

Зоологический журнал, 2020, T. 99, № 8, стр. 843-865
Закономерности распределения макрозообентоса в некоторых реках европейской части России и сопредельных территорий
Л. В. Воробьева a, *, И. С. Решетов b, **, А. И. Азовский b, ***, Н. П. Ковачева a, Е. С. Чертопруд b, c, ****
a Институт рыбного хозяйства и океанографии
107140 Москва, Россия
b Биологический факультет Московского государственного университета им. М.В. Ломоносова
119991 Москва, Россия
c Институт экологии и эволюции им. А.Н. Северцова РАН
119071 Москва, Россия
* E-mail: vorobjeva.lada@yandex.ru
** E-mail: reshetov137@gmail.com
*** E-mail: aiazovsky@mail.ru
**** E-mail: horsax@yandex.ru
Поступила в редакцию 03.04.2019
После доработки 16.06.2019
Принята к публикации 07.08.2019
Аннотация
Проведен статистический анализ закономерностей пространственного распределения макрозообентоса ряда водотоков территории европейской части России и Латвии (бассейны Оки, Дона, Гауи). На основании неметрического многомерного шкалирования (NMDS) показано, что распределение макрозообентоса в изученных водотоках континуально, а четко выраженные типы сообществ отсутствуют. На основании процедуры SIMPER выявлены статистически достоверные видовые комплексы, связанные с типами субстрата, скоростью течения, размерным классом водотоков, а также с водосборными бассейнами. При помощи процедуры ANOSIM для каждого видового комплекса выделены характерные и дифференцирующие таксоны ранга рода или вида, оценены жизненные формы этих таксонов. Обосновано, что, хотя таксономический состав видовых комплексов текучих вод слабо прогнозируем, возможно статистически достоверное выявление группировок таксонов, связанных с ключевыми факторами среды. Проведение подобных исследования в водоемах различных гидрологических типов в разных географических регионах является перспективным направлением экологического мониторинга пресных вод.
Один из основных вопросов экологии: существуют ли сообщества как дискретные, хорошо определяемые единицы, или эти сообщества образуют континуум, плавно переходя друг в друга (Heino et al., 2003; Жирков, 2017)?
Ряд подходов к классификации водных сообществ основан именно на предпосылке о существовании дискретных видовых комплексов. Однако есть мнение, что четко выделяемые типы сообществ связаны с применением для классификации методов кластеризации и являются отчасти искусственными (Heino et al., 2003). Другой подход к пониманию организации сообществ связан с концепцией “правил сборки” (“assembly rules”). В рамках этой концепции определяется, какой набор видов из регионального пула будет встречаться в данном местообитании. Современное понимание этой концепции основную роль в организации сообществ отводит истории колонизации местообитания и локальным экологическим фильтрам. Факторы среды служат фильтрами, позволяя заселить местообитание только тем видам из регионального пула, которые способны сохранять жизнеспособность при данных условиях. Серия таких фильтров должна задавать четкие рамки для состава сообществ, благодаря чему структура сообществ вдоль экологического градиента может быть хорошо прогнозируемой (Keddy, 1992; Heino et al., 2003; Diamond, 1975).
Экологические фильтры разнятся по масштабу и представляют собой иерархическую систему. В случае водотоков выделяют четыре пространственных уровня таких фильтров: бассейн, долина реки, участок русла, микробиотоп (Poff, 1997). На уровне бассейна значимы такие факторы, как климат, геология, преобладающий тип хозяйственной деятельности. На уровне речной долины – размер реки, ширина долины, тип землепользования, морфология русла. На уровне руслового участка – распределение субстратов, вертикальная циркуляция воды. На уровне микробиотопа – размер частиц грунта и скорость течения. Биотические взаимодействия влияют на состав сообществ на локальном уровне, организмы, вступающие в них, должны преодолеть экологические фильтры более крупного масштаба (Poff, 1997).
Возможность прогнозирования состава сообществ важна для адекватной оценки состояния водотока. Исследования ненарушенных и слабо нарушенных антропогенной деятельностью участков рек позволяют составить представление о диапазоне изменчивости экосистем в естественных условиях среды. Райт с соавторами (Wright et al., 1984) в работе по изучению водотоков Великобритании применил методы ординации (бестрендовый анализ соответствий) и кластерного анализа (двухфакторный анализ индикаторных видов или TWINSPAN) на основе данных по видовому составу бентосных макробеспозвоночных. В этой же работе был применен подход к прогнозированию ожидаемого состава сообществ на основании базы данных о видовом составе и наборе экологических переменных в каждом из местообитаний (Wright et al., 1984; Reynoldson et al., 2001). Методы ординации и кластерного анализа с использованием множества экологических переменных (физические, химические и биологические характеристики местообитания) также широко применяются для классификации (Wright et al., 1984; Ormerod, Edwards, 1987; Reynoldson et al., 2001; Heino et al., 2003).
В отечественной литературе распространен топологический, или биотопический подход к классификации сообществ, предложенный В.И. Жадиным. В основу классификации здесь положены тип субстрата и скорость течения. По этим признакам для рек выделяются лито-, псаммо-, пело-, аргилло- и фитореофильные сообщества (или сообщества каменистых, песчаных, илистых, глинистых грунтов и сообщества беспозвоночных водных макрофитов соответственно) (Жадин, 1940; Жадин, Герд, 1961).
Классификация М.В. Чертопруда основана на наборах жизненных форм, характерных для сообществ и их классов (Чертопруд, 2011; Палатов и др., 2016; Чертопруд, Палатов, 2017; Палатов, Чертопруд, 2018). В этой классификации тип субстрата является одним из основных абиотических факторов, определяющих формирование сообщества. Для Европейской части России автор выделяет 39 типов сообществ, относящихся к четырем комплексам: креналь, ритраль, фиталь и пелаль. Каждый тип сообщества характеризуется определенным набором видов, для выделения сообществ используется модификация метода Браун-Бланке. Креналь (ручьевой комплекс) – водотоки с меженным водорасходом до 0.01 м3/с и шириной до 1 м, отличается мелкой мозаикой различных субстратов детритно-опадно-песчаного ряда, очень маленькой глубиной, относительно слабым течением (не более 0.1–0.2 м/с), крупнодисперсным органическим веществом, сильной затененностью. Ритраль – сообщества плотных, главным образом каменистых, грунтов с сильным течением и невысокой температурой воды. Фиталь – комплекс зарослевых сообществ. Пелаль – сообщества мягких грунтов (ил, заиленный песок, заиленный детрит) (Чертопруд, 2011).
В рамках данной работы проанализирована связь таксономического состава водных сообществ макрозообентоса с речным бассейном, типом субстрата, скоростью течения. Параллельно оценено, насколько велика степень дискретности выделяемых видовых комплексов, приуроченных к разным биотопам. Кроме того, интерес представляет оценка применимости некоторых распространенных в макроэкологии математических методов анализа данных для анализа сообществ макрозообентоса пресных текучих вод.
МАТЕРИАЛЫ И МЕТОДИКА
Районы исследования. В работе использован материал, собранный на р. Гауя и в ее бассейне (Латвия) в октябре 2013 г., р. Поля в июне–июле 2013 г., р. Угра в июле 2014 г, р. Тихая Сосна на территории заповедника “Дивногорье” в июне 2015 г. (Воронежская обл.), р. Жиздра и в ее бассейне, а также на р. Ока в июле 2015 г. (Калужская обл.), на р. Ворона в августе 2015 г. (Тамбовская обл.). В основном пробы отбирали на средних реках длиной 100–500 км и в устьях их притоков. Пробы на ручьях отбирали в основном в бассейне р. Гауи. Перечень рек и число проб с каждой из них приведен в табл. 1. Всего обработано 182 пробы. Помимо указанных в таблице, были собраны пробы с ручьев, названия которых не указаны. Это водотоки шириной 0.2–1 м, глубиной менее 0.5 м, вероятно, пересыхающие. В бассейне Гауи насчитывалось 8 таких проб, в бассейне Оки – 5.
Таблица 1.
Характеристика обследованных водотоков
| Название реки | Географическое положение точек отбора проб | Русло | Бассейн | Примечание | Число проб | ||
|---|---|---|---|---|---|---|---|
| ширина, м | длина, км | площадь, км2 | название | ||||
| Гауя | Латвия | 10–30 | 460 | 8.9 тысяч | Гауя | – | 17 |
| Вельюпите | Латвия | 3–4 | – | – | Гауя | Лесной овраг | 3 |
| Дауда | Латвия | 1.5–2 | – | – | Гауя | – | 1 |
| руч. Тилберга | Латвия | Менее 1 | – | – | Гауя | Лесной овраг | 1 |
| Ратниеку | Латвия | 2–3 | – | – | Гауя | Лесной овраг | 1 |
| Скаульпе | Латвия | 2–3 | – | – | Гауя | Лесной овраг | 3 |
| Амата | Латвия | 7–10 | 68 | – | Гауя | – | 6 |
| Упмали | Латвия | 2–3 | – | – | Гауя | Лесной овраг | 3 |
| Ракшупе | Латвия | 2.5–3 | – | – | Гауя | Лесной овраг | 2 |
| Перлюпе | Латвия | 2.4–3 | – | – | Гауя | – | 3 |
| Кумада | Латвия | 3–4 | – | – | Гауя | Лесной овраг | 3 |
| Поля | РФ, Московская обл. | 5–10 | 92 | 1560 | Ока | – | 5 |
| Клязьма | Московская обл., близ устья р. Поли | 10–30 | 686 | 42.5 тысяч | Ока | – | 6 |
| Угра | Смоленская обл., Калужская обл. | 10–30 | 399 | 15.7 тысяч | Ока | – | 29 |
| Тихая Сосна | Воронежская обл. | 5–10 | 161 | 4350 | Дон | Правый приток Дона. Пробы отобраны в окрестностях музея-заповедника “Дивногорье” | 10 |
| Жиздра | Калужская обл. | 5–10 | 223 | 9170 | Ока | Левый приток Оки | 32 |
| Серена | Калужская обл. | 5–10 | 108 | 1130 | Ока | Левый приток Жиздры, пробы отобраны в устьевом участке | 2 |
| Вытебеть | Калужская обл. | 5–10 | 133 | 1760 | Ока | Правый приток Жиздры, пробы отобраны в устьевом участке | 3 |
| Рессета | Калужская обл. | 5–10 | 123 | 2270 | Ока | Правый приток Жиздры, пробы отобраны в устьевом участке | 6 |
| Ока | Калужская обл. | 1500 | 245 тысяч | Ока | Пробы отобраны на участке от устья р. Жиздры до г. Калуга | 6 | |
| Вырка | Калужская обл. | 43 | 260 | Ока | Левый приток Оки, пробы отобраны в устьевом участке | 2 | |
| Ворона | Тамбовская обл. | 454 | 13.2 тысяч | Дон | – | 25 | |
Отбор проб
Для отбора проб использовали гидробиологический сачок площадью 0.02 м2. На каждой станции длиной 10–20 м пробы отбирали в точках с однородными условиями среды (грунт, наличие или отсутствие макрофитов, скорость течения). В каждой точке организмы отбирали до тех пор, пока число видов не выходило на плато, затем их фиксировали в 70% растворе этилового спирта.
