

Зоологический журнал, 2020, T. 99, № 8, стр. 938-950
Периферический отдел слуховой системы морских птиц (чистиковые – Alcidae). 1. Морфометрия наружного уха с ушной птерилией и их функциональные возможности
В. Д. Анисимов *
Биологический факультет Московского государственного университета
имени М.В. Ломоносова
119991 Москва, Россия
* E-mail: flora_oa@mail.ru
Поступила в редакцию 06.09.2019
После доработки 24.12.2019
Принята к публикации 21.02.2020
Аннотация
Для трех видов семейства чистиковых (Alcidae) проведено морфологическое описание с метрическими измерениями наружного уха и перьев ушной птерилии. Приведены линейные размеры структур, представлены данные о взаимном их расположении. Наружное слуховое отверстие окружает неширокая аптериальная кожная поверхность. Канал слухового прохода за наружным слуховым отверстием в кожном покрове резко расширяется в широкую подкожную полость, которая в сечении превышает площадь слухового отверстия в 4.5–5.0 раз. Проекция наружного слухового отверстия на крупное отверстие в предтимпанальную полость перекрывает его на 1/3. По топографическим и размерным параметрам канала слухового прохода под водой исключается возможность смыкания его стенок и перекрытия слухового прохода. Ушные перья образуют вокруг слухового отверстия плотную стенку с невысоким перьевым сводом и имеют воздушную полость над слуховым отверстием. Структура бородок апикальных участков опахала ушных и контурных перьев такова, что образует водоотталкивающий наружный слой, исключающий быстрое намокание перьев. Под сводом ушной птерилии и в слуховом проходе наружного уха сохраняется воздушный канал, который в водной среде может создавать воздушную “пробку”. В целом, у изученных чистиковых наружное ухо с ушной птерилией имеет морфологическую водозащитную адаптацию, с сохранением воздушной звукопроводящей функции.
Анализ анатомического и морфологического строения слуховой системы групп птиц, разных по таксономическому положению, но сходных по среде обитания, позволяет выявить общие закономерности морфологической адаптации их слухового анализатора (Ильичев, 1962; Анисимов, 1978; Барсова, 1980, 1984; Голубева, Ямалова, 1980 и др.). Чистиковые и пингвины, высоко специализированные к водной среде, являются удобными объектами для сравнения морфологической и функциональной адаптаций слуховой системы к условиям среды (Ильичев, 1961; Анисимов, 1976, 1986; Kartaschew, Iljitschew, 1964).
Первые работы, которые включали описания анатомии периферического отдела слуховой системы птиц, в том числе чистиковых и пингвинов, проводили на материале с продолжительным сроком хранения в формалиновом фиксаторе (Ильичев, 1961, 1962 и др.). Морфометрические данные, полученные на фиксированном материале, не пригодны для использования в сравнительной и функциональной морфологии.
У абсолютного большинства морских птиц слабо изучены не только частотно-пороговая чувствительность слуха, но и анатомическое строение периферического отдела слуховой системы. До настоящего времени считается, что птицы, адаптированные к водной среде, обладают “плохим”, нечувствительным слухом, а их наружное ухо относится к примитивному, водозащитному типу (Ильичев, 1962, 1966; Wever et al., 1969). При этом у чистиковых птиц звуковая сигнализация и голосовое общение хорошо развиты, что указывает на акустическую чувствительность их периферического отдела слуховой системы (Карташев, Ильичев, 1966; Олексенко, 1982; Klenova, Kolesnikova, 2013 и др.).
Ушная птерилия птиц как часть перьевого покрова по функциональной значимости представляет собой аналог ушной раковины млекопитающих и адаптирована к акустической функции наружного уха птиц (Ильичев, Извекова, 1961; Ильичев, 1962, 1966). В работах по изучению различных характеристик оперения пингвинов и некоторых других птиц отсутствуют упоминания об ушной птерилии (Штефан, 1977 и др.).
До недавнего времени оставалось не известным, слышат ли птицы в воде (Dooling, Therrien, 2012). В поведенческом эксперименте (Johansen et al., 2016) показано наличие слуховой чувствительности в воде у большого баклана (Phalacrocorax carbo sinensis (Shaw et Nodder 1801)). Для морских и водных видов птиц нет четких и однозначных морфометрических и электрофизиологических доказательств способности воспринимать звук под водой, не определены и пути его возможной передачи к слуховым рецепторам внутреннего уха.
Предлагаемая работа по сравнительной морфологии и размерным характеристикам наружного уха и ушной птерилии трех видов чистиковых птиц выполнена на основе эколого-морфологических методик, разработанных в лаборатории орнитологии МГУ им. М.В. Ломоносова (Ильичев, 1971). Эти методики апробированы в работах по сравнительной морфологии слуховой системы птиц, в том числе и водных (Барсова, 1973; Анисимов, 1978, 1986 и др.). Результаты морфометрических исследований на свежем, не фиксированном в формалине, наружном ухе чистиков более точно характеризуют строение этого участка слуховой системы птиц.
МАТЕРИАЛЫ И МЕТОДИКА
Весь материал по периферическому отделу слуховой системы чистиковых собран и сразу обработан в течение экспедиционных сезонов 1985–1987 гг. на побережье Чукотского п-ова. Морфометрические данные по строению наружного уха получены только на свежем материале в течение первых суток после гибели птицы. Измерения и сравнительное морфологическое описание наружного уха проводили на четырех особях каждого из трех видов чистиковых птиц: ипатки (Fratercula corniculata (J.F. Naumann 1821)), толстоклювой кайры ((Uria lomvia (Linnaeus 1758), чукотский подвид U. l. heckeri (Portenko 1944)) и тихоокеанского чистика (Cepphus columba (Pallas 1881)).
По разработанной нами методике для морфометрических исследований структур периферического отдела слуховой системы птиц (Анисимов, 1974; 1978) выполнены промеры и описание анатомического строения наружного уха чистиковых. Голову мертвой птицы жестко фиксировали за клюв специальным зажимом, который крепился на шарнирной подставке и обеспечивал возможность препарировать, измерять и фотографировать нужные участки наружного уха и перьев под любым наклоном.
Все угловые измерения, измерение площади проекции контура слухового отверстия, площади аптериальной поверхности кожного покрова рядом с отверстием, площади проекции контура поперечного сечения канала слухового прохода, получены по прорисовкам их проекций на бумаге с миллиметровой сеткой. Для этого использовали бинокулярный микроскоп МБС-2 при 10-, 20- и 40-кратном увеличении. Микроскоп был снабжен измерительной линейкой окуляр-микрометра с точностью до 0.01 мм, рисовальным аппаратом РА-1 и фотонасадкой для микроскопа. Расстояние между контурными и ушными перьями измеряли с точностью до 0.02 мм, линейные характеристики перьев ушной птерилии – с точностью до 0.5 мм, их наклон к плоскости кожного покрова – с точностью до 2°.
