

Зоологический журнал, 2020, T. 99, № 7, стр. 745-756
Многолетняя динамика биомассы макрозообентоса в восточной части Баренцева моря (за период с 1924 по 2014 гг.)
И. Е. Манушин a, *, Н. А. Стрелкова a, П. А. Любин b, Н. Е. Журавлёва c, Д. В. Захаров a, d, В. С. Вязникова a
a Полярный филиал ФГБНУ “Всероссийский научно-исследовательский институт
рыбного хозяйства и океанографии”
183038 Мурманск, Россия
b Институт проблем экологии и недропользования АН РТ
420087 Казань, Россия
c Зоологический институт РАН
199034 Санкт-Петербург, Россия
d Мурманский морской биологический институт КНЦ РАН
183010 Мурманск, Россия
* E-mail: manushyn@gmail.com
Поступила в редакцию 25.11.2019
После доработки 15.01.2020
Принята к публикации 09.03.2020
Аннотация
Со времени начала регулярных бентосных съемок Баренцева моря ведется дискуссия о влиянии климатических изменений на бентос морских акваторий. На основе архивных данных показано изменение биомассы макрозообентоса в восточной части Баренцева моря в различные климатические периоды. В холодные периоды биомасса макрозообентоса достоверно меньше, чем в теплые. Обсуждается роль стабильности климатических условий в изменении уровня биомассы. Приведены примеры реагирования на климатические изменения некоторых массовых видов макрозообентоса.
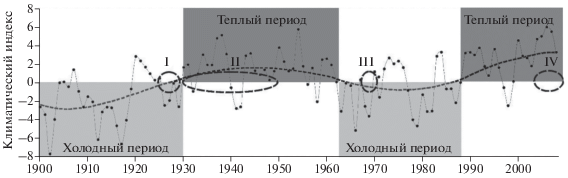
Баренцево море, расположенное на границе Бореальной и Арктической биогеографических областей, является одним из наиболее продуктивных районов арктического шельфа. Важным компонентом экосистемы этого относительно мелководного моря является бентос. Изучение жизни донной фауны необходимо как для комплексного описания экосистемы моря, так и для понимания реакции популяций бентосных видов на внешние воздействия. Опубликовано достаточно много работ о влиянии факторов среды как в отдельные годы, так и в течение многих лет на макрозообентос Баренцева моря (Несис, 1960; Галкин, 1964; Kiyko, Pogrebov, 1997; Денисенко, 2013; Manushin et al., 2013; Манушин и др., 2014 и др.). Регистрация изменений количественных показателей бентоса на протяжении ряда лет позволяет проанализировать его ответ на изменяющиеся климатические условия. Существует точка зрения, согласно которой реакция бентоса на климатические изменения происходит через изменения пополнения популяций (Несис, 1960; Денисенко, 2013). Полярный институт (ПИНРО) проводит исследования бентоса в Баренцевом море уже почти столетие. Накопленный за это время материал может быть использован для выявления воздействия климатических изменений на бентосные сообщества. Для Баренцева моря с 1900 г. были выделены четыре климатических периода (Boitsov et al., 2012) (рис. 1). Первый (I), холодный, период начался в 19 веке и длился до 1929 г. Второй (II), теплый, охватывает период с 1930 по 1962 г. Третий (III), холодный – с 1963 по 1988 г. Современный теплый период (IV) начался в 1989 г. и продолжается до сих пор (González-Pola et al., 2018). Таким образом, сравнивая качественные и количественные характеристики бентоса в каждый из этих периодов, можно оценить воздействие климатических изменений на бентосные сообщества Баренцева моря.
Рис. 1.
Климатические периоды Баренцева моря (по: Boitsov et al., 2012) и сроки проведения съемок макрозообентоса на востоке этой акватории (I–IV – порядковые номера климатических периодов и соответствующих анализируемых съемок).
Основной целью работы является оценка влияния климатических изменений на макрозообентос восточной части Баренцева моря. Проанализирована реакция отдельных видов на изменение океанографических условий в разные климатические периоды.
