

Зоологический журнал, 2020, T. 99, № 7, стр. 772-783
Разнообразие и многолетняя динамика макробентоса в мангровых посадках и естественных ассоциациях провинции Кхань Хоа, Вьетнам
С. С. Звонарева a, *, Ю. И. Кантор a, Т. Т. Х. Нгуйен b, Т. А. Бритаев a
a Институт проблем экологии и эволюции им. А.Н. Северцова РАН
119071 Москва, Россия
b Совместный Российско-Вьетнамский тропический центр
650000 Ня Чанг, Вьетнам
* E-mail: sonyazv@mail.ru
Поступила в редакцию 10.11.2019
После доработки 17.01.2020
Принята к публикации 09.03.2020
Аннотация
Стремительное сокращение площадей, занимаемых мангровыми лесами, убедило Вьетнамское правительство в необходимости программы по сохранению и восстановлению мангровых экосистем по всей стране. Одним из проектов этой программы стал эксперимент по высадке саженцев Rhizophora apiculata в заливе Дам Бай в провинции Кхань Хоа. В ходе настоящей работы был проведен мониторинг состояния макробентоса в новых мангровых посадках, а также было произведено сравнение его количественных и качественных характеристик с таковыми в естественных мангровых ассоциациях этого района. Основной объем материала был собран в трех горизонтах литорали в районе мангровых посадок в заливе Дам Бай. Кроме того, были исследованы естественные мангровые ассоциации: одна – также в заливе Дам Бай, а другая (самая зрелая и большая ассоциация района) – в соседнем заливе Ня Фу. Естественная мангровая ассоциация в заливе Дам Бай была представлена узким поясом Rhizophora apiculata. Верхний горизонт в районе мангровых посадок также представлен узким поясом Rhizophora apiculata. До 2004 года средний и нижний горизонты были образованы илистой литоралью. В 2004 году в среднем горизонте ниже взрослых деревьев верхнего горизонта были высажены мангровые саженцы. По мере роста саженцы стали замедлять приливно-отливные течения, и биогенные элементы стали задерживаться в экосистеме. Развивающаяся крона молодых деревьев начала давать опад и тень, благоприятствующую развитию микроводорослей, кроме того, молодые деревья образовали твердый субстрат. Вследствие этого произошли рост видового богатства макробентосных видов-эврибионтов и рост обилия видов-оппортунистов. Молодые деревья в среднем горизонте по мере роста образовали единый мангровый лес большой площади вместе со взрослыми деревьями верхнего горизонта, что стало причиной появления специализированных видов брюхоногих и двустворчатых моллюсков, а также крабов. В течение периода мониторинга состав и структура макробентоса мангровых посадок все больше приближались к таковым в зрелой естественной мангровой ассоциации в Ня Фу, но после десяти лет наблюдений за ростом мангровых саженцев между этими ассоциациями сохраняются существенные различия: макробентос Ня Фу представлен бóльшим числом специализированных мангровых видов брюхоногих моллюсков, а также он имеет совершено другую структуру таксоцена двустворчатых моллюсков. Вероятно, это связано с высоким флористическим разнообразием в Ня Фу и со зрелостью этой мангровой ассоциации. В результате качественных и количественных сборов макробентоса в трех исследованных мангровых ассоциациях было найдено 139 видов беспозвоночных. Ключевыми группами макробентоса стали брюхоногие, двустворчатые моллюски и крабы. Полученные данные о составе и разнообразии макробентоса вписываются в общую картину мангровой фауны Индо-Вест-Пацифики и Юго-Восточной Азии.
Мангровые заросли произрастают на границе моря и суши на побережьях тропического и экваториального пояса. Мангры Юго-Восточной Азии представляют наибольший интерес, поскольку они занимают большие площади и обладают более высоким разнообразием флоры и фауны по сравнению с другими регионами (Ellison et al., 1999). В последние десятилетия площади мангровых лесов повсеместно сокращаются и Вьетнам не исключение (Walters et al., 2000). Ранее мангры Вьетнама занимали около 4000 км2, но гербициды и напалм, использованные во время войны (1962–1971 гг.), уничтожили около 1050 км2 (Hong, San, 1993). Популяционный и экономический рост после Вьетнамской войны также привели к значительному сокращению площадей, занимаемых мангровыми лесами. Развитие креветочных и крабовых хозяйств, строительство соляных прудов, сельское хозяйство, добыча угля и загрязнение прибрежных вод нефтью приводит к тому, что многие ассоциации деградируют и местами превращаются в пустоши (Hong, San, 1993). Большая часть сохранившихся мангров расположена в дельте Меконга, и даже они представляют собой преимущественно вторичные леса и посадки (Spalding et al., 1997).
Однако, давно известно, насколько велика роль мангровых лесов для функционирования других экосистем прибрежных вод и для экономики стран побережья. Мангровые заросли служат местообитанием богатой фауны беспозвоночных, рыб, рептилий и птиц, а также поддерживают продуктивность и других прибрежных сообществ, обеспечивая пищей и местом для роста личинок и молоди многих, в т.ч. промысловых видов рыб и беспозвоночных (Kathiresan, Bingham, 2001; Twilley et al., 1992). Они стабилизируют береговую линию, фильтруют пресные стоки, а также значительно снижают ущерб от тайфунов и цунами (Mazda et al., 2002; Mazda et al., 2006). Мангры могут являться объектом экотуризма (Ellison, 2008). Кроме того, по всему побережью Вьетнама распространены традиционные формы использования мангров: в качестве стройматериала, топлива, как место добычи пищи и т.д. (Hong, San, 1993).
В связи с этим, на государственном уровне осознана необходимость сохранения и восстановления мангровых лесов. Вьетнамское правительство создало несколько национальных парков и охраняемых зон, а также искусственных мангровых посадок (Field 1998; Hong, San, 1993).
В настоящее время собран большой объем информации, опубликовано множество работ и обзоров, посвященных отдельным направлениям изучения мангров.
Мангровые деревья образуют среду обитания для разнообразной фауны беспозвоночных. Ключевыми группами беспозвоночных в мангровой экосистеме являются моллюски и крабы, имеющие высокое разнообразие, плотность поселения и дающие значительную биомассу. Кроме того, функционирование экосистемы в значительной степени зависит от жизнедеятельности этих групп (Berry, 1963; Kathiresan, Bingham, 2001; Lee, 1998, 1999; Macnae, 1968).