Идентификация организмов
Таксономическое определение вели в лабораторных условиях по возможности до вида, но в некоторых случаях – до группы видов или рода по определителю пресноводных беспозвоночных России и сопредельных территорий под ред. Цалолихина (тома 3–6).
Статистический анализ данных
Выделение сообществ. Математическую обработку данных проводили на основании относительного обилия организмов каждой таксономической группы в пробах (процент от общей численности). Для предварительного выявления закономерностей распределения видов в пробах использовался метод непрямой ординации – неметрическое многомерное шкалирование (MDS). Метод MDS заключается в визуализации матрицы сходства между пробами, при этом данные о факторах среды и модели отклика видов на изменение факторов не используются. Данный метод подходит для работы с большими биогеографическими матрицами с сильными шумами (Шитиков и др., 2011) и позволяет ответить на вопрос, образуют ли пробы дискретные группы, четко отделенные друг от друга.
Для построения матрицы сходства использовали индекс сходства Чекановского (индекс процентного сходства), вычисленный на основе данных по относительному обилию организмов в пробах.
Для оценки статистической достоверности выделенных групп проб использован метод ANOSIM, основанный на ранжировании индексов сходства между пробами внутри группы и пробами по всей выборке. Вычисляется на основании диагональной матрицы сходства по формуле:
Показатель R может принимать значения (–1; 1), если R = 1, то все пробы внутри группы более сходны между собой, чем пробы из разных групп. Если R = 0, то различий между показателями внутри- и межгруппового сходства нет. Для оценки статистической значимости используется перестановочный тест (Clarke, Warwick, 2001).
Представляет интерес оценка сходства между пробами, учитывающая уровень таксономического определения организмов, т.е. то, насколько распределение таксонов уровня выше видового соответствует распределению видов. Идентификация организмов до вида зачастую требует значительных затрат времени, а также достаточно высокого уровня подготовки специалистов. При этом в экологических исследованиях, особенно прикладного характера, часто необходима обработка большого объема материала в сжатые сроки. Решением этой проблемы может быть идентификация организмов до уровня рода или семейства. Если распределение проб по степени сходства при определении организмов до уровня выше видового, примерно, соответствует таковому на уровне вида, то при экспресс-оценке сообществ достаточно определения до рода или семейства. С помощью индекса гамма-плюс рассчитывали расстояние между видами, основанное на линнеевской таксономической классификации, при этом шаги от вида до рода, от рода до семейства и т.д. равны между собой. Расстояние между идентичными видами в сравниваемых пробах равно нулю, разными видами одного рода – 1, разными родами одного семейства – 2 и т.д. Значение индекса гамма-плюс является средним всех расстояний между каждым видом в одной пробе и ближайшим к нему видом в другой. Вычисляется по формуле:
Анализ таксономической структуры. Для определения видов, внесших наибольший вклад в картину сходства и различия между пробами, использовалась процедура SIMPER. Для каждой группы проб выделены дифференцирующие (вносящие наибольший вклад в показатель различия между группами проб при попарном сравнении групп) и характерные (вносящие наибольший вклад в показатель сходства между пробами внутри группы) таксоны. Подробно процедура описана в работе Кларка и Уорвика (Clarke, Warwick, 2001).
Анализ таксономического разнообразия. Для каждого типа субстрата было рассчитано среднее число видов в пробе, среднее ожидаемое число таксонов ранга рода и вида на 30 особей ES(30) (минимальное число особей в пробе) и среднее значение индекса Шеннона в пробе. Ожидаемое число видов на количество особей рассчитывалось по методу разрежения. Методы расчета приводятся у Кларка и Уорвика (Clarke, Warwick, 2001). Проверка статистической достоверности различий средних значений этих показателей для каждого типа субстрата проводилась с помощью критерия Крускала-Уоллиса.
Математические операции, связанные с выделением сообществ, анализом таксономической структуры и анализом таксономического обилия и разнообразия проведены в программе PRIMER 6. Расчет критерия Крускала-Уоллиса и описательная статистика для распределения числа видов, ожидаемого числа видов на стандартное число особей и индекса Шеннона проведены в программе Statistica 13.
РЕЗУЛЬТАТЫ
Анализируемые данные были разделены на четыре группы по следующим факторам:
– бассейны рек: Ока, Дон, Гауя;
– тип субстрата: песок, ил, глина, пресноводная губка, макрофиты, детрит, камни, коряги;
– скорость течения (0 – отсутствие течения, 1 – низкая скорость (0–0.1 м/с), 2 – средняя скорость (0.1–0.3 м/с), 3 – высокая скорость (более 0.3 м/с));
– размер водотока (1 – ручьи (ширина до 1 м), 2 – малые (ширина 1–5 м), 3 – средние малые (ширина 5–10 м), 4 – средние крупные (ширина 10–30 м), 5 – крупные (ширина свыше 30 м).
Для первоначального выявления закономерностей распределения видов в пробах была проведена процедура многомерного шкалирования (MDS). Данные о распределении точек отбора проб в пространстве ординации не позволяют выявить дискретные группы проб, однако позволяет сделать заметными закономерности распределения сходства таксономического состава по группам “бассейн” и “тип субстрата” (рис. 1, 2).
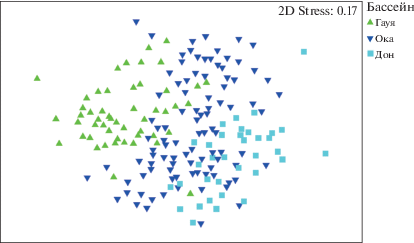
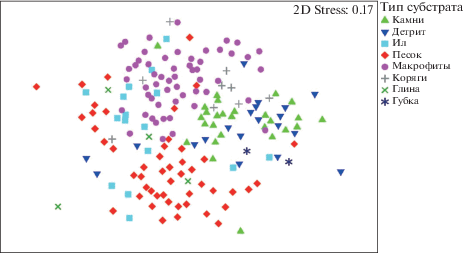
На основании диаграмм ординации можно заключить, что наиболее отчетливо пробы сгруппированы по водосборным бассейнам. Типы субстрата образуют три более или менее хорошо различимые группы: макрофиты, песок, камни + + детрит. Пробы с типом субстрата “ил” распределены более или менее равномерно между песком и макрофитами, пробы с типом субстрата “коряги” группируются большей частью вблизи группы “камни–детрит”, но также распределены в группах “макрофиты” и “песок”. Типы субстратов “глина” и “губка” насчитывают малое число проб, и оценить их расположение затруднительно.
Далее для проверки статистической достоверности распределения проб по группам и проверки результатов ординации была проведена процедура ANOSIM.
Оценка влияния фактора бассейна на видовое распределение
Однофакторный ANOSIM показал, что все различия между бассейнами статистически достоверны. В наибольшей степени различались бассейны Гауи и Дона, что подтверждает картину диаграммы ординации, на которой эти группы удалены друг от друга в наибольшей степени, в то время как группы проб из Оки и Дона значительно перекрываются.
Оценка влияния типа субстрата
При попарном сравнении типов субстрата недостоверными (уровень статистической значимости p более 5%) оказались различия между парами: камни – губка, детрит – губка, ил – глина, коряги – губка, глина – губка. Проб, отобранных с губки, насчитывалось только две, так что в дальнейшем эта группа проб была исключена из анализа. Субстраты ил и глина были объединены в одну группу проб.
Оценка влияния размера водотока
ANOSIM для групп проб, выделенных по размеру водотока, показал отсутствие достоверных различий видового состава для всех размерных классов, кроме одного (1 – ручьи).
Оценка влияния скорости течения
ANOSIM для групп, выделенных по фактору скорости, показал отсутствие достоверных различий между группами 0 и 1 (0 и 0–0.1 м/с), при низком значении R. Наибольшие различия наблюдались между группами 3 и 0 (более 0.3 и 0 м/с). Между остальными парами групп, выделенных на основании скорости течения, наблюдались статистически достоверные различия при низком значении R.Таким образом, в наибольшей степени различались группы, отобранные на быстром течении, от групп, отобранных на слабом течении или на участках без течения. Между группами, отобранными на участках с низким течением и на участках без течения, достоверных различий не выявлено.
Значения показателя R и уровня достоверности p при попарном сравнении групп проб в ходе процедуры однофакторного ANOSIM представлены в табл. 2.
Таблица 2.
Значения показателя R и уровня статистической достоверности для пар групп проб при попарном сравнении в ходе процедуры однофакторного ANOSIM*
| Группы проб | Значение R | Уровень достоверности p, % |
|---|---|---|
| Бассейн | ||
| Гауя–Ока | 0.280 | 0.1 |
| Гауя–Дон | 0.611 | 0.1 |
| Ока–Дон | 0.143 | 0.1 |
| Тип субстрата | ||
| Камни–Детрит | 0.209 | 0.1 |
| Камни–Ил | 0.441 | 0.1 |
| Камни–Песок | 0.407 | 0.1 |
| Камни–Макрофиты | 0.403 | 0.1 |
| Камни–Коряги | 0.175 | 4.7 |
| Камни–Губка | 0.314 | 10.6 |
| Камни–Глина | 0.654 | 0.05 |
| Детрит–Ил | 0.365 | 0.1 |
| Детрит–Песок | 0.388 | 0.1 |
| Детрит–Макрофиты | 0.481 | 0.1 |
| Детрит–Коряги | 0.281 | 0.9 |
| Детрит–Губка | 0.378 | 8.2 |
| Детрит–Глина | 0.527 | 0.3 |
| Ил–Песок | 0.185 | 0.1 |
| Ил–Макрофиты | 0.347 | 0.1 |
| Ил–Коряги | 0.151 | 2 |
| Ил–Губка | 0.492 | 1.3 |
| Ил–Глина | 0.116 | 17.7 |
| Песок–Макрофиты | 0.430 | 0.1 |
| Песок–Коряги | 0.395 | 0.1 |
| Песок–Губка | 0.483 | 0.2 |
| Песок–Глина | 0.349 | 0.4 |
| Макрофиты–Коряги | 0.300 | 0.1 |
| Макрофиты–Губка | 0.543 | 0.07 |
| Макрофиты–Глина | 0.457 | 0.06 |
| Коряги–Губка | 0.227 | 18.2 |
| Коряги–Глина | 0.296 | 3.2 |
| Глина–Губка | 0.357 | 6.7 |
| Размер водотока | ||
| Ручьи–Прочие | 0.197 | 0.1 |
| Скорость течения | ||
| 0 м/с–0–0.1 м/с | 0.031 | 24.1 |
| 0 м/с–0.1–0.3 м/с | 0.077 | 2.3 |
| 0 м/с–более 0.3 м/с | 0.302 | 0.1 |
| 0–0.1 м/с–0.1–0.3 м/с | 0.042 | 1.3 |
| 0–0.1 м/с–более 0.3 м/с | 0.206 | 0.1 |
| 0.1–0.3 м/с–более 0.3 м/с | 0.044 | 1.8 |
Разграничение влияния фактора бассейна и других факторов
Наиболее четко выражены различия между бассейнами. Чтобы исключить влияние фактора бассейна на распределение видового состава в остальных группах проб, был проведен двухфакторный перекрестный ANOSIM для групп, выделенных на основании факторов субстрата, размера водотока и скорости течения, с группами, выделенными на основании фактора бассейна.