МАТЕРИАЛЫ И МЕТОДЫ
Для оценки влияния климата на макрозообентос, была выбрана восточная часть Баренцева моря (между 71° и 79° с.ш. и восточнее 44° в.д. до архипелага Новая Земля или до 65° в.д.), которая наименее подвержена антропогенному воздействию в отличие от остальной части (Денисенко, 2008). Бентосные данные были проанализированы по четырем климатическим периодам. Под климатом обычно понимают многолетний (порядка нескольких десятилетий) режим погоды. Соответственно, менее масштабные по продолжительности изменения в данной работе отдельно не рассматриваются. Большинство рейсов было выполнено в период гидрологического лета – с июля по сентябрь. Рейсы бентосной съемки Баренцева моря первой половины 20 века выполнялись почти каждый год, (самый длительный перерыв пришелся на годы Второй мировой войны), поэтому данные станций начала второго (теплого) периода частично отражают ситуацию в донных сообществах, сохранившуюся после первого (холодного) периода. В работе нумерация периодов бентосных съемок (I, II, III, IV) совпадает с нумерацией климатических периодов, в которые проводились рейсы.
В бентосных исследованиях первой половины 20 века использовали дночерпатели Петерсена (c площадью захвата 0.1 м2 в 5-кратной повторности на станции и площадью захвата 0.25 м2 в 2-кратной повторности на станции), а во второй половине 20 века – дночерпатели “Океан” (площадь захвата 0.25 м2 в 2-кратной повторности на каждой станции). Полученные пробы промывали через железное сито с ячеей в 1 мм, разбирали на таксономические группы на борту судна, затем консервировали спиртом для последующей лабораторной обработки.
Таблица 1.
Средний вес особей и количество видов в бентосных пробах, отобранных на востоке Баренцева моря в разные климатические периоды
| Показатель | I период 1924–1929 гг. |
II период 1930–1950 гг. |
III период 1968–1970 гг. |
IV период 2003–2014 гг. |
|---|---|---|---|---|
| Средний вес особи на станциях, г | 0.265 ± 0.033 | 0.936 ± 0.090 | 0.997 ± 0.098 | 0.252 ± 0.019 |
| Общее количество зарегистрированных видов | 141 | 184 | 153 | 701 |
В съемках ПИНРО 2003–2014 гг. пробы были получены с использованием дночерпателя ван Вина (площадь захвата 0.1 м2). На каждой станции отбирали по 5 проб, которые промывали через мягкое капроновое сито с размером ячеи 0.5 мм. Оставшийся грунт с животными фиксировали в 4% растворе формальдегида. Полученные таким образом пробы сортировали по таксономическим группам в лаборатории и после этого консервировали в 75% спирте. Организмы идентифицировали, по возможности, до видового уровня. Животные каждого таксона были просчитаны и взвешены. Биомассу определяли для организмов, фиксированных 75% этиловым спиртом. Таким образом, биомасса бентоса для всех периодов представлена в спиртовом весе.
Предварительный анализ показал, что различия в сборе и обработке материала не позволяют корректно сравнивать видовой состав, численность и, в отдельных случаях, биомассу выборок, полученных в различные периоды. Одним из показателей тщательности первичной обработки бентосных проб, собранных на одной акватории, является средний вес особей – чем меньше средний вес, тем тщательнее выбраны мелкие особи. Наиболее сходны по тщательности обработки сборы 1924–1929 и 2003–2014 гг., о чем можно судить по среднему весу особей в пробе (таблица). Наименее детально обработан материал в 1930–1950 и 1968–1970 гг., что сказалось не только на показателях численности, но и на общей биомассе животных. Так как биомасса менее всего зависит от метода и качества обработки материала. Именно этот показатель использован для анализа влияния климата на зообентос Баренцева моря. Таксономическая обработка произведена более тщательно в последний рассматриваемый период (см. таблицу).
Станционная информация (координаты, глубина, придонная температура воды) регистрировалась при отборе проб. Статистическая обработка данных по глубине и температуре проводилась по станционным данным без учета количества особей макрозообентоса на станции. В разные периоды положение станций существенно различалось (рис. 2), поэтому, для сравнения, средние значения биомассы макрозообентоса на рассматриваемой акватории рассчитывались двумя способами:
Рис. 2.
Расположение бентосных станций на востоке Баренцева моря в различные периоды исследований. Пунктирной рамкой ограничен район исследований.