Проведены измерения длины и площади проекции контура отверстия слухового прохода в предтимпанальную полость в направлении к барабанной перепонке.
На живой птице измерен наклон продольной оси канала слухового прохода к поверхности кожного покрова с отверстием наружного уха. Выполнено топографическое описание того, как совмещаются наружное слуховое отверстие с более крупным отверстием слухового прохода в предтимпанальную полость. Определены координаты слухового отверстия наружного уха на голове у живых птиц.
Из собственно птерилии, передних, верхних, нижних и задних перьев первого ряда вокруг слухового отверстия и аптериальной поверхности одного уха каждой птицы взято по 2 пера. На правом и левом ухе каждой птицы измерены по 4 контурных пера и по 16 перьев ушной птерилии. В общей сложности от одной птицы взято для измерения по 20 перьев. У трех видов чистиков в сумме использовано 240 перьев. Таким образом, на наружном ухе и ушной птерилии 12 птиц выполнено 960 промеров.
Результаты основных измерений ушной птерилии и участков слухового прохода наружного уха чистиковых представлены в таблицах. Сравнительные параметры размеров контурных и перьев ушной птерилии чистиков приведены на диаграммах.
РЕЗУЛЬТАТЫ
Наружное ухо птиц – функциональное понятие. Это канал проведения звукового сигнала из внешней среды к барабанной перепонке (Ильичев, 1962, 1966). Наружное слуховое отверстие птиц контактирует с внешней, воздушной средой. Противоположный конец слухового прохода заканчивается предтимпанальной полостью с барабанной перепонкой. У видов птиц, адаптированных к водной среде, наружное слуховое отверстие прикрыто перьевым покровом и не имеет непосредственного контакта с водой (Анисимов, 1986; Анисимов, Ильичев, 1980).
Контурные перья на голове чистиковых птиц прикрывают рыхлым слоем ушную птерилию и скрывают слуховое отверстие. У птиц место слухового отверстия на голове определено координатами Х и Y. По оси Z–Х слуховое отверстие расположено со смещением от центра глазного яблока в каудальном направлении и параллельно вектору линии смыкания нижней и верхней частей клюва. Ось X–Y направлена перпендикулярно вниз от точки Х (рис. 1). У ипатки координаты слухового отверстия по оси Z–Х 17 ± 2 мм, по оси X–Y 16 ± ± 2 мм, у толстоклювой кайры по оси Z–Х 18 ± 2 мм, по оси X–Y 10 ± 2 мм, у чистика по оси Z–Х 12 ± ± 2 мм, по оси X–Y 10 ± 2 мм.
Рис. 1.
Голова толстоклювой кайры с координатами расположения слухового отверстия. Пунктирной линией обозначена продольная ось клюва, кругом ‑ место наружного слухового отверстия. Z–X – расстояние от центра глазного яблока (Z) в каудальном направлении параллельно продольной оси клюва. X–Y – расстояние до слухового отверстия с вертикальным смещением вниз от точки Х.

Наружное слуховое отверстие располагается на аптериальном участке поверхности кожи. Внутренний контур первого ряда ушных перьев определяет место аптериальной поверхности и слухового отверстия наружного уха (рис. 2А). По размерам площадь аптериальной поверхности меньше площади поверхности слухового отверстия. Относительно общей площади поверхности слухового отверстия, ограниченной первым рядом перьев ушной птерилии (рис. 2а, 2с), площадь аптериальной поверхности составляет в среднем 27% у ипатки, 39% у толстоклювой кайры, 36% у обыкновенного чистика. Размеры слухового отверстия правого и левого уха чистиковых птиц могут различаться в пределах 3–10% (табл. 1).
Рис. 2.
Наружное слуховое отверстие и канал слухового прохода ипатки: А – освобожденное от перьев отверстие наружного уха. а – диаметр слухового отверстия. В – начало расширенного канала слухового прохода со стороны слухового отверстия. b – диаметр сечения канала слухового прохода. с – граница первого ряда перьев ушной птерилии вокруг слухового отверстия.
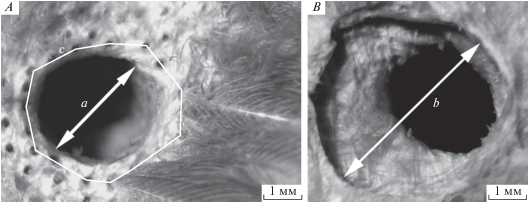
Таблица 1.
Размеры площади аптериальной поверхности, наружного слухового отверстия и участков слухового прохода наружного уха чистиковых птиц
| Площадь, мм2 | Номер особи, ухо | Среднее значение | |||||||
|---|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | ||||||
| правое | левое | правое | левое | правое | левое | правое | левое | ||
| Ипатка (Fratercula corniculata) | |||||||||
| Аптериальной поверхности и слухового отверстия | 5.2 | 5.2 | 4.1 | 4.2 | 5.1 | 4.8 | 6.3 | 6.6 | 5.2 |
| Проекции наружного слухового отверстия | 3.8 | 3.7 | 3.2 | 3.6 | 3.9 | 3.1 | 4.2 | 4.8 | 3.78 |
| Проекции поперечного сечения слухового прохода | 14.1 | 14.8 | 20.0 | 19.2 | 15.4 | 16.0 | 20.0 | 21.8 | 17.66 |
| Проекции отверстия слухового прохода в предтимпанальную полость | 8.1 | 8.0 | 6.7 | 6.6 | 7.0 | 6.5 | 7.8 | 8.0 | 7.33 |
| Толстоклювая кайра (Uria lomvia heckeri) | |||||||||
| Аптериальной поверхности и слухового отверстия | 6.0 | 6.9 | 8.0 | 8.5 | 7.8 | 7.7 | 6.0 | 6.6 | 7.2 |
| Проекции наружного слухового отверстия | 4.1 | 3.9 | 4.8 | 4.6 | 5.0 | 4.8 | 4.0 | 3.9 | 4.38 |
| Проекции поперечного сечения слухового прохода | 26.0 | 25.0 | 22.3 | 20.0 | 19.3 | 18.5 | 20.4 | 19.2 | 21.33 |
| Проекции отверстия слухового прохода в предтимпанальную полость | 6.0 | 6.5 | 7.0 | 6.9 | 7.0 | 7.2 | 6.5 | 6.0 | 6.63 |
| Тихоокеанский чистик (Cepphus columba) | |||||||||
| Аптериальной поверхности и слухового отверстия | 6.4 | 6.5 | 7.5 | 8.0 | 8.2 | 6.7 | 6.8 | 6.7 | 7.1 |
| Проекции наружного слухового отверстия | 4.2 | 4.4 | 5.0 | 4.8 | 4.4 | 4.1 | 5.2 | 5.0 | 4.63 |
| Проекции поперечного сечения слухового прохода | 19.4 | 19.6 | 20.0 | 21.0 | 24.1 | 23.5 | 20.0 | 20.3 | 20.98 |
| Проекции отверстия слухового прохода в предтимпанальную полость | 7.0 | 7.1 | 8.0 | 8.0 | 6.7 | 6.8 | 7.9 | 7.7 | 7.7 |
Рельеф слухового прохода не имеет перегибов и сужений. Длина канала слухового прохода составляет 8–14 мм. Слуховой проход за слуховым отверстием начинается расширенной подкожной полостью и заканчивается крупным отверстием в замкнутую предтимпанальную полость с барабанной перепонкой. Вид расширенного участка канала в подкожном слое, сразу за наружным слуховым отверстием, показан на рис. 2B.