Живущие здесь виды беспозвоночных имеют разную приуроченность к мангровой экосистеме, есть как специализированные виды, встречающиеся только в манграх, так и виды-эврибионты, т.е. виды, приспособленные к широкому спектру условий и населяющие разные типы экосистем. Специализированными видами брюхоногих моллюсков является большинство представителей семейства Potamididae, род Littoraria (Littorinidae), некоторые представители семейств Neritidae, Cerithiidae, Assimineidae и др. (Frey, 2010; Houbrick, 1985; Houbrick, 1992; Reid, 1985, 1986; Reid et al., 2008; Reid et al., 2010; Reid, 2014). Среди двустворчатых моллюсков это представители рода Geloina, некоторые представители семейства Lucinidae и др. (Glover, Taylor, 2016; Kathiresan, Bingham, 2001; Morton, 1976). Для крабов нет детальной информации о специализированных мангровых видах, но известно, что высокое разнообразие и обилие представителей надсемейства Grapsoidea характерны для зрелых и ненарушенных мангров, тогда как высокое обилие видов надсемейства Ocypodoidea присуще молодым и нарушенным мангровым экосистемам (Macintosh et al., 2002).
Кроме трех ключевых групп макробентоса, мангровые ассоциации населяют представители и других таксонов. В манграх различных районов Индо-Вест Пацифики встречаются медузы (Tsingalia, 2014), роющие актинии (Mitra, Pattanayak, 2013), аннелиды (Metcalfe, Glasby, 2008), сипункулиды (Adrianov, Maiorova, 2012; Gopal, Chauhan, 2006; Macnae, 1968;), раки-отшельники (Gherardi, Vannini, 1993; Kathiresan, Bingham, 2001; Macnae, 1968), раки-богомолы (Ng, 2013), креветки (Kathiresan, Bingham, 2001; Macnae, 1968; Yu et al., 1997), брахиоподы (Mitra, Pattanayak, 2013a; Printrakoon, Kamlung-ek, 2013), кишечнодышашие (Dittmann, 2002; Kumar, 2000) и др.
Публикаций, касающихся различных аспектов мангровых экосистем Вьетнама, сравнительно немного. Кроме того, значительная их часть написана на вьетнамском языке, потому малодоступна. Фауна вьетнамских мангров изучена очень плохо. Хонг и Сан (Hong, San, 1993) приводят данные работ по крабам, полихетам и моллюскам; Чертопруд и др. (Chertoprud et al., 2012) анализируют разнообразие и распределение крабов, найденных в манграх в окрестностях Ня Чанга. Опубликованы таксономические работы о креветках из вьетнамских мангров (Anker et al., 2006; Dworschak et al., 2006; Marin, 2008; Marin, 2014). Кроме этого, есть ряд исследований, посвященных мейобентосу (Mokievsky et al., 2011; Tchesunov et al., 2010; Xuan et al., 2007).
Разнообразие и структура фауны беспозвоночных изменяются под влиянием нарушений, эксплуатации, менеджмента или восстановления мангровых зарослей. Таким образом, эти параметры могут отражать статус мангровой ассоциации и могут быть использованы в качестве индикатора изменений, как в естественных мангровых лесах, так и в посадках (Macintosh et al., 2002).
В 2005 г. в бухте Дам Бай (о-в. Че, залив Ня Чанг) стартовал эксперимент по высадке саженцев Rhizophora apiculata Blume. Целью работы было проследить качественные и количественные изменения макробентоса в посадках с течением времени, а также сравнить макробентос посадок и естественных мангровых ассоциаций. Нами проведен десятилетний мониторинг фауны макробентоса в трех горизонтах литорали в районе мангровых посадок залива Дам Бай, состояние макробентоса этих горизонтов соотнесено с состоянием ближайших естественных мангровых ассоциаций. В рамках этого масштабного проекта уже опубликованы некоторые данные, касающиеся брюхоногих моллюсков. В статье Звонаревой и др. (Zvonareva et al., 2016) приведен полный список брюхоногих моллюсков, кроме того, опубликована часть данных о количественных характеристиках брюхоногих до 2012 года (Zvonareva et al., 2015).
Данное исследование охватывает все ключевые группы макробентоса естественных и искусственных мангровых зарослей центрального Вьетнама, которые не часто попадают в поле зрения исследователей. Впервые зафиксированы все этапы становления сообщества макробентосных беспозвоночных в мангровых посадках с момента высадки саженцев до их десятилетнего возраста, что не имеет аналогов в литературе.
МАТЕРИАЛЫ И МЕТОДЫ
Берег центрального Вьетнама с заливами Ня Чанг и Ня Фу (рис. 1) часто испытывает штормовое воздействие, поэтому мангровые заросли здесь встречаются только в хорошо защищенных бухтах и лагунах (Hong, San, 1993). Но и в заливах Ня Чанг и Ня Фу мангры скудны и часто находятся в плохом состоянии из-за антропогенного влияния. На данный момент они присутствуют вокруг марикультурных прудов в устье реки Бэ в границах города Ня Чанг и в куту залива Ня Фу. Самая крупная естественная мангровая ассоциация без марикультурных хозяйств расположена на северо-восточном побережье залива Ня Фу. Кроме того, мангры, представленные узкими поясами растительности, присутствуют на островах залива Ня Фу и во внутренней части залива Дам Бай.
Рис. 1.
Карта исследованных районов (по: Zvonareva et al., 2015): а – карта района исследования с указанием его местоположения на карте Вьетнама, б – схема мангровой ассоциации в заливе Ня Фу, в – спутниковый снимок естественной мангровой ассоциации и мангровых посадок в заливе Дам Бай (2014 г.), г – схема мангровых посадок в заливе Дам Бай с указанием расположения исследованных станций.
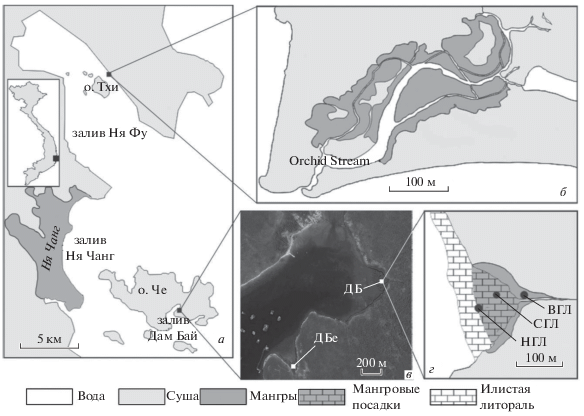
В заливе Дам Бай (о-в Че, залив Ня Чанг, рис. 1) до высадки мангровых саженцев верхний горизонт в небольшом углублении береговой линии восточного побережья залива был занят узким поясом мангровых деревьев Rhizophora apiculata с небольшой примесью Avicennia marina (Forsk.) Vierh. (координаты: 12°13.116′ с.ш., 109°18.230′ в.д.). В 2004 г. в среднем горизонте песчано-илистой литорали (12°12.097′ с.ш., 109°18.213′ в.д.) были высажены саженцы Rhizophora apiculata, а затем в 2007 г. – саженцы высажены и в нижнем горизонте литорали (12°12.086′ с.ш., 109°18.166′ в.д.).
