

Зоологический журнал, 2020, T. 99, № 9, стр. 1002-1013
Хромосомная изменчивость Chironomus plumosus (Linnaeus 1758) (Diptera, Chironomidae) из прибрежной лагуны Балтийского моря (Куршский залив)
М. Ф. Маркиянова a, *, Н. А. Петрова b, **
a Институт океанологии им. П.П. Ширшова РАН
117997 Москва, Россия
b Зоологический институт РАН
199034 Санкт-Петербург, Россия
* E-mail: markiyanovamarina@gmail.com
** E-mail: chironom@zin.ru
Поступила в редакцию 14.02.2019
После доработки 22.04.2020
Принята к публикации 22.04.2020
Аннотация
Изучена хромосомная изменчивость популяции Chironomus plumosus (Linnaeus 1758) из прибрежной лагуны (Куршский залив Балтийского моря). Давленые препараты политенных хромосом из слюнных желез готовили по этил-орсеиновой методике. Кариофонд включает 19 последовательностей дисков хромосом, что превышает их среднее число в Палеарктике (12.5 ± 0.4). В кариофонде присутствуют уникальные, впервые описанные для вида последовательности: p'pluВ15, p'pluВ16, р'pluС7, р'pluD19, р'pluЕ3 и р'pluF7, последние четыре последовательности возникли в результате редких для кариофонда Сh. plumosus перицентрических инверсий. Выявлены 21 зиготическое сочетание и 19 геномных сочетаний последовательностей дисков хромосом. Показатели хромосомной изменчивости – доля гетерозиготных особей (54.43 ± 7.08%) и число гетерозиготных инверсий на одну особь (0.71 ± 0.10) – ниже, чем в центре палеарктической части ареала, что характерно для периферических популяций. Структура хромосомной изменчивости стабильна в течение всего периода исследования (1975 г., 1998–2009 гг.), что, вероятно, обусловлено благоприятными условиями жизни в заливе.
Chironomus plumosus (Linnaeus 1758) s. str. широко распространен в Голарктике (Белянина, Логинова, 1993; Шобанов, 1994, 1994а; Шилова, Шобанов, 1996; Голыгина, Кикнадзе, 2001; Большаков, 2013; Kiknadze et al., 2016), обитает в континентальных пресных водоемах различного типа и играет важную роль в их функционировании (Тодераш, 1984; Балушкина, 1987).
В Палеарктике хромосомная изменчивость изучена в более чем восьмидесяти природных популяциях Ch. plumosus (Максимова, Петрова, 1978; Шобанов, 1994, 1994а, 1994б; Шилова, Шобанов, 1996; Голыгина, Кикнадзе, 2001; Kiknadze et al., 2016). Данные о хромосомной изменчивости в популяциях Ch. plumosus, обитающих в прибрежных лагунах, единичны и ограничены двумя заливами Балтийского моря – Куршским (Максимова, 1980) и Заалер (залив Дарс-Цингст) (Шобанов, 1994б). Материал в этих исследованиях был собран локально, только на одной станции, что не позволяет выявить особенности хромосомной изменчивости в популяциях, обитающих в среде с большим разнообразием условий.
Прибрежные лагуны относят к мелководным эстуарным экосистемам (Bird, 2008), в которых при смешении пресной и соленой воды возникает градиент солености (Хлебович, 1974) и формируется уникальный тип экосистемы, которая характеризуется изменчивым гидрологическим режимом со специфическими элементами, свойствами и взаимодействиями (Столяров, 2011). В зоне транзита с материка в океан терригенный и органический осадки не только аккумулируются, но и сильно трансформируются, формируется мощный биологический барьер на пути загрязненного материкового стока (Столяров, 2011). В эстуариях формируются “физически контролируемые сообщества”, в организации которых роль факторов среды повышается, а роль межвидовых взаимодействий снижается (Столяров, 2011). Кроме этого, они испытывают сильное антропогенное воздействие, обусловленное активной хозяйственной деятельностью на их водосборной территории.
Куршский залив – крупнейшая мелководная лагуна в юго-восточной части Балтийского моря (Географический атлас …, 2002). Гидрохимический режим залива определяется мелководностью, ветровым перемешиванием, речным стоком и водообменом с отрытым морем (Гуделис, 1959; Червинскас, 1959). Куршский залив является большей частью пресноводным полузакрытым водоемом, в котором присутствует плавный соленостный градиент (Гидрометеорологические …, 1985; Dailidienė, Davulienė, 2007). Донные осадки представлены преимущественно различными илами, которые богаты органическими веществами – детритом и гумусами (Чечко, 2002). Содержание взвешенного вещества во всей водной толще в среднем составляет более 30 мг/л, в придонном слое воды этот показатель несколько выше (Чечко, 2002). Из-за активной сельскохозяйственной деятельности на водосборной территории в Куршский залив со стоком реки Неман поступают большие объемы азота и фосфора. Начиная с 1990-х годов, в заливе происходят экстремальные “цветения” микроводорослей, после которых отмечают дефицит кислорода на дне, вызванный интенсификацией деструкции мертвого органического вещества (Александров, 2010a). С 2000-х гг. в заливе стали обычными токсичные “цветения” с присутствием в воде цианотоксинов (микроцестины, анабенопептиды) в концентрациях, превышающих пределы, допустимые для вод рекреационного использования по квалификации ВОЗ (Ежова и др., 2012).
В Куршском заливе личинки Ch. plumosus являются доминирующим видом в зообентосе, их средняя численность и биомасса составляют соответственно 584 экз./м2 и 16.4 г/м2 (Рудинская, 2004). Личинки обитают в пресноводной части залива (южный и центральный районы) и части северного района, где соленость вод не превышает 2‰, а в районе Клайпедского пролива, где влияние Балтийского моря наибольшее и соленость составляет в среднем 5.1 ± 2.1‰ (Dailidienė, Davulienė, 2007), не встречаются (Маркиянова, 2015).
Многолетнее изучение хромосомной изменчивости в популяции Ch. plumosus Куршского залива, начатое в 1998 г. (Маркиянова, 2009), необходимо для понимания ее роли в адаптации к специфическим условиям мелководных эстуарных экосистем, а также для пополнения данных о видовом кариофонде. Кроме этого, подобные сведения могут служить базисом для экологического мониторинга водных экосистем.
Цель настоящей работы – изучить хромосомную изменчивость Ch. plumosus, обитающего в Куршском заливе Балтийского моря.
МАТЕРИАЛ И МЕТОДЫ
Личинки Ch. plumosus IV возраста были собраны в открытой части Куршского залива на 17 станциях с глубины 2.5–5.1 м в период 1998–2009 гг. (рис. 1). Личинок фиксировали на месте сбора смесью 96% этилового спирта и ледяной уксусной кислоты (3 : 1).
Давленые препараты политенных хромосом из слюнных желез готовили по этил-орсеиновой методике (Демин, Шобанов, 1990). Изучение кариотипов проводили под микроскопами “Labobal 4” (Carl Zeiss Jena, Германия), “Olympus СX 41” (Япония) при увеличении 400× и 1000×. Проанализировано 238 препаратов Ch. plumosus.
Установление последовательностей дисков хромосом (ПДХ) проводили по системе Максимовой (1976), усовершенствованной Шобановым (1994, 1994а) с использованием стандартных цитофотокарт (Кикнадзе и др., 1991; Голыгина, Кикнадзе, 2001). При обозначении последовательностей дисков хромосом указывали сокращенное латинское название вида (plu), прописной буквой символ плеча (A, B, C …) и номер последовательности дисков; префиксом обозначали географическое распространение последовательности дисков: p' – в Палеарктике, h' – в Голарктике (Голыгина, Кикнадзе, 2001). При картировании новых последовательностей дисков инвертированный участок плеча подчеркивали, в квадратные скобки заключали районы другого плеча, вошедшие в состав данного плеча в результате перицентрической инверсии.
Основываясь на работах Голыгиной и Кикнадзе (2001), Кикнадзе (2008), ПДХ разделили на четыре категории: основные общие последовательности –присутствуют в кариофондах всех популяций и доминируют по частоте встречаемости (p'pluA1, p'pluB1, h'pluC1, h'pluD1, p'pluE1, h'pluF1); альтернативные основные (p'pluA2, h'pluB2, h'pluC2, h'pluD2 и h'pluE2) – присутствуют в большинстве популяций с частотой ниже, чем основные; редкие последовательности – встречаются с низкой частотой и только в гетерозиготе и уникальные последовательности – присутствуют только в данной популяции и с очень низкими частотами.
В работе использовали показатели хромосомной изменчивости: состав и частоты ПДХ, их зиготические сочетания и геномные комбинации; показатели гетерозиготности: доля гетерозиготных особей (%) и число гетерозиготных инверсий на 1 особь.
Соответствие частот встречаемости зиготических сочетаний последовательностей дисков равновесию Харди-Вайнберга было проверено с помощью критерия χ2. Статистическая значимость различий частот ПДХ и зиготических сочетаний оценивали с помощью одностороннего варианта точного критерия Фишера (P). Средние значения приведены со стандартными ошибками.
Математическая обработка результатов осуществлялась с использованием пакета прикладных программ Excel.
РЕЗУЛЬТАТЫ
Вид имеет 2n = 8. Сочетание плеч хромосом: AB, CD, EF и G (thummi– комплекс) (Максимова, 1976; Keyl, 1962). Хромосомы AB, CD, EF – мета- и субметацентрические, G – телоцентрическая. За центромеру хромосомы АВ принят диск 12u, хромосомы СD – 14o, хромосомы EF – 11b, хромосомы G – диск 1a (Сигарева, 1981). В хромосоме G имеются одно ядрышко и 4 кольца Бальбиани, гомологи всегда не спарены.
В плече А обнаружено три ПДХ: h'pluА1, p'pluА2 и p'pluА3.
Последовательности h'pluА1 и p'pluА2 (Максимова, 1976; Шобанов, 1994, 1994а; Кикнадзе и др., 1991) имеют альтернативную нумерацию (Голыгина, Кикнадзе, 2001; Kiknadze et al., 2016), которая приведена после знака “равно”, и следующий порядок дисков:
h'pluА1=h'pluА2 1-2c 10-12a 13ba 4f-c 2g-d 9-4d 2h-3 12cd 13c-19 K
p'pluА2=p'pluА1 1-2c 10-12 3-2h 4d-9 2d-g 4c-f 13-19 K
Последовательность h'pluА1 доминирует по частоте встречаемости, остальные встречаются значительно реже (табл. 1). ПДХ h'pluА1и p'pluА2 представлены в популяции в гетеро- и гомозиготном состоянии, p'pluА3 обнаружена только в гетерозиготе c h'pluА1 (табл. 2).
Таблица 1.
Состав и частота встречаемости (%) последовательностей дисков хромосом (ПДХ) в популяции Ch. plumosus Куршского залива
| ПДХ | Май 1998 г. N = 104 |
Июль 2005 г. N = 39 |
Май 2007 г. N = 50 |
Май 2008 г. N = 34 |
Май 2009 г. N = 11 |
Среднее |
|---|---|---|---|---|---|---|
| А1 | 91.0 | 93.6 | 92.0 | 95.6 | 81.8 | 90.8 |
| А2 | 8.6 | 6.4 | 8.0 | 4.4 | 18.2 | 9.1 |
| А3 | 0.4 | – | – | – | – | 0.1 |
| В1 | 18.6 | 20.5 | 19.0 | 19.1 | 18.2 | 19.0 |
| В2 | 81.4 | 79.5 | 81.0 | 76.5 | 81.8 | 80.0 |
| В15 | – | – | – | 3.0 | – | 0.6 |
| В16 | – | – | – | 2.0 | – | 0.4 |
| С1 | 98.0 | 100.0 | 99.0 | 98.5 | 100.0 | 99.2 |
| С2 | 1.0 | – | 1.0 | 1.5 | – | 0.7 |
| С7 | 0.5 | – | – | – | – | 0.1 |
| D1 | 99.5 | 100.0 | 100.0 | 100.0 | 100.0 | 99.9 |
| D19 | 0.5 | – | – | – | – | 0.1 |
| E1 | 97.1 | 89.7 | 94.0 | 91.2 | 81.8 | 90.8 |
| E2 | 2.9 | 10.3 | 6.0 | 5.9 | 18.2 | 8.7 |
| E3 | – | – | – | 3.0 | – | 0.6 |
| F1 | 100.0 | 100.0 | 99.0 | 97.0 | 100.0 | 99.2 |
| F2 | – | – | 1.0 | – | – | 0.2 |
| F7 | – | – | – | 3.0 | – | 0.6 |
| G1 | 100.0 | 100.0 | 100.0 | 100.0 | 100.0 | 100.0 |
Таблица 2.
Состав и частота встречаемости (%) зиготических сочетаний последовательностей дисков хромосом в популяции Ch. plumosus Куршского залива
| Зиготические сочетания ПДХ |
Май 1998 г. |
Июль 2005 г. |
Май 2007 г. |
Май 2008 г. |
Май 2009 г. |
Среднее |
|---|---|---|---|---|---|---|
| А1.1 | 82.7 | 87.2 | 84.0 | 91.2 | 63.6 | 81.7 |
| А1.2 | 15.4 | 12.8 | 16.0 | 8.8 | 36.4 | 17.9 |
| А2.2 | 1.0 | – | – | – | – | 0.2 |
| А1.3 | 1.0 | – | – | – | – | 0.2 |
| B1.1 | 4.8 | 2.6 | 2.0 | 5.9 | – | 3.1 |
| B1.2 | 27.9 | 36.0 | 34.0 | 17.6 | 36.4 | 30.4 |
| B2.2 | 67.3 | 61.4 | 64.0 | 67.6 | 63.6 | 66.6 |
| B2.15 | – | – | – | 5.9 | – | 1.2 |
| B2.16 | – | – | – | 2.9 | – | 0.6 |
| C1.1 | 97.1 | 100.0 | 98.0 | 97.1 | 100.0 | 98.4 |
| C1.2 | 1.9 | – | 2.0 | 2.9 | – | 1.4 |
| С1.7 | 1.0 | – | – | – | – | 0.2 |
| D1.1 | 99.0 | 100.0 | 100.0 | 100.0 | 100.0 | 99.8 |
| D1.19 | 1.0 | – | – | – | – | 0.2 |
| E1.1 | 95.2 | 79.5 | 88.0 | 85.3 | 63.6 | 82.9 |
| E1.2 | 4.8 | 20.5 | 12.0 | 11.8 | 36.4 | 17.1 |
| E1.3 | – | – | – | 2.9 | – | 0.6 |
| F1.1 | 100.0 | 100.0 | 98.0 | 97.0 | 100.0 | 99.6 |
| F1.2 | – | – | 2.0 | – | – | 0.4 |
| F1.7 | – | – | – | 3.0 | – | 0.6 |
| G1.1 | 100.0 | 100.0 | 100.0 | 100.0 | 100.0 | 100.0 |
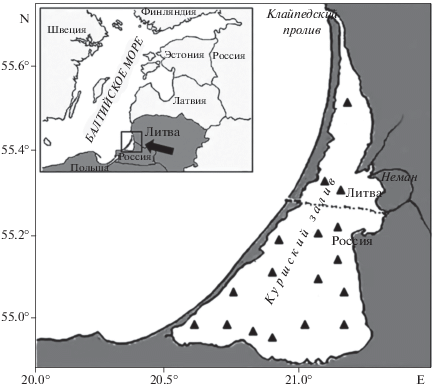
В плече В обнаружено четыре ПДХ: h'pluB1, h'pluB2, р'pluВ15 и р'pluВ16 (табл. 1). Последовательности дисков р'pluВ15, р'pluВ16 ‒ уникальные, впервые обнаружены и описаны для кариофонда вида.
Последовательность дисков р'pluВ15 возникла, возможно, на основе последовательности h'pluB2 в результате парацентрической инверсии небольшого участка хромосомы 13r‒14j в прицентромерном районе:
р'pluВ15 25‒23f 15g‒23e 15f‒14k 13r‒14j 13q‒a 12v 12u (рис. 2).
Рис. 2.
Гетерозигота h'pluB2.р'pluВ15. Здесь и далее черной линией показаны границы инверсии, стрелка указывает центромеру.

Последовательность р'pluВ16 возникла, вероятно, также на основе последовательности h'pluB2 в результате парацентрической инверсии участка хромосомы 22‒13 в прицентромерном районе:
р'pluВ16 25‒23f 15g‒21 13‒15f 23e‒22 12v12u (рис. 3).
Частота встречаемости последовательности h'pluB2 значительно превышала частоты других последовательностей дисков (табл. 1). Последовательности дисков h'pluB1, h'pluB2 обнаружены в гомо- и гетерозиготном состояниях (табл. 2). Последовательности р'pluВ15, р'pluВ16 встречаются в популяции только в гетерозиготе с h'pluB2.
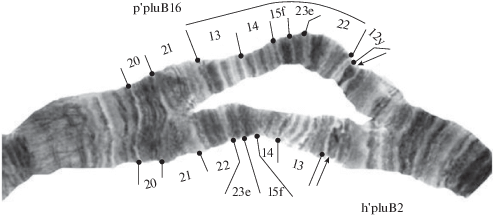
Плечо С представлено тремя последовательностями дисков: p'pluC1, p'pluC2 и р'pluС7 (табл. 1). Последовательность р'pluС7 впервые обнаружена и описана для кариофонда вида. Она возникла в результате перицентрической инверсии:
р'pluС7 25‒16 [5‒14о] (рис. 4).
Последовательность p'pluC1 преобладала в гомозиготном состоянии, p'pluC2 и р'pluС7 встречались только в гетрозиготе с p'pluC1 и с низкими частотами (табл. 1, 2).
В плече D обнаружено две последовательности дисков: p'pluD1 и р'pluD19 (табл. 1). Последовательность р'pluD19 описана для видового кариофонда впервые, возникла в результате перицентрической инверсии:
р'pluD19 1‒4 [15‒14о] (рис. 4).
Последовательность p'pluD1 доминировала в гомозиготном состоянии, р'pluD19 встречалась только в гетерозиготе с р'pluD1 с очень низкой частотой (табл. 1, 2).
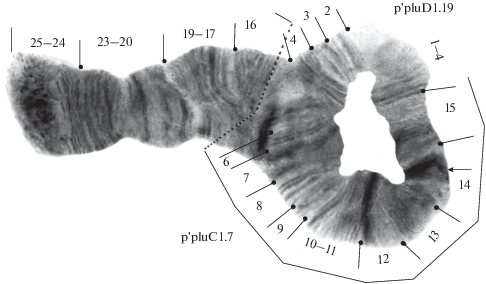
В плече Е найдено три последовательности дисков: p'pluE1, h'pluE2 и р'pluЕ3 (табл. 1). Последовательность р'pluЕ3 описана впервые, возникла в результате перицентрической инверсии:
р'pluЕ3 1‒6 [16‒11b] (рис. 5).
По частоте встречаемости доминировала последовательность p'pluE1, реже встречалась h'pluE2, а р'pluЕ3 отмечена с очень низкой частотой (табл. 1). Последовательность p'pluE1 отмечена в гомо- и гетерозиготном состояниях, последовательности h'pluE2 и р'pluЕ3 только в гетерозиготе с p’pluE1 (табл. 2).
В плече F обнаружено три последовательности: h'pluF1, р'pluF2 и р'pluF7 (табл. 1). Последовательность р'pluF7 описана впервые, возникла в результате перицентрической инверсии:
р'pluF7 22‒17 [7‒11b] (рис. 5).
По частоте встречаемости доминировала h'pluF1, последовательности р'pluF2 и р'pluF7 встречались очень редко. Последовательность h'pluF1 присутствовала в гомо- и гетерозиготном состоянии, р'pluF2 и р'pluF7 только в гетерозиготе с h'pluF1 (табл. 2).
Плечо G представлено одной последовательностью h'pluG1.
Данные о составе и частоте встречаемости зиготических сочетаний последовательностей дисков представлены в табл. 2. Всего было обнаружено 21 зиготическое сочетание. Доминировали гомозиготные основные последовательности во всех хромосомных плечах, кроме плеча В, в котором преобладала гомозиготная альтернативная последовательность h'pluB2.2.
В кариофонде изученной популяции зарегистрировано 19 геномных комбинаций (табл. 3). Среди них, с частотой 43.6%, преобладало сочетание A1.1 B2.2 C1.1 D1.1 E1.1 F1.1.
Таблица 3.
Состав и частота встречаемости (%) геномных комбинаций последовательностей дисков хромосом в популяции Ch. plumosus Куршского залива
| Геномные комбинации ПДХ | Май 1998 г. |
Июль 2005 г. |
Май 2007 г. |
Май 2008 г. |
Май 2009 г. |
Среднее | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| A | B | C | D | E | F | G | ||||||
| 1.1 | 2.2 | 1.1 | 1.1 | 1.1 | 1.1 | 1.1 | 51.0 | 46.1 | 50.0 | 50.0 | 18.2 | 43.6 |
| 1.1 | 1.2 | 1.1 | 1.1 | 1.1 | 1.1 | 1.1 | 22.1 | 23.1 | 18.0 | 11.8 | 27.2 | 20.4 |
| 1.1 | 1.1 | 1.1 | 1.1 | 1.1 | 1.1 | 1.1 | 3.8 | 2.6 | 2.0 | 5.9 | – | 2.8 |
| 1.2 | 2.2. | 1.1 | 1.1 | 1.1 | 1.1 | 1.1 | 8.6 | 2.6 | 4.0 | 5.9 | 18.2 | 7.8 |
| 1.2 | 2.2 | 1.1 | 1.1 | 1.2 | 1.1 | 1.1 | – | 5.1 | – | – | 9.1 | 2.8 |
| 1.2 | 1.2 | 1.1 | 1.1 | 1.1 | 1.1 | 1.1 | 4.7 | 5.1 | 8.0 | – | – | 3.5 |
| 2.2 | 2.2 | 1.1 | 1.1 | 1.1 | 1.1 | 1.1 | 1.0 | – | – | – | – | 0.2 |
| 1.2 | 1.1 | 1.1 | 1.1 | 1.2 | 1.1 | 1.1 | 1.0 | – | – | – | – | 0.2 |
| 1.1 | 1.2 | 1.1 | 1.1 | 1.2 | 1.1 | 1.1 | – | 7.7 | 4.0 | 2.9 | – | 2.9 |
| 1.1 | 2.2 | 1.1 | 1.1 | 1.2 | 1.1 | 1.1 | 3.8 | 7.7 | 8.0 | 5.9 | 18.2 | 8.7 |
| 1.2 | 1.2 | 1.1 | 1.1 | 1.1 | 1.2 | 1.1 | 1.0 | – | 2.0 | 2.9 | 9.1 | 3.0 |
| 1.1 | 2.2 | 1.2 | 1.1 | 1.1 | 1.1 | 1.1 | 1.0 | – | – | 2.9 | – | 0.8 |
| 1.3 | 2.2 | 1.1 | 1.1 | 1.1 | 1.1 | 1.1 | 1.0 | – | – | – | – | 0.2 |
| 1.2 | 1.1 | 1.1 | 1.1 | 1.1 | 1.1 | 1.1 | – | – | 2.0 | – | – | 0.4 |
| 1.1 | 1.2 | 1.2 | 1.1 | 1.1 | 1.1 | 1.1 | – | – | 2.0 | – | – | 0.4 |
| 1.1 | 2.15 | 1.1 | 1.1 | 1.1 | 1.1 | 1.1 | – | – | – | 5.9 | – | 1.1 |
| 1.2 | 2.16 | 1.1 | 1.1 | 1.1 | 1.1 | 1.1 | – | – | – | 2.9 | – | 0.5 |
| 1.1 | 2.2 | 1.7 | 1.19 | 1.1 | 1.1 | 1.1 | 1.0 | – | – | – | – | 0.2 |
| 1.1 | 2.2 | 1.1 | 1.1 | 1.3 | 1.7 | 1.1 | – | – | – | 3.0 | – | 0.6 |
Доля гетерозиготных особей варьировала от 44.23 до 81.82%, в среднем составила 54.43 ± 7.08% (табл. 4). Число гетерозиготных инверсий на одну особь изменялось от 0.50 до 1.09, в среднем составляло 0.71 ± 0.10 (табл. 4).
Таблица 4.
Показатели гетерозиготности в популяции Ch. plumosus Куршского залива
| Показатель гетерозиготности | Май 1998 г. N = 104 |
Июль 2005 г. N = 39 |
Май 2007 г. N = 50 |
Май 2008 г. N = 34 |
Май 2009 г. N = 11 |
Среднее | SD |
|---|---|---|---|---|---|---|---|
| Число гетерозиготных инверсий на 1 особь | 0.53 | 0.69 | 0.66 | 0.56 | 1.09 | 0.71 | 0.10 |
| Доля гетерозиготных особей, % | 44.23 | 54.00 | 48.00 | 44.12 | 81.82 | 54.43 | 7.08 |
Проверка соблюдения равновесия Харди-Вайнберга, показала, что данная популяция в период исследования была равновесна по частотам зиготических сочетаний ПДХ.
ОБСУЖДЕНИЕ
Всего за период исследования у личинок Сh. plumosus из Куршского залива было обнаружено 19 ПДХ (табл. 1), что выше среднего числа ПДХ в ареале, которое составляет 12.5 ± 0.4 (Голыгина, Кикнадзе, 2001). Последовательности p'pluВ15, p'pluВ16, р'pluС7, р'pluD19, р'pluЕ3 и р'pluF7 для кариофонда Сh. plumosus описаны впервые, найдены только в популяции Куршского залива и относятся к категории уникальных ПДХ. Последовательности р'pluС7 и р'pluD19, р'pluЕ3 и р'pluF7 возникли в результате редких для кариофонда Сh. plumosus перицентрических инверсий.
Из всех хромосомных плеч наиболее полиморфно плечо В, оно представлено четырьмя ПДХ (табл. 1). Остальные плечи, кроме мономорфного плеча G, имели две–три последовательности.
В изученной популяции со средней частотой от 80до 100% доминировали основные ПДХ во всех хромосомных плечах, кроме плеча В, в котором преобладала альтернативная последовательность h'pluВ2 (табл. 1). Альтернативные последовательности p'pluA2 и h'pluE2 встречались с гораздо более низкими средними частотами – около 10%. Средние частоты остальных ПДХ были менее 1%. По данным Голыгиной и Кикнадзе (2001), подобная структура хромосомной изменчивости, когда в плечах A, C, D, E, F, G доминируют основные ПДХ, в плече B – альтернативная h'pluВ2, а прочие ПДХ встречаются редко, отмечена в 21% изученных популяций.
За период исследования выявлено 21 зиготическое сочетание, среди них преобладали основные гомозиготы. Только в плече В на протяжении всего периода исследования отмечено устойчивое доминирование альтернативной гомозиготы h'pluВ2.2 (табл. 2).
В кариофонде популяции было обнаружено 19 различных геномных комбинаций. Чаще других встречались: А1.1 B2.2 C1.1 D1.1 E1.1 F1.1 (с частотой 43.6%) и A1.1 B1.2 C1.1 D1.1 E1.1 F1.1 (20.4%). Геномная комбинация, состоящая только из стандартных последовательностей дисков, встречалась редко, в среднем с частотой 2.8%.
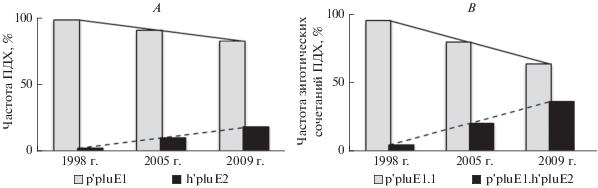
Анализ хромосомной изменчивости в популяции Сh. plumosus Куршского залива за период с 1975 (Максимова, 1980) по 2009 г. показал, что частоты основных, альтернативных ПДХ и их зиготических сочетаний в плечах A, B, C, D и F, согласно точному критерию Фишера (Р > 0.05), статистически не различаются. Только в плече Е сравнение данных 1998 г. и 2005, 2009 гг. выявлено статистически значимое (Р < 0.05) снижение частоты последовательности р'pluE1 (рис. 6А) и гомозиготы р'pluE1.1 (рис. 6В) при возрастании частоты h'pluE2 (рис. 6А) и гетерозиготы р'pluE1.h'pluE2 (рис. 6В). Подобный анализ данных для 2007 и 2008 гг. статистически значимых различий не выявил (Р > 0.05). Таким образом, за тридцатилетний период исследований структура хромосомной изменчивости популяции Сh. plumosus Куршского залива существенно не изменялась, хотя в заливе в 1990–2000 гг. происходит интенсивный прогрев воды, обусловленный ростом числа “теплых” лет (Александров, 2010). Это стало одной из основных причин развития до уровня “гиперцветения” потенциально токсичных Cyanobacteria (Aphanizomenon flos-aquae и др.), роста среднемноголетней первичной продукции в 1.5 раза и перехода водной системы в гиперэвтрофное состояние (Александров, 2010, 2010a).
Рис. 6.
Хромосомная изменчивость плеча Е и ее линейный тренд в популяции Ch. plumosus Куршского залива, приведены данные, полученные в разные годы со статистически значимыми различиями частот (Р < 0.05); А – частоты ПДХ, В – частоты зиготических сочетаний ПДХ.
В популяции Куршского залива доля гетерозиготных особей составила в среднем 54.43 ± 7.08%, число гетерозиготных инверсий на одну особь – в среднем 0.71 ± 0.10 (табл. 4). С 1998 г. по 2008 г. эти показатели варьировали незначительно (табл. 4), в 2009 г. при малочисленности выборки их значения были почти в два раза выше (табл. 4). В центре палеарктической части ареала в популяциях Сh. plumosus в 1988–1998 гг. доля гетерозиготных особей изменялась от 0 до 100% и в среднем составляла 65.85 ± 3.78, число гетерозиготных инверсий на 1 особь – от 0 до 2, в среднем – 0.99 ± ± 0.08 (Голыгина, Кикнадзе, 2001). Таким образом, в изученной популяции в 1998–2009 гг. показатели гетерозиготности имели значения ниже средних в Палеарктике, что характерно для периферических популяций (Майр, 1968).
Популяции хирономид прибрежных лагун Балтийского моря находятся на морском побережье, где их распространение ограничено соленостью воды. Такие популяции считают периферическими (Старобогатов, 1970; Левонтин, 1978) и для большинства из них характерна низкая численность и биомасса особей (Майр, 1968). Но, в прибрежных лагунах Балтийского моря численность и биомасса личинок рода Chironomus высокие, что обусловлено благоприятными условиями обитания (Рудинская, 2004; Arndt, 1994). Левонтин (1978) предложил различать периферические популяции по двум аспектам – географическому и экологическому. Если они совпадают, то такая популяция неустойчива и имеет низкую численность. В случае, когда популяция периферическая только в географическом отношении, а в экологическом – нет, она имеет высокую численность. Исходя из этого, популяции хирономид прибрежных лагун (в том числе Куршский и Заалер заливы) можно считать периферическими только в географическом аспекте.
На хромосомную изменчивость в популяциях Сh. plumosus оказывают влияние тип водоема и локальные условия обитания (Петрова и др., 1996). Хромосомная изменчивость популяций Куршского и Заалер заливов, как популяций эвтрофных мелководных прибрежных лагун, характеризуется доминированием основных ПДХ в плечах A, C, D, E, F, G и их зиготических сочетаний (рис. 7А, 7В), а также снижением показателей гетерозиготности по сравнению с популяциями из центральной части ареала в Палеарктике (рис. 8А, 8В). В Куршском и Заалер заливах, различающихся по основным характеристикам (табл. 5), соответственно различны локальные условия обитания и хромосомная изменчивость популяций Сh. plumosus (рис. 7А, 7В). Так, в популяции залива Заалер частота h'pluВ2 составляла всего 2.65% и отсутствовала гомозигота h'pluВ2.2 (Шобанов, 1994б), а в популяции Куршского залива h'pluВ2 доминировала со средней частотой 80% и соответствующая гомозигота с частотой 66.6% (табл. 1 и 2; рис. 7А, 7В). Кроме этого, в кариофонде популяции Заалер залива отсутствовали уникальные ПДХ, а в популяции Куршского залива они представлены и составляют почти треть всех ПДХ (табл. 1).
Рис. 7.
Хромосомная изменчивость в популяциях Ch. plumosus Куршского залива в 1998–2009 гг. и Заалер залива (Дарс-Цингст залив) в 1989 г. (Шобанов, 1994б; А – частоты основных и альтернативных ПДХ, В – частоты зиготических сочетаний основных и альтернативных ПДХ.
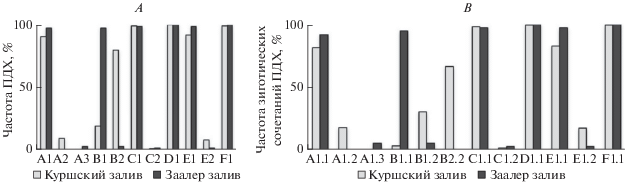
Рис. 8.
Показатели гетерозиготности Ch. plumosus и их линейные тренды: Палеарктика (центр ареала), 1988–1998 гг.рассчитано по данным Голыгиной и Кикнадзе (2001); Куршский залив, 1998–2008 гг. (в расчет не включены данные 2009 г. из-за малочисленности выборки); Заалер залив, 1989 г. (Шобанов, 1994б); А – доля гетерозиготных особей, В – число гетерозиготных инверсий на 1 особь.
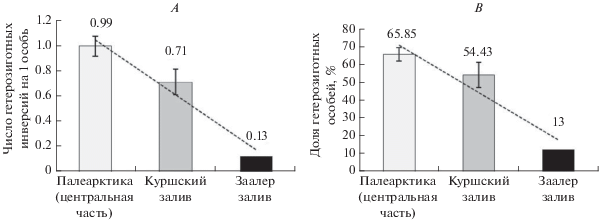
Таблица 5.
Основные характеристики Заалер и Куршского заливов
| Залив | Площадь, м2 | Глубина, м cредняя многолетняя (max) |
Соленость, ‰ средняя многолетняя (min/max) |
рН |
|---|---|---|---|---|
| Заалер (Schlungbaum et al., 1994) | 80.9 | 2.2 (4) | 3.5 (0.4/5.4) | 9.07 |
| Куршский (Гидрометеорологические…, 1985; Dailidienė, Davulienė, 2007) | 1564 | 3.8 (5.8) | 0.04–2.6 (0.01/7.7) | 8.3 |
ЗАКЛЮЧЕНИЕ
Хромосомная изменчивость популяции Ch. plumosus прибрежной лагуны, в которой присутствует градиент солености, характеризуется, с одной стороны, широким составом ПДХ (одним из самых больших среди всех изученных популяций), с другой стороны, – сниженными по сравнению со средними значениями в ареале показателями гетерозиготности. Только в данной популяции почти треть выявленных ПДХ относятся к категории уникальных и впервые описанных для вида, причем бо̀льшая их часть (четыре из шести) возникла в результате редких для кариофонда Сh. plumosus перицентрических инверсий. Структура хромосомной изменчивости и инверсионного полиморфизма стабильна в течение всего периода исследования (1975 г., 1998–2009 гг.), что, вероятно, обусловлено благоприятными условиями жизни в заливе.
Список литературы
Александров С.В., 2010. Влияние климатических изменений на уровень эвтрофирования Куршского залива // Вестник Российского государственного университета им. И. Канта. Вып. 1. С. 49–57.
Александров С.В., 2010a. Первичная продукция планктона в лагунах Балтийского моря (Вислинский и Куршский заливы). Калининград: Атлант НИРО. 228 с.
Балушкина Е.В., 1987. Функциональное значение личинок хирономид в континентальных водоемах. Л.: Наука. 179 с.
Белянина С.И., Логинова Н.В., 1993. Кадастр порядков дисков в политенных хромосомах видов Chironomus группы plumosus. II. Кариофонд Chironomus plumosus // Цитология. Т. 35. № 8. С. 65–70.
Географический атлас Калининградской области, 2002. Калининград: Изд-во КГУ. С. 116–135.
Гидрометеорологические условия шельфовой зоны морей СССР, 1985. Под ред. Теризиева Ф.С. // Т. 1. Балтийское море. Вып. 3. Куршский и Вислинский заливы. Л.: Гидрометеоиздат. 72 с.
Большаков В.В., 2013. Адаптивная роль хромосомных инверсий у личинок рода Chironomus (Diptera, Chironomidae). Автореф. дис. … канд. биол. наук. Борок. 25 с.
Голыгина В.В., Кикнадзе И.И., 2001. Кариофонд вида Chironomus plumosus (Diptera, Chironomidae) в Палеарктике // Цитология. Т. 43. № 5. С. 507–519.
Гуделис В., 1959. Геологические и физико-географические условия залива Куршю марес // Куршю Марес: итоги комплексного исследования. Вильнюс. С. 7–47.
Демин С.Ю., Шобанов Н.А., 1990. Кариотип комара Chironomus entis из группы plumosus в европейской части СССР // Цитология. Т. 32. № 10. С. 1046–1054.
Ежова Е.Е., Е.К. Ланге Е.К., Я.В. Русских Я.В, Жаковская З.А, Чернова Е.Н., 2012. Вредоносные цветения микроводорослей в Куршском заливе Балтийского моря в 2008–2011 гг. // Проблемы изучения и охраны природного и культурного наследия национального парка “Куршская коса”. Калининград: БФУ им. И. Канта. Вып. 8. С. 81–95.
Кикнадзе И.И., 2008. Роль хромосомного полиморфизма в дивергенции популяций и видов в роде Chironomus (Diptera) // Зоологический журнал. Т. 87. № 6. С. 686–701.
Кикнадзе И.И., Шилова А.И., Керкис И.Е., Шобанов Н.А., Зеленцов Н.И. и др., 1991. Кариотипы и морфология личинок трибы Chironomini. Атлас. Новосибирск: Наука. 115 с.
Левонтин Р., 1978. Генетические основы эволюции. М.: Мир. С. 157–161.
Майр Э., 1968. Зоологический вид и эволюция. Перевод с английского Гептнера В.Г., Орловой В.Н. М.: Мир. С. 310–315.
Максимова Ф.Л., 1976. К вопросу о кариотипе Chironomus plumosus L. усть-ижорской популяции Ленинградской области // Цитология. Т. 18. № 10. С. 1264–1269.
Максимова Ф.Л., 1980. Инверсионный полиморфизм природных популяций Chironomus plumosus L. Новые данные по кариосистематике двукрылых насекомых // Труды Зоологического института АН СССР. Т. 95. С. 31–39.
Максимова Ф.Л., Петрова Н.А., 1978. Географическая изменчивость кариотипа Chironomus plumosus L. // Зоологический журнал. Т. 54. № 12. С. 1816–1825.
Маркиянова М.Ф., 2009. Цитогенетическая структура популяции Chironomus plumosus (L.) (Diptera, Chironomidae) Куршского залива Балтийского моря в связи с изменением его трофического статуса // Биология внутренних вод. № 4. С. 44‒47.
Маркиянова М.Ф., 2015. Состав и распространение видов-двойников рода Chironomus Meigen, 1803 (Diptera, Chironomidae). В Куршском заливе Балтийского моря // Поволжский экологический журнал. № 4. С. 400–408.
Петрова Н.А., Ильинская Н.Б., Кайданов Л.З., 1996. Адаптивный характер инверсионного полиморфизма у мотыля Chironomus plumosus (Díptera, Chironomidae). Пространственное распределение инверсий по ареалу // Генетика. Т. 32. № 12. С. 1629–1642.
Рудинская Л.В., 2004. Макрозообентос Куршского залива // Закономерности гидрологического режима водоемов различного типа. Под ред. Алимова А.Ф., Ивановой М.Б. М.: Научный мир. С. 129–135.
Сигарева Л.Е., 1981. Дифференциальная окраска хромосом хирономид. II. C-диски политенных хромосом трех видов хирономусов // Цитология. Т. 23. № 3. С. 270–274.
Старобогатов Я.И., 1970. Фауна моллюсков и зоогеографическое районирование континентальных водоемов земного шара. Л.: Наука. С. 5–10.
Столяров А.П., 2011. Структурно-функциональная организация эстуарных экосистем Белого моря. Автореф. дис. … докт. биол. наук. М.: Московский государственный университет имени М.В. Ломоносова. 51 с.
Тодераш И.К., 1984. Функциональное значение хирономид в экосистемах водоемов Молдавии. Отв. ред. Владимиров М.З. Кишинев: Штиинца. С. 145–171.
Хлебович В.В., 1974. Критическая соленость биологических процессов. Л.: Наука. 236 с.
Червинскас Э., 1959. Основные черты гидрологического режима // Куршю Марес: итоги комплексного исследования. Вильнюс. С. 47–69.
Чечко В.А., 2002. Анализ пространственно-временной изменчивости взвешенного вещества Калининградского залива Балтийского моря // Водные ресурсы. Т. 29. № 4. С. 425–432.
Шилова А.И., Шобанов Н.А., 1996. Каталог хирономид рода Chironomus Meigan 1803 (Diptera, Chironomidae) России и бывших республик СССР // Экология, эволюция и систематика хирономид. Под ред. Шобанова Н.А., Зинченко Т.Д. Тольятти, Борок: ИБВВ и ИЭВБ РАН. С. 28–44.
Шобанов Н.А., 1994. Кариофонд Chironomus plumosus (L.) (Diptera, Chironomidae). I. Стандартизация дисков политенных хромосом в системе Максимовой // Цитология. Т. 36. № 1. С. 117–122.
Шобанов Н.А., 1994а. Кариофонд Chironomus plumosus (L.) (Diptera, Chironomidae). II. Инверсионные варианты хромосомных плеч // Цитология. Т. 36. № 1. С. 123–127.
Шобанов Н.А., 1994б. Кариофонд Chironomus plumosus (L.) (Diptera, Chironomidae). IV. Внутри- и межпопуляционный полиморфизм // Цитология. Т. 36. № 11. С. 1129–1143.
Arndt E.A., 1994. Struktur und Dynamik des Makrozoobenthos in der Darß-Zingster Boddenkette im Laufe der letzten 25 Jahre unterbesonderer Berücksichtigung der Makrozoobenthos – Entwicklungim Saaler Bodden von 1986–1990 // Rostocker Meeresbiologische Beiträge. V. 2. S. 93–120.
Bird E., 2008. Coastal geomorphology: an introduction. Second edition. England: John Willey & Sons Ltd. P. 395–331.
Dailidienė I., Davulienė L., 2007. Long-term mean salinity in the Curonian Lagoon in 1993–2005 // Acta Zoologica Lituanica. V. 17. № 2. P. 172–181.
Kiknadze I., Istomina A., Golygina V., Gunderina L., 2016. Karyotypes of Palearctic and Holarctic species of the genus Chironomus. Novosibirsk: Academic Publishing House “GEO”. 489 p.
Keyl H.-G., 1962. Chromosomenevolution bei Chironomus. II. Chromosomenumbanten und phylogenetische Beziehungen der Arten // Chromosoma. V. 13. № 4. P. 464–514.
Schlungbaum G., Baudler H., Nausch G., 1994. Die Darß-Zingster Boddenkette – ein typisches Flachwasserästuar an der südlichen Ostseeküste // Rostocker Meeresbiologische Beiträge. V. 2. S. 5–26.
Дополнительные материалы отсутствуют.
Инструменты
Зоологический журнал