты литорали (West et al., 2003). Анатомия, включая репродуктивную систему, изучена в деталях только у одного раздельнополого вида S. lacustris (Mandahl-Barth, 1954; Strong, Glaubrecht, 2007). Мужской паллиальный гонодукт у них состоит из расширенной трубки, выполняющей функцию простаты, и органа, формирующего сперматофор. Женский гонодукт представлен проксимальной белковой и дистальной капсульной железами. Железы соединены вентральным желобком, в который открываются сперматофорная бурса и проток семяприемника (Strong, Glaubrecht, 2007). Строение гонады не изучено ни у одного из представителей трибы Syrnolopsini.
Данное сообщение посвящено не исследованному ранее виду S. minuta, который широко распространен в оз. Танганьика (West et al., 2003) и доминирует на мягких грунтах в мелководной зоне озера (Keating, Wing, 2007).
Материалом для исследований послужили 25 экз. S. minuta. Четыре из них предоставлены проф. Ф. Бандель (Prof. F. Bandel, Германия) с этикеткой “оз. Танганьика”. Двадцать один экземпляр собран Эллинор Мичел (Dr. Ellinor Michel, Великобритания) с помощью легкой водолазной техники с песчаного грунта на глубине 15 м в заливе Катабэ оз. Танганьика, 21.08.2009.
Улитки зафиксированы 80% этиловым спиртом, перед вскрытием все раковины были сфотографированы с помощью цифрового фотоаппарата Cannon 60OES с макрообъективом 65 мм. Половые органы исследованы на тотальных препаратах, окрашенных водным раствором азокармина, и гистологических парафиновых срезах, толщиной 5–7 мкм, окрашенных гематоксилином и заключенных в канадский бальзам (Лилли, 1969). Исследования препаратов и фотографирование выполнено с помощью светового микроскопа Axio Imager M1, с цифровой камерой Axio Cm Mrc5 (K. Zeiss, Germany).
Выборка гастропод с высотой раковины 5.8–6.25 мм состояла из двух морфотипов: с гладкой раковиной (S. m. morph minuta) и со скульптурой в виде продольных килей (S. m. morph carinifera Smith 1889) (рис. 1). Только у пяти улиток на последнем обороте раковины у базального края устья были хорошо выражены от 2 до 4 “линий остановки роста” (рис. 1B–1E), что косвенно свидетельствовало об их взрослом, половозрелом, возрасте. На некоторых раковинах находились кладки с одиночными яйцами, собранными в группы по 5–7 штук (рис. 1G).
Рис. 1.
Раковины Syrnolopsis minuta из оз. Танганьика: A–C – типичный морфотип, D–F –морфотип carinifera, G – яйцевые массы. Масштаб 1 мм.

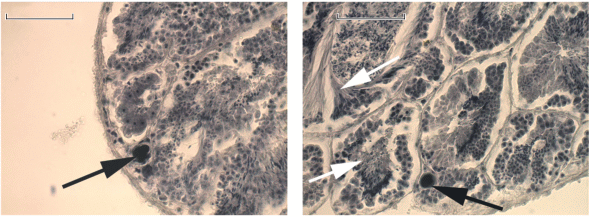
Общий план строения половой системы пяти экземпляров (рис. 1A–1E) вполне соответствовал строению самок S. lacustris (Strong, Glaubrecht, 2007), т.е. включал белковую и капсульную железы, соединенные с латеральной стороны вентральным желобком. Вдоль желобка проходил длинный проток крупного семяприемника (рис. 2A, 3A). На вентральной стороне капсульной железы присутствовал “мешок” или сперматофорная бурса, согласно Шторм и Глаубрехт (Strong, Glaubrecht, 2007) для Anceya giraudi Pilsbry et Beecher 1929 (рис. 3). Исследования гонад двух из пяти особей на гистологических препаратах выявили необычную картину, а именно наличие многочисленных ацинусов с развивающимися сперматоцитами и уже зрелыми сперматозоидами, при этом присутствовали в незначительных количествах и молодые ооциты (рис. 4). Таким образом, гонада у особей, первоначально идентифицированных как самки, имела типичное для гермафродитов строение.
Рис. 2.
Паллиальные половые органы Syrnolopsis minuta: A – вентральная сторона; B – дорсальная сторона гермафродитных особей; С – вентральная сторона молодой особи, пол которой определить не удалось; ag – белковая железа, cg – капсульная железа, i – кишка, mp – мужские половые органы, srd – проток семяприемника, st – стилет. Масштаб 1 мм.