Площадь проекции контура поперечного сечения слухового прохода больше площади наружного слухового отверстия в 4.5–4.8 раза. Размер отверстия в предтимпанальную полость больше размера наружного слухового отверстия, у ипатки в 1.9 раза, у толстоклювой кайры в 1.5 раза, у тихоокеанского чистика в 1.6 раза (табл. 1).
Угол наклона продольной оси канала слухового прохода к плоскости наружного слухового отверстия на аптериальной поверхности составляет 40°–55°. Проекция наружного слухового отверстия на отверстие слухового прохода в предтимпанальную полость топографически совпадает на 1/3. Эти параметры могут служить подтверждением отсутствия у чистиковых птиц топографической несовместимости наружного и внутреннего отверстий в слуховом проходе наружного уха.
Ипатка
Морфологическое описание основных параметров контурного пера и ушной птерилии чистиков мы более подробно рассматриваем на примере ипатки – модельного вида чистиковых птиц. Метрические параметры ушных и контурных перьев ипатки даны в табл. 2.
Таблица 2.
Размеры перьев ушной птерилии и контурных перьев рядом с ними у ипатки
| Номер особи, ухо | Перо | ||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| контурное | переднее | верхнее | нижнее | заднее | |||||||||||||||||
| Длина опахала, мм | Ширина опахала, мм | Длина очина, мм | Угол бородки, ° | Длина опахала, мм | Ширина опахала, мм | Длина очина, мм | Угол бородки, ° | Длина опахала, мм | Ширина опахала, мм | Длина очина, мм | Угол бородки, ° | Длина опахала, мм | Ширина опахала, мм | Длина очина, мм | Угол бородки, ° | Длина опахала, мм | Ширина опахала, мм | Длина очина, мм | Угол бородки, ° | ||
| 1 | Правое | 18.0 | 6.0 | 1.4 | 40 | 9.5 | 2.5 | 1.6 | 17 | 9.8 | 2.0 | 1.6 | 20 | 12.0 | 1.8 | 1.8 | 15 | 10.0 | 1.4 | 1.5 | 18 |
| 18.5 | 5.3 | 1.3 | 45 | 10.6 | 2.5 | 1.6 | 18 | 8.1 | 1.2 | 1.3 | 18 | 11.7 | 2.6 | 2.1 | 17 | 11.2 | 2.8 | 1.2 | 17 | ||
| Левое | 16.8 | 8.0 | 1.1 | 32 | 14.0 | 2.8 | 1.5 | 20 | 9.8 | 2.0 | 1.6 | 17 | 11.0 | 1.9 | 1.8 | 14 | 9.8 | 1.5 | 1.4 | 24 | |
| 16.5 | 7.8 | 1.2 | 45 | 12.6 | 3.0 | 1.7 | 15 | 10.5 | 1.8 | 1.6 | 18 | 14.5 | 2. 0 | 2.0 | 19 | 8.5 | 1.5 | 1.5 | 20 | ||
| 2 | Правое | 16.5 | 6.3 | 1.1 | 40 | 13.6 | 3.6 | 1.4 | 18 | 10.5 | 2.0 | 1.1 | 24 | 11.6 | 2.1 | 2.0 | 20 | 8.6 | 1.3 | 1.4 | 19 |
| 17.0 | 6.8 | 1.4 | 42 | 13.1 | 2.2 | 1.4 | 20 | 9.2 | 1.3 | 1.4 | 20 | 11.2 | 1.9 | 1.9 | 18 | 9.3 | 1.3 | 1.3 | 18 | ||
| Левое | 21.0 | 7.5 | 1.0 | 44 | 9.6 | 1.5 | 1.2 | 12 | 8.5 | 1.7 | 1.6 | 20 | 11.2 | 2.2 | 2.1 | 18 | 11.5 | 2.2 | 1.2 | 18 | |
| 19.0 | 7.1 | 1.0 | 30 | 13.1 | 2.1 | 1.6 | 20 | 14.0 | 2.1 | 1.4 | 18 | 10.2 | 2.0 | 1.8 | 23 | 9.7 | 1.3 | 1.4 | 14 | ||
| 3 | Правое | 15.8 | 7.0 | 1.1 | 30 | 14.1 | 3.0 | 1.2 | 22 | 8.2 | 1.2 | 1.5 | 22 | 11.3 | 2.1 | 2.0 | 25 | 9.7 | 1.6 | 1.3 | 16 |
| 16.8 | 5.8 | 1.6 | 35 | 11.0 | 1.7 | 1.8 | 19 | 8.0 | 1.5 | 1.2 | 19 | 10.7 | 1.6 | 1.7 | 16 | 11.8 | 3.4 | 1.2 | 21 | ||
| Левое | 13.0 | 7.0 | 0.9 | 42 | 11.3 | 1.9 | 1.8 | 20 | 11.4 | 1.8 | 1.3 | 20 | 12.4 | 1.7 | 1.6 | 18 | 9.8 | 2.3 | 1.2 | 29 | |
| 11.5 | 6.2 | 0.7 | 43 | 10.3 | 1.7 | 1.2 | 17 | 9.3 | 1.3 | 1.1 | 23 | 11.5 | 2.3 | 2.1 | 16 | 10.6 | 1.7 | 1.1 | 17 | ||
| 4 | Правое | 12.5 | 8.0 | 1.0 | 44 | 12.1 | 1.8 | 1.9 | 21 | 7.5 | 1.2 | 1.5 | 20 | 12.7 | 1.6 | 1.5 | 19 | 11.4 | 1.7 | 0.8 | 14 |
| 14.7 | 9.9 | 0.9 | 41 | 14.3 | 2.9 | 1.5 | 18 | 11.8 | 1.3 | 1.2 | 21 | 10.7 | 2.0 | 1.4 | 20 | 11.3 | 2.6 | 1.1 | 22 | ||
| Левое | 19.2 | 11 | 1.0 | 39 | 14.9 | 3.4 | 1.4 | 15 | 12.0 | 1.9 | 2.1 | 14 | 12.1 | 1.7 | 1.6 | 21 | 11.4 | 2.4 | 1.1 | 19 | |
| 16.5 | 8.4 | 1.0 | 35 | 12.3 | 1.6 | 1.3 | 18 | 13.1 | 2.8 | 1.6 | 18 | 10.6 | 1.5 | 1.6 | 18 | 9.6 | 2.9 | 1.0 | 20 | ||
| Среднее значение | 16.4 | 7.4 | 1.1 | 39 | 12.3 | 2.38 | 1.5 | 18 | 10.1 | 1.7 | 1.4 | 19 | 11.6 | 1.9 | 1.8 | 18 | 10.2 | 2.0 | 1.2 | 19 | |