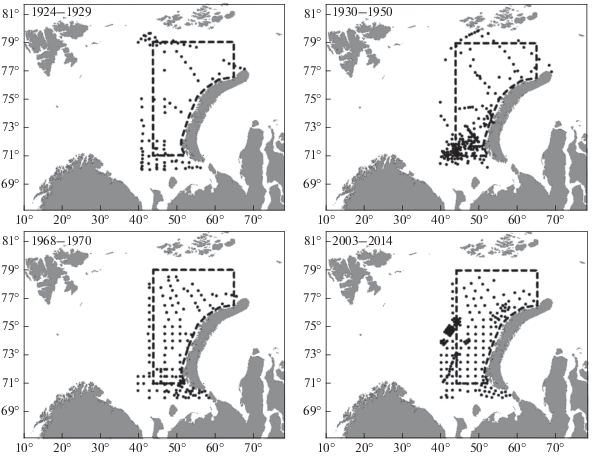
1 – по фактическим станционным данным (43 станции за период 1924–1929 гг., 150 станций за 1930–1950 гг., 80 станций за 1968–1970 гг. и 109 станций за 2003–2014 гг.; учитывались станции, расположенные только на исследуемой акватории);
2 – способом интерполяции: станционные данные конвертировались в метаданные узлов регулярной решетки (100 × 100) методом “Inverse Distance to Power” (98 станций за период 1924–1929 гг., 234 станции за 1930–1950 гг., 141 станция за 1968–1970 гг. и 218 станций за 2003–2014 гг.; в данном случае для большей точности учитывалось влияние станций, расположенных не только на исследуемой акватории, но и рядом с ее границами). Узлы, не относящиеся к рассматриваемому району, не учитывались. Средняя биомасса на исследуемой акватории рассчитывалась как средняя биомасса в оставшихся узлах интерполяционной решетки, всего в расчетах использовались экстраполированные данные 6730 узлов.
РЕЗУЛЬТАТЫ
Результаты, полученные и традиционным и интерполяционным методами, показали прямую зависимость величины общей биомассы бентоса от климатических изменений в Баренцевом море (рис. 3).
Рис. 3.
Средняя биомасса макрозообентоса на востоке Баренцева моря в разные климатические периоды, рассчитанная по фактическим (a) и интерполированным (b) данным. Черная вертикальная линия – стандартная ошибка (при интерполированных расчетах мала и скрыта толщиной маркера).
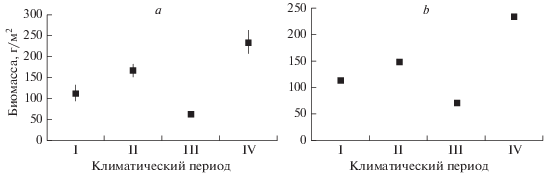
В конце холодного периода начала 20 века (I климатический период) средняя биомасса бентоса в восточной части Баренцева моря составляла 112(±20.5)/ 114(±0.6) г/м2 (традиционный/интерполяционный методы, в скобках – ошибка среднего). В последовавший за ним теплый период (II) она увеличилась до 168(±14.9)/149(±0.6) г/м2. Во второй холодный период (III) биомасса бентоса снизилась до 63(±8.1)/71(±0.3) г/м2. Современный период (IV) является самым теплым за весь ряд наблюдений в Баренцевом море. Средняя биомасса макрозообентоса в 2003–2014 гг. увеличилась до 234(±29.7)/234(±1.1) г/м2. Хотя биомассы во второй и третий периоды могут быть недооценены, тем не менее ясно, что в восточной части Баренцева моря в теплые периоды средняя биомасса макрозообентоса была от 1.5 до 3 раз выше, чем в холодные.
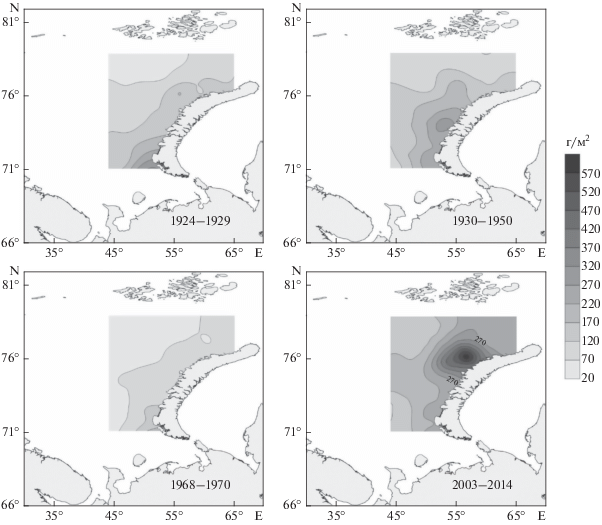
Анализ распределения общей биомассы макрозообентоса показал, что район повышенной биомассы во все климатические периоды был в основном приурочен к прибрежью архипелага Новая Земля (рис. 4). В холодные периоды самая высокая биомасса бентоса отмечается на юге исследованного района, а в теплые периоды она смещается на север.