На основании двухфакторного ANOSIM выделение размерной группы 1 (ручьи) остается статистически достоверным.
Для групп, выделенных на основании фактора скорости течения, достоверных различий не выявлено для пар. В наибольшей степени различались пары 3–0 и 3–1 (более 3 и 0 м/с; более 3 и 0–0.1 м/с). Достоверно, но с низким значением R различалась пара 3–2 (более 0.3 и 0.1–0.3 м/с). Таким образом, можно выделить группу проб с высокой скоростью течения (более 0.3 м/с), как достоверно отличающуюся от остальных групп, выделенных по признаку скорости течения. Для дальнейшего анализа группы 0, 1 и 2 были объединены.
Двухфакторный ANOSIM для групп, выделенных по типу субстрата, выявил отсутствие статистической значимости для пар детрит–ил, камни–коряги, ил–коряги.
Достоверные различия во всех случаях попарного ANOSIM наблюдались для следующих типов субстрата: макрофиты, песок; в четырех случаях из пяти: камни; в трех случаях из пяти: ил, детрит, коряги. Наибольшая степень несходства видового состава наблюдалась в парах камни–песок, детрит–песок, детрит–макрофиты, песок–макрофиты, песок–коряги.
Значения показателя R и уровня достоверности p при попарном сравнении групп проб в ходе процедуры двухфакторного ANOSIM представлены в табл. 3.
Таблица 3.
Значения показателя R и уровня статистической достоверности для пар групп проб при попарном сравнении в ходе процедуры двухфакторного ANOSIM*
| Группы проб | Значение R | Уровень достоверности p, % |
|---|---|---|
| Тип субстрата | ||
| Камни–Детрит | 0.232 | 0.1 |
| Камни–Ил | 0.370 | 0.4 |
| Камни–Песок | 0.535 | 0.1 |
| Камни–Макрофиты | 0.341 | 0.1 |
| Камни–Коряги | 0.152 | 15.6 |
| Детрит–Ил | 0.064 | 32.5 |
| Детрит–Песок | 0.578 | 0.1 |
| Детрит–Макрофиты | 0.511 | 0.1 |
| Детрит–Коряги | 0.312 | 3.7 |
| Ил–Песок | 0.255 | 0.7 |
| Ил–Макрофиты | 0.419 | 0.1 |
| Ил–Коряги | 0.218 | 8.6 |
| Песок–Макрофиты | 0.551 | 0.1 |
| Песок–Коряги | 0.682 | 0.1 |
| Макрофиты–Коряги | 0.318 | 0.2 |
| Размер водотока | ||
| Ручьи–Прочие | 0.248 | 0.3 |
| Скорость течения | ||
| 0 – 0–0.1 м/с | 0.034 | 26 |
| 0 – 0.1–0.3 м/с | 0.076 | 8.7 |
| 0 – более 0.3 м/с | 0.397 | 0.1 |
| 0–0.1 – 0.1–0.3 м/с | 0.01 | 35.7 |
| 0–0.1 – более 0.3 м/с | 0.270 | 0.1 |
| 0.1–0.3 – более 0.3 м/с | 0.068 | 1 |
Процедура MDS и ANOSIM была проведена и для матрицы сходства, основанной на индексе гамма-плюс. Результаты сходны с полученными на основании матрицы Брея-Кертиса, однако для диаграмм ординации, полученных на основании индекса гамма-плюс, характерно более сильное перекрывание групп проб (рис. 3, рис. 4). Двухфакторный ANOSIM для групп проб, выделенных по бассейнам и типам субстрата, показал достоверный уровень различий для каждой группы проб, выделенной по бассейну. Для типов субстрата достоверно выделение всех групп, кроме пар камни–коряги (p – 9.15%), детрит–ил (p – 36.8%), детрит–коряги (p – 14.4%) и ил–коряги (p – 6.2%).
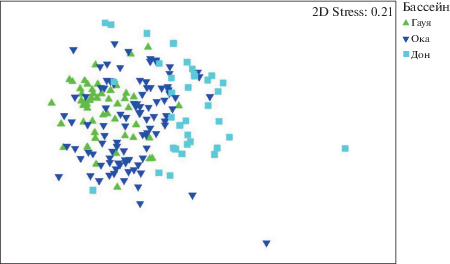
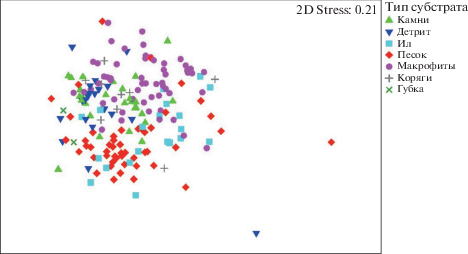
Для каждой группы проб, выделенных на основании типа субстрата, а также для группы проб, отобранных на ручьях, и группы проб, отобранных на участках с высокой скоростью течения, были выделены характерные и дифференцирующие таксоны ранга рода и вида на основании процедуры SIMPER. Для исключения влияния фактора бассейна на распределение видов по типам субстрата проведен двухфакторный перекрестный анализ для групп, выделенных по факторам бассейна и типа субстрата, размера водотока и скорости течения.
Таксоны ранга рода и вида, характерные для каждого типа субстрата, приведены в табл. 4.
Таблица 4.
Характерные таксоны для групп проб, выделенных по типу субстрата
| Тип субстрата | Перечень характерных таксонов |
|---|---|
| Камни | Baetis rhodani (Pictet 1843) (15.68)*, Hydropsyche sp. Pictet 1834 (7.97), Leuctra hippopus Kempny 1899 (2.95), Ancylus fluviatilis O.F. Muller 1774 (5.71), Heptagenia sp. Walsh 1863 (4.43) |
| Детрит | Baetis rhodani (Pictet 1843) (10.04), Brillia bifida (Kieffer 1909) (5.92), Leuctra hippopus Kempny 1899 (4.17), Elodes sp. Latreille 1796 (5.07) |
| Ил | Lithoglyphus naticoides (C. Pfeiffer 1828) (16.43), Chironomus sp. Meigen 1803 (4.64), Caenis horaria (Linnaeus 1758) (2.99) |
| Песок | Polypedilum scalaenum (Schrank 1803) (7.71), Lithoglyphus naticoides (C. Pfeiffer 1828) (11.26), Cryptochironomus gr. defectus (4.47), Cladotanytarsus gr. mancus (5.01), Tubifex newaensis (Michaelsen 1903) (3.59), Henslowiana (Supiniana) sp. Korniushin 1996 (4.60) |
| Макрофиты | Cloeon gr. dipterum (11.99), Baetis vernus Curtis 1834 (11.75), Cloeon luteolum (Muller 1776) (8.10), Laccophilus hyalinus (De Geer 1774) (4.71) Platycnemis pennipes (Pallas 1771) (3.19), Simuliidae gen. sp. (6.04) |
| Коряги | Paratanytarsus confusus Palmen 1960, Hydropsyche sp. Pictet 1834 (6.84), Macronychus quadrituberculatus Muller 1806 (5.80), Heptagenia flava Rostock 1878 (7.57) |
В табл. 5 приведены дифференцирующие виды для каждой из пар групп проб, выделенных по типу субстрата. В качестве дифференцирующих для субстрата были отобраны таксоны с отношением среднего показателя несходства к стандартному отклонению (${{\bar {\delta }}_{i}}$/SDi) более 0.5. Приведены значения среднего относительного численного обилия (в процентах от общей численности) для каждого таксона в каждой рассматриваемой группе проб и процент вклада каждого из них в объяснение различий между группами проб в паре. Для каждого дифференцирующего таксона ранга вида, группы видов или рода приведены таксоны высшего ранга (семейство, а также отряды для насекомых, подкласс – для кольчатых червей и класс – для моллюсков – ввиду неразработанности общепринятой системы отрядов для этих групп). Также для каждого дифференцирующего таксона приведена его жизненная форма (Приложение 1 ). Жизненные формы для всех таксонов, кроме личинок хирономид и жука Macronychus quadrituberculatus Muller 1806, приводятся по данным Чертопруда (Чертопруд М.В., 2006; Чертопруд, Песков, 2003). Жизненная форма M. quadrituberculatus приводится по данным Джонса (Johns, 2005).
Таблица 5.