Контурные перья формируют плотный наружный слой перьевого покрова над слуховым отверстием (рис. 1). Эти перья расположены рядом с перьями ушной птерилии и длиннее их, с симметричным, широким опахалом (рис. 3A). Расстояние между перьями на уровне очина 0.9–1.0 мм. Наклон базальной части пера к кожному покрову 60°–65°, наклон медиальной части пера – 40°–50°. Вершины опахал контурных перьев прикрывают перья ушной птерилии. Длина очина контурного пера 0.7–1.6 мм. Бородки в средней части опахала контурного пера расположены к стержню под углом 30°–45°. Лучи на бородках опахала вблизи и вдоль всего стержня пера удлиненные, серповидные, с длиной в среднем 0.6 мм. Сцепленные бородки выражены только по месту расположения удлиненных лучей. Вдоль всего стержня эта узкая часть опахала и формирует уплотненный участок поверхности пера. Бородки, направленные к внешнему краю опахала, нитевидной формы, не сцеплены. Лучи на таких участках бородок укорачиваются до 0.02 мм и приобретают вид мелких ворсинок (рис. 3E). Между такими лучами сохраняется тонкая прослойка воздуха.
Рис. 3.
Ушное и контурное перья ипатки (А, B, E) и чистика (C, D): А, C – контурное перо; В, D – ушное перо, E – лучи на вершинах бородок опахала пера.

Ушные перья ипатки двумя рядами плотно окружают наружное слуховое отверстие и аптериальную поверхность кожи (рис. 2А). В первом ряду расположено от 16 до 18 перьев. Расстояние между перьями от 0.4 до 0.6 мм. По расположению вокруг слухового отверстия перья можно охарактеризовать как передние, верхние, нижние и задние ушные. Ушная птерилия характеризуется перьями с более коротким, узким опахалом и с плотным расположением бородок к стержню пера. Очины ушных перьев длиннее очинов контурных перьев (табл. 2). Перья дифференцированы от соседних контурных перьев. Бородки ушных перьев сохраняют структурное строение бородок контурного пера (рис. 3B, 3D).
Передние ушные перья первого ряда растут по самому краю стенки слухового отверстия в количестве до 4 штук, и расположены плотнее по сравнению с другими участками. Расстояние между перьями 0.40–0.45 мм. Просвет между перьями на уровне кожного покрова до 0.2 мм.
Эти перья имеют небольшой сагиттальный изгиб. Опахала передних ушных перьев короче опахал контурного оперения, и они более узкие. Длина очина может достигать 1.9 мм. Угол наклона бородки к стержню в средней части опахала 12°–22° (табл. 2). Наклон базальной части пера к поверхности кожного покрова и слуховому отверстию – от 55° до 65°. В основании пера бородки не опушены (рис. 3).
Не сцепленные концы бородок в апикальной части ушных перьев достигают нижней поверхности накрывающих их контурных перьев. Удлиненные лучи на бородках опахала передних ушных перьев остаются только на участке опахала вдоль стержня пера (рис. 3E). На конце бородки лучи укорочены (рис. 3E), расстояние между ними не превышает 0.02 мм.
Только между верхними, нижними, задними перьями ушной птерилии и слуховым отверстием располагается аптериальная поверхность кожи (рис. 2A). Длина и ширина этих ушных перьев меньше размеров контурных перьев (табл. 2). Наклон верхних, нижних и задних ушных перьев к кожной поверхности более пологий по сравнению с наклоном передних ушных перьев и не превышает 60°. Вершины перьев достигают опахал накрывающих их контурных перьев. В отличие от контурных, бородки опахала других ушных перьев остаются узкими. В медиальной части опахала бородки отходят от стержня под более острым углом 18°–19° и не опушены в базальной части опахала.
Перья второго ряда ушной птерилии не отличаются структурой и размерами от ушных перьев первого ряда.
Передние и верхние перья ушной птерилии ипатки формируют вертикальную уплотненную переднюю и боковую верхнюю части стенки свода над слуховым отверстием, а нижние и задние ушные перья – более пологую (угол наклона 55°–60°) нижнюю боковую и заднюю стенки воздушного свода над слуховым отверстием. Высота перьевого свода над слуховым отверстием у разных особей от 2.0 до 2.5 мм.
У пера ушной птерилии имеется укороченное дополнительное опахало из 4–5 бородок, растущее от стержня на уровне его очина. Отсутствуют пуховые перья рядом с ушной птерилией.
Толстоклювая кайра
Контурные перья и перья ушной птерилии по расположению вокруг слухового отверстия и строению совпадают с характеристиками тех же перьев у ипатки.
У кайры, как у ипатки, два ряда ушных перьев. В первом ряду от 16 до 20 перьев. Расстояние между перьями 0.4–0.7 мм. Передние ушные перья расположены плотнее остальных ушных перьев. Эти перья укорочены, с узким опахалом, их очины длиннее очина контурного пера (табл. 3). Наклон бородок опахала к стержню пера 19°–32°.
Таблица 3.