Рис. 4.
Распределение средней биомассы макрозообентоса на востоке Баренцева моря в различные климатические периоды.
При рассмотрении изменений биогеографической структуры макрозообентоса восточной части Баренцева моря можно проследить следующие закономерности. За период исследования доля арктических и бореальных видов сократилась, доля космополитов не показала какого-либо тренда (рис. 5). В то же время доля бореально-арктических видов, составляя в начале прошлого века немногим более половины общей биомассы бентоса, неуклонно росла и в современный период составляет более 90%.
Рис. 5.
Доля видов различной биогеографической принадлежности в общей биомассе бентоса восточной части Баренцева моря за период с 1924 по 2014 г.
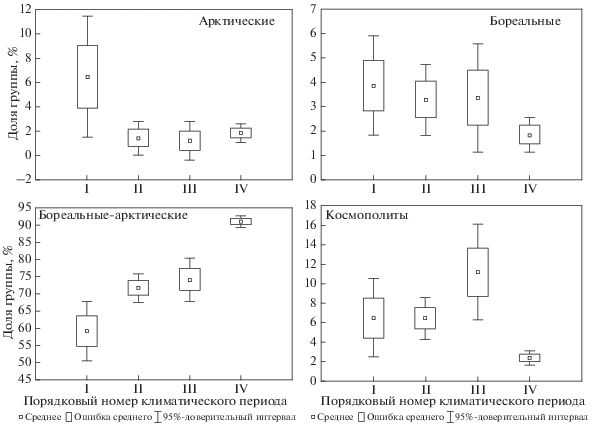
В макрозообентосе восточной части Баренцева моря доля эпифауны оставалась почти всегда неизменной (за исключением периода 1930–1950 гг.), доля инфауны была выше в теплые годы, чем в холодные, а доля онфауны была выше в холодные годы (рис. 6).
Рис. 6.
Доля видов различной жизненной формы в общей биомассе восточной части Баренцева моря за период с 1924 по 2014 г.
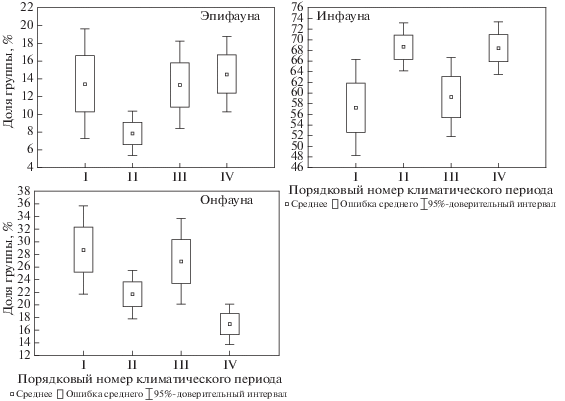
Доля детритофагов-грунтоедов в общей биомассе за все время наблюдений была неизменной, за исключением холодного периода 1963–1988 гг., когда она увеличилась вдвое, в основном за счет инфаунного вида-космополита Golfingia (Golfingia) margaritacea (Sars 1851) (рис. 7). Однако это может быть артефактом, так как эта сипункулида относится некоторыми авторами к детритофагам-собирателям. Возможно, преобладающий тип питания зависит от нескольких факторов. Доля детритофагов-собирателей была выше в теплые климатические периоды, в то время как сестонофаги стабильно составляли около трети биомассы макрозообентоса. Доля плотоядных видов также была достаточно стабильной, за исключением первого холодного периода. Это произошло из-за большого количества офиур, которые имеют смешанный тип питания (могут быть как плотоядными, так и детритофагами-собирателями). Возможно, в первый климатический период у них преобладал второй тип питания.
Рис. 7.
Доля видов с различным типом питания в общей биомассе восточной части Баренцева моря за период с 1924 по 2014 г.