Материал собирали не только в мангровых посадках, но и в двух естественных ассоциациях. Южнее посадок в заливе Дам Бай в небольшой бухте расположена естественная мангровая ассоциация (координаты: 12°11.858′ N; 109°17.949′ E, рис. 1), которая по типу и размеру соответствует верхнему горизонту в районе посадок до высадки саженцев в среднем и нижнем горизонтах. Кроме того, была исследована крупная естественная ассоциация в заливе Ня Фу рядом с парком Orchid Stream (станция НФ, координаты: 12°24.362′ N; 109°14.407′ E рис. 1). Она отличается от мангров в заливе Дам Бай структурой растительности, деревья здесь растут вдоль ручья, который несколько распресняется стоками с гор (до 18 ‰ в верховьях). Мангровые деревья представлены семью видами с доминированием видов семейства Rhizophoreacea в низовьях и Xylocarpus granatum и Excoecaria agallocha в верховьях. Эта ассоциация подвергается антропогенному влиянию, поскольку расположена рядом с рекреационным парком.
Начиная с 2005 г., в районе посадок проводили сбор количественных проб. Использовали разные орудия лова – дночерпатели, трубки и рамки различной площади, но в большинстве случаев использовали рамку площадью 0.11 м2. Количество проб на станции так же варьировало от года к году, в большинстве случаев собирали по четыре пробы. Из рамки извлекали грунт на глубину 15–20 см, затем отделяли животных от грунта путем отмучивания в ведре. Оставшуюся крупную фракцию осадка промывали через сито с размером ячеи 3 мм. Начиная с 2014 г., грунт из рамки площадью 0.11 м2 сразу помещали на сито, а отмучивали только грунт из маленькой рамки площадью 0.01 м2. Из полученных проб животных выбирали в лаборатории в Ня Чанге. Червеобразных животных фиксировали 4% формалином, остальных – 75% спиртом. Для каждой количественной пробы затем были подсчитаны плотность и биомасса.
Помимо количественных проб, собирали и качественные пробы со стволов и корней деревьев, также собирали крупных и редко встречающихся животных (ручной сбор). В табл. 1 приведен список станций и площадей проб в заливах Дам Бай и Ня Фу.
Таблица 1.
Список проведенных мангровых съемок
| Время работ | Метод отбора проб | |||
|---|---|---|---|---|
| Мангровые посадки в заливе Дам Бай | ||||
| ВГЛ | СГЛ | НГЛ | ||
| Октябрь 2005 | 3 × 0.11 | 3 × 0.11 | 3 × 0.11 | |
| Ноябрь 2006 | 5 × 0.0225 | 5 × 0.0225 | 5 × 0.0225 | |
| Октябрь 2008 | 3 × 0.11 | 2 × 0.11, 3 × 0.026 | 3 × 0.025 | |
| Май 2009 | 3 × 0.11 | 3 × 0.11 | 3 × 0.11 | |
| Октябрь 2009 | 4 × 0.11 | 4 × 0.11 | 4 × 0.11 | |
| Май 2010 | 4 × 0.11 | 4 × 0.11 | 4 × 0.11 | |
| Октябрь 2010 | 4 × 0.11 | 4 × 0.11 | 3 × 0.11 | |
| Май 2011 | 4 × 0.11 | 4 × 0.11 | 4 × 0.11 | |
| Октябрь 2011 | 4 × 0.11 | 4 × 0.11 | 4 × 0.11 | |
| Сентябрь 2012 | 4 × 0.11 | 4 × 0.11 | 4 × 0.11 | |
| Май 2013 | 8 × 0.11 | 7 × 0.11 | 4 × 0.11 | |
| Май 2014 | 4 × 0.11, 4 × 0.01 | 4 × 0.11, 4 × 0.01 | 4 × 0.11, 4 × 0.01 | |
| Октябрь 2014 | 4 × 0.11, 4 × 0.01 | 4 × 0.11, 4 × 0.01 | 4 × 0.11, 4 × 0.01 | |
| Октябрь 2015 | 4 × 0.11, 4 × 0.01 | 4 × 0.11, 4 × 0.01 | 4 × 0.11, 4 × 0.01 | |
| Естественные мангровые ассоциации | ||||
| ДБе | НФ | |||
| Сентябрь 2012 | – | 4 × 0.11 | ||
| Май 2013 | 7 × 0.11 | кач. | ||
| Май 2014 | 4 × 0.11, 4 × 0.11 | кач. | ||
| Октябрь 2014 | 4 × 0.11, 4 × 0.11 | кач. | ||
| Октябрь 2015 | 4 × 0.11, 4 × 0.11 | 3 × 0.11, 3 × 0.01 | ||
Примечания. Указаны месяц и год проведения полевой работы, станция и метод отбора количественных проб (число проб × × площадь пробы, м2), отмечен сбор качественных проб. ВГЛ – верхний горизонт литорали в районе мангровых посадок, СГЛ – средний горизонт литорали посадок, НГЛ – нижний горизонт литорали посадок, ДБе – естественная мангровая ассоциация в заливе Дам Бай, НФ – естественная мангровая ассоциация в заливе Ня Фу, кач. – в съемке собраны только качественные пробы, во всех остальных случаях качественные пробы были собраны в дополнение к количественным.
В данную работу было решено не включать данные по типу Annelida, поскольку это очень сложная для видовой идентификации группа, определение требует больших трудозатрат и их обработка не входила в задачи данной работы.
Все собранные коллекции хранятся в лаборатории морфологии и экологии морских беспозвоночных ИПЭЭ РАН.
РЕЗУЛЬТАТЫ
В исследованных нами мангровых ассоциациях было найдено 139 видов макробеспозвоночных (исключая Annelida, видовая идентификация которых оказалась фактически невозможной). Наибольшим разнообразием обладали брюхоногие и двустворчатые моллюски и крабы, другие же группы представлены 1–2 видами.
На рис. 3 показано видовое богатство групп макробентоса на исследованных станциях.
Брюхоногие моллюски
Брюхоногие моллюски – самая богатая группа, представленная 64 видами из 24 семейств. Наиболее разнообразными семействами были Neritidae (8 видов), Cerithiidae (7 видов), Potamididae (6 видов), Littorinidae (6 видов), Onchidiidae (6 видов) и Ellobiidae (5 видов).