Размеры перьев ушной птерилии и контурных перьев рядом с ними у толстоклювой кайры
| Номер особи, ухо | Перо | ||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| контурное | переднее | верхнее | нижнее | заднее | |||||||||||||||||
| Длина опахала, мм | Ширина опахала, мм | Длина очина, мм | Угол бородки, ° | Длина опахала, мм | Ширина опахала, мм | Длина очина, мм | Угол бородки, ° | Длина опахала, мм | Ширина опахала, мм | Длина очина, мм | Угол бородки, ° | Длина опахала, мм | Ширина опахала, мм | Длина очина, мм | Угол бородки, ° | Длина опахала, мм | Ширина опахала, мм | Длина очина, мм | Угол бородки, ° | ||
| 1 | Правое | 10.8 | 5.2 | 0.9 | 40 | 12.4 | 4.9 | 1.5 | 32 | 9.5 | 3.0 | 1.2 | 18 | 13.0 | 4.0 | 0.8 | 19 | 13.0 | 6.0 | 0.8 | 24 |
| 12.5 | 5.4 | 0.7 | 38 | 9.7 | 5.0 | 1.0 | 21 | 10.1 | 2.7 | 1.0 | 17 | 10.0 | 4.2 | 1.2 | 15 | 16.0 | 6.5 | 0.9 | 20 | ||
| Левое | 12.6 | 4.8 | 0.6 | 44 | 13.5 | 6.0 | 1.0 | 19 | 9.8 | 3.6 | 1.2 | 24 | 12.3 | 3.5 | 1.5 | 16 | 11.0 | 4.0 | 1.1 | 18 | |
| 14.5 | 4.8 | 0.8 | 41 | 10.9 | 2.1 | 1.5 | 28 | 14.0 | 2.1 | 1.3 | 20 | 10.6 | 3.8 | 1.3 | 18 | 13.5 | 4.1 | 1.6 | 18 | ||
| 2 | Правое | 13.9 | 4.6 | 0.8 | 39 | 9.8 | 2.2 | 2.1 | 15 | 9.5 | 2.2 | 1.4 | 19 | 12.6 | 3.0 | 1.2 | 20 | 10.8 | 5.6 | 1.2 | 19 |
| 14.8 | 6.6 | 0.7 | 39 | 9.2 | 1.6 | 1.6 | 18 | 10.3 | 2.0 | 1.4 | 18 | 12.8 | 3.4 | 1.6 | 17 | 9.8 | 4.1 | 0.8 | 21 | ||
| Левое | 15.6 | 6.2 | 0.7 | 40 | 9.9 | 1.9 | 1.5 | 32 | 8.8 | 1.8 | 1.5 | 18 | 12.6 | 3.9 | 1.5 | 21 | 13.0 | 4.5 | 1.1 | 22 | |
| 14.2 | 5.9 | 0.7 | 36 | 12.5 | 2.6 | 1.5 | 30 | 9.3 | 2.7 | 1.2 | 14 | 11.4 | 4.4 | 1.2 | 18 | 12.0 | 3.8 | 1.0 | 19 | ||
| 3 | Правое | 16.0 | 6.5 | 0.8 | 38 | 12.5 | 3.4 | 1.3 | 20 | 10.2 | 3.3 | 1.1 | 16 | 11.1 | 4.6 | 1.2 | 16 | 14.0 | 6.6 | 1.2 | 20 |
| 14.0 | 7.0 | 0.8 | 41 | 12.1 | 3.4 | 1.2 | 30 | 9.4 | 2.9 | 1.0 | 21 | 10.0 | 5.6 | 0.9 | 19 | 9.5 | 6.7 | 1.2 | 15 | ||
| Левое | 11.2 | 6.0 | 0.7 | 43 | 10.3 | 4.0 | 0.9 | 21 | 10.8 | 2.8 | 1.1 | 29 | 11.4 | 5.5 | 1.2 | 17 | 10.5 | 5.9 | 1.0 | 15 | |
| 10.8 | 5.8 | 0.6 | 40 | 8.4 | 2.2 | 0.8 | 22 | 11.1 | 3.4 | 1.0 | 17 | 9.6 | 4.9 | 0.9 | 25 | 10.2 | 4.8 | 0.7 | 28 | ||
| 4 | Правое | 10.9 | 8.0 | 0.7 | 44 | 10.9 | 4.2 | 1.4 | 19 | 8.8 | 2.9 | 1.2 | 14 | 9.5 | 4.7 | 0.8 | 23 | 8.8 | 3.7 | 1.0 | 24 |
| 11.1 | 7.3 | 0.7 | 45 | 10.8 | 4.4 | 1.4 | 16 | 9.9 | 2.8 | 1.1 | 22 | 9.9 | 3.9 | 0.9 | 20 | 10.0 | 5.0 | 0.8 | 19 | ||
| Левое | 10.4 | 5.7 | 0.6 | 41 | 10.4 | 4.9 | 1.3 | 18 | 11.0 | 3.0 | 1.3 | 19 | 10.7 | 6.0 | 0.6 | 18 | 9.5 | 4.0 | 0.9 | 21 | |
| 11.4 | 7.3 | 0.8 | 39 | 11.6 | 4.0 | 1.2 | 20 | 9.9 | 2.6 | 1.0 | 20 | 11.2 | 5.0 | 0.8 | 16 | 10.3 | 3.8 | 0.8 | 20 | ||
| Среднее значение | 12.8 | 6.1 | 0.7 | 40 | 10.9 | 3.5 | 1.3 | 22 | 10.1 | 2.7 | 1.2 | 19 | 11.2 | 4.4 | 1.1 | 19 | 11.1 | 4.9 | 0.8 | 20 | |
Наклон всех ушных перьев к отверстию уха у кайры совпадает с наклоном перьев ушной птерилии ипатки. Высота перьевого свода над слуховым отверстием до 2.50 мм. Вершины ушных перьев упираются в накрывающие их контурные перья.
Имеется укороченное дополнительное опахало, растущее от стержня на уровне очина пера. Отсутствуют пуховые перья рядом с ушными перьями. Метрические характеристики ушных и контурных перьев толстоклювой кайры даны в табл. 3.
Тихоокеанский чистик
Контурные перья с широким опахалом. Эти перья по длине и ширине опахала отличаются от контурного пера ипатки (рис. 3С). Лучи на концах бородок мелкие, ворсистой структуры. Расстояние между контурными перьями 0.6–0.8 мм. Наклон бородки к стержню пера в среднем 36°, что способствует ослаблению пера на изгиб.