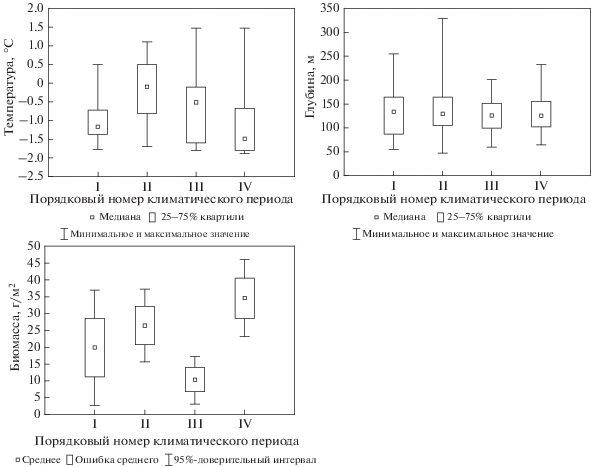
Для оценки влияния климатических изменений было выбрано 5 видов, массовое развитие которых наблюдалось в течение одного или более периодов: один – арктический и четыре – бореально-арктических вида. Одним из наиболее массовых видов на востоке Баренцева моря во все периоды наблюдений является бореально-арктический двустворчатый моллюск Macoma calcarea (Gmelin 1791). Динамика его биомассы изменялась синхронно с изменением общей биомассы макрозообентоса (рис. 8). В этой части моря он обитает примерно на одной и той же глубине (100–150 м) и при одной и той же температуре (почти всегда отрицательной) при изменении климатических условий.
Рис. 8.
Температура (°C), глубина распространения (м) и биомасса (г/м2) Macoma calcarea на востоке Баренцева моря.
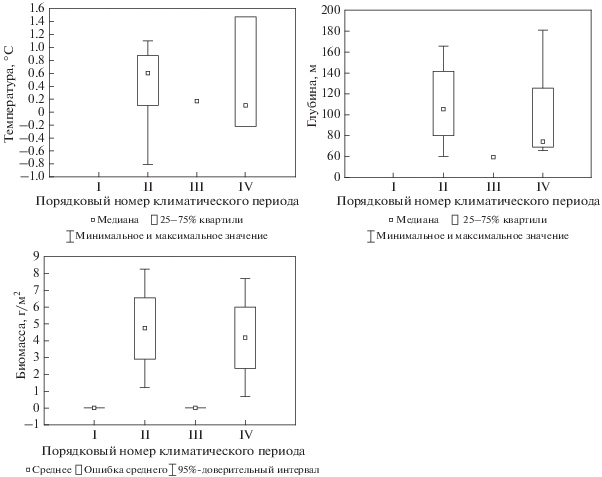
Существуют еще два бореально-арктических вида со сходной динамикой величин биомассы, но живущие в разных экологических условиях. Первый – многощетинковый червь Spiochaetopterus typicus M. Sars 1856, – значимо увеличивает свою биомассу в теплые климатические периоды и почти исчезает в холодные (рис. 9). Второй вид – двустворчатый моллюск Chlamys islandica (O. F. Müller 1776) – также становится массовым в теплые периоды и практически исчезает в холодные (рис. 10). Но если полихета встречается в основном при отрицательных температурах на глубинах от 150 до 300 м, то моллюск обитает при положительных температурах на мелководьях (от 70 до 140 м).
Рис. 9.
Температура (°C), глубина распространения (м) и биомасса (г/м2) Spiochaetopterus typicus на востоке Баренцева моря.
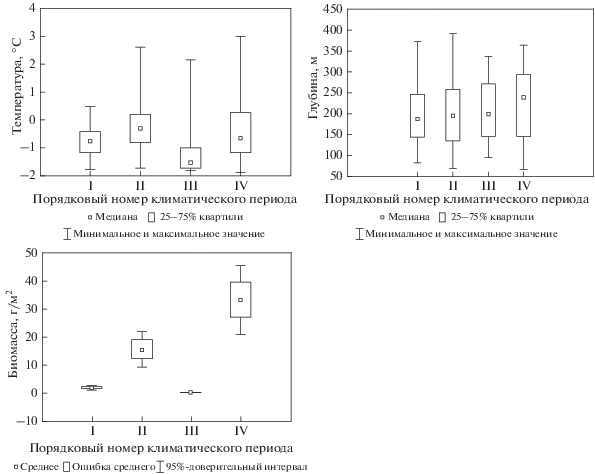
Рис. 10.