Краткая аутэкологическая характеристика хирономид, являющихся дифференцирующими или характерными таксонами для анализируемых типов субстрата
| Название таксона | Краткая характеристика |
|---|---|
| Brillia bifida (Kieffer 1909) | Представители рода отмечены как ксилофаги-измельчители, обитающие на погруженной растительности (Armitage et al., 1995; Merrit et al., 1996) |
| Chironomus sp. Meigen 1803 | Преимущественно пелофилы (развиваются на илах и заиленных песках). Адаптированы к условиям высокого содержания органического вещества в воде и недостатка кислорода. Малоподвижны (строят трубки). Виды рода преимущественно являются детритофагами-фильтраторами или детритофагами-собирателями (Зинченко, 2011; Armitage et al., 1995; Merrit et al., 1996) |
| Cladotanytarsus gr. mancus | Обитает на заиленных участках со слабым течением, редко в обрастаниях камней (Зинченко, 2011). Малоподвижный фильтратор (Merrit et al., 1996) |
| Cryptochironomus gr. defectus | Эвритоп, преимущественно пелофил. Указан для илов, заиленного песка, глинистой почвы в зарослях макрофитов. (Зинченко, 2011). Хищник (Merrit et al., 1996). В качестве основного источника питания указываются олигохеты (Armitage et al., 1995) |
| Paratanytarsus confusus Palmen 1960 | Эвритоп. Отмечен на илах, мхе, макрофитах, обрастаниях камней. Высокой численности достигает на твердых субстратах, часто является частью литореофильного комплекса (Зинченко, 2011). Малоподвижный фильтратор (Armitage, 1995; Merrit et al., 1996) |
| Polypedilum scalaenum (Schrank 1803) | Указывается для заиленных, преимущественно песчанистых, биотопов в умеренно загрязненных реках, а также для заиленной почвы в зарослях макрофитов (Зинченко, 2011) |
| Procladius (Holotanypus) sp. Roback 1982 | Эвритопен, встречается на участках с замедленным течением на заиленных субстратах (илы, песок, гравий, глина, затопленные почвы) (Зинченко, 2011). Преимущественно хищник, в качестве основных компонентов питания указаны планктонные ракообразные, мелкие хирономиды, олигохеты. Однако есть данные о всеядности представителей данного рода, может быть собирателем. Подвижен (Armitage et al., 1995; Merrit, 1996) |
| Prodiamesa olivacea (Meigen 1818) | Встречается на разнообразных биотопах (указываются заиленный гравий, песок, почвы и глина), часто является массовым (Зинченко, 2011). Преимущественно детритофаг (Armitage et al., 1995) |
| Stenochironomus sp. Kieffer 1919 | Минирует древесные остатки (Зинченко, 2011; Armitage et al., 1995) |
Принимая во внимание видовое богатство семейства Chironomidae, а также широкий спектр условий, в которых обитают личинки, определить жизненную форму личинок хирономид представляется затруднительным. В табл. 5 приведена краткая аутэкологическая характеристика каждого таксона, являющегося характерным или дифференцирующим для каждого типа субстрата, основанная на литературных данных. Спектр питания и способ добывания пищи могут сильно варьировать даже у близких видов и в пределах одного вида в зависимости от доступности пищевых ресурсов, поэтому приведенная характеристика является ориентировочной.
В табл. 6 приведены характерные таксоны ранга вида и рода для ручьев и группы проб, отобранных на участках с высокой скоростью течения. Средний показатель внутригруппового сходства для ручьев составляет 13.07%, для высокой скорости течения – 13.74%. Показатель внутригруппового сходства для объединенной группы проб, отобранных на водотоках, более крупных, чем ручьи, составляет 8.18%, для групп проб, отобранных на течении скоростью менее 0.3 м/с – 8.09%.
Таблица 6.
Таксоны, характерные для ручьев и участков с высокой скоростью течения
| Характеристика участка отбора проб | Перечень характерных таксонов |
|---|---|
| Ручьи | Elodes sp. Latreille 1796 (10.27)*, Brillia bifida (Kieffer 1909) (5.89), Leuctra hippopus Kempny 1899 (3.50), Dixa sp. Meigen 1818 (6.52) |
| Участки с высокой скоростью течения (0.3–0.5 м/с) | Baetis rhodani (Pictet 1843) (11.18), Hydropsyche sp. Pictet 1834 (5.30), Simuliidae gen. sp. (8.10), Baetis vernus Curtis 1834 (3.42), Heptagenia sp. Walsh 1863 (2.78) |
В табл. 7 представлен перечень таксонов ранга вида и рода. Эти таксоны дифференцируют пробы, отобранные в ручьях и на участках с высокой скоростью течения, от других проб. Также приведены их жизненные формы. Из таблицы видно, что характерные и дифференцирующие таксоны для ручьев в целом совпадают и представлены в основном формами детритофагов, а также хирономидой – ксилофагом Brillia bifida (Kieffer 1909). Для ручьев характерны мелкомозаичные биотопы с большим количеством аллохтонной органики, и такие жизненные формы наилучшим образом адаптированы к обитанию в подобных условиях. Что касается форм, которые дифференцируют участки с высокой скоростью течения от участков с низкой скоростью течения или его отсутствием, то здесь с высоким течением ожидаемо ассоциированы прикрепленные формы, а плавающие собиратели скорее избегают высоких скоростей течения. Также с участками с высокой скоростью течения ассоциирована ритральная поденка-соскребатель (Baetis rhodani (Pictet 1843)), что также ожидаемо, т.к. высокие скорости течения характерны дли ритрали. Для таксонов, которые дифференцируют группы, выделенные по скорости течения, можно отметить в целом невысокие значения отношения ${{\bar {\delta }}_{i}}$/SDi. По этому показателю наилучшим дифференцирующим видом является ассоциированный с высокой скоростью течения хищный прикрепленный ручейник Hydropsyche sp. Pictet 1834.
Таблица 7.
Дифференцирующие таксоны для групп проб, отобранных на ручьях и на участках с высокой скоростью течения
| Вид, группа видов или род | Семейство | Отряд (подкласс – для Oligochaeta, класс – для моллюсков) | Жизненная форма | Относительное обилие, % от общей численности организмов в пробе | ${{\bar {\delta }}_{i}}$/SDi | Вклад в несходство между группами проб, % |
|---|---|---|---|---|---|---|
| Размер водотока* | ||||||
| Elodes sp. Latreille 1796 | Scirtidae | Coleoptera | Детритофаг-собиратель | 10.27а | 0.54 | 3.98 |
| 0.04б | ||||||
| Brillia bifida (Kieffer 1909) | Chironomidae | Diptera | См. табл. 4 | 5.89а | 0.51 | 2.53 |
| 0.74б | ||||||
| Leuctra hippopus | Leuctridae | Plecoptera | Подвижный | 3.50а | 0.75 | 1.71 |
| Kempny 1899 | детритофаг-собиратель | 0.92б | ||||
| Скорость течения** | ||||||
| Simuliidae gen. sp. | Simuliidae | Diptera | Малоподвижный | 8.10а | 0.44 | 5.21 |
| фильтратор | 0.33б | |||||
| Baetis vernus Curtis 1834 | Baetidae | Ephemeroptera | Плавающий | 3.42а | 0.50 | 4.62 |
| собиратель | 4.99б | |||||
| Baetis rhodani (Pictet 1843) | Baetidae | Ephemeroptera | Плавающий | 11.18а | 0.42 | 3.65 |
| альгофаг-соскребатель | 2.07б | |||||
| Hydropsyche sp. Pictet 1834 | Hydropsychidae | Trichoptera | Малоподвижный | 5.30а | 0.69 | 3.30 |
| хищник | 0.89б | |||||
| Cloeon luteolum | Baetidae | Ephemeroptera | Плавающий собиратель | 1.49а | 0.45 | 2.88 |
| (Muller 1776) | 3.44б | |||||
| Cloeon gr. dipterum | Baetidae | Ephemeroptera | Плавающий собиратель | 0.83а | 0.42 | 2.82 |
| 5.33б | ||||||
| Tubifex newaensis | Tubificidae | Oligochaeta | Роющий грунтоед | 0.66а | 0.40 | 1.67 |
| (Michaelsen 1903) | 1.89б | |||||
Анализ характерных и дифференцирующих таксонов ранга вида и рода был также проведен для водосборных бассейнов. Характерные таксоны приведены в табл. 8, дифференцирующие – в табл. 9. Следует отметить, что таксоны, включенные в списки характерных и дифференцирующих для бассейна, широко распространены и не являются эндемиками данных бассейнов. В нашем случае преобладание тех или иных таксонов в бассейне объясняется скорее неравномерным распределением субстратов в сериях проб. Так, таксоны, дифференцирующие бассейн Гауи от бассейнов Оки и Дона, представлены в основном формами детритофагов, а также плавающим соскребателем Baetis rhodani, а в наших сборах пробы с каменистых грунтов и детрита в основном сосредоточены в бассейне Гауи. Отдельно следует отметить вид брюхоногих моллюсков Lithoglyphus naticoides (C. Pfeiffer 1828), являющийся в наших сборах характерным и дифференцирующим для бассейна Дона. Это вид, происходящий из бассейна западного Черного моря, является активным инвайдером. Во второй половине 19 в. он проник в Западную Европу (Германия и Нидерланды), в начале 20 в. – во Францию (Mouthon, 2007). В бассейне Волги впервые отмечен в 1971 г., в дельте (Пирогов, 1972), в настоящее время распространился до Верхней Волги (Tyutin, Slynko, 2010; Perova et al., 2018). Однако в наших сборах он массово представлен только в реках бассейна Дона, т.е. в нативном для этого вида Азово-Черноморской бассейне.
Таблица 8.
Таксоны, характерные для бассейнов
| Название бассейна | Перечень характерных таксонов |
|---|---|
| Ока | Baetis vernus Curtis 1834 (6.88)*, Cloeon gr. dipterum (4.20), Cloeon luteolum (Muller 1776) (4.49), Polypedilum scalaenum (Schrank 1803) (3.86), Cryptochironomus gr. defectus (2.39), Laccophilus hyalinus (De Geer 1774) (2.72) |
| Дон | Lithoglyphus naticoides (C. Pfeiffer 1828) (28.45) |
| Гауя | Baetisrhodani (Pictet 1843) (15.68), Nemoura sahlbergi Morton 1896, Leuctra hippopus Kempny 1899 (3.32) |
Таблица 9.
Дифференцирующие таксоны для бассейнов
| Вид, группа видов или род | Семейство | Отряд (подкласс – для Oligochaeta, класс – для моллюсков) | Жизненная форма | Относительное обилие, % от общей численности организмов в пробе | ${{\bar {\delta }}_{i}}$/SDi | Вклад в несходство между группами проб, % |
|---|---|---|---|---|---|---|
| Гауяа–Окаб | ||||||
| Baetisrhodani (Pictet 1843) | Baetidae | Ephemeroptera | Плавающий альгофаг-соскребатель | 15.34а; 0.00б | 0.60 | 6.69 |
| Polypedilum scalaenum (Schrank 1803) | Chironomidae | Diptera | См. табл. 4 | 0.53а; 3.86б | 0.47 | 2.90 |
| Henslowiana (Supiniana) sp. Korniushin 1996 | Sphaeriidae | Bivalvia | Роющий фильтратор | 3.95а; 1.31б | 0.57 | 2.87 |
| Cryptochironomus gr. defectus | Chironomidae | Diptera | См. табл. 4 | 0.05а; 2.39б | 0.48 | 1.74 |
| Tubifex newaensis (Michaelsen 1903) | Tubificidae | Oligochaeta | Роющий грунтоед | 0.00а; 2.89б | 0.44 | 1.68 |
| Leuctra hippopus Kempny 1899 | Leuctridae | Plecoptera | Подвижный детритофаг-собиратель | 3.32а; 0.31б | 0.64 | 1.33 |
| Гауяа–Донв | ||||||
| Lithoglyphus naticoides (C. Pfeiffer 1828) | Lithoglyphidae | Gastropoda | Малоподвижный детритофаг | 0.00а; 28.45в | 0.94 | 18.73 |
| Cloeon gr. dipterum | Baetidae | Ephemeroptera | Плавающий собиратель | 0.43а; 9.73в | 0.55 | 5.16 |
| Baetis rhodani (Pictet 1843) | Baetidae | Ephemeroptera | Плавающий альгофаг-соскребатель | 15.34а; 0.00в | 0.53 | 5.13 |
| Henslowiana (Supiniana) sp. Korniushin 1996 | Sphaeriidae | Bivalvia | Роющий фильтратор | 3.95а; 0.00в | 0.60 | 3.46 |
| Caenis horaria (Linnaeus 1758) | Caenidae | Ephemeroptera | Роющий детритофаг | 0.00а; 2.52в | 0.56 | 2.01 |
| Окаб–Донв | ||||||
| Lithoglyphus naticoides (C. Pfeiffer 1828) | Lithoglyphidae | Gastropoda | Малоподвижный детритофаг | 0.00б; 28.45в | 0.74 | 13.94 |
Для групп проб, выделенных по типу субстрата, дополнительно был проведен анализ таксономического разнообразия. Были рассчитаны средние значения для каждой группы по следующим показателям: общее число таксонов ранга рода и вида, ожидаемое число таксонов на 30 особей (численность самой малочисленной пробы), индекс Шеннона. Средние значения показателей для каждой из групп проб приведены в табл. 10 (ранжированы по убыванию).