Ушные перья вокруг слухового отверстия представлены одним рядом. Количество перьев в ряду от 22 до 24 штук. Размеры опахала и очина ушных перьев совпадают с размерными данными контурных перьев (рис. 3C, 3D). Передние ушные перья плотно расположены по краю слухового отверстия. Наклон бородок к стержню пера 31°–32°. Расстояние между перьями до 0.55–0.6 мм. Наклон стенок перьевого свода ушной птерилии над слуховым отверстием уха совпадает с наклоном ушной птерилии ипатки и кайры. Высота перьевого свода над слуховым отверстием в среднем 2.5 мм. Существует дополнительное опахало на внутренней стороне ушного пера. В основании опахала пера бородки опушены. Рядом с передними и верхними ушными перьями находятся длинные нитевидные перья, которых не имеют кайра и ипатка. Метрические характеристики ушных и контурных перьев даны в табл. 4.
Таблица 4.
Размеры перьев ушной птерилии и контурных перьев рядом с ними у тихоокеанского чистика
| Номер особи, ухо | Перо | ||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| контурное | переднее | верхнее | нижнее | заднее | |||||||||||||||||
| Длина опахала, мм | Ширина опахала, мм | Длина очина, мм | Угол бородки, ° | Длина опахала, мм | Ширина опахала, мм | Длина очина, мм | Угол бородки, ° | Длина опахала, мм | Ширина опахала, мм | Длина очина, мм | Угол бородки, ° | Длина опахала, мм | Ширина опахала, мм | Длина очина, мм | Угол бородки, ° | Длина опахала, мм | Ширина опахала, мм | Длина очина, мм | Угол бородки, ° | ||
| 1 | Правое | 9.5 | 7.5 | 1.0 | 30 | 10.3 | 3.9 | 0.9 | 28 | 12.0 | 4.5 | 0.9 | 34 | 10.9 | 4.6 | 1.1 | 30 | 11.6 | 5.0 | 1.0 | 30 |
| 9.4 | 7.3 | 0.7 | 42 | 10.0 | 4.7 | 0.9 | 32 | 8.7 | 4.4 | 0.8 | 33 | 10.7 | 4.5 | 1.0 | 32 | 9.0 | 5.7 | 0.6 | 32 | ||
| Левое | 10.9 | 3.4 | 1.1 | 32 | 9.9 | 3.7 | 0.9 | 33 | 10.1 | 3.7 | 0.9 | 37 | 10.9 | 4.6 | 1.0 | 29 | 8.9 | 4.8 | 0.6 | 29 | |
| 10.2 | 3.0 | 1.0 | 41 | 10.1 | 3.6 | 1.0 | 32 | 10.8 | 3.0 | 1.0 | 34 | 10.8 | 5.5 | 0.8 | 30 | 8.7 | 6.0 | 0.6 | 30 | ||
| 2 | Правое | 8.3 | 4.6 | 0.8 | 31 | 9.5 | 3.4 | 1.0 | 23 | 8.7 | 2.3 | 0.7 | 32 | 10.7 | 5.2 | 1.0 | 32 | 0.4 | 5.9 | 0.7 | 32 |
| 9.6 | 5.3 | 0.6 | 36 | 9.8 | 4.4 | 0.8 | 35 | 9.3 | 5.3 | 0.8 | 27 | 9.1 | 4.5 | 0.8 | 34 | 10.4 | 5.0 | 0.8 | 34 | ||
| Левое | 9.2 | 5.3 | 0.7 | 31 | 10.3 | 5.4 | 0.8 | 39 | 9.4 | 5.4 | 0.7 | 28 | 9.7 | 5.3 | 0.9 | 27 | 8.8 | 5.8 | 0.8 | 27 | |
| 9.2 | 6.7 | 0.5 | 30 | 10.0 | 4.1 | 0.9 | 33 | 8.3 | 5.2 | 0.6 | 30 | 9.0 | 4.2 | 0.8 | 33 | 9.0 | 5.1 | 0.7 | 33 | ||
| 3 | Правое | 10.0 | 3.9 | 1.1 | 40 | 10.2 | 3.4 | 1.0 | 32 | 9.2 | 4.5 | 1.5 | 35 | 11.5 | 3.6 | 0.9 | 35 | 8.5 | 4.4 | 0.6 | 35 |
| 10.1 | 5.8 | 0.6 | 32 | 8.7 | 4.0 | 1.2 | 33 | 9.3 | 5.2 | 0.6 | 33 | 10.0 | 3.3 | 0.6 | 42 | 9.2 | 4.4 | 0.8 | 42 | ||
| Левое | 9.4 | 5.3 | 0.8 | 31 | 9.3 | 4.5 | 0.8 | 27 | 10.0 | 4.8 | 0.7 | 37 | 10.7 | 3.7 | 1.0 | 31 | 9.4 | 4.0 | 0.8 | 31 | |
| 9.2 | 4.5 | 0.7 | 33 | 11.0 | 4.8 | 1.0 | 29 | 9.6 | 5.0 | 0.7 | 35 | 10.8 | 4.1 | 0.9 | 41 | 9.7 | 4.1 | 1.0 | 41 | ||
| 4 | Правое | 10.5 | 5.0 | 0.8 | 40 | 9.9 | 2.8 | 0.7 | 31 | 10.1 | 4.0 | 0.8 | 32 | 11.2 | 3.8 | 1.2 | 31 | 8.6 | 5.0 | 0.8 | 31 |
| 9.2 | 4.5 | 0.7 | 45 | 10.4 | 3.1 | 0.8 | 27 | 9.4 | 4.7 | 0.6 | 27 | 10.7 | 3.3 | 1.2 | 32 | 9.7 | 5.1 | 0.7 | 32 | ||
| Левое | 10.2 | 4.2 | 0.8 | 42 | 10.2 | 3.2 | 0.9 | 31 | 10.1 | 3.8 | 0.7 | 29 | 11.0 | 3.3 | 1.0 | 31 | 8.8 | 4.0 | 0.9 | 31 | |
| 10.5 | 4.6 | 0.8 | 41 | 10.4 | 2.7 | 0.9 | 32 | 9.9 | 4.8 | 0.6 | 33 | 11.8 | 3.0 | 1.2 | 30 | 9.5 | 4.8 | 0.8 | 30 | ||
| Среднее значение | 9.7 | 5.0 | 0.8 | 36 | 10.0 | 3.8 | 0.9 | 31 | 9.7 | 4.4 | 0.8 | 33 | 10.6 | 4.1 | 1.0 | 32 | 9.3 | 4.9 | 0.8 | 32 | |
ОБСУЖДЕНИЕ
Адаптация к водной среде у морских птиц и полуводных млекопитающих характеризуется минимально возможными размерами наружного слухового отверстия, сужением и удлинением канала слухового прохода наружного уха (Ильичев 1962; Анисимов, Ильичев, 1980; Солнцева, 2006 и др.).
