

Зоологический журнал, 2020, T. 99, № 9, стр. 1023-1035
Периферический отдел слуховой системы морских птиц (чистиковые – Alcidae). 2. Морфология и функциональные возможности колумеллярного комплекса среднего уха
В. Д. Анисимов *
Биологический факультет Московского государственного университета
имени М.В. Ломоносова
119991 Москва, Россия
* E-mail: flora_oa@mail.ru
Поступила в редакцию 02.10.2019
После доработки 24.12.2019
Принята к публикации 21.02.2020
Аннотация
У трех представителей семейства чистиковых (Alcidae) – ипатки, толстоклювой кайры и тихоокеанского чистика – исследованы морфометрия и функциональные возможности колумеллярного комплекса среднего уха. Описаны особенности строения барабанной перепонки, слухового столбика и колумеллярной мышцы. Строение, расположение, способы фиксации, соотношение элементов, угловые характеристики барабанной перепонки, слухового столбика у этих видов не имеют существенных различий. Размеры, строение и способ фиксации колумеллярной мышцы – musculus tensor tympani – характеризуют возможности ее участия в периодическом процессе оптимизации натяжения поверхности барабанной перепонки и в смещении слухового столбика. На стенках барабанной полости среднего уха имеются многочисленные сплетения крупных кавернозных сосудов и венозных синусов, способных увеличиваться в размерах от избытка в них крови. С учетом морфометрии и экспериментов со свежими блоками звукопередающей системы среднего уха описаны процессы движения барабанной перепонки и слухового столбика при имитации сокращения колумеллярной мышцы среднего уха. Сделано заключение о сходном механизме рефлекторного сокращения колумеллярной мышцы среднего уха у исследованных видов. Предложены схемы рычажной настройки и оптимизации движения слухового столбика с барабанной перепонкой. У чистиковых птиц морфологические водозащитные адаптации структур барабанной полости и колумеллярного комплекса среднего уха могут участвовать в процессе трансформации звука, поступающего из водной среды к слуховым рецепторам улитки внутреннего уха.
Описательная морфология слуховой системы птиц не позволяет выявить весь диапазон функциональных возможностей слухового анализатора, адаптированного к экологическим условиям среды обитания вида (Freye-Zumpfe, 1952–1953; Голубева, Ямалова, 1980; Барсова и др., 1985 и др.). Мало работ с описанием морфометрии и возможной функции колумеллярного комплекса среднего уха птиц (Ильичев, 1966; Анисимов, 1978; Анисимов, Ильичев 1980). Канал звукопередачи в среднем ухе слухового анализатора морских птиц долгое время считали не эффективным, но звуковое общение у этих птиц существует и хорошо изучено (Ильичев, 1966; Карташев, Ильичев, 1966; Олексенко, 1982; Klenova, Kolesnikova, 2013; Jouventin, Aubin, 2002 и др.).
Функция оптимизации механизма звукопередачи в среднем ухе птиц рассмотрена в разных вариантах рычажного колебания слухового столбика, но без учета в этом процессе рефлекторных сокращений колумеллярной мышцы (Schwartzkopff, 1955; Werner, 1960; Ильичев, 1966, 1972 и др.). Исследованиями функции среднего уха на живых птицах показаны сложные процессы настройки и защиты звукопередачи в колумеллярной комплексе под контролем рефлекторного сокращения колумеллярной мышцы (Анисимов, 1971, 1979; Анисимов, Ильичев, 1982, 1985).
В предлагаемой работе, на основе морфофункциональных методик (Анисимов, 1974, 1974а) представлены результаты морфометрического описания колумеллярного комплекса среднего уха трех видов чистиковых птиц. Рассматривается коррелятивная связь строения барабанной полости среднего уха, барабанной перепонки и слухового столбика с водозащитной адаптацией чистиков к водной среде. Проведен экспериментальный анализ биомеханических процессов изменения функциональной настройки колумеллярного комплекса среднего уха чистиковых при имитации возможных рефлекторных сокращений колумеллярной мышцы.
МАТЕРИАЛ И МЕТОДИКА
Морфометрический материал по чистиковым птицам собран и обработан в полевых условиях, в сезон его сбора на побережье Чукотского п-ова в районе Берингова пролива и Чукотского моря, в период с 1984 по 1987 гг. Морфометрические исследования колумеллярного комплекса среднего уха чистиков проводили на свежем материале. Изучали строение и функциональные возможности среднего уха у четырех особей каждого вида: ипатки (Fratercula corniculata (J.F. Naumann 1821)), толстоклювой кайры (Uria lomvia (Linnaeus 1758)), чукотский подвид U. l. heckeri (Portenko 1944)) и тихоокеанского чистика (Cepphus columba (Pallas 1881)).
На свежих блоках слуховой капсулы среднего уха птиц, по разработанной методике (Анисимов, 1974; Анисимов, Барсова, 2010), проводили морфологические описания с линейными, угловыми и весовыми измерениями участков и частей колумеллярного комплекса среднего уха. Колумеллярный комплекс образован барабанной перепонкой (membrana tympani), слуховым столбиком (columella auris) и тимпанальной мышцей (musculus tensor tympani–stapedius). В связи с участием мышцы в оптимизации функции звукопроведения барабанной перепонкой и слуховым столбиком (Анисимов, 1971, 1978; 1979), мы сочли правомерным тимпанальную мышцу в данной статье называть колумеллярной мышцей.
В работе использовали микроскоп МБС–2, снабженный измерительной линейкой окуляр-микрометра с точностью до 0.01 мм, рисовальный аппарат РА-1. По прорисовкам с 10-, 20- и 40-кратным увеличением на бумаге с миллиметровой сеткой измеряли площади проекций поверхности барабанной перепонки, колумеллярной мышцы, овального и круглого окон слуховой капсулы улитки внутреннего уха птицы с точностью до 0.05 мм2.
По разработанной методике (Анисимов, 1974а), в эксперименте на свежих ушных блоках капсулы среднего уха чистиковых изучали векторы смещений поверхности барабанной перепонки, деформацию участков шатра барабанной перепонки и слухового столбика под воздействием натянутого сухожилия колумеллярной мышцы. На фотоснимках, по наклеенным точечным блестящим меткам на перепонке, слуховом столбике и сухожилии мышцы, измеряли величину и определяли направление векторных смещений (Анисимов, 1974б).
Схема измерения метрических параметров барабанной перепонки и хрящевого отдела (extracolumtlla), а также костного отдела (stapes) слухового столбика чистиковых птиц представлена на рис. 1I, 1II. Размерные данные, масса и пропорциональные соотношения частей колумеллярного комплекса чистиковых даны в таблицах. Диаграммы сопоставления размеров частей слухового столбика, параметры коэффициентов усиления звукового давления на подошву столбика чистиковых и других видов птиц отражены на рисунках.
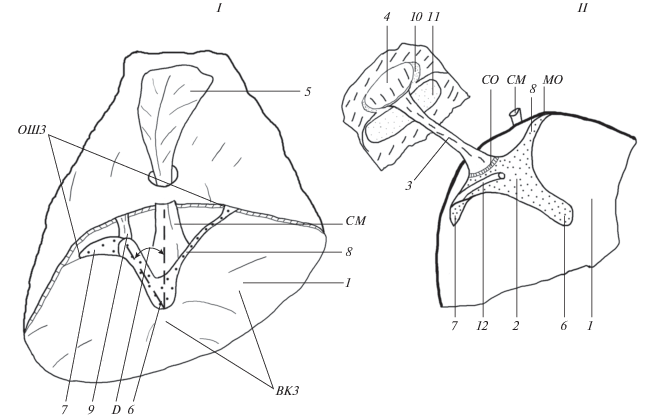
Рис. 1.
Схема строения колумеллярного комплекса среднего уха чистиковых. Прорисовка контуров с препаратов. I – барабанная перепонка со стороны слухового прохода наружного уха и расположение колумеллярной мышцы. II – слуховой столбик и его подошва, вид с фронтальной стороны столбика. 1 – membrana tympani, 2 – extracolumella, 3 – stapes, 4 – clipeolus, 5 – musculus tensor tympani (колумеллярная мышца), 6 – processus extracolumellaris, 7 – pr. supracolumellaris, 8 – pr. infacolumellaris, 9 – ligamentum acsendens, 10 – fen. ovale, 11 – fen. rotunda, 12 – lig. platneri; ОШЗ – опорно-шарнирная зона хрящевого отдела слухового столбика на барабанной перепонке, ВКЗ – вибрационно-колебательная зона свободной поверхности шатра, СМ – сухожилие мышцы, CO – синхондрозное образование, МО – место опоры pr. infacolumellaris на стенке барабанной полости среднего уха.
Представлена схема векторных смещений поверхности шатра барабанной перепонки и слухового столбика под воздействием натяжения сухожилия колумеллярной мышцы звукопередающей системы среднего уха чистиковых птиц.
Следует отметить методическую особенность сбора морфологического материала по морским птицам. Перья на голове чистиковых не намокали даже после 30–40 мин нахождения убитых птиц в воде. При фиксации структур колумеллярного комплекса среднего уха в растворе 10% формалина, на голове убитой птицы необходимо сразу удалять часть контурных и ушных перьев над слуховым отверстием наружного уха. В этом случае формалиновый фиксатор быстрее проникает в слуховой проход к барабанной перепонке.
Структуры среднего уха
Среднее ухо в периферическом отделе слуховой системы чистиковых птиц отражает морфологическую адаптацию к экологической среде обитания (Анисимов, 1986). В барабанной полости (cavum tympani) среднего уха чистиковых имеются небольшие по объему воздушные камеры. В этих камерах под водой может сохраняться воздушная “пробка”. Канал евстахиевой трубы правого и левого ушей соединяется одной узкой щелью с носоглоткой и ротовой полостью птицы. Этот канал может плотно перекрываться мускулатурой со стороны ротовой полости птицы. Описание строения среднего уха птиц дается в обзорной публикации Ильичева (1972).
Ниже в сравнительном плане дается морфометрическая характеристика колумеллярного комплекса в барабанной полости среднего уха трех видов чистиковых птиц, на примере ипатки.
Барабанная перепонка
Перепонка расположена на вентральном участке наружного края барабанной полости среднего уха. Проекция основания барабанной перепонки имеет форму овала (рис. 2). Отсутствует упругое фиброзное кольцо по краю стенки барабанной полости, на котором у птиц с чувствительным слухом крепится основание шатра барабанной перепонки (Ильичев, 1975; Анисимов, 1978).
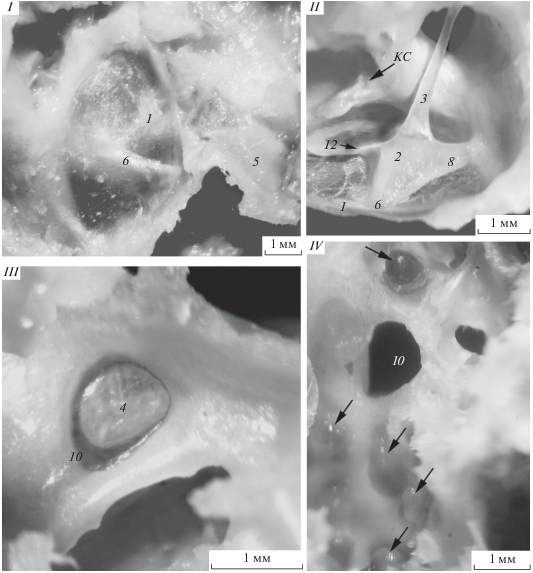
Рис. 2.
Фрагменты частей колумеллярного комплекса и участка барабанной полости среднего уха Cepphus columba (I, III, IV) и Uria lomvia (II). Макрофотографии. I – барабанная перепонка снаружи и колумеллярная мышца на внешней стенке боковой затылочной кости; II – участок слуховой столбика со стороны барабанной полости, вид сбоку; III – подошва слухового столбика в овальном окне, вид со стороны полости улитки; IV – барабанная полость среднего уха с участками вздутых от крови венозных синусов. Стрелки – места расположения венозных синусов. КС – кровеносный сосуд. Остальные обозначения как на рис. 1.
На поверхности барабанной перепонки птиц условно можно выделить две функциональные зоны (Анисимов, 1979; Анисимов, Барсова, 2010). Участок поверхности перепонки, в которую упирается и на которой фиксируется слуховой столбик (columella), сухожилие колумеллярной мышцы и восходящая связка (ligamentum ascendens), образуют участок опорно-шарнирной зоны (ОШЗ) шатра (рис. 1I, 2I).
Площадь шарнирной зоны относительно площади остальной поверхности барабанной перепонки составляет в среднем 24% у ипатки, 21% у чистика и 20% у толстоклювой кайры. Остальная часть поверхности составляет вибрационно-колебательную зону (ВКЗ) поверхности шатра барабанной перепонки. Вибрационно-колебательная зона шатра выполняет главную функцию восприятия энергии звуковой волны и передает ее на слуховой столбик (Ильичев, Черный, 1973; Анисимов, 1979 и др.).
У чистиковых птиц вибрационно-колебательная зона в основании шатра не имеет жесткой фиксации с костной стенкой барабанной полости. Отсутствует и четкая граница между наружной поверхностью шатра перепонки и кожного покрова предтимпанальной полости наружного уха (рис. 2I).
Барабанная перепонка толстая, волокнистой структуры и не эластичная на растяжение. Шатер перепонки имеет асимметричную вершину и выпячивается в предтимпанальную полость наружного уха на высоту до 0.6 мм. Поверхность наружного эктодермального слоя имеет рыхлый контакт с внутренним фиброзным слоем перепонки только по всему периметру края основания шатра. По месту фиксации барабанной перепонки с хрящевым отделом столбика, эктодермальный и фиброзный участки шатра перепонки плотно контактируют, что ограничивает подвижность одного относительно другого (Анисимов, 1978).
Со стороны наружного уха эктодермальный слой барабанной перепонки покрыт дополнительными наслоениями отмершего эпидермиса и сальных выделений. Шатер перепонки непрозрачный, что затрудняет определение точной границы по месту фиксации слухового столбика на ее поверхности. После удаления отмершего слоя эпидермиса барабанная перепонка становится полупрозрачной. В этом случае более отчетливо определяется граница фиксации хрящевого отдела слухового столбика на барабанной перепонке.
Вибрационно-колебательная зона перепонки по краю основания шатра имеет прогиб поверхности в барабанную полость до 1.0 мм. Рядом с прогибом по краю барабанной перепонки имеются морщинистые складки. Здесь, между слоями барабанной перепонки, сохраняется толстая прослойка рыхлой соединительной ткани. Такая структура основания шатра способствует смещению поверхности барабанной перепонки при сокращении колумеллярной мышцы (Анисимов, 1971, 1974б, 1978). Прогиб и фиксация поверхности барабанной перепонки отсутствуют только в опорно-шарнирной зоне шатра. В этом месте основание шатра барабанной перепонки прочно крепится к стенке барабанной полости. Слабо развитая связка – ligamentum ascendens – на опорно-шарнирной зоне шатра дополнительно закрепляет барабанную перепонку и хрящевую часть столбика на костном участке стенки барабанной полости (рис. 1I).
На внешней поверхности барабанной перепонки и по месту фиксации сухожилия мышцы на опорно-шарнирной зоне шатра отмечаются наиболее значительные (в виде концентрических валиков) наслоения и скопления отмершего эпидермиса и секреторных выделений. Аналогичные скопления сальных выделений и отмершего эпидермиса известны и для пингвинов и чаек (Анисимов, Ильичев, 1980; Анисимов, Барсова, 2010). Очертание наслоения и количество отмершего эпидермиса рядом с местом расположения сухожилия мышцы могут служить подтверждением наличия рефлекторных сокращений колумеллярной мышцы у живых чистиковых птиц.
Площадь проекции основания барабанной перепонки ипатки, толстоклювой кайры и тихоокеанского чистика дана в табл. 1.
Таблица 1.
Метрические параметры колумеллярного комплекса звукопередающей системы среднего уха чистиковых птиц
| Показатель | Номер особи, ухо | Среднее значение | |||||||
|---|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | ||||||
| правое | левое | правое | левое | правое | левое | правое | левое | ||
| Ипатка (Fratercula corniculata) | |||||||||
| Площадь барабанной перепонки, мм2 | 15.1 | 14.8 | 15.3 | 14.9 | 14.7 | 15.1 | 13.2 | 13.8 | 14.61 |
| Площадь подошвы слухового столбика, мм2 | 0.82 | 0.81 | 0.87 | 0.86 | 0.84 | 0.87 | 0.70 | 0.77 | 0.81 |
| Площадь овального окна в слуховой капсуле улитки, мм2 | 1.65 | 1.67 | 1.63 | 1.52 | 1.42 | 1.36 | 1.36 | 1.63 | 1.53 |
| Площадь круглого окна в слуховой капсуле улитки, мм2 | 2.10 | 2.16 | 2.03 | 1.98 | 1.82 | 2.05 | 1.92 | 2.03 | 2.01 |
| Соотношение площади круглого окна и площади овального окна | 1.27 | 1.29 | 1.24 | 1.30 | 1.28 | 1.50 | 1.41 | 1.24 | 1.31 |
| Площадь поверхности колумеллярной мышцы на боковой затылочной кости, мм2 | 4.9 | 4.7 | 6.4 | 6.1 | 6.3 | 6.8 | 4.8 | 4.5 | 5.56 |
| Длина слухового столбика, мм | 4.55 | 4.60 | 4.53 | 4.10 | 4.85 | 4.38 | 4.35 | 4.33 | 4.46 |
| Толстоклювая кайра (Uria lomvia heckeri) | |||||||||
| Площадь барабанной перепонки, мм2 | 15.8 | 15.2 | 14.8 | 14.3 | 16.8 | 16.2 | 13.,9 | 14.2 | 15.27 |
| Площадь подошвы слухового столбика, мм2 | 0.86 | 0.81 | 0.87 | 0.84 | 0.93 | 0.88 | 0.82 | 0.80 | 0.85 |
| Площадь овального окна в слуховой капсуле улитки, мм2 | 1.60 | 1.58 | 1.62 | 1.65 | 1.62 | 1.40 | 1.50 | 1.55 | 1.56 |
| Площадь круглого окна в слуховой капсуле улитки, мм2 | 2.30 | 2.10 | 2.30 | 2.40 | 2.40 | 2.04 | 2.06 | 1.96 | 2.19 |
| Соотношение площади круглого окна и площади овального окна | 1.43 | 1.32 | 1.41 | 1.45 | 1.48 | 1.45 | 1.37 | 1.26 | 1.39 |
| Площадь поверхности колумеллярной мышцы на боковой затылочной кости, мм2 | 8.3 | 7.9 | 7.4 | 7.6 | 8.2 | 8.4 | 6.7 | 7.3 | 7.72 |
| Длина слухового столбика, мм | 5.15 | 5.20 | 4.95 | 5.10 | 5.00 | 4.90 | 4.73 | 4.90 | 4.99 |
| Чистик (Cepphus columba) | |||||||||
| Площадь барабанной перепонки, мм2 | 11.9 | 11.8 | 12.7 | 12.5 | 12,8 | 12.3 | 11.8 | 12.0 | 12.22 |
| Площадь подошвы слухового столбика, мм2 | 0.66 | 0.64 | 0.78 | 0.77 | 0.68 | 0.66 | 0.67 | 0.69 | 0.69 |
| Площадь овального окна в слуховой капсуле улитки, мм2 | 1.10 | 1.20 | 1.19 | 1.21 | 1.26 | 1.22 | 1.20 | 1.25 | 1.20 |
| Площадь круглого окна в слуховой капсуле улитки, мм2 | 1.60 | 1.66 | 1.60 | 1.70 | 1.92 | 1.90 | 1.69 | 1.76 | 1.72 |
| Соотношение площади круглого окна и площади овального окна | 1.45 | 1.38 | 1.34 | 1.40 | 1.52 | 1.55 | 1.40 | 1.41 | 1.43 |
| Площадь поверхности колумеллярной мышцы на боковой затылочной кости, мм2 | 4.8 | 5.5 | 4.9 | 4.8 | 4.5 | 3.8 | 3.8 | 3.9 | 4.50 |
| Длина слухового столбика, мм | 3.05 | 2.90 | 3.00 | 3.13 | 2.90 | 3.00 | 3.12 | 3.20 | 3.03 |
Колумеллярная мышца
Долгое время функция тимпанальной мышцы колумеллярного комплекса в среднем ухе птиц оставалась недостаточно изученной (Breuer, 1907; Wada, 1924 и др.). В дальнейшем экспериментами было доказано, что рефлекторное сокращение мышцы происходит в ответ на звуковой сигнал возрастающей интенсивности (Анисимов, 1971; Голубева, 1972; Анисимов, Ильичев, 1982, 1985).
Колумеллярная мышца чистиковых птиц расположена на наружной стороне боковой затылочной кости черепа заднего дивертикула костной капсулы среднего уха (рис. 2I). Небольшая колумеллярная мышца под волокнистой тканью прикрыта сверху толстым слоем мускулатуры затылочной области головы птицы. Аналогичное расположение и крепление колумеллярной мышцы отмечено нами у сов, врановых, пингвинов и чайковых птиц (Анисимов, 1978; Анисимов, Ильичев, 1980а, 1982; Анисимов, Барсова, 2010). У всех чистиковых птиц эта мышца перистая по структуре, с короткими волокнами, расположенными к апоневрозу под углом до 45°.
К колумеллярной мышце, со стороны наружной поверхности затылочной кости, подходят и соприкасаются с ней две тонкие веточки нервных окончаний. Одна веточка нервного окончания отходит от лицевого нерва (n. fascialis), другая веточка, возможно, – от тройничного нерва (n. trigeminus). Место прохождения второй веточки нерва и возможность его контакта с тройничным нервом нами не изучены.
Апоневроз мышцы переходит в сухожилие, которое через отверстие в крыльях боковой затылочной кости проникает в предтимпанальную полость наружного уха. Часть сухожильных волокон крепится на опорно-шарнирной зоне шатра (рис. 1I). Одна порция волокон сухожилия крепится к фиброзному слою перепонки, другая – на центральном отростке (pr. exstracolumellaris) и средней части нижнего бокового отростка (pr. infracolumellaris) хрящевого слухового столбика. Ширина сухожилия достигает у ипатки и кайры 0.45 мм, у чистика до 0.35 мм. Продольная ось сухожилия мышцы расположена под углом 30°–35° к плечу центрального отростка на поверхности опорно-шарнирной зоне шатра (рис. 1, угол D). Площадь фиксации мышцы на боковой затылочной кости относительно площади поверхности барабанной перепонки в среднем составляет 50% у ипатки, 38 у кайры и 37% у чистика. Масса слухового столбика и колумеллярной мышцы приведена в табл. 2. Топография взаимного расположения барабанной перепонки и колумеллярной мышцы чистиковых птиц представлена на рис. 2I.
Таблица 2.
Масса слухового столбика и колумеллярной мышцы среднего уха чистиковых птиц
| Масса | Номер особи, ухо | Среднее значение | |||||||
|---|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | ||||||
| правое | левое | правое | левое | правое | левое | правое | левое | ||
| Ипатка (Fratercula corniculata) | |||||||||
| Слухового столбика с участком барабанной перепонки, мг | 1.50 | 1.40 | 1.50 | 1.60 | 1.62 | 1.64 | 1.40 | 1.35 | 1.50 |
| Костной части столбика, мг | 0.50 | 0.50 | 0.55 | 0.50 | 0.50 | 0.40 | 0.50 | 0.45 | 0.48 |
| Колумеллярной мышцы, мг | 4.9 | 4.7 | 6.4 | 6.1 | 6.3 | 6.8 | 4.8 | 4.5 | 5.56 |
| Толстоклювая кайра (Uria lomvia heckeri) | |||||||||
| Слухового столбика с участком барабанной перепонки, мг | 1.30 | 1.40 | 1.40 | 1.30 | 1.30 | 1.30 | 1.45 | 1.40 | 1.35 |
| Костной части столбика, мг | 0.50 | 0.50 | 0.40 | 0.50 | 0.50 | 0.40 | 0.50 | 0.50 | 0.47 |
| Колумеллярной мышцы, мг | 8.3 | 7.9 | 7.4 | 7.6 | 8.2 | 8.4 | 6.7 | 7.3 | 7.72 |
| Чистик (Cepphus columba) | |||||||||
| Слухового столбика с участком барабанной перепонки, мг | 0.90 | 0.70 | 0.80 | 0.80 | 0.65 | 0.50 | 0.90 | 0.90 | 0.77 |
| Костной части столбика, мг | 0.40 | 0.30 | 0.30 | 0.30 | 0.25 | 0.30 | 0.45 | 0.45 | 0.34 |
| Колумеллярной мышцы, мг | 4.8 | 5.5 | 4.9 | 4.8 | 4.5 | 3.8 | 3.8 | 3.9 | 4.50 |
Слуховой столбик
Слуховой столбик состоит из хрящевого и костного отделов (рис. 2II). Их форма и способ фиксации на барабанной перепонке у разных видов чистиковых птиц не были изучены. Хрящевой и костный отделы слухового столбика крепятся встык синхондрозным образованием (рис. 1II). Толщина костного участка столбика в средней части наименьшая и составляет 0.30–0.35 мм. Существует заметный перегиб продольной оси слухового столбика по месту сближения костного участка столбика с проходящей рядом крупной артерией на стенке барабанной полости среднего уха (рис. 2II).
Центральный отросток образует продольную ось хрящевого отдела и опору столбика на барабанной перепонке (рис. 2II). Этот отросток – самый крупный в хрящевом отделе столбика. Рельеф и структура центрального отростка формируют вершину шатра барабанной перепонки. Угол вершины шатра у чистиковых, расположенный в плоскости сечения продольной оси слухового столбика, равен 125°–130° (рис. 3I, угол A). Наклон костного отдела столбика к плечу фронтальной поверхности шатра достигает в среднем 104° у ипатки, 94° у кайры и 97° у чистика (рис. 3I, угол B).
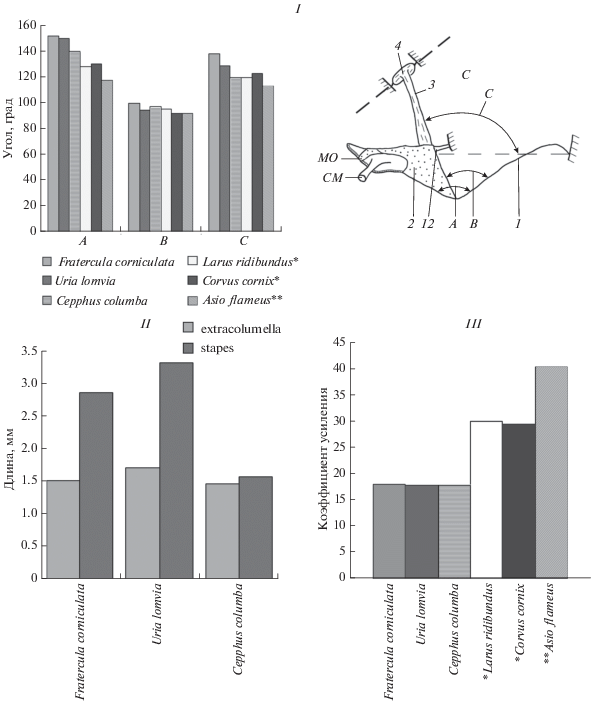
Рис. 3.
Сравнительные размеры и схема расположения углов в колумеллярном комплексе среднего уха птиц. I – углы между барабанной перепонкой и слуховым столбиком птиц. А – угол вершины шатра барабанной перепонки, B – угол наклона слухового столбика к фронтальной поверхности барабанной перепонки, C – угол наклона слухового столбика к плоскости основания шатра барабанной перепонки. Остальные обозначения как на рис. 1. II – диаграмма размеров длины хрящевой и костной части слухового столбика чистиковых птиц; 2 – extracolumella, 3 – stapes. III – коэффициент усиления звукового давления на подошву слухового столбика у разных видов птиц. * по: Анисимов, Барсова, 2010; ** по: Анисимов, Ильичев, 1980а.
Верхний боковой отросток (pr. supracolumellaris) – самый короткий – и формирует верхнюю часть базального участка хрящевого отдела столбика. Дистальный конец этого отростка в виде неширокой пластинки крепится только на барабанной перепонке в 0.3–0.4 мм от края шатра.
Нижний боковой отросток (pr. infracolumellaris) сужается в сторону своего дистального конца и упирается в стенку барабанной полости капсулы среднего уха (рис. 1). Нижний боковой отросток экстраколумеллы имеет постоянную фиксацию с фиброзным слоем перепонки и с волокнами сухожилия колумеллярной мышцы.
Платнерова связка (lig. platneri) отходит от верхнего бокового отростка в сторону отростка квадратной кости на противоположной стенке барабанной полости. У чистиковых птиц по связке, в том же направлении, тянется тонкая веточка нерва – chorda tympani (рис. 2II).
Костный отдел слухового столбика без воздушных полостей, с расширением по месту синхондроза и в области подошвы (clipiolus). У ипатки и кайры хрящевой отдел столбика всегда меньше костного, тогда как у чистика эти два отдела слухового столбика по длине выровнены (рис. 3II).
Костная часть столбика подошвой крепится в овальном окне (fen. ovale) улитки кольцевой связкой (lig. annulare) (рис. 2III). У чистиковых птиц кольцевая связка имеет разный просвет между подошвой и стенками овального окна. Плоскость подошвы столбика расположена под углом 70°–75° к продольной оси костной части столбика. Площадь поверхности барабанной перепонки превышает площадь подошвы столбика почти в 18 раз (рис. 3III).
Со стороны барабанной полости подошву к овальному окну прочно связывает дополнительная, тонкая волокнистая мембрана. Неравномерная ширина кольцевой связки между подошвой и стенкой овального окна характерна для всех исследованных видов чистиковых птиц. Площадь круглого окна (fen. rotunda) улитки превышает площадь поверхности овального окна в 1.30–1.4 раза (табл. 1). Подошва со стороны полости улитки плоская и имеет слабовыраженную вогнутую поверхность в виде присоски.
Костная часть столбика приближена к стенке барабанной полости. В месте сближения костной части столбика со стенкой барабанной полости расположена крупная кровеносная артерия (рис. 2II). Отмечены особи, у которых костная часть столбика плотно соприкасается и удерживается эластичной выстилкой на стенке барабанной полости по месту прохождения кровеносного сосуда.
На стенках барабанной полости среднего уха расположены многочисленные крупные кавернозные сплетения и венозные синусы. Нами было отмечено, что у чистиковых птиц, отстрелянных при подлете со стороны моря, кровеносные сплетения и венозные синусы на стенках барабанной полости были вздутыми и набухшими от избытка крови. Места вздутых венозных синусов, расположенных на стенке барабанной полости вблизи овального окна улитки, отмечены стрéлками (рис. 2IV). У таких птиц происходит выпячивание поверхности фронтальной части барабанной перепонки в сторону предтимпанальной полости наружного уха. Сухожилие тимпанальной мышцы тоже натянуто, а боковые отростки хрящевой части слухового столбика соприкасаются со стенкой барабанной полости. При этом и подошва столбика вместе с кольцевой связкой выпячиваются в барабанную полость среднего уха. Таких изменений не наблюдается у чистиковых с не вздутыми венозными синусами и кавернозными сплетениями.
Площадь проекции подошвы, размеры овального и круглого окна улитки чистиковых птиц, а также отношение площади круглого окна к площади овального окна даны в табл. 1.
Изменение динамического состояния колумеллярного комплекса под воздействием колумеллярной мышцы
Опыты с воздействием звуковых сигналов на слуховую систему птиц показали функциональную связь между рефлекторными сокращениями колумеллярной мышцы и процессом восприятия звука слуховыми рецепторами внутреннего уха (Голубева, 1972; Анисимов, 1974а; Голубева, Ильичев, 1977).
В биотопах горных рек Алтая и морского побережья Чукотки, где гнездятся околоводные и морские птицы, шумовой фон постоянно колеблется в пределах 50–90 дБ (Анисимов, Барсова, 2009). В эксперименте, когда на живых птиц воздействовали звуковым давлением в переменном шумовом фоне, происходило и изменение порога начального рефлекторного сокращения колумеллярной мышцы (Анисимов, Ильичев, 1982). Оптимизация функции звукопроведения в колумеллярном комплексе среднего уха птиц при нарастании интенсивности звукового стимула, в условиях переменного шумового фона, происходит под контролем рефлекторного сокращения мышцы (Анисимов, Ильичев, 1985).
В эксперименте на свежих блоках капсулы среднего уха трех видов чистиковых птиц исследован механизм смещения поверхности барабанной перепонки и слухового столбика под воздействием натяжения сухожилия колумеллярной мышцы. По движению блестящих точечных меток на перепонке и слуховом столбике прослежены направления смещений участков барабанной перепонки и слухового столбика (рис. 4I, 4II).
Рис. 4.
Схема направлений движения поверхности барабанной перепонки и слухового столбика при натяжении сухожилия колумеллярной мышцы у чистиковых птиц. I – движение поверхности участков барабанной перепонки и сухожилия колумеллярной мышцы, II – движение барабанной перепонки, хрящевого и костного отдела слухового столбика. Стрелки – направление движения участков барабанной перепонки и слухового столбика. Остальные обозначения как на рис. 1.
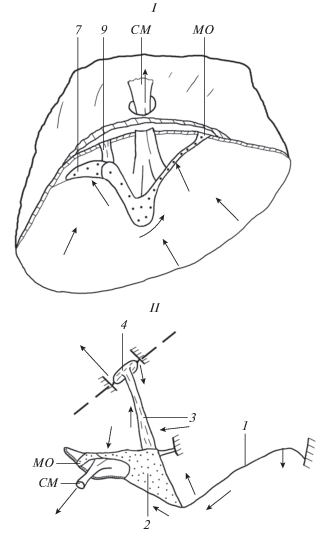
Нижний боковой отросток экстраколумеллы до натяжения сухожилия мышцы всегда упирается в стенку барабанной полости, а верхний боковой отросток крепится только на поверхности шатра и не контактирует со стенкой барабанной полости (рис. 4MO). При максимально натянутом сухожилии мышцы оба боковых отростка экстраколумеллы упираются в стенку барабанной полости. Величина смещения верхнего бокового отростка в этом случае достигает 0.35 мм. Восходящая сухожильная связка между хрящевым отделом столбика и стенкой барабанной полости в этом случае расслабляется (рис. 4I).
Натяжение и смещение перепонки с блестящими метками происходит в сторону натянутого сухожилия мышцы (рис.4I). Высота шатра барабанной перепонки при этом снижается. Прогиб поверхности шатра в барабанную полость и рыхлая прослойка соединительной ткани между слоями перепонки дает возможность осуществляться таким смещениям. Смещение поверхности перепонки происходит к вершине шатра и в направлении предтимпанальной полости наружного уха на 0.75–0.92 мм (рис. 4II).
Постепенное увеличение натяжения сухожилия мышцы создает вращательный поворот вершины столбика. Одновременно происходит поперечный изгиб нижнего бокового отростка по месту его фиксации с натянутым сухожилием мышцы. Опора отростков хрящевого отдела слухового столбика на стенке барабанной полости блокирует поступательные и поперечные смещения костного отдела столбика. При слабо натянутом сухожилии мышца подошвы слухового столбика в овальном окне улитки совершает рычажно–колебательное смещение.
Величина рычажного движения подошвы в полость улитки может достигать 0.1 мм. Движение продолжается до появления плотного контакта столбика со стенкой барабанной полости и происходит с большей амплитудой именно в области широкого участка кольцевой связки. Вектор таких колебательных движений совпадает с направлением продольной оси полости улитки.
Максимальное натяжение сухожилия блокирует поступательное смещение слухового столбика. В этом случае столбик не только хрящевым, но и костным участком начинает надавливать на стенку барабанной полости. Дальнейшее, поступательное смещение столбика в овальном окне блокируется вращательным поворотом подошвы и прекращается.
Выявленные в эксперименте изменения величины колебательных движений подошвы столбика возможны только при условии функционирования колумеллярной мышцы. Ограниченная величина колебательных движений слухового столбика в овальном окне связана с малыми размерами круглого окна, что характеризуется и высоким входным сопротивлением жидкости улитки.
Процесс расслабления сухожилия мышцы сопровождается возвратом в естественное первоначальное состояние поверхности шатра барабанной перепонки и слухового столбика в овальном окне.
В целом, эксперимент указывает на вероятность воздействия рефлекторного сокращения мышцы на изменение взаимного положения элементов колумеллярного комплекса среднего уха. В этом случае может включаться и рычажный механизм защиты звукопередающей системы при появлении избыточного внешнего давления на барабанную перепонку и подошву столбика. Такой процесс функционирования колумеллярного комплекса среднего уха отмечен нами у живых сов и врановых, на свежих колумеллярных комплексах костной капсулы среднего уха пингвинов и, возможно, существует у живых морских птиц (Анисимов, 1971, 1974б, 1979).
Сравнительный морфометрический анализ среднего уха
У исследованных видов чистиковых птиц в структурах среднего уха в колумеллярном комплексе среднего уха сохраняется звукопроводящий канал (Анисимов, 1974б, 1986).
Водозащитная адаптация в строении среднего уха проявляется в уменьшении размеров барабанной перепонки, повышении плотности и жесткости поверхности наружного слоя с уплотненной эктодермой. На поверхности перепонки расширяется опорно-шарнирная зона (становится равной 20–24%), при небольших размерах всей остальной, воспринимающей энергию звуковой волны, поверхности перепонки (Анисимов, 1986). Барабанная перепонка чистиковых по сравнению с этой структурой птиц, обладающих развитыми звуковым общением и слухом, имеет наименьший коэффициент усиления для поршневой передачи звукового давления на подошву слухового столбика и высокое ответное входное сопротивление жидкости в полости улитки (рис. 3III).
Водозащитные адаптации, связанные с уменьшением размеров барабанной перепонки, с дополнительными связками между слуховым столбиком и стенкой барабанной полости среднего уха в колумеллярном комплексе чистиковых, могут влиять на ослабление чувствительности слуха птиц к звукам низкой и средней интенсивности (Ильичев, 1972, 1975).
Весь колумеллярный комплекс чистиковых птиц характеризуется снижением амплитудных колебаний небольшой барабанной перепонки, ограниченной подвижностью поперечных смещений экстраколлумелы и костного отдела столбика в овальном окне (рис. 2II).
Одним из основных показателей поршневого усиления колебания слухового столбика в овальном окне улитки чистиковых птиц может служить наклон фронтальной поверхности перепонки к продольной оси слухового столбика. Наклон столбика к фронтальной поверхности составляет у кайры 94°, чистика 97°, ипатки 104°. Оптимальный угол передачи звукового давления с фронтальной поверхности шатра барабанной перепонки на слуховой столбик отмечен у сов, так, у болотной совы угол наклона равен 87° (рис. 3I).
Наклон столбика к плоскости основания шатра барабанной перепонки составляет у чистиковых 119°–138° (рис. 3I, угол C) и при движении колумеллярной мышцы происходят широкоамплитудные смещения столбика в поперечном направлении (Анисимов, 1978, 1979).
Известно, что колумеллярная мышца сов и врановых птиц может рефлекторно сокращаться при нарастании давления на барабанную перепонку не только звуковых стимулов, но и при нарастании давления воздуха на перепонку со стороны слухового прохода наружного уха (Анисимов, 1974; Claes et al., 2018). Продолжительное рефлекторное сокращение колумеллярной мышцы сохраняется и в условиях фонового шума, и при воздействии звуков высокой интенсивности на структуры колумеллярного комплекса (Анисимов, Ильичев, 1982, 1985). Возможно, избыточное давление воздуха на барабанную перепонку со стороны наружного уха передается на слуховую косточку и ее подошву, вызывая ответные защитные рефлекторные сокращения колумеллярной мышцы (Ильичев, 1975; Голубева, Ильичев, 1977).
В эксперименте с имитацией рефлекторного сокращения колумеллярной мышцы получены векторные смещения поверхности шатра перепонки с рычажным перегибом слухового столбика. Рефлекторная функция колумеллярной мышцы чистиковых птиц может указывать на то, что в среднем ухе процессы защиты и оптимизации звукопередачи находятся под контролем слуховых центров головного мозга (Голубева, 1972; Барсова, 1984).
Глубоководные, скоростные погружения и всплытия чистиковых птиц (Piatt, Nettleship, 1985; Burger, Simpson, 1986 и др.), по всей видимости, могут сопровождаться вздутием крупных кровеносных сосудов и венозных синусов на стенках барабанной полости среднего уха. Такие образования сосудистых расширений в кровеносной системе существуют в среднем ухе пингвинов родов Eudyptes и Pygoscelis (Анисимов, 1986; Анисимов, Ильичев, 1980) и морских млекопитающих (Солнцева, 2006).
В целом, у чистиковых птиц развитие морфологической водозащитной адаптации в периферическом отделе слуховой системы сохраняет регуляторную звукопроводящую функцию колумеллярного комплекса среднего уха в воздушной среде обитания.
ЗАКЛЮЧЕНИЕ
Периферический отдел слуховой системы чистиковых, непосредственно контактирующий с воздушной и водной средами, характеризуется развитием водозащитной адаптацией на уровне барабанной перепонки и в капсуле среднего уха.
Адаптивный характер строения среднего уха чистиковых птиц определяется сходной топографией барабанной перепонки, расположением и строением колумеллярной мышцы, конструктивными особенностями слухового столбика и расположением его подошвы в овальном окне внутреннего уха.
В колумеллярном комплексе среднего уха сохраняется механизм оптимизации функции барабанной перепонки и слухового столбика под контролем рефлекторных сокращений колумеллярной мышцы.
Векторная нагрузка основных смещений поверхности барабанной перепонки, при рефлекторном сокращении колумеллярной мышцы, направлена на вершину хрящевого отдела слуховой столбик и его костную подошву в овальном окне.
Функционирование колумеллярной мышцы сопровождается смещением барабанной перепонки и слухового столбика и включением механизма оптимизации передачи звукового давления от поверхности барабанной перепонки к мембране овального окна слуховой капсулы внутреннего уха.
Периферический отдел слуховой системы чистиковых, адаптированный к условиям водной среды, отражает специфику морфологической перестройки колумеллярного комплекса среднего уха морских птиц. Этот факт осложняет использование строения периферического отдела слуховой системы морских птиц в качестве критерия для филогенетических выводов.
В целом, структурно-морфологическая организация колумеллярного комплекса и барабанной полости среднего уха чистиковых (Alcidae) связана со специфическими экологическими и акустическими условиями среды их обитания.
Список литературы
Анисимов В.Д., 1971. Смещение звукопередающей системы среднего уха сов при действии тимпанальной мышцы // Анализаторные системы и ориентационное поведение птиц. М.: Изд-во Московского Университета. С. 44–46.
Анисимов В.Д., 1974. Изучение строения и функционирования звукопередающей системы среднего уха обыкновенной неясыти методом моделирования // Орнитология. М.: Изд-во Московского Университета. Вып. 11. С. 76–87.
Анисимов В.Д., 1974а. Методика биомеханического исследования среднего уха птиц // Материалы VI Всесоюзной орнитологической конференции. М.: Изд-во Московского Университета. Ч. 1. С. 63–64.
Анисимов В.Д., 1974б. Морфо-функциональные особенности среднего уха пингвинов // Материалы VI Всесоюзной орнитологической конференции. М.: Изд-во Московского Университета. Ч. 1. С. 64–67.
Анисимов В.Д., 1978. Морфофункциональный анализ звукопередающей системы птиц. Автореф. дис. … канд. биол. наук. М.: МГУ. 24 с.
Анисимов В.Д., 1979. Рычажный механизм в звукопередающей системе птиц // Доклады АН СССР. Т. 248. № 5. С. 1238–1241.
Анисимов В.Д., 1986. Структурная организация наружного отдела слуховой системы птиц, адаптированных к глубоководному плаванию // Изучение птиц СССР, их охрана и рациональное использование. Ленинград. Ч. 1. С. 33–34.
Анисимов В.Д., Барсова Л.И., 2009. Зашумленность звуковой среды биотопов птиц в природных ландшафтах России и сопредельных территорий // Бюллетень московского общества испытателей природы. Отд. биол. Т. 114. Вып. 4. С. 37–43.
Анисимов В.Д., Барсова Л.И., 2010. Морфология структур и механика звукопередачи в среднем ухе чаек рода Larus // Зоологический журнал. Т. 89. № 5. С. 598–608.
Анисимов В.Д., Ильичев В.Д., 1980. Периферия слуховой системы пингвинов // Морфологические аспекты эволюции. М.: Наука. С. 130–147.
Анисимов В.Д., Ильичев В.Д., 1980а. Звукопередающие функции среднего уха птиц. 1. Структурные адаптации специализированных систем // Зоологический журнал. Т. 59. Вып. 11. С. 1706–1717.
Анисимов В.Д., Ильичев В.Д., 1982. Звукопередающие функции среднего уха птиц. 2. Среднеушная мышца как регулятор звукопередачи // Зоологический журнал. Т. 61. Вып. 5. С. 747–754.
Анисимов В.Д., Ильичев В.Д., 1985. Звукопередающие функции среднего уха птиц. 3. Режимы динамического состояния звукопередающей системы и рефлекторная активность среднеушной мышцы // Зоологический журнал. Т. 64. Вып. 11. С. 1672–1684.
Барсова Л.И., 1984. Сравнительный анализ слуховых центров продолговатого мозга чистиковых птиц // Зоологический журнал. Т. 63. Вып. 3. С. 411–420.
Барсова Л.И., Богословская Л.С., Васильев Б.Д., 1985. Слуховые центры продолговатого мозга наземных позвоночных. М.: Наука. 198 с.
Голубева Т.Б., 1972. Рефлекторная активность тимпанальной мышцы совы Asio otus // Журнал эволюционной биохимии и физиологии. Т. 8. № 2. С. 173–181.
Голубева Т.Б., Ильичев В.Д., 1977. Сравнительная характеристика активности периферического отдела слуховой системы птиц // Сенсорные системы. Отв. ред. Гершуни Г.В. Ленинград: Наука. С. 162–177.
Голубева Т.Б., Ямалова Г.В., 1980. Структурно-функциональные особенности периферического отдела слуховой системы птиц // Сенсорные системы и головной мозг птиц. М.: Наука. С. 84–113.
Ильичев В.Д., 1966. Функциональные особенности и факторы эволюции звукопередающей системы птиц // Зоологический журнал. Т. 14. Вып. 9. С. 1421–1435.
Ильичев В.Д., 1972. Биоакустика птиц. М.: Издательство Московского Университета. 286 с.
Ильичев В.Д., 1975. Локация птиц. М.: Наука. 196 с.
Ильичев В.Д., Черный А.Г., 1973. Характеристики слуховой системы и некоторые характеристики пространственного слуха ушастой совы // Адаптивные механизмы акустической ориентации. Под редакцией Н.П. Наумова. М.: Изд-во Московского Университета. С. 67–140.
Карташев Н.Н., Ильичев В.Д., 1966. Некоторые особенности слухового аппарата чистиковых птиц и звуковая среда их колоний // Научные доклады высшей школы. Биологические науки. № 3. С. 36–43.
Олексенко А.И., 1982. Акустическая сигнализация атлантического чистика. // Сборник “Экологические исследования и охрана птиц Прибалтийских республик”. Каунас. С. 127–129.
Солнцева Г.Н., 2006. Орган слуха морских млекопитающих. М.: Наука. 204 с.
Breuer J., 1907. Über das Gehörorgan der Vögel // Sitzungsberichte. Akademie dre Wissenschaften in Wien. V. 116. № 3. S. 249–291.
Burger Alan E., Simpson Murk, 1986. Diving Depths of Atlantic Puffins and Common Murres // Auk. V. 103. № 4. P. 828–830.
Claes R., Muyshondt P.G.G., Dirckx J.J.J., Aerts P., 2018. Deformation of avian middle ear structures under static pressure loads, and potential regulation mechanisms // Zoology. V. 126. P. 128–136.
Freye-Zumpfe H., 1952–1953. Befunde in Mittelohr der Vögel. Halle-Wittenberg: Wiss. Z. Martin-Luter-Univ. H. 8. P. 445–461.
Jouventin P., Aubin T., 2002. Acoustic systems are adapted to breeding ecologies: individual recognition in nesting penguins // Animal behaviour. V. 64. P. 747–757.
Klenova A.V., Kolesnikova Y.A., 2013. Evidence for a non-gradual pattern of call development in auks (Alcidae, Charadriiformes) // Journal of Ornithology. V. 154. № 3. P. 705–716.
Piatt J., Nettleship D., 1985. Diving depths of four alcids // Auk. V. 102. № 2. P. 293–297.
Schwatzkopff J., 1955. Schallsinnesorgane, ihre Function und biologische Bedeutung bei Vogeln. Proc. XI Intern. Congr. Basel. S. 189–208.
Wada J., 1924. Bieträge zur vergleichenden Physiologie des Gehörorgans // Pflűgers Archiv. V. 202. № 1–2. S. 46–69.
Werner C., 1960. Das Gehőrorgan der Wirbeltiere und Menschen. Beispiel für eine vergleichende Morphologie der Lagebeziehungen. Leipzig: VEB Georg Thieme. 311 p.
Дополнительные материалы отсутствуют.
Инструменты
Зоологический журнал