Таблица 10.
Средние показатели таксономического разнообразия и выравненности для каждого типа субстрата
| Тип субстрата | Число проб | Значение показателя ± ± стандартное отклонение |
|---|---|---|
| Общее число таксонов | ||
| Камни | 27 | 14.33 ± 5.31 |
| Коряги | 9 | 12.78 ± 5.17 |
| Детрит | 20 | 12.50 ± 4.70 |
| Макрофиты | 55 | 11.49 ± 4.57 |
| Ил | 20 | 10.75 ± 4.13 |
| Песок | 49 | 9.84 ± 5.52 |
| Ожидаемое число таксонов на 30 особей (ES(30)) | ||
| Камни | 27 | 8.22 ± 2.31 |
| Коряги | 9 | 8.15 ± 2.51 |
| Детрит | 20 | 7.74 ± 2.33 |
| Макрофиты | 55 | 7.33 ± 2.62 |
| Ил | 20 | 6.89 ± 2.23 |
| Песок | 49 | 6.53 ± 2.69 |
| Индекс Шеннона | ||
| Камни | 27 | 1.85 ± 0.46 |
| Коряги | 9 | 1.83 ± 0.36 |
| Детрит | 20 | 1.79 ± 0.39 |
| Макрофиты | 55 | 1.65 ± 0.51 |
| Ил | 20 | 1.58 ± 0.46 |
| Песок | 49 | 1.54 ± 0.62 |
Наибольший уровень достоверности был определен для распределения средних значений общего числа видов (p = 0.19%), низкий уровень достоверности определен для показателя среднего ожидаемого числа видов (p = 6.68%), по среднему значению индекса Шеннона группы различались с очень низким уровнем достоверности (p = 14.94%), однако ранжирование групп по убыванию значения по всем трем показателям оказалось одинаковым. Наибольшим таксономическим разнообразием отличались группы ритрали (камни и коряги), наименьшим – пелали (ил и песок).
ОБСУЖДЕНИЕ
Выявлено, что сходство видового состава внутри групп проб, выделенных на основании типа субстрата и водосборного бассейна, а также отобранных на ручьях и на участках с высокой скоростью течения (более 0.3 м/с), достоверно выше, чем в целом по всей выборке проб. Однако наблюдалось значительное перекрывание этих групп на диаграмме NMDS и высокая степень несходства внутри групп (низкое значение R). В опубликованных работах указывается на высокое разнообразие и слабую прогнозируемость видового состава сообществ текучих вод (Wright et al., 1984; Reynoldson et al., 2001; Heino et al., 2003; Heino et al., 2015). Это связано с рядом причин. Экосистемы текучих вод характеризуются частыми и мало предсказуемыми нарушениями, такими как наводнения и засухи. Животные организмы в таких системах могут с высокой степенью случайности элиминироваться из биотопа и реколонизировать его. Водные макробеспозвоночные обладают сравнительно высокой способностью к расселению и колонизации новых биотопов, а также зачастую характеризуются толерантностью к широкому спектру условий и широким ареалом. Эти факторы ослабляют действие локальных фильтров (Heino et al., 2003; Heino, Mykrä, 2008).
Приложение 1 . Дифференцирующие таксоны для групп проб, выделенных по типу субстрата
| Вид, группа видов или род | Семейство | Отряд (подкласс – для Oligochaeta, класс – для моллюсков) | Жизненная форма | Относительное обилие, % от общей численности организмов в пробе | ${{\bar {\delta }}_{i}}$/SDi | Вклад в несходство между группами проб, % |
|---|---|---|---|---|---|---|
| Камниа–детритб | ||||||
| Baetis rhodani (Pictet 1843) | Baetidae | Ephemeroptera | Плавающий альгофаг-соскребатель | 15.68а; 10.04б | 1.00 | 12.81 |
| Brillia bifida (Kieffer 1909) | Chironomidae | Diptera | См. табл. 4 | 2.93а; 5.29б | 0.72 | 4.50 |
| Elodes sp. Latreille 1796 | Scirtidae | Coleoptera | Детритофаг-собиратель | 1.38а; 5.07б | 0.51 | 3.80 |
| Hydropsyche sp. Pictet 1834 | Hydropsychidae | Trichoptera | Малоподвижный хищник | 7.97а; 0.67б | 0.78 | 3.19 |
| Leuctra hippopus Kempny 1899 | Leuctridae | Plecoptera | Подвижный детритофаг-собиратель | 2.95а; 4.17б | 0.90 | 3.14 |
| Heptagenia sp. Walsh 1863 | Heptageniidae | Ephemeroptera | Ползающий альгофаг-соскребатель | 4.43а; 0.34б | 0.57 | 2.26 |
| Камниа–илв | ||||||
| Hydropsyche sp. Pictet 1834 | Hydropsychidae | Trichoptera | Малоподвижный хищник | 7.97а; 0.00в | 0.99 | 5.75 |
| Chironomus sp. Meigen 1803 | Chironomidae | Diptera | См. табл. 4 | 0.00а; 4.64в | 0.52 | 3.84 |
| Tubifex newaensis (Michaelsen 1903) | Tubificidae | Oligochaeta | Роющий грунтоед | 0.61а; 4.12в | 0.50 | 3.66 |
| Ancylus fluviatilis O.F. Müller 1774 | Ancylidae | Gastropoda | Малоподвижный альгофаг-соскребатель | 5.71а; 0.00в | 0.60 | 3.44 |
| Caenis horaria (Linnaeus 1758) | Caenidae | Ephemeroptera | Роющий детритофаг | 0.82а; 2.99в | 0.53 | 2.50 |
| Детритб–илв | ||||||
| Prodiamesa olivacea (Meigen 1818) | Chironomidae | Diptera | См. табл. 4 | 0.06б; 1.96в | 0.73 | 4.96 |
| Henslowiana (Supiniana) sp. Korniushin 1996 | Sphaeriidae | Bivalvia | Роющий фильтратор | 2.71б; 1.91в | 0.58 | 4.83 |
| Baetisrhodani (Pictet 1843) | Baetidae | Ephemeroptera | Плавающий альгофаг-соскребатель | 10.04б; 0.80в | 0.57 | 3.44 |
| Камниа–песокг | ||||||
| Hydropsyche sp. Pictet 1834 | Hydropsychidae | Trichoptera | Малоподвижный хищник | 7.97а; 0.47г | 0.97 | 5.36 |
| Polypedilum scalaenum (Schrank 1803) | Chironomidae | Diptera | См. табл. 4 | 0.00а; 7.71г | 0.60 | 4.45 |
| Ancylus fluviatilis O.F. Müller 1774 | Ancylidae | Gastropoda | Малоподвижный альгофаг-соскребатель | 5.71а; 0.00г | 0.56 | 3.40 |
| Henslowiana (Supiniana) sp. Korniushin 1996 | Sphaeriidae | Bivalvia | Роющий фильтратор | 0.05а; 4.60г | 0.61 | 3.05 |
| Cryptochironomus gr. defectus | Chironomidae | Diptera | См. табл. 4 | 0.05а; 4.47г | 0.61 | 2.52 |
| Tubifex newaensis (Michaelsen 1903) | Tubificidae | Oligochaeta | Роющий грунтоед | 0.61а; 3.59г | 0.56 | 2.13 |
| Детритб–песокг | ||||||
| Henslowiana (Supiniana) sp. Korniushin 1996 | Sphaeriidae | Bivalvia | Роющий фильтратор | 2.71б; 4.60г | 0.78 | 4.13 |
| Baetis rhodani (Pictet 1843) | Baetidae | Ephemeroptera | Плавающий альгофаг-соскребатель | 10.04б; 0.02г | 0.52 | 3.91 |
| Polypedilum scalaenum (Schrank 1803) | Chironomidae | Diptera | См. табл. 4 | 0.00б; 7.71г | 0.53 | 3.25 |
| Leuctra hippopus Kempny 1899 | Leuctridae | Plecoptera | Подвижный детритофаг-собиратель | 4.17б; 0.28г | 0.58 | 1.81 |
| Илв–песокг | ||||||
| Polypedilum scalaenum (Schrank 1803) | Chironomidae | Diptera | См. табл. 4 | 0.00в; 7.71г | 0.59 | 4.99 |
| Chironomus sp. Meigen 1803 | Chironomidae | Diptera | См. табл. 4 | 4.64в; 3.56г | 0.67 | 4.98 |
| Tubifex newaensis (Michaelsen 1903) | Tubificidae | Oligochaeta | Роющий грунтоед | 4.12в; 3.59г | 0.65 | 4.97 |
| Pisidium amnicum (O.F. Müller 1774) | Sphaeriidae | Bivalvia | Роющий фильтратор | 5.42в; 2.48г | 0.54 | 4.28 |
| Cryptochironomus gr. defectus | Chironomidae | Diptera | См. табл. 4 | 0.35в; 4.47г | 0.66 | 3.00 |
| Caenis horaria (Linnaeus 1758) | Caenidae | Ephemeroptera | Роющий детритофаг | 2.99в; 0.65г | 0.53 | 2.45 |
| Procladius (Holotanypus) sp. Roback 1982 | Chironomidae | Diptera | См. табл. 4 | 1.65в; 1.85г | 0.61 | 2.18 |
| Камниа–макрофитыд | ||||||
| Baetis vernus Curtis. 1834 | Baetidae | Ephemeroptera | Плавающий собиратель | 1.82а; 11.75д | 0.63 | 6.79 |
| Hydropsyche sp. Pictet 1834 | Hydropsychidae | Trichoptera | Малоподвижный хищник | 7.97а; 0.83д | 1.10 | 6.46 |
| Cloeon gr. dipterum | Baetidae | Ephemeroptera | Плавающий собиратель | 1.23а; 11.99д | 0.64 | 5.34 |
| Cloeon luteolum (Muller 1776) | Baetidae | Ephemeroptera | Плавающий собиратель | 0.86а; 8.10д | 0.63 | 5.02 |
| Ancylus fluviatilis O.F. Müller 1774 | Ancylidae | Gastropoda | Малоподвижный альгофаг-соскребатель | 5.71а; 0.01д | 0.69 | 3.86 |
| Laccophilus hyalinus (De Geer. 1774) | Dytiscidae | Coleoptera | Плавающий хвататель | 0.00а; 4.71д | 0.51 | 3.10 |
| Bithynia tentaculata (Linnaeus 1758) | Bithyniidae | Gastropoda | Малоподвижный собиратель | 1.56а; 2.71д | 0.56 | 2.37 |
| Детритб–макрофитыд | ||||||
| Baetis vernus Curtis 1834 | Baetidae | Ephemeroptera | Плавающий собиратель | 2.61б; 11.75д | 0.62 | 7.53 |
| Cloeon gr. dipterum | Baetidae | Ephemeroptera | Плавающий собиратель | 0.12б; 11.99д | 0.61 | 4.57 |
| Cloeon luteolum (Muller 1776) | Baetidae | Ephemeroptera | Плавающий собиратель | 1.81б; 8.10д | 0.52 | 4.13 |
| Илв–макрофитыд | ||||||