Брюхоногие моллюски были наиболее разнообразной группой на всех станциях, кроме ниженго горизонта литорали. В мангровых посадках было найдено 45 видов, причем верхний и средний горизонты имели 27 и 28 видов соответственно, а нижний горизонт несколько меньше – 22 вида. В естественной ассоциации в Ня Фу было найдено 35 видов, а в естественной ассоциации в Дам Бае – 27.
Специализированными можно назвать 17 из всех найденных видов. Наиболее богата ими естественная ассоциация в Ня Фу (16 видов, см. рис. 2). В посадках самым бедным был нижний горизонт – всего 3 специализированных вида, а верхний и средний имели 10 и 11 видов соответственно. Следует отметить, что в последней съемке в среднем горизонте посадок найдено два специализированных мангровых вида: Terebralia sulcata (Born 1778) и Cassidula nucleus (Gmelin 1791), которые до этого не встречались в посадках, но присутствовали во всех сборах из естественной мангровой ассоциации в Ня Фу. В естественной мангровой ассоциации в Дам Бае обнаружено 8 специализированных мангровых видов.
Рис. 2.
Видовое богатство макробентосных беспозвоночных (кроме аннелид) в трех горизонтах литорали мангровых посадок и в двух естественных мангровых ассоциациях, найденных за весь период исследований. ВГЛ – верхний горизонт литорали в районе мангровых посадок в заливе Дам Бай, СГЛ – средний горизонт литорали в посадках, НГЛ – нижний горизонт литорали в посадках, ДБе – естественная мангровая ассоциация в заливе Дам Бай, НФ – естественная мангровая ассоциация в заливе Ня Фу.
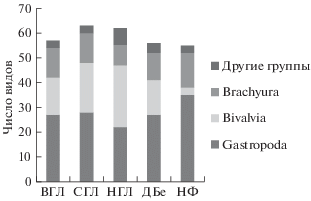
Рис. 3.
Количество специализированных мангровых видов и неспециализированных видов-эврибионтов в мангровых посадках с 2005 по 2015 гг. и двух естественных мангровых ассоциациях.
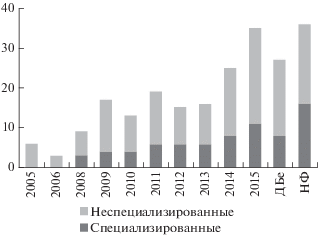
На рис. 3 видно, что во всех мангровых ассоциациях преобладают эврибионтные виды, характерные, в основном, для эпифауны мягких грунтов литорали. Однако в естественной мангровой ассоциации в Дам Бае значительную долю видов составляют виды-эврибионты, характерные для каменистой литорали, например, Pictocolumbella ocellata (Link 1807), Planaxis sulcatus (Born 1780) и т.д. В нижнем горизонте большинство найденных моллюсков – виды, характерные для сублиторали, например, Lataxiena blosvillei (Deshayes 1832), представители семейств Rannelidae, Cypraeidae и др.
Видовое богатство в мангровых посадках менялось с течением времени. На рис. 4 проиллюстрирован рост разнообразия в трех горизонтах. На рис. 3 видно, что этот рост происходит как за счет специализированных видов, так и за счет видов-эврибионтов.
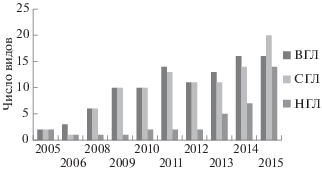
Рис. 4.
Динамика видового богатства брюхоногих моллюсков в трех горизонтах литорали в районе мангровых посадок.
По количественным характеристикам брюхоногие моллюски, опять же, лидируют среди других групп беспозвоночных на всех станциях, кроме нижнего горизонта литорали. В верхнем и среднем горизонтах литорали показатели плотности и биомассы имеют ярко выраженную тенденцию к росту (рис. 5). Обилие и биомасса в естественной мангровой ассоциации в Дам Бае были заметно ниже показателей верхнего и среднего горизонтов литорали в поздний период наблюдений. Нижний же горизонт был крайне беден брюхоногими моллюсками на протяжении всего периода наблюдений.
Рис. 5.
Динамика средней плотности (экз./м2) и биомассы (г/м2) брюхоногих моллюсков в трех горизонтах мангровых посадок и в двух естественных ассоциациях.
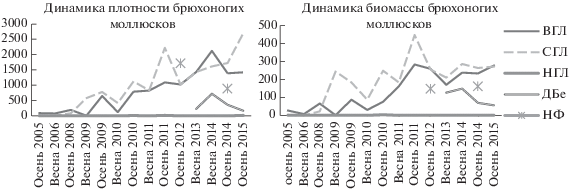
Обилие и биомасса брюхоногих моллюсков в некоторых съемках имели очень высокие значения. Так, максимальная плотность брюхоногих зарегистрирована в 2015 г. в среднем горизонте посадок – 2698 ± 1127 экз./м2, максимальная биомасса зарегистрирована осенью 2011 г. – 447 ± ± 318 г/м2.
На всех станциях доминирующими видами были эврибионты и виды-оппортунисты Battilaria sp., Clithon oualaniense (Lesson 1831) и Pirenella cingulata (Gmelin 1791). В некоторых съемках на позднем этапе мониторинга посадок их суммарный вклад в плотность (рис. 6а) и биомассу превышает 90%. Рост численности и биомассы в верхнем и среднем горизонтах литорали также происходит исключительно благодаря этим трем видам, способным образовывать очень плотные скопления (рис. 6б).
Рис. 6.
Массовые виды брюхоногих моллюсков в посадках и естественных мангровых ассоциациях: а – процентное соотношение плотности массовых видов брюхоногих моллюсков в двух верхних горизонтах литорали в районе мангровых посадок в Дам Бае (нижний горизонт не приводится, поскольку обилие брюхоногих моллюсков здесь ничтожно) и в двух естественных мангровых ассоциациях на момент последней съемки (для Ня Фу – 2014 г., для остальных станций – 2015 г.); б – скопление брюхоногих моллюсков Battilaria sp., Clithon oualaniense и Pyrenella cingulata в среднем горизонте литорали.

Двустворчатые моллюски
В ходе работы было найдено 43 вида двустворчатых моллюсков, относящихся к 18 семействам. Эта группа по видовому богатству следует за брюхоногими моллюсками на всех станциях, кроме нижнего горизонта литорали, где разнообразие двустворчатых моллюсков превышает разнообразие брюхоногих, и Ня Фу, где двустворчатых моллюсков крайне мало.