Адаптация к водной среде у чистиковых птиц затрагивает в первую очередь наружное слуховое отверстие, размеры которого, вместе с размерами аптериальной поверхности, немного меньше размеров слухового отверстия наружного уха пингвинов (Анисимов, 1986). Слуховой проход сразу за слуховым отверстием начинается расширенным каналом (рис. 2B). Площадь поперечного сечения слухового прохода превышает площадь проекции контура наружного слухового отверстия в 4.5–4.8 раза (табл. 1). Для сравнения, у пингвинов площадь поперечного сечения слухового прохода на выходе в предтимпанальную полость только в 0.3 раза превышает размер площади наружного слухового отверстия (Анисимов, Ильичев, 1980).
Наличие подкожной расширенной полости в канале слухового прохода и в предтимпанальной полости, по всей видимости, может указывать на то, что у чистиковых птиц существует резонансное усиление звукового давления на барабанную перепонку в наружном ухе (Красильников, 1954; Ильичев, Черный, 1973).
Аптериальная поверхность рядом с отверстием наружного уха небольшая (рис. 2A). Она впервые подробно описана на примере чистиковых, и только отмечена у гагар и пингвинов (Анисимов, 1986). Аптериальная поверхность, ее существование рядом с отверстием наружного уха и функциональные возможности данных структур у других групп птиц остаются неизученными. Известно, что у пингвинов и гагар, так же как у чистиковых, под перьями ушной птерилии, над аптериальной поверхностью и слуховым отверстием наружного уха формируется воздушный свод.
Ушные перья укороченные, имеют нитевидные, не сцепленные наружными участками бородки. На таких бородках опахала пера расположены лучи ворсистой структуры, до 0.02 мм длиной (рис. 3Е). Если птицы, отстрелянные исследователями на морском побережье, падали в море, оперение этих птиц в течение 30–40 мин (до момента извлечения из воды) не намокало, включая и перья на голове. У чистиков имеющие ворсистую структуру концы перьев удерживают частички воздуха и, как волосяной покров околоводных млекопитающих, могут обладать водоотталкивающими и звукоэкранирующими свойствами (Анисимов, 1986; Солнцева, 2006).
У чистиковых из родов Fratercula и Uria ушные перья по размерам отличаются от контурных перьев рода Cepphus. У ипатки и толстоклювой кайры опахало контурных перьев длиннее и шире, а очины короче, по сравнению с размерными характеристиками перьев ушной птерилии (рис. 4). Угол наклона бородок ушных перьев к стержню пера в средней части опахала у ипатки и толстоклювой кайры составляет в среднем 18°–22°. Угол между бородками и стержнем контурного пера 36°–45° (табл. 2–3).
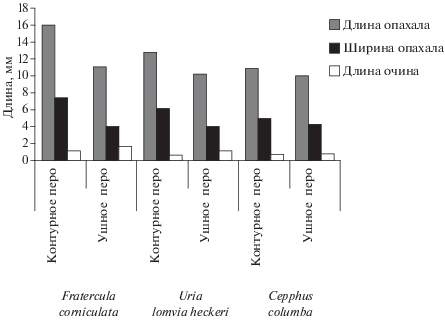
По угловым параметрам и форме очина пера (табл. 2–3) можно ожидать, что ушные перья ипатки и толстоклювой кайры обладают повышенным сопротивлением на изгиб, который возникает при давлении воды на контурные перья и ушную птерилию.
У представителя рода Cepphus ушные и контурные перья по своим метрическим параметрам слабо дифференцированы (рис. 3C–3D). У этого рода чистиковых контурные и ушные перья близки по показателю наклона бородки опахала к стержню пера (табл. 4) и обладают меньшей упругостью на изгиб. По метрическим параметрам перья ушной птерилии ипатки и толстоклювой кайры более отчетливо, чем у тихоокеанского чистика, отражают повышенную адаптацию к водной среде (рис. 4).
Плотное расположение передних ушных и верхних ушных перьев по краю слухового отверстия (0.40–0.60 мм), с просветом между перьями до 0.2 мм, исключает возможность смыкания стенок наружного слухового отверстия. Широкий слуховой проход и крутой (40°–55°) наклон его продольной оси к наружному слуховому отверстию создают топографическую совместимость наружного слухового отверстия и крупного отверстия слухового прохода в предтимпанальную полость. Их взаимное расположение совпадает на 1/3. По этим и другим морфометрическим данным, у живых чистиковых птиц исключается возможность смыкания стенок слухового прохода, т.е. существует механизм защиты барабанной перепонки от воды. Ильичев (1962, 1966, 1972), учитывая топографическую несовместимость указанных выше отверстий в слуховом проходе наружного уха, представил предположительный механизм защитного перекрывания слухового прохода наружного уха у чистиковых птиц.
В замкнутом слуховом проходе со стороны предтимпанальной полости наружного уха сохраняется воздух. Под водой у живых птиц из воздуха над слуховым отверстием и в слуховом проходе может образовываться единая воздушная “пробка–пузырь”. До настоящего времени неизвестно, может ли сохраняться под водой воздушная “пробка” в наружном ухе у чистиковых птиц и каковы функции этой пробки.
У гагар, чистиковых и пингвинов в полостях барабанной полости среднего уха расположены венозные синусы и кавернозные сплетения, которые могут увеличиваться в размере и набухать от притока крови, в результате чего повышается давление воздуха на барабанную перепонку со стороны барабанной полости среднего уха (Анисимов, Ильичев, 1980; Анисимов, 1986). Этот факт необходимо учитывать при рассмотрении возможных вариантов защитной функции воздушной “пробки–пузыря” в слуховом проходе наружного уха морских птиц.
Таким образом, результаты строения и сравнительной морфометрии наружного уха и “ушной” птерилии трех видов чистиковых птиц рассматриваются нами не только как вариант водозащитной адаптации, но и как высокая степень специализации к сохранению чувствительности их слуховой системы.
ЗАКЛЮЧЕНИЕ
Сравнительный морфометрический анализ строения наружного уха и ушной птерилии у трех видов чистиковых птиц показал, что эти структуры имеют адаптации к условиям водной среды. В наружном ухе чистиковых имеется наружное слуховое отверстие на аптериальном участке кожного покрова, но размеры этого отверстия невелики. Канал слухового прохода широкий, с большим отверстием в предтимпанальную полость с барабанной перепонкой.