| Cloeon gr. dipterum | Baetidae | Ephemeroptera | Плавающий собиратель | 1.29в; 11.99д | 0.63 | 6.67 |
| Baetis vernus Curtis 1834 | Baetidae | Ephemeroptera | Плавающий собиратель | 0.00в; 11.75д | 0.55 | 6.55 |
| Cloeon luteolum (Muller 1776) | Baetidae | Ephemeroptera | Плавающий собиратель | 0.22в; 8.10д | 0.55 | 4.56 |
| Chironomus sp. Meigen 1803 | Chironomidae | Diptera | См. табл. 4 | 4.64в; 0.58д | 0.55 | 3.95 |
| Песокг–макрофитыд | ||||||
| Baetis vernus Curtis 1834 | Baetidae | Ephemeroptera | Плавающий собиратель | 1.47г; 11.75д | 0.58 | 7.05 |
| Cloeon gr. dipterum | Baetidae | Ephemeroptera | Плавающий собиратель | 0.67г; 11.99д | 0.62 | 5.60 |
| Polypedilum scalaenum (Schrank 1803) | Chironomidae | Diptera | См. табл. 4 | 7.71г; 0.22д | 0.67 | 5.44 |
| Cloeon luteolum (Muller 1776) | Baetidae | Ephemeroptera | Плавающий собиратель | 0.44г; 8.10д | 0.60 | 4.90 |
| Cladotanytarsus gr. mancus | Chironomidae | Diptera | См. табл. 4 | 5.01г; 0.00д | 0.50 | 3.77 |
| Cryptochironomus gr. defectus | Chironomidae | Diptera | См. табл. 4 | 4.47г; 0.00д | 0.74 | 3.32 |
| Laccophilus hyalinus (De Geer 1774) | Dytiscidae | Coleoptera | Плавающий хвататель | 0.01г; 4.71д | 0.52 | 3.07 |
| Tubifex newaensis (Michaelsen 1903) | Tubificidae | Oligochaeta | Роющий грунтоед | 3.59г; 0.20д | 0.62 | 2.71 |
| Камниа–корягие | ||||||
| Paratanytarsus confusus Palmen 1960 | Chironomidae | Diptera | См. табл. 4 | 1.33а; 14.57е | 0.94 | 11.55 |
| Baetis rhodani (Pictet 1843) | Baetidae | Ephemeroptera | Плавающий альгофаг-соскребатель | 15.68а; 5.47е | 0.66 | 6.87 |
| Hydropsyche sp. Pictet 1834 | Hydropsychidae | Trichoptera | Малоподвижный хищник | 7.97а; 6.84е | 1.18 | 5.55 |
| Heptagenia flava Rostock 1878 | Heptageniidae | Ephemeroptera | Ползающий альгофаг-соскребатель | 0.00а; 7.57е | 0.55 | 4.81 |
| Heptagenia sp. Walsh 1863 | Heptageniidae | Ephemeroptera | Ползающий альгофаг-соскребатель | 4.43а; 2.32е | 0.62 | 3.71 |
| Macronychus quadrituberculatus Muller 1806 | Elmidae | Coleoptera | Малоподвижный ксилофаг | 0.00а; 5.80е | 0.61 | 3.69 |
| Atherix ibis (Fabricus 1798) | Athericidae | Diptera | Малоподвижный хищник | 2.47а; 3.09е | 0.86 | 2.70 |
| Детритб–корягие | ||||||
| Paratanytarsus confusus Palmen 1960 | Chironomidae | Diptera | См. табл. 4 | 0.15б; 14.57е | 1.12 | 14.37 |
| Baetis rhodani (Pictet 1843) | Baetidae | Ephemeroptera | Плавающий альгофаг-соскребатель | 10.04б; 5.47е | 0.92 | 6.77 |
| Hydropsyche sp. Pictet 1834 | Hydropsychidae | Trichoptera | Малоподвижный хищник | 0.67б; 6.84е | 1.57 | 4.92 |
| Brillia bifida (Kieffer 1909) | Chironomidae | Diptera | См. табл. 4 | 5.29б; 0.30е | 0.56 | 2.77 |
| Heptagenia sp. Walsh 1863 | Heptageniidae | Ephemeroptera | Ползающий альгофаг-соскребатель | 0.34б; 2.32е | 0.76 | 2.73 |
| Stenochironomus sp. Kieffer 1919 | Chironomidae | Diptera | См. табл. 4 | 0.00б; 2.23е | 0.57 | 2.71 |
| Atherix ibis (Fabricus 1798) | Athericidae | Diptera | Малоподвижный хищник | 0.32б; 3.09е | 0.76 | 2.55 |
| Leuctra hippopus Kempny 1899 | Leuctridae | Plecoptera | Подвижный детритофаг-собиратель | 4.17б; 0.76е | 0.60 | 2.20 |
| Илв–корягие | ||||||
| Chironomus sp. Meigen 1803 | Chironomidae | Diptera | См. табл. 4 | 4.64в; 5.24е | 0.68 | 6.90 |
| Heptagenia flava Rostock 1878 | Heptageniidae | Ephemeroptera | Ползающий альгофаг-соскребатель | 0.09в; 7.57е | 0.64 | 5.32 |
| Paratanytarsus confusus Palmen 1960 | Chironomidae | Diptera | См. табл. 4 | 0.00в; 14.57е | 0.60 | 4.25 |
| Macronychus quadrituberculatus Muller 1806 | Elmidae | Coleoptera | Малоподвижный ксилофаг | 0.00в; 5.80е | 0.71 | 4.07 |
| Pisidium amnicum (O. F. Müller 1774) | Sphaeriidae | Bivalvia | Роющий фильтратор | 5.42в; 1.94е | 0.54 | 3.79 |
| Hydropsyche sp. Pictet 1834 | Hydropsychidae | Trichoptera | Малоподвижный хищник | 0.00в; 6.84е | 0.78 | 2.92 |
| Песокг–корягие | ||||||
| Paratanytarsus confusus Palmen. 1960 | Chironomidae | Diptera | Малоподвижный фильтратор | 0.02г; 14.57е | 0.73 | 6.18 |
| Heptagenia flava Rostock 1878 | Heptageniidae | Ephemeroptera | Ползающий альгофаг-соскребатель | 0.10г; 7.57е | 0.72 | 6.05 |
| Macronychus quadrituberculatus Muller 1806 | Elmidae | Coleoptera | Малоподвижный ксилофаг | 0.00г; 5.80е | 0.80 | 4.60 |
| Polypedilum scalaenum (Schrank 1803) | Chironomidae | Diptera | См. табл. 4 | 7.71г; 0.00е | 0.60 | 4.52 |
| Hydropsyche sp. Pictet 1834 | Hydropsychidae | Trichoptera | Малоподвижный хищник | 0.47г; 6.84е | 1.04 | 3.80 |
| Chironomus sp. Meigen 1803 | Chironomidae | Diptera | См. табл. 4 | 3.56г; 5.24е | 0.50 | 3.06 |
| Cryptochironomus gr. defectus | Chironomidae | Diptera | См. табл. 4 | 4.47г; 0.00е | 0.63 | 2.66 |
| Henslowiana (Supiniana) sp. Korniushin 1996 | Sphaeriidae | Bivalvia | Роющий фильтратор | 4.60г; 0.00е | 0.52 | 2.41 |
| Tubifex newaensis (Michaelsen 1903) | Tubificidae | Oligochaeta | Роющий грунтоед | 3.59г; 0.00е | 0.53 | 2.13 |
| Макрофитыд–корягие | ||||||
| Simuliidae gen. sp. | Simuliidae | Diptera | Малоподвижный фильтратор | 6.04д; 5.95е | 0.61 | 7.81 |
| Heptagenia flava Rostock 1878 | Heptageniidae | Ephemeroptera | Ползающий альгофаг-соскребатель | 0.20д; 7.57е | 0.80 | 7.10 |
| Baetis vernus Curtis 1834 | Baetidae | Ephemeroptera | Плавающий собиратель | 11.75д; 0.83е | 0.58 | 6.60 |
| Cloeon gr. dipterum | Baetidae | Ephemeroptera | Плавающий собиратель | 11.99д; 0.30е | 0.60 | 5.86 |
| Macronychus quadrituberculatus Muller 1806 | Elmidae | Coleoptera | Малоподвижный ксилофаг | 0.00д; 5.80е | 0.89 | 5.42 |
| Cloeon luteolum (Muller 1776) | Baetidae | Ephemeroptera | Плавающий собиратель | 8.10д; 0.00е | 0.55 | 4.63 |
| Paratanytarsus confusus Palmen 1960 | Chironomidae | Diptera | Малоподвижный фильтратор | 0.23д; 14.57е | 0.73 | 4.50 |
| Hydropsyche sp. Pictet 1834 | Hydropsychidae | Trichoptera | Малоподвижный хищник | 0.83д; 6.84е | 0.95 | 3.53 |
В современной литературе уделяется значительное внимание концепции мета-сообществ (metacommunity concept), согласно которой мета-сообщество – набор локальных сообществ, объединенных миграционными потоками особей (Leibold et al., 2004). Структура мета-сообществ определяется взаимодействием таких факторов, как отбор видов (species sorting), эффект массы (mass effects), ограничение распространения (dispersal limitation) и динамика пятен (patch dynamics) (Heino, 2011). Отбор видов происходит под воздействием факторов среды, позволяя закрепляться в сообществе видам, способным существовать при данных условиях. Эффект массы проявляется, когда локальное сообщество влияет на видовой состав соседних сообществ за счет распространения видов с высокой расселительной способностью, независимо от биотических и абиотических факторов среды. Ограничение расселения влияет на состав сообществ в случае, если организмы с низкой расселительной способностью не способны достичь местообитания, благоприятного с точки зрения факторов среды, из-за его удаленности. Динамика пятен связана с взаимодействием процессов колонизации местообитаний и конкуренции, при этом изолированные или сильно нарушенные местообитания заселяются более успешными организмами-колонизаторами, которые в менее изолированных или более зрелых сообществах могут замещаться более конкурентоспособными видами (Leibold et al., 2004; Heino et al., 2015a). На структуру большинства метасообществ эти факторы влияют в совокупности, при этом их выраженность может быть различной в разных пространственных масштабах. Так, ограничение распространения может превалировать в масштабе от 100 тыс. км2 до континентального. Отбор видов с наибольшей силой проявляется в масштабе от 100 до 100 тыс. км2, в котором наиболее выражен градиент факторов среды. Значимость эффекта массы возрастает в масштабе 1–100 км2, в случае, когда обмен видами между соседними сообществами за счет их дисперсии может перекрыть действие факторов среды. Наконец, динамика пятен влияет на состав сообществ в самых малых пространственных масштабах (Heino, 2011).