Температура (°C), глубина распространения (м) и биомасса (г/м2) Chlamys islandica на востоке Баренцева моря.
Типичную реакцию арктических видов на изменения климата можно проследить на примере двустворчатого моллюска Portlandia arctica (Gray 1824). Вид был массовым в первый период, холодный и длительный. Во время второго периода (длительное потепление) вид почти исчез на исследуемой акватории, и даже в последовавший затем холодный период его биомасса не восстанавливалась.
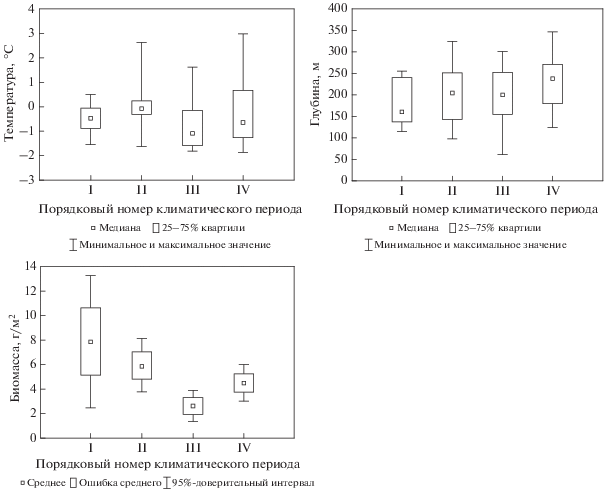
Такой массовый для Баренцева моря борельно-арктический вид, как Ctenodiscus crispatus (Bruzelius 1805), имел более низкие величины биомассы в теплые периоды, чем в первый холодный период, что отличает его от других бореально-арктических видов, рассмотренных выше (рис. 11). Глубина обитания вида за весь этот период практически не менялась, значения температуры обитания становились положительными в теплые периоды.
ОБСУЖДЕНИЕ
Во второй половине прошлого и начале текущего веков было предпринято несколько попыток оценки и объяснения реакции бентоса Баренцева моря на климатические изменения.
Несис, изучая динамику встречаемости бореальных и арктических видов бентоса на разрезе “Кольский меридиан”, пришел к выводу о том, что “… реакция донной фауны на изменение температурных условий происходит с некоторым запозданием. Может быть, это связано с тем, что донная фауна реагирует не на температурные условия одного какого-нибудь года (как было указано выше, такой связи мы не обнаружили), а на тенденцию повышения или понижения температуры в течение нескольких лет подряд” (Несис, 1960; с. 135–136).
Антипова (1975) объясняет снижение биомассы в 1968–1970 гг. по сравнению с данными довоенной съемки похолоданием вод Баренцева моря.
На основании исследований 1991–1994 гг. Кийко и Погребов (Kiyko, Pogrebov, 1997) констатируют, что в начале 1990-х годов общая биомасса бентоса не отличалась значимо от таковой 1920–1930-х годов, но была существенно меньше в конце 1960-х годов. Так как съемка конца 1960-х годов приходится на холодный период, съемка 1920–1930-х годов – на переходный от холодного к теплому, а съемка начала 1990-х приурочена к началу теплого периода, то данное исследование также подтверждает тенденцию увеличения средней биомассы бентоса в Баренцевом море в теплые периоды.
Большое внимание вопросу влияния климата на донную фауну Баренцева моря уделил Денисенко (2003, 2007, Denisenko, 2001). В первой из этих работ (Denisenko, 2001) автор утверждает, что основным фактором, влияющим на биомассу макрозообентоса в Баренцевом море, является промысел рыбы донными тралами, а температурный фактор выходит на первое место только в тех районах, где траловый промысел отсутствует.
Сопоставляя изменения температуры и биомассы макрозообентоса на разрезе “Кольский меридиан”, Денисенко констатирует: “Анализ количественного распределения и видовой структуры сообществ зообентоса в различные периоды исследований вдоль разреза не выявил каких-либо закономерностей изменений общей биомассы в зависимости от температурных условий” (Денисенко, 2003; с. 70).