Топография и размеры слухового прохода наружного уха чистиковых птиц указывают на сохранение функционального, звукового канала связи между окружающей средой и средним ухом.
В наружном ухе имеется топографическое совпадение наружного слухового отверстия и внутреннего отверстия слухового прохода в предтимпанальную полость. Исключается перекрывание слухового прохода для защиты от воды наружного уха.
Под водой, в замкнутом канале наружного уха чистиковых птиц формируется воздушная “пробка–пузырь”, способная выполнять водозащитную функцию.
Ушная птерилия вокруг наружного слухового отверстия адаптирована к водозащитной функции наружного уха. Перья формируют над наружным слуховым отверстием воздушный свод, который связан с воздухом в канале слухового прохода. Лучи на бородках опахал ушных и контурных перьев обладают водоотталкивающими свойствами и, вероятно, позволяют данному участку перьев не намокать.
Таким образом, выявлены водозащитная адаптация в строении наружного уха и сохранение канала акустической связи между внешней средой и периферическим отделом слуховой системы чистиковых птиц.
Список литературы
Анисимов В.Д., 1974. Изучение строения и функционирования звукопередающей системы среднего уха обыкновенной неясыти методом моделирования // Орнитология. Вып. 11. С. 76–87.
Анисимов В.Д., 1976. К морфологии среднего уха пингвинов // Вестник Московского Университета. № 3. С. 16–19.
Анисимов В.Д., 1978. Морфофункциональный анализ звукопередающей системы птиц. Автореф. дис. … канд. биол. наук. М.: МГУ. 24 с.
Анисимов В.Д., 1986. Структурная организация наружного отдела слуховой системы птиц, адаптированных к глубоководному плаванию // Изучение птиц СССР, их охрана и рациональное использование. Ленинград. Ч. 1. С. 33–34.
Анисимов В.Д., Ильичев В.Д., 1980. Периферия слуховой системы пингвинов // Морфологические аспекты эволюции. Отв. ред. Соколов В.Е., Лебедкина Н.С. М.: Наука. С. 130–147.
Барсова Л.И., 1973. Морфо-экологический анализ слуховых ядер продолговатого мозга сов. Автореф. дис. … канд. биол. наук. М.: МГУ. 22 с.
Барсова Л.И., 1980. Слуховые центры продолговатого мозга пингвинов // Сенсорные системы и головной мозг птиц. Ред. Ильичев В.Д., Богословская Л.С. М.: Наука. С. 165–180.
Барсова Л.И., 1984. Сравнительный анализ слуховых центров продолговатого мозга чистиковых птиц (ALCIDAE) // Зоологический журнал. Т. 63. № 3. С. 411–420.
Голубева Т.Б., Ямалова Г.В., 1980. Структурно-функциональные особенности периферического отдела слуховой системы птиц // Сенсорные системы и головной мозг птиц. Ред. Ильичев В.Д., Богословская Л.С. М.: Наука. С. 84–113.
Ильичев В.Д., 1961. Некоторые приспособления к водному образу жизни в строении наружного отдела слухового анализатора у птиц // Вестник Московского университета. Биология, почвоведение. № 1. С. 22–25.
Ильичев В.Д., 1962. Морфо-функциональный анализ наружного отдела слухового анализатора птиц. Автореф. дис. …канд. биол. наук. Киев: Академия наук Украинской ССР Институт зоологии. 17 с.
Ильичев В.Д., 1966. Биоакустика птиц. Автореф. дис. … докт. биол. наук. М.: МГУ. 22 с.
Ильичев В.Д., 1971. Экологические подходы в изучении анализаторной системы // Анализаторные системы и ориентационное поведение птиц. Под редакцией В.Д. Ильичева. М.: Изд-во Московского Университета. С. 28–31.
Ильичев В.Д., 1972. Биоакустика птиц. М.: Изд-во Московского Университета. 286 с.
Ильичев В.Д., Извекова Л.М., 1961. Некоторые особенности функционирования наружного отдела слухового анализатора птиц // Зоологический журнал. Т. 40. № 11. С. 1704–1713.
Ильичев В.Д., Черный А.Г. 1973. Характеристики слуховой системы и некоторые характеристики пространственного слуха ушастой совы // Адаптивные механизмы акустической ориентации. Под редакцией. Н.П. Наумова. М.: Изд-во Московского Университета. С. 67–140.
Карташев Н.Н., Ильичев В.Д., 1966. Некоторые особенности слухового аппарата чистиковых птиц и звуковая среда их колоний // Научные доклады высшей школы. Биологические науки. № 3. С. 36–43.
Красильников В.А., 1954. Звуковые волны. М.: Государственное издательство технико-теоретической литературы. 440 с.
Олексенко А.И., 1982. Акустическая сигнализация атлантического чистика // Экологические исследования и охрана птиц Прибалтийских республик. Каунас. С. 127–129.
Солнцева Г.Н., 2006. Орган слуха морских млекопитающих. М.: Наука. 204 с.
Штефан Б., 1977. Птерилография пингвинов // Сборник. Адаптация пингвинов. М.: Наука. С. 55–101.
Dooling R.J., Therrien S.C., 2012. Hearing in Birds: What Changes from Air to Water // Advances in Experimental Medicine and Biology. V. 730. Eds Popper A.N., Hawkins A. New York: Springer Science+Business Media, LLC. P. 77–82.
Johansen S., Larsen O.N., Christensen-Dalsgaard J., Seidelin L., Huulvej T. et al., 2016. In-Air and Underwater Hearing in the Great Cormorant (Phalacrocorax carbo sinensis) // The Effects of Noise on Aquatic Life II, Advances in Experimental Medicine and Biology. V. 875. Eds A.N. Popper, A. Hawkins. New York: Springer Science+Business Media. P. 505–512.
Kartaschew N.N., Iljitschew W.D., 1964. Das Gehororgan der Alkenvogel // Journal für Ornithologie. V. 105. P. 113–136.
Klenova A.V., Kolesnikova Y.A., 2013. Evidence for a non-gradual pattern of call development in auks (Alcidae, Charadriiformes) // Journal of Ornithology. V. 154. № 3. P. 705–716.
Wever E.G., Herman P.N., Simmons J.A., Hrtzler D.R., 1969. Hearing in the blackfooted penguin, Spheniscus demersus, as represented by the cochlear potentials // Proceedings of the National Academy of Sciences of the USA. V. 63. № 3. P. 676–680.
Дополнительные материалы отсутствуют.
Инструменты
Зоологический журнал