Таким образом, на состав сообщества влияют как факторы среды, так и положение сообщества в пространстве (его удаленность от других сообществ, расположение на территории водосбора), и выраженность их влияния зависит от масштаба, на котором рассматривается структура данного сообщества. При увеличении пространственного масштаба возрастает роль ограниченности расселительной способности организмов, на малом масштабе играет роль эффект массы, гомогенизирующий состав сообществ, причем для организмов с высокой расселительной способностью масштаб, на котором может проявляться эффект массы, больше, чем для организмов с низкой расселительной способностью (Heino et al., 2015a).
На локальном уровне состав сообществ в первую очередь определяется локальными факторами среды (тип субстрата, химический состав воды, скорость течения и т.д.). Однако и факторы среды обнаруживают достаточно слабую связь с видовым составом сообщества. На основании канонического анализа соответствий в работе Хейно и Микра (Heino, Mykrä, 2008) подсчитано, что локальными факторами объясняется от 9.8 до 31.7% видового состава сообществ. В региональном масштабе возрастает влияние фактора расселительной способности организмов. Пространственные факторы могут в меньшей степени влиять на распределение организмов с высокой расселительной способностью (среди гидробионтов это в первую очередь крылатые амфибиотические насекомые). Поскольку на разных пространственных масштабах видовой состав связан с различными группами факторов, в исследованиях стоит учитывать географический охват. Модели прогнозирования сообществ, пригодные для одного региона, могут снижать свою эффективность в другом (Mykrä et al., 2007).
На основании материалов данной работы удалось выделить набор характерных и дифференцирующих таксонов ранга вида или рода для каждого типа субстрата, а также для ручьев и участков с высокой скоростью течения. Эти списки таксонов представлены типичным набором жизненных форм, описанных ранее М.В. Чертопрудом (Чертопруд, Песков, 2003; Чертопруд, 2006) (табл. 3–6).
Для субстратов, представляющих собой твердую опору (камни и коряги) дифференцирующими являются такие жизненные формы, как плавающий альгофаг-соскребатель, ползающий альгофаг-соскребатель, малоподвижный альгофаг-соскребатель, малоподвижный хищник, т.е. различные типы соскребателей и прикрепленных организмов. В табл. 3 можно видеть, что для камней и коряг приведен сходный набор жизненных форм, за исключением малоподвижного ксилофага (жук семейства Elmidae Macronychus quadrituberculatus), представленного на корягах, но не на камнях. Для коряг в качестве характерного и дифференцирующего вида отмечена хирономида Paratanytarsus confusus Palmen 1960. Согласно литературным источникам, этот вид часто достигает высокой численности на твердых субстратах, являясь частью литореофильного комплекса.
Для субстратов, образующих комплекс пелали (ил и песок), дифференцирующими являются следующие жизненные формы: роющий фильтратор, роющий грунтоед, такие рода хирономид как Polypedilum Kieffer 1912, Cryptochironomus Kieffer 1918, Cladotanytarsus Kieffer 1921, Chironomus Meigen 1803, Procladius Skuse 1889 (табл. 3). Для песка и ила характерен сходный набор жизненных форм, однако по нашим данным хирономиды родов Polypedilum, Cryptochironomus и Cladotanytarsus характерны для песка, хирономида рода Chironomus – в большей степени для ила, а поденка рода Caenis Stephens 1835 является характерным и дифференцирующим таксоном только для ила.
Фиталь представлена в первую очередь плавающими жизненными формами, такими как плавающий собиратель (в первую очередь, поденки семейства Baetidae), плавающий хвататель (имаго и личинки плавунцов Dytiscidae). Характерной жизненной формой для растительности является также малоподвижный хвататель, в наших сборах представленный личинкой стрекозы Platycnemis pennipes (Pallas 1771), а также малоподвижные фильтраторы – личинки семейства Simuliidae – общая форма для растительности и реофильного комплекса камней и коряг.
Для детрита характерные и дифференцирующие формы – различные детритофаги-собиратели (веснянки рода Leuctra Stephens 1836, личинки жуков семейства Scirtidae), хирономиды, питающиеся на затопленной древесине (в наших сборах – представитель подсемейства Orthocladiinae Brillia bifida).
Характерные и дифференцирующие формы для ручьев, формирующих мелкомозаичные биотопы с большим количеством грубого аллохтонного детрита, ожидаемо представлены детритофагами-собирателями, для участков с высокой скоростью течения – прикрепленными фильтраторами, малоподвижными хищниками и разновидностями соскребателей.
Таким образом, для каждого типа субстрата выявлен набор жизненных форм, приуроченных к данному типу субстрата и максимально адаптированных нему. Это различные прикрепленные, ползающие и малоподвижные формы, а также соскребатели для ритрали (камни и коряги), предоставляющей плотную опору для обитающих на этом субстрате организмов; плавающие малоподвижные формы для фитали, роющие фильтраторы и собиратели для рыхлых грунтов (ил и песок), детритофаги-собиратели, а также ксилофаги для детрита. Эти результаты позволяют предположить, что тип субстрата является эффективным экологическим фильтром, на локальном уровне определяя присутствие жизненных форм, адаптированных к условиям этого типа субстрата.
Полученные нами результаты подтверждают результаты ряда работ, опубликованных ранее (Чертопруд, Песков, 2003; Чертопруд, 2006, 2011; Палатов и др., 2016; Чертопруд, Палатов, 2017; Палатов, Чертопруд, 2018 и др.), что указывает на применимость использованного нами математического аппарата к сообществам макрозобентоса пресных текучих вод.
Для каждого типа субстрата удалось выделить набор жизненных форм, приуроченных к нему, однако группы проб, выделенных по этому фактору, сильно перекрываются между собой, причем каждая группа характеризуется высокой гетерогенностью. Это может объясняться наличием в речном макробентосе значительного числа эврибионтных видов, не приуроченных строго к какому-либо определенному биотопу. Кроме того, по табл. 3 видно, что даже дифференцирующие для типа субстрата жизненные формы могут встречаться на различных типах субстрата, не всегда характерных для данной жизненной формы. Вероятно, в этом проявляется “эффект массы”, когда одно сообщество влияет на видовой состав соседнего за счет распространения видов за пределы участка с пригодными для этих видов абиотическими условиями.
Наиболее хорошо различимые типы субстрата – макрофиты и песок, по результатам ANOSIM достоверно отличающиеся от других типов во всех случаях попарного сравнения. Камни и коряги представляют собой группу субстратов литореофильного комплекса со сходным набором жизненных форм. Их отличительная черта – наличие твердой опоры для беспозвоночных и связанный с этим набор жизненных форм соскребателей и прикрепленных фильтраторов. Статистически недостоверными оказалось выделение групп проб “детрит–ил” и “ил–коряги”. Первый случай, вероятно, объясняется неравномерным распределением групп, связанных с данными типами субстратов, по водосборным бассейнам. Второй – заселением заиленных древесных остатков организмами пелофильного комплекса. Также зачастую, особенно в ручьях, сложно разграничить сообщества таких субстратов как камни и детрит, т.к. они представляют собой единый мелкомозаичный комплекс.
На основании факторов размера водотока и скорости течения в наших сборах достоверно выделяются только группы проб, отобранных на ручьях и на высокой (более 0.3 м/с) скорости течения. Большая часть проб отбиралась на равнинных водотоках среднего размера, и для них характерна более или менее плавная смена таксономического состава сообществ. Также в наших сборах мал разброс по скорости течения, но даже здесь пробы, отобранные на участках с относительно высоким течением (0.3–0.5 м/с), образуют достоверно отличающуюся группу.
Также при двухфакторном ANOSIM по факторам “бассейн” и “тип субстрата” не выявлено статистически значимых различий между типами субстратов “детрит” и “ил”: по-видимому, в серии проб из бассейна Гауи недостаточно представлен тип субстрата “ил”. Хорошо различимы типы субстрата “песок”, “камни” и “макрофиты”, что подтверждает диаграмму неметрического многомерного шкалирования.
Картина сходства проб и значимость выделения сообществ на основании матрицы сходства, построенной с использованием индекса гамма-плюс, т.е. с учетом уровня таксономического определения организмов, достаточно близка к картине, построенной с использованием индекса сходства Брея-Кертиса. Можно заключить, что для экспресс-анализа бентосных сообществ допустимо использовать определение уровня рода или семейства. Это подтверждается литературными данными (Heino, Soininen, 2007).
Необходимо отметить, что в данной работе не учитывался фактор сезонности формирования сообществ макрозообентоса, поэтому затруднительно сказать, каков в данном случае вклад сезонности в формирование различий между сообществами. Безусловно, этот вопрос требует дополнительных исследований в дальнейшем. Общие закономерности формирования сообществ на уровне жизненных форм не противоречат опубликованным ранее литературным данным, поэтому использованные математические методы можно использовать для анализа макрозообентоса пресных текучих вод. Однако требуется совершенствование методики и апробация ее на более широком материале, с учетом большего спектра факторов.