В другой своей работе Денисенко (2007) приходит к выводу, что имевшие место флуктуации донного населения в Печорском море (юго-восток Баренцева) были вызваны естественными причинами преимущественно климатического характера. Выявленное им увеличение общей биомассы зообентоса в Печорском море он связывает с усилением циркуляции водных масс при потеплении, которое должно сопровождаться общим повышением биопродуктивности морских экосистем.
При описании состояния зообентоса в прибрежье архипелага Новая Земля (район исследования примерно совпадает с нашим) Денисенко высказывает мнение, что “способность климатических изменений существенно сказываться на суммарной биомассе бентоса, по нашему мнению, маловероятна” (Денисенко, 2013, с. 198). Следует, однако, отметить, что перед этим высказыванием было написано нечто противоположное: “Тем не менее понижение биомассы в конце 60-х годов могло быть вызвано именно предшествовавшим сильным похолоданием” (Денисенко, 2013, с. 195).
Результаты нашего исследования показывают, что в теплые климатические периоды величина общей биомассы макрозообентоса на востоке Баренцева моря достоверно выше, чем в холодные. При выявлении возможной причины этого мы, также как и Денисенко, считаем, что в теплые годы происходит повышение биопродуктивности Баренцева моря (за счет увеличения первичной продукции). Период с более высокой продуктивностью, длящийся несколько лет подряд, дает возможность особям, пережившим похолодание, не только повысить уровень воспроизводства, но и с низкой популяционной смертностью дожить до взрослого состояния, что повышает общую биомассу.
Также нельзя исключить существование связи между величиной биомассы и стабильностью условий, предшествовавших съемке. Так, съемке 1924–1929 гг. предшествовали несколько десятилетий холодных лет, в течение которых донная фауна имела возможность успешно адаптироваться к климатической ситуации. В результате этого показатели средней биомассы оказались почти в два раза выше, чем в аналогичный холодный период съемки 1968–1970 гг., которому, однако, предшествовало длительное потепление Арктики 1930–1960-х годов. Аналогично, станции 1930– 1950-х годов первого теплого периода были выполнены, в основном, сразу после холодного периода, вследствие чего они показали биомассу в полтора раза меньшую, чем станции второго теплого периода, отбору которых предшествовало 15 теплых лет.
При рассмотрении изменения вклада особей разных биогеографических группировок в величину общей биомассы можно отметить увеличение за весь период наблюдения доли бореально-арктических видов при одновременном уменьшении доли бореальных видов (хотя отчасти его можно объяснить некорректной видовой идентификацией в первые периоды исследования). Также обращает на себя внимание уменьшение в 3 раза доли арктических видов в общей биомассе после завершения в 1929 г. длительного холодного периода.
При рассмотрении изменений долей различных жизненных форм в общей биомассе бентоса в восточной части Баренцева моря в различные климатические периоды статистически значимых изменений не выявлено.
Анализ изменений в трофической структуре выявил увеличение доли детритофагов-собирателей в теплые периоды. Возможной причиной этого является повышение первичной продукции и, соответственно, нарастание массы органического вещества, поступающего на дно. Также была отмечена тенденция к увеличению доли детритофагов-собирателей за весь период наблюдений.
При рассмотрении влияния климатических изменений на бентос необходимо учитывать воздействие этих изменений на отдельные массовые виды. Наибольшую биомассу в районе исследования во все периоды образует двустворчатый моллюск Macoma calcarea. Его биомасса увеличивается в теплые годы. При изучении его экологии можно видеть, что он во все периоды занимает одни те же глубины и наиболее часто встречается при отрицательных температурах. Исключением является второй, теплый, период, когда на станциях с положительной придонной температурой были зарегистрированы особи, оставшиеся после длительного периода похолодания. Повышение в теплые периоды биомассы вида, который постоянно обитает при отрицательных температурах, может свидетельствовать в пользу предположения о влиянии именно трофических условий на изменение биомассы.
Еще два массовых для Баренцева моря вида – седентарная полихета Spiochaetopterus typicus и двустворчатый моллюск Chlamys islandica – имеют сходные с макомой изменения в биомассе, с той лишь разницей, что в холодные периоды эти виды практически полностью исчезают из донных сообществ. Представляется интересным, что показатели биомассы и глубоководного холодноводного и мелководного тепловодного видов синхронно увеличиваются в теплые периоды и уменьшаются в холодные. Т.е. виды с противоположными, по сути, экологическими предпочтениями одинаково реагируют на изменения климата. Такие результаты могут помочь как при интерпретации изменений, происходящих в донных биоценозах, так и при моделировании будущих сценариев влияния изменений климата на сообщества бентоса.