Наиболее богат в видовом отношении из исследованных мангровых ассоциаций нижний горизонт литорали (26 видов), в среднем горизонте литорали найдено несколько меньше видов (21 вид), а самым бедным в мангровых посадках был верхний горизонт литорали (15 видов). В естественной мангровой ассоциации в заливе Дам Бай видовое богатство примерно соответствовало богатству верхнего горизонта (14 видов). В естественной мангровой ассоциации в Ня Фу зарегистрировано самое низкое видовое богатство, здесь найдено всего 3 вида двустворчатых моллюсков.
В исследованных ассоциациях найдено всего два специализированных вида Geloina erosa (Lightfoot 1786) и Pegophysema philippiana (Reeve 1850). Оба они встречены и в естественной мангровой ассоциации в Ня Фу, и в мангровых посадках залива Дам Бай, а в естественной мангровой ассоциации в Дам Бае была найдена только P. philippiana. Остальные обнаруженные в ходе работы двустворчатые моллюски – это виды-эврибионты, характерные для инфауны мягких грунтов литорали и сублиторали (например, представители семейств Tellinidae и Cardiidae). Кроме инфауны, на корнях и стволах мангровых деревьев встречаются представители эпифауны, например, Saccostrea cuccullata (Born 1778) и представители семейства Mytilitdae. В нижнем горизонте литорли доля сублиторальных видов выше, чем на других станциях. Кроме того, также на этой станции в качественных пробах встречаются крупные формы, например, представители семейств Veneridae и Pinnidae.
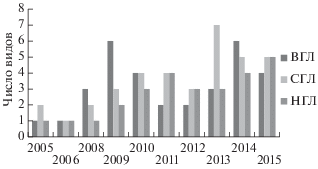
На начальном этапе мониторинга (2005–2006 гг.) разнообразие двустворчатых моллюсков не превышало 1–2 вида, тогда как в последующие годы их разнообразие достигает 8 видов в верхнем, 9 в среднем и 12 видов в нижнем горизонте литорали (см. рис. 7).
Рис. 7.
Динамика видового богатства двустворчатых моллюсков в трех горизонтах литорали в районе мангровых посадок.
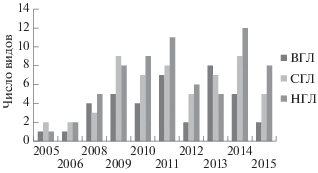
Плотность и биомасса двустворчатых моллюсков испытывали сильные колебания на всем протяжении работ без видимых тенденций к увеличению или уменьшению с течением времени. Наибольшая плотность была зарегистрирована в нижнем горизонте в 2008 г. (153 ± 138 экз./м2). Наибольшая биомасса была зарегистрирована в 2013 г. в естественной мангровой ассоциации в Дам Бае (338 ± 151 экз./м2). В верхнем и среднем горизонтах литорали, а также в естественной мангровой ассоциации в Дам Бае – наибольший вклад в плотность и биомассу вносит вид-эврибионт Gafrarium pectinatum (Linnaeus 1758).
Крабы
В трех мангровых ассоциациях за весь период наблюдений было найдено 18 видов крабов. В верхнем горизонте литорали обнаружено 11 видов крабов, в среднем – 14 видов, в нижнем – 8 видов. Самыми бедными в видовом отношении были естественные мангровые ассоциации, и в заливе Дам Бай и в Ня Фу разнообразие в обоих случаях было представлено шестью видами. Надсемейство Ocypodoidea в исследованных ассоциациях представлено 9 видами. Надсемейство Grapsoidea представлено всего двумя видами: 1. Metopograpsus latifrons (White 1847) найден на всех станциях, 2. единственный экземпляр Sesarmidae gen. sp., найден в последней съемке (октябрь 2015 г.) в СГЛ.
Видовое богатство выросло с течением времени от 1–2 видов в первых двух съемках до 6 видов в верхнем, 7 в среднем и 5 видов в нижнем горизонте литорали в разные годы позднего этапа мониторинга (рис. 8).
Биомасса и плотность крабов испытывали значительные колебания без видимых тенденций на протяжении всего периода мониторинга, но в целом их значения были невелики по сравнению с обилием моллюсков. В большинстве случаев значения плотности крабов не превышали 40 экз./м2, а значения биомассы – 16 г/м2.
На всех исследованных станциях наибольший вклад в плотность и биомассу вносили представители надсемейства Ocypodoidea, причем в нижнем горизонте литорали это был вид Macrophthalmus miloti (Crosnier 1965), тогда как на остальных станциях доминировали представители рода Uca.
Другие группы беспозвоночных
Кроме трех ключевых групп беспозвоночных и аннелид, в исследованных мангровых ассоциациях были обнаружены немногочисленные представители других таксономических групп. Всего их обнаружено 13 видов: медуза Cassiopea sp., роющая актиния Edwarsiidae gen. sp., хитон Acanthochitona ostreaphila (Sirenko et Saito 2017), 3 вида раков-отшельников (Clibanarius sp., Paguroidea gen. sp., Coenobitidae gen. sp.), 2 вида креветок (Caridea), 1 вид Stomatopoda, 2 вида Sipunculida, брахиопода Lingula anatina Lamarck 1801 и один представитель кишечнодышащих (Ptychoderidae gen. sp.). Креветки и раки-отшельники были встречены на всех исследованных станциях, остальные виды из перечисленных встречались только в нижнем горизонте литорали в мангровых посадках залива Дам Бай. Ни одна из этих таксономических групп не обладала высокими значениями обилия, и в динамике их количественных показателей не было выявлено тенденций.
ОБСУЖДЕНИЕ
Разнообразие брюхоногих моллюсков данных ассоциаций подробно обсуждалось в статье Звонаревой и др. (Zvonareva et al., 2016). Проведенное ими сравнение списков видов ряда работ по Тайланду, Филиппинам, Малайзии, Вьетнаму и др. показало, что для мангровых ассоциаций заливов Дам Бай и Ня Фу характерно довольно высокое разнообразие брюхоногих моллюсков.
Оценка разнообразия двустворчатых моллюсков проведена в основном по количественным пробам (в отличие от оценки брюхоногих моллюсков и крабов), т.к. представители этой группы принадлежат преимущественно к инфауне, находятся в толще грунта и визуально плохо заметны. Т.е. обнаруженное видовое богатство в большой степени зависит от количества проб. В работе по естественным манграм северного Вьетнама приведен список из 91 вида двустворчатых моллюсков (Hong, San, 1993), из них всего три вида присутствуют в материале, собранном в ходе настоящего исследования, однако большая часть видов из этого списка относится к фауне сублиторали. В различных работах из других районов Юго-Восточной Азии было найдено от 4 до 44 видов двустворчатых моллюсков (Jiang, Lee, 1995; Macintosh et al. 2002; Lozouet, Plaziat, 2008; Wells, 1983). Таким образом, видовой список двустворчатых моллюсков данной работы является одним из самых длинных. Возможно, это связано с высокой интенсивностью отбора проб и с тем, что в ходе работы пробы отбирали и в нижнем горизонте литорали, где попадалось много видов, характерных для сублиторали, например, Vasticardium angulatum, Ruditapes philippinarum и др.