ЗАКЛЮЧЕНИЕ
Сообщества донных макробеспозвоночных на различных субстратах статистически достоверно различаются. Проведенный анализ позволил выявить характерные и дифференцирующие для каждого субстрата жизненные формы. Однако группы проб на разных типах субстрата отличаются высокой степенью несходства проб внутри группы и значительно перекрываются, что указывает на постепенность изменения видового состава.
Статистически достоверно различаются группы проб из различных водосборных бассейнов. Различия могут быть связаны с расселительной способностью организмов. Однако этот вопрос требует более полного изучения с охватом широкого спектра водотоков различных типов из разных водосборных бассейнов.
Распределение таксономического состава на уровне более высоких таксонов, чем вид и род, в целом соответствует распределению на видовом уровне и для экспресс-анализа сообществ может использоваться определение до семейства. Однако диаграмма ординации, полученная на основании индекса сходства гамма-плюс, который учитывает таксономический ранг, отличается более высокой степенью перекрывания групп проб, и для полного понимания закономерностей формирования сообществ определение до вида предпочтительно.
Несмотря на выявленные статистически достоверные закономерности, сообщества текучих вод отличаются высоким разнообразием и слабой предсказуемостью видового состава. На их состав и структуру влияет множество факторов, обусловленных не только условиями среды, но и положением сообщества относительно других сообществ водотока и водосборного бассейна. Сообщества водотоков образуют систему, оказывая влияние на видовой состав друг друга посредством расселения видов, и выраженность этого влияния возрастает с уменьшением пространственного масштаба, на котором рассматривается распределение видового состава. Таксономический состав сообществ текучих вод слабо прогнозируем. Тем не менее, можно выявить ассоциации видов и жизненных форм, достоверно связанные с изучаемыми факторами среды. Несмотря на высокое таксономическое разнообразие текучих вод, возможно составить достаточно короткий перечень “ядерных” таксонов, наиболее характерных для изучаемых условий. Составление таких перечней для различных географических зон и типов водных объектов может представлять практический интерес для природоохранной деятельности.
Список литературы
Жадин В.И., 1940. Фауна рек и водохранилищ // Труды зоологического института АН СССР. Т. 5. № 3–4. С. 519–919.
Жадин В.И., Герд С.В., 1961. Реки, озера и водохранилища СССР, их фауна и флора. М.: Государственное учебно-педагогическое издательство Министерства просвещения РСФСР. 599 с.
Жирков И.А., 2017. Био-география общая и частная: суши, моря и континентальных водоемов. М.: Товарищество научных изданий КМК. 568 с.
Зинченко Т.Д., 2011. Эколого-фаунистическая характеристика хирономид (Diptera, Chironomidae) малых рек бассейна Средней и Нижней Волги (атлас). Тольятти: Кассандра. 258 с.
Определитель пресноводных беспозвоночных России и сопредельных территорий. Т. 3. Паукообразные. Низшие насекомые. 1997. Цалолихин С.Я. (ред.). СПб.: ЗИН РАН. 439 с.
Определитель пресноводных беспозвоночных России и сопредельных территорий. Т. 4. Высшие насекомые. Двукрылые. 1999. Цалолихин С.Я. (ред.). СПб.: ЗИН РАН. 998 с.
Определитель пресноводных беспозвоночных России и сопредельных территорий. Т. 5. Высшие насекомые. 2001. Цалолихин С.Я. (ред.). СПб.: ЗИН РАН. 836 с.
Определитель пресноводных беспозвоночных России и сопредельных территорий. Т. 6. Моллюски. Полихеты. Немертины. 2004. Цалолихин С.Я. (ред.). СПб.: Наука. 528 с.
Палатов Д.М., Чертопруд М.В., 2018. Сообщества макрозообентоса плотных грунтов водотоков восточного Причерноморья // Экология. № 1. С. 66–73.
Палатов Д.М., Чертопруд М.В., Фролов А.А., 2016. Фауна и типы сообществ макрозообентоса мягких грунтов водотоков горных районов восточного Причерноморья // Биология внутренних вод. № 2. С. 45–55.
Пирогов В.В., 1972. О нахождении Lithoglyphus naticoides в дельте Волги // Зоологический журнал. Т. 51. № 6. С. 912–913.
Чертопруд М.В., 2006. Анализ жизненных форм реофильного макробентоса: новый подход к классификации сообществ // Журнал общей биологии. Т. 67. № 3. С. 190–197.
Чертопруд М.В., 2011. Разнообразие и классификация реофильных сообществ макробентоса средней полосы Европейской России // Журнал общей биологии. Т. 72. № 1. С. 51–73.
Чертопруд М.В., Палатов Д.М., 2017. Сообщества макрозообентоса малых водотоков восточных Балкан // Биология внутренних вод. № 3. С. 46–46.
Чертопруд М.В., Песков К.В., 2003. Географические параллели организации литореофильных сообществ малых рек Восточной Европы и Северной Азии // Журнал общей биологии. Т. 64. № 1. С. 78–87.
Шитиков В.К., Зинченко Т.Д., Розенберг Г.С., 2011. Макроэкология речных сообществ: концепции, методы, модели. Тольятти: Кассандра. 255 с.
Armitage P.D., Cranston P.S., Pinder L.C.V., 1995. The Chironomidae: Biology and ecology of non-biting midges. London: Chapman & Hall. 572 p.
Clarke K.R., Somerfield P.J., Chapman M.G., 2006. On resemblance measures for ecological studies, including taxonomic dissimilarities and a zero-adjusted Bray–Curtis coefficient for denuded assemblages // Journal of Experimental Marine Biology and Ecology. V. 330. Issue 1. P. 55–80.
Clarke K.R., Warwick R.M., 2001. Change in marine communities: an approach to statistical analysis and interpretation. Plymouth: Plymouth Marine Laboratory. 176 p.
Diamond J.M., 1975. Assembly rules of species communities // Ecology and evolution of communities. Cody M.L., Diamond J.M. (Ed.). Cambridge, Massachusetts, USA: Harvard University Press. P. 342–344.
Heino J., 2011. A macroecological perspective of diversity patterns in the freshwater realm // Freshwater Biology. V. 56. Issue 9. P. 1703–1722.
Heino J., Melo A.S., Bini L.M., Altermatt F., Al-Shami S.A. et al., 2015. A comparative analysis reveals weak relationships between ecological factors and beta diversity of stream insect metacommunities at two spatial levels // Ecology and Evolution. V. 5. Issue 6. P. 1235–1248.
Heino J., Melo A.S., Siqueira T., Soininen J., Valanko S., Bini L.M., 2015a. Metacommunity organisation, spatial extent and dispersal in aquatic systems: patterns, processes and prospects // Freshwater Biology. V. 60. Issue 5. P. 845–869.
Heino J., Muotka T., Mykra H., Paavola R., Hamalainen H., Koskenniemi E., 2003. Defining macroinvertebrate assemblage types of headwater streams: implications for bioassessment and conservation // Ecological Applications. V. 13. Issue 3. P. 842–852.
Heino J., Mykrä H., 2008. Control of stream insect assemblages: roles of spatial configuration and local environmental factors // Ecological Entomology. V. 33. Issue 5. P. 614–622.
Heino J., Soininen J., 2007. Are higher taxa adequate surrogates for species-level assemblage patterns and species richness in stream organisms // Biological Conservation. V. 137. Issue 1. P. 78–89.
Johns T., 2005. A study of the aquatic riffle beetle Macronychus quadrituberculatus on the Holy Brook, Berkshire. A dissertation submitted to the University of Bristol in accordance with the requirements of the masters degree of Ecology and Management of the Natural Environment in the Faculty of Biological Sciences. 83 p.
Keddy P.A., 1992. Assembly and response rules: two goals for predictive community ecology // Journal of Vegetation Science. V. 3. Issue 2. P. 157–164.
Leibold M.A., Holyoak M., Mouquet N., Amarasekare P., Chase J.M., et al., 2004. The metacommunity concept: a framework for multi-scale community ecology // Ecology Letters. V. 7. Issue 7. P. 601–613.
Merritt R.W., Wallace J.R., Higgins M.J., Alexander M.K., Berg M.B., Morgan W.T., Cummins K.W., Vandeneeden B., 1996. Procedures for the functional analysis of invertebrate communities of the Kissimmee river-floodplain ecosystem // Florida Scientist. V. 59. № 4. P. 216–274.
Mouthon J., 2007. Lithoglyphus naticoides (Pfeiffer) (Gastropoda: Prosobranchia): distribution in France, population dynamics and life cycle in the Saône river at Lyon (France) // Annales de Limnologie – International Journal of Limnology. V. 43. № 1. P. 53–59.
Mykrä H., Heino J., Muotka T., 2007. Scale-related patterns in the spatial and environmental components of stream macroinvertebrate assemblage variation // Global Ecology and Biogeography. V. 16. Issue 2. P. 149–159.
Ormerod S.J., Edwards R.W., 1987. The ordination and classification of macroinvertebrate assemblages in the catchment of the River Wye in relation to environmental factors // Freshwater Biology. V. 17. Issue 3. P. 533–546.
Perova S.N., Pryanichnikova E.G., Tyutin A.V., 2018. Expansion of the Range of the Black Sea Snail Lithoglyphus naticoides (C. Pfieffer, 1828) (Mollusca: Gastropoda: Lithoglyphidae) and Associated Trematode Species in the Upper Volga Basin // Inland Water Biology. V. 11. Issue 2. P. 234–235.
Poff N.L., 1997. Landscape filters and species traits: towards mechanistic understanding and prediction in stream ecology // Journal of the North American Benthological Society. V. 16. № 2. P. 391–409.
Reynoldson T.B., Rosenberg D.M., Resh V.H., 2001. Comparison of models predicting invertebrate assemblages for biomonitoring in the Fraser River catchment, British Columbia // Canadian Journal of Fisheries and Aquatic Sciences. V. 58. № 7. P. 1395–1410.
Tyutin A.V., Slynko Yu.V., 2010. The First Finding of the Black Sea Snail Lithoglyphus naticoides (Gastropoda) and Its Associated Species Specific Trematoda in the Upper Volga Basin // Russian Journal of Biological Invasions. V. 1. Issue 1. P. 45–49.
Wright J.F., Moss D., Armitage P.D., Furse M.T., 1984. A preliminary classification of running-water sites in Great Britain based on macro-invertebrate species and the prediction of community type using environmental data // Freshwater Biology. V. 14. Issue 3. P. 221–256.
Дополнительные материалы отсутствуют.
Инструменты
Зоологический журнал