ВЫВОДЫ
В восточной части Баренцева моря биомасса макрозообентоса в теплые климатические периоды выше, чем в холодные. Вторым по значимости фактором может являться стабильность условий: чем продолжительнее часть климатического периода перед съемкой, тем отчетливее проявляется прямая связь между фазой климатического индекса и величиной биомассы макрозообентоса. Динамики биомасс тепловодных и холодноводных видов могут показывать одинаковые тренды при изменениях климата.
Список литературы
Антипова Т.В., 1975. Распределение биомассы бентоса Баренцева моря // Труды ПИНРО. Вып. 35. С. 121–124.
Галкин Ю.И., 1964. Многолетние изменения в распределении двустворчатых моллюсков в южной части Баренцева моря // Новые исследования планктона и бентоса Баренцева моря. Труды ММБИ. Вып. 6 (10). С. 22–40.
Денисенко С.Г., 2003. Многолетние изменения донной фауны Баренцева моря и гидрологические флюктуации вдоль разреза “Кольский меридиан” // Материалы симпозиума, посвященного 100-летию океанографических наблюдений на разрезе “Кольский меридиан” (Мурманск, 1999). Мурманск: Изд-во ПИНРО. С. 65–76.
Денисенко С.Г., 2007. Зообентос Баренцева моря в условиях изменяющегося климата и антропогенного воздействия // Динамика морских экосистем и современные проблемы сохранения биологического потенциала морей России. Владивосток: Дальнаука. С. 418–511.
Денисенко С.Г., 2008. Макрозообентос Баренцева моря в условиях меняющегося климата и антропогенного воздействия. Дис. … докт. биол. наук. СПб. ЗИН. 465 с.
Денисенко С.Г., 2013. Биоразнообразие и биоресурсы макрозообентоса Баренцева моря: Структура и многолетние изменения. СПб.: Наука. 284 с.
Манушин И.Е., Анисимова Н.А., Любин П.А., Вязникова В.С., Захаров Д.В., 2014. Изменения в биомассе макрозообентоса в восточной части Баренцева моря за последнее столетие // Чтения памяти Дерюгина (Материалы XVI научного семинара “Чтения памяти К.М. Дерюгина” (СПбГУ, 06.12.2013). СПб.: СПбГУ, каф. ихтиологии и гидробиологии. С. 52–59.
Несис К.Н., 1960. Изменения донной фауны Баренцева моря под влиянием колебаний гидрологического режима // Советские рыбохозяйственные исследования в морях Европейского Севера. М.: Рыбное хозяйство. С. 129–138.
Boitsov V.D., Karsakov A.L., Trofimov A.G., 2012. Atlantic water temperature and climate in the Barents Sea, 2000–2009. ICES Journal of Marine Science. V. 69. Iss. 5. P. 833–840.
Denisenko S.G., 2001. Long-term changes of zoobenthos biomass in the Barents Sea // Zoological sessions (Annual reports 2000). Proceedings of the Zoological Institute RAS. V. 289. P. 59–66.
González-Pola C., Larsen K. M. H., Fratantoni P., Beszczynska-Möller A., Hughes S. L. (Eds), 2018. ICES Report on Ocean Climate 2016. ICES Cooperative Research Report. № 339. 110 pp. [Электронный ресурс]. Режим доступа: https://doi.org/10.17895/ices.pub.4069 Дата обновления 20.06.2018.
Kiyko O.A., Pogrebov V.B., 1997. Long-term benthic population changes (1920–1930-s – present) in the Barents and Kara seas // Marine Pollution Bulletin. V. 35. № 7–12. P. 322–332.
Manushin I.E., Strelkova N.A., Lubin P.A., Vyaznikova V.S., Zakharov D.V., 2013. Changes in biomass of macrozoobenthos in the eastern part of the Barents Sea // ICES Annual Science Conference 2013. Conference proceedings. International Council for the Exploration of the Sea. B-16. [Электронный ресурс]. Режим доступа: http://www.ices.dk/sites/pub/CM%20Doccuments/ CM-2013/Theme%20Session%20B%20contributions/ B1613.pdf Дата обновления 27.06.2018.
Дополнительные материалы отсутствуют.
Инструменты
Зоологический журнал