В статье Чертопруд (Chertoprud et al., 2012) приведен список видов из мангровых посадок в заливе Дам Бай. Нам удалось обнаружить все виды из этого списка, кроме Parasesarma ungulatum, поскольку он встречается уже в супралиторали, а в нашей работе сборы проводили только на литорали. Но нами было обнаружено пять видов, отсутствующих в работе Чертопруд и коллег, включая 1 вид семейства Sesarmidae. Для фауны залива Ня Фу в той же статье приведены еще пять видов, отсутствующих в наших сборах (включая P. ungulatum). В указанной работе сборы проводили в другой мангровой ассоциации в кутовой части залива, вероятно, поэтому и существует разница в видовых списках. Хонг и Сан (Hong, San, 1993) сообщают о 44 видах крабов в мангровых лесах северного Вьетнама, причем с нашей работой общими являются лишь 4 вида. В естественных ассоциациях Малайзии в районе Саравак был найден 31 вид крабов (Ashton, 2002), 30 видов крабов найдено в районе Селангор (Sasekumar, 1974); 25 видов крабов было найдено в естественных мангровых ассоциациях, 23 вида в восьмилетних посадках (10 из них принадлежат семейству Grapsidae), 13 видов в однолетних посадках в Тайланде (7 из них принадлежат семейству Grapsidae) (Macintosh et al., 2002). Таким образом, видовое богатство крабов в данной работе уступает видовому богатству крабов и в естественных ассоциациях, и в посадках других районов Юго-Восточной Азии. Число видов надсемейства Ocypodoidea сравнимо с их количеством в других районах, но семейства Grapsidae и Sesarmidae представлены очень слабо. Эти различия в таксономическом составе и видовом богатстве могут быть обусловлены несовершенством методики отбора проб, (крабы очень подвижны и быстро прячутся в глубокие норы, и, возможно, для более полного охвата всего разнообразия крабов требуются дополнительные методы лова, например, крабовые ловушки). Но не исключено и то, что мангровые ассоциации других районов сильно отличаются по структуре и степени сложности от исследованных в данной работе и потому обладают более богатой фауной.
В исследованных ассоциациях наиболее разнообразными и обильными являются те же ключевые группы, что и в других районах Индо-Вест Пацифики (Berry, 1963; Kathiresan, Bingham, 2001; Lee, 1998, 1999; Macnae, 1968). Состав менее значимых групп макробентоса также типичен для мангров Индо-Вест Пацифики (Kathiresan, Bingham, 2001). Таким образом, полученные данные о составе и разнообразии макробентоса вписываются в общую картину мангровой фауны Индо-Вест-Пацифики и Юго-Восточной Азии.
Обобщив полученные данные по динамике видового состава и обилия, можно восстановить события, произошедшие за 10 лет существования мангровых посадок. До высадки саженцев в СГЛ мангры представляли собой очень узкий пояс Rhizophora apiculata с включением нескольких деревьев Avicennia marina в верхнем горизонте литорали. Эта мангровая ассоциация имела небольшую площадь и была просто организована, и биогенные элементы свободно вымывались приливами. Этим можно объяснить низкое разнообразие макробентоса и низкое обилие моллюсков на начальном этапе наблюдения мангровых посадок. В отличие от моллюсков, крабы обладали высоким обилием в первый год наблюдений, причем эти значения достигаются исключительно благодаря крабам Uca spp., которые (по литературным данным) характерны для незрелых мангровых посадок и фрагментированных мангров (Macintosh et al., 2002).
Высадка в 2004 г. монокультуры R. apiculata в СГЛ привела к ряду изменений, как на этой станции, так и в верхнем горизонте. В среднем горизонте саженцы хорошо прижились и росли довольно быстро, в связи с чем появилось большое количество свободной органики в среднем горизонте напрямую – виде опада, а также опосредованно – появление затененных манграми участков создало благоприятные условия для развития микроводорослей и макрофитов. Появление в экосистеме дополнительной органики вызвало резкий рост разнообразия неспециализированных видов моллюсков и крабов, рост обилия видов-оппортунистов, неспециализированных к манграм хищников и падальщиков. Привлечению новых видов также способствовало появление твердого субстрата в виде подрастающих деревьев. О подобном росте обилия макробентоса сообщается в работе, где сравниваются посадки разного возраста в Таиланде (Macintosh et al., 2002). Саженцы в среднем горизонте замедляют приливно-отливные течения и препятствуют вымыванию органики из верхнего горизонта. Также здесь происходит рост численности видов-оппортунистов и неспециализированных видов. Таким образом, теперь мангры занимают значительную площадь и предоставляют большое количество микросред. Вследствие этого, в фауне начинают появляться специализированные мангровые виды. По литературным данным (что подтверждают наши наблюдения), площадь мангровой ассоциации и разнообразие ее растительности являются главными факторами, определяющими разнообразие мангровой фауны и особенно специализированных видов (Ellison et al., 1999, Hutchings, Saenger, 1987; Lee, 1998). Резкие колебания биомассы и плотности разных групп макробентоса можно объяснить естественной вариабельностью.
Естественная мангровая ассоциация в заливе Дам Бай имела много общего с верхним горизонтом литорали в районе посадок до 2004 г. (небольшая площадь, структура растительности). И этим объясняется низкое разнообразие специализированной мангровой фауны в этой ассоциации, а также относительно невысокое обилие брюхоногих моллюсков.
Естественная мангровая ассоциация в Ня Фу была самой обширной, имела сложную пространственную структуру и обладала высоким флористическим разнообразием. Вероятно, именно с этим связано высокое видовое богатство специализированных брюхоногих моллюсков. Эта ассоциация отличается от остальных исследованных станций крайне низким разнообразием и обилием двустворчатых моллюсков. Это может быть связано с тем, что мангровые заросли – не самая благоприятная среда для жизни двустворчатых моллюсков. Большинство двустворчатых моллюсков – фильтраторы, и для многих из них проблематично жить в условиях редкого затопления в верхних горизонтах, а в естественных условиях мангровые деревья способны заселить только верхнюю часть литоральной зоны (Morton, 1976). Кроме того, известно, что в зрелом мангровом лесу грунт имеет кислую реакцию и в нем способны жить двустворчатые моллюски лишь с очень толстой раковиной (Morton, 1976). Таким образом, в зрелых манграх могут жить лишь специализированные двустворчатые моллюски, которые имеют определенные приспособления к этим сложным условиям, остальные же живут здесь на пределе своей экологической пластичности (Morton, 1976).
Фауна нижнего горизонта литорали даже через 8 лет после высадки саженцев резко отличалась по всем показателям от верхнего и среднего горизонтов в районе мангровых посадок, а также от обеих естественных мангровых ассоциаций. Рост саженцев здесь угнетен, а фауна больше похожа по составу на фауну илистой литорали и верхней сублиторали, нежели на фауну мангров. Вероятно, это происходит из-за влияния слишком частого и продолжительного затопления грунта, т.к. здесь создаются условия, не подходящие ни для саженцев ризофоры, ни для нормальной мангровой фауны.
На момент последней съемки можно заключить, что после 10 лет развития нового сообщества структура и состав фауны в нем все еще сильно отличаются от фауны зрелой мангровой ассоциации в Ня Фу. Но в то же время фауна верхнего и среднего горизонтов мангровых посадок уже больше похожа на фауну зрелых мангров, чем фауна естественной ассоциации в заливе Дам Бай, характеризующаяся небольшой площадью, простотой пространственной структуры и низким разнообразием растительности.
Список литературы
Adrianov A., Maiorova A., 2012. Peanut worms of the phylum Sipuncula from the Nha Trang Bay (South China Sea) with a key to species // Zootaxa. V. 3166. P. 41–58.
Anker A., Poddoubtchenko D., Marin I.N., 2006. On the presence of the alpheid shrimp genus Bermudacaris Anker and Iliffe, 2000 (Crustacea: Decapoda: Caridea) in the Pacific Ocean, with description of a new species from Vietnam // Journal of Natural History. V. 40. Issue 27–28. P. 1675–1686.
Ashton E.C., 2002. Mangrove sesarmid crab feeding experiments in Peninsular Malaysia // Journal of Experimental Marine Biology and Ecology. V. 273. Issue 1. P. 97–119.
Berry A.J., 1963. Faunal zonation in Mangrove swamps // Bulletin of the National Museum State of Singapore. V. 32. P. 90–98.
Chertoprud E.S., Spiridonov V.A., Marin I.N., Mokievsky V.O., 2012. Brachyuran crabs (Crustacea Decapoda Brachyura) of the mangrove intertidal zone of southern Vietnam // Benthic fauna of the Bay of Nhatrang, Southern Vietnam. V. 2. P. 258–295.
Dittmann S., 2002. Benthic fauna in tropical tidal flats – a comparative perspective // Wetlands Ecology and Management. V. 10. P. 189–195.
Dworschak P.C., Marin I., Anker A., 2006. A new species of Naushonia Kingsley, 1897 (Decapoda: Thalassinidea: Laomediidae) from Vietnam and the Philippines with notes on the genus Espeleonaushonia Juarrero & Martínez-Iglesias, 1997 // Zootaxa. V. 16. P. 1–16.
Ellison A.M., 2008. Managing mangroves with benthic biodiversity in mind: Moving beyond roving banditry // Journal of sea research. V. 59. P. 2–15.
Ellison A.M., Fransworth E.J., Merkt R.E., 1999. Origins of mangrove ecosystems and the mangrove biodiversity anomaly // Global Ecology and Biogeography. V. 8. Issue 2. P. 95–115.
Field C.D., 1998. Rehabilitation of Mangrove Ecosystems an Overview // Marine Pollution Bulletin. V. 37. Issue 8–12. P. 383–392.
Frey M.A., 2010. A revised classification of the Gastropod genus Nerita // The Veliger. V. 51. Issue 1. P. 1–7.
Gherardi F., Vannini M., 1993. Hermit crabs in a mangrove swamp: proximate and ultimate factors in the clustering of Clibanarius laevimanus // Journal of Experimental Marine Biology and Ecology. V. 168. Issue 2. P. 167–187.
Glover E., Taylor J., 2016. Lucinidae of the Philippines: highest known diversity and ubiquity of chemosymbiotic bivalves from intertidal to bathyal depths (Mollusca: Bivalvia) // Tropical Deep-Sea Benthos. Héros V., Strong E., Bouchet P. (Eds). Paris: Mémoires du Museum national d’Histoire Naturelle. P. 65–234.
Gopal B., Chauhan M., 2006. Biodiversity and its conservation in the Sundarban mangrove ecosystem // Aquatic Sciences. V. 68. Issue 3. P. 338–354.
Hong P.N., San H.T., 1993. Mangroves of Vietnam. Bangkok: IUCN. 173 p.
Houbrick R.S., 1985. Genus Clypeomorus Jousseaume (Cerithiidae–Prosobranchia) // Smithsonian Contributions to Zoology. V. 403. P. 1–131.
Houbrick R.S., 1992. Monograph of the Genus Cerithium Bruguiere in the Indo-Pacific (Cerithiidae–Prosobranchia) // Smithsonian Contributions to Zoology. V. 510. P. 1–211.
Hutchings P., Seanger P., 1987. Ecology of mangroves. Heatwall H. (Ed.). St. Lucia: University of Queensland Press. 370 p.
Jiang J.X., Li R.G., 1995. An ecological study on the Mollusca in mangrove areas in the estuary of the Jiulong River // Hydrobiologia. V. 295. Issue 1–3. P. 213–220.
Kathiresan K., Bingham B.L., 2001. Biology of mangroves and mangrove Ecosystems // Advances in Marine Biology. V. 40. P. 81–251.
Kumar R.S., 2000. A review of biodiversity studies of soil dwelling organisms in indian mangroves // Zoos’ Print Journal. V. 15. P. 221–227.
Lee S.Y., 1998. Ecological role of grapsid crabs in mangrove ecosystems: a review // Marine and Freshwater Research. V. 49. Issue 4. P. 335–343.
Lee S.Y., 1999. Tropical mangrove ecology: Physical and biotic factors influencing ecosystem structure and function // Australian Ecology. V. 24. Issue 4. P. 355–366.
Lozouet P., Plaziat J.-C., 2008. Mangrove environments and mollusks Abatan River, Bohol and Panglao Islands, Central Philippines. Hackenheim: ConchBooks. 166 p.
Macintosh D.J., Ashton E.C., Havanon S., 2002. Mangrove rehabilitation and intertidal biodiversity: a study in the Ranong mangrove ecosystem, Thailand // Estuarine, Coastal and Shelf Science. V. 55. Issue 3. P. 331–345.
Macnae W., 1968. A general account of the fauna and flora of mangrove swamps and forests in the Indo-West-Pacific region // Advances in Marine Biology. V. 6. P. 73–270.
Marin I.N., 2008. Description of two new species from the genera Palaemonella Dana, 1852 and Vir Holthuis, 1952 (Crustacea: Caridea: Palaemonidae: Pontoniinae) // Zoologische Mededelingen. V. 82. Issue 35. P. 375–390.
Marin I., 2014. The first record of an association between a pontoniine shrimp (Crustacea: Decapoda: Palaemonidae: Pontoniinae) and a thalassematid spoon worm (Echiura: Thalassematidae), with the description of a new shrimp species // Zootaxa. V. 3847. Issue 4. P. 557–566.
Mazda Y., Magi M., Ikeda Y., Kurokawa T., Asano T., 2006. Wave reduction in a mangrove forest dominated by Sonneratia sp. // Wetlands Ecology and Management. V. 14. Issue 4. P. 365–378.
Mazda Y., Magi M., Nanao H., Kogo M., Miyagi T. et al., 2002. Coastal erosion due to long-term human impact on mangrove forests // Wetlands Ecology and Management. V. 10. Issue 1. P. 1–9.
Metcalfe K.N., Glasby C.J., 2008. Diversity of Polychaeta (Annelida) and other worm taxa in mangrove habitats of Darwin Harbour, northern Australia // Journal of Sea Research. V. 59. Issue 1–2. P. 70–82.
Mitra S., Pattanayak J., 2013. Diversity and distribution of sea-anemones (Cnidaria: Actiniaria) in the estuaries and mangroves of Odisha, India // Zoological Survey of India. V. 113. Issue 3. P. 113–118.
Mitra S., Pattanayak J., 2013a. Studies on Lingula anatina (Brachiopoda: Inarticulata) in Subarnarekha estuary, Odisha with special reference to habitat // Zoological Survey of India. V. 113. Issue 3. P. 49–53.
Mokievsky V.O., Tchesunov A.V., Udalov A.A., Nguen D.T., 2011. Meiobenthos of a mangrove intertidal area of Nha Trang Bay, Vietnam, South China Sea: quantitative distribution and community structure with special reference to free-living nematodes // Biologiya Morya. V. 37. P. 262–273.
Morton B., 1976. The biology, ecology and functional aspects of the organs of feeding and digestion of the S.E. Asian mangrove bivalve, Enigmonia aenigmatica (Mollusca: Anomiacea) // Journal of Zoology. V. 179. P. 437–466.
Ng Y.P., 2013. The Ecology of Stomatopods in Matang waters with emphasis on Miyakea nepa and Oratosquillina perpensa. PhD thesis. Kuala Lumpur: University of Malaya, Faculty of sciensce. 180 p.
Printrakoon C., Kamlung-ek A., 2013. Socioeconomic study and economic value of living fossil, aLingula sp. in mangrove ecosystem in Trat Province, Thailand // Chinese Journal of Population Science. V. 11. Issue 3. P. 187–199.
Reid D.G., 1985. Habitat and zonation patterns of Littoraria species (Gastropoda: Littorinidae) in Indo-Pacific mangrove forests // Biological Journal of the Linnean Society. V. 26. P. 39-68.
Reid D.G., 1986. The littorinid molluscs of mangrove forests in the Indo-Pacific region: the genus Littoraria. London: British Museum (Natural History). 227 p.
Reid D.G., 2014. The genus Cerithidea Swainson, 1840 (Gastropoda: Potamididae) in the Indo-West Pacific region // Zootaxa. V. 3775. Issue 1. P. 1–65.
Reid D.G., Dyal P., Lozouet P., Glaubrecht M., Williams S.T., 2008. Mudwhelks and mangroves: the evolutionary history of an ecological association (Gastropoda: Potamididae) // Molecular phylogenetics and evolution. V. 47. Issue 2. P. 680–99.
Reid D.G., Dyal P., Williams S.T., 2010. Global diversification of mangrove fauna: a molecular phylogeny of Littoraria (Gastropoda: Littorinidae) // Molecular phylogenetics and evolution. V. 55. Issue 1. P. 185–201.
Spalding M., Blasco F., Field C., 1997. World mangrove atlas. Okinawa: International Society for Mangrove Ecosystems. 178 p.
Tchesunov A.V., Mokievsky V.O., Thanh N.V., 2010. Three new free-living nematode species (Nematoda, Enoplida) from mangrove habitats of Nha Trang, Central Vietnam // Russian Journal of Nematology. V. 18. Issue 1. P. 155–172.
Tsingalia M.H., 2014. Population and spatial dynamics mangrove jellyfish Cassiopeia sp. at Kenya’s Gazi bay // American Journal of Life Sciences. V. 2. Issue 6. P. 395–399.
Twilley R.R., Chen R., Hargis T., 1992. Carbon sinks in mangroves and their implications to carbon budget of tropical coastal ecosystems // Water, Air Soil Pollution. V. 64. P. 265–288.
Walters B.B., 2000. Local Mangrove Planting in the Philippines: Are Fisherfolk and Fishpond Owners Effective Restorationists? // Restoration Ecology. V. 8. Issue 3. P. 237–246.
Wells F., 1983. An analysis of marine invertebrate distributions in a mangrove swamp in northwestern Australia // Bulletin of Marine Science. V. 33. Issue 3. P. 736–744.
Xuan Q., Vanreusel A., Thanh N.V., Smol N., 2007. Biodiversity of meiofauna in the intertidal Khe Nhan mudflat, Can Gio mangrove forest, Vietnam with special emphasis on free living nematodes // Ocean Science Journal. V. 42. Issue 3. P. 135–152.
Yu R., Chen G.Z., Wong Y.S., Tam N.F.Y., Lan C.Y., 1997. Benthic macrofauna of the mangrove swamp treated with municipal wastewater // Hydrobiologia. V. 347. P. 127–137.
Zvonareva S.S., Kantor Yu.I., Li X., Britayev T.A., 2015. Long-term monitoring of Gastropoda (Mollusca) fauna in planted mangroves in central Vietnam // Zoological Studies. V. 54. P. 39.
Zvonareva S.S., Kantor Yu.I., 2016. Checklist of gastropod molluscs in mangroves of Khanh Hoa province, Vietnam // Zootaxa. V. 4162. Issue 3. P. 401.
Дополнительные материалы отсутствуют.
Инструменты
Зоологический журнал