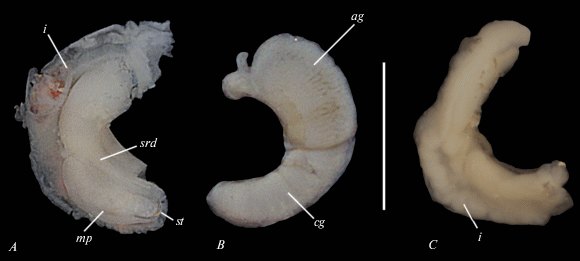
Рис. 3.
Паллиальный гонодукт Syrnolopsis minuta: A – женские половые органы на гистологическом срезе, B – мужские органы, C – стилет или сперматофор; sr – семяприемник, vd – семяпровод (остальные обозначения как на рис. 2). Масштаб (мкм): A – 200; B, C – 100.
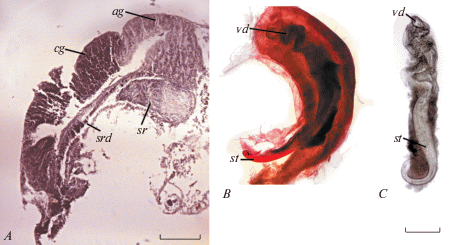
Рис. 4.
Гонада Syrnolopsis minuta (особь на pис. 1С, 3А) на гистологических срезах. Белые стрелки – ацинусы с развивающимися сперматозоидами, черные стрелки – молодые ооциты. Масштаб 50 мкм.
Эта находка позволяет иначе интерпретировать функцию “мешка”, занимающего позицию сперматофорной бурсы у других Syrnolopsini, т.е. считать его мужским гонодуктом. Стенки “мешка”, вероятно, выполняют функцию простаты, внутренняя его часть содержит пенис со стилетом (рис. 3B–3С), острый кончик которого торчит наружу (рис. 2A). Отметим, что у 18 других вскрытых особей (напр., рис. 1F) паллиальные органы по ширине были в 2–2.5 раза меньше, подразделенность на белковую и капсульную железы не была выражена, стилет отсутствовал (рис. 2С); и их половую принадлежность определить не удалось. Гонада и паллиальные органы у этих особей не были изучены на гистологических срезах.
Для выяснения вопроса – является ли гермафродитизм характерной особенностью только для S. minuta – требуются дополнительные исследования. Кроме того, необходимо выяснить тип гермафродитизма (протерандрия или протерогония), место подразделения гермафродитного протока на яйцевод и семяпровод, а также существует ли у протока семяприемника самостоятельное отверстие.
Список литературы
Лилли Р., 1969. Патогистологическая техника и практическая гистохимия. Москва: Мир, 1969. 639 с.
Bouchet P., Rocroi J-P., Hausdorf B., Kaim A., Kano Y., Nützel A., Parkhaev P., Schrödl M., Strong E.E., 2017. Revised classification, nomenclator and typification of gastropod and monoplacophoran families // Malacologia, V. 61. № 1–2. P. 1–526.
Keating C., Wing D., 2007. Diving into the unknown world of soft substrate snails. https://www.geo.arizona.edu/ nyanza/pdf/2007/7%20Bitonti&Wing.pdf
Mandahl-Barth G., 1954. The anatomy and systematic position of the Tanganyika snails Syrnolopsis and Anceya // Annales du Musée Royal de L’afrihue Centrale, Sciences, Zoologiques. Tervuren, Belgium. V. 1. P. 217–221.
Strong E.E., Glaubrecht M., 2007. Anatomy and systematics of the minute syrnolopsine gastropods from Lake Tanganyika (caenogastropoda, Cerithiodea, Paludomidae) // Acta Zoologica (Stockholm). V. 88. P. 1–21.
West K., Michel E., Todd J., Brown D., Clabaugh J., 2003. The Gastropods of Lake Tanganyika. Diagnostic key, classification and notes on the fauna. Dorchester: Dorset Press. 128 p.
Wilson A.B., Claubrecht M., Meyer A., 2004. Ancient lakes as evolutionary reservoirs: evidence from the thalassoid gastropods of Lake Tanganyika // Proceedings of the Royal Society of London. B. 271. P. 529–536.
Дополнительные материалы отсутствуют.
Инструменты
Зоологический журнал