

Зоологический журнал, 2020, T. 99, № 9, стр. 1062-1076
Определение популяционной плотности мелких млекопитающих путем мечения на линиях живоловок
Н. А. Щипанов *
Институт проблем экологии и эволюции имени А.Н. Северцова РАН
119071 Москва, Россия
* E-mail: shchipa@mail.ru
Поступила в редакцию 23.03.2019
После доработки 11.10.2019
Принята к публикации 19.12.2019
Аннотация
По сравнению с традиционным мониторингом мелких млекопитающих на основе отлова с изъятием, наблюдение за индивидуально мечеными животными менее инвазивно и более соответствует современному этическому стандарту. Ограничением традиционного метода мечения с повторным отловом на площадке является относительно бóльшая трудоемкость. Расстановка ловушек в линию позволяет одновременно наблюдать население нескольких участков и получать репрезентативные выборки, но для этого необходима унификация данных. Сопоставимой для любых видов оценкой обилия является абсолютная плотность популяции – число особей, живущих на 1 га. Предложен метод расчета плотности и индексов нерезидентности на основе распределения частот встречаемости особей с разным количеством поимок. На примере обыкновенной бурозубки (Sorex araneus (L. 1758)) показано соответствие расчетных и эмпирических данных, полученных на одной и той же территории. Обсуждаются индексы нерезидентности. Сделаны выводы об их пригодности для оценки многолетней динамики у отдельных видов и ограниченной применимости для межвидовых сравнений.
Изучение реакции биоты на антропогенную трансформацию ландшафтов (потерю или изменение местообитаний, их фрагментацию, загрязнение, вспышки зоонозных болезней, распространение инвазивных видов и глобальные климатические процессы) основано на продолжающихся наблюдениях – мониторинге (Spellerberg, 2005; Siddig et al., 2016). Объектами наблюдений являются особи, популяции или группы популяций и сообщества видов, состояние которых отражает состояние среды (Соколов, Ильенко, 1977; Landers et al., 1988; Bartell, 2006; Burger, 2006; Siddig et al., 2016). Основными требованиями к индикационным видам являются их адекватный масштаб и скорость реакции. Беспозвоночные животные и травянистые растения могут оказаться более чувствительными к мелкомасштабным изменениям, чем к реакции биогеоценоза в целом, а крупные виды могут реагировать с запаздыванием из-за низких темпов оборота поколений (Carignan, Villard, 2002). Мелкие млекопитающие – один из наиболее удобных объектов мониторинга (Бердюгин, Большаков, 2005; Мухачева и др., 2009; Истомин, 2014; Ильяшенко и др., 2015; Катаев, 2015; Бобрецов и др., 2017). Основными показателями качества среды их обитания являются относительное обилие вида/видов, демографическая структура популяции и/или состояние организма, включая физиологию, уровень накопления поллютантов в органах и пр. Сбор данных, в основном, производится при использовании различных способов отлова с изъятием (линии давилок, ловчие канавки, конуса, цилиндры). При этом для получения репрезентативных выборок необходимы достаточно большой объем учетов (Карасева и др., 2008) и, соответственно, неизбежное при таких учетах умерщвление большого количества зверьков (обычно несколько сотен), что не соответствует современным этическим нормам (Veatch, 2016).
Демографические характеристики, характер использования пространства и уровень нерезидентности могут быть оценены с применением индивидуального мечения зверьков – одного из наименее инвазивных способов наблюдения, не влияющего на локальную структуру популяций отдельных видов и сообщества в целом. В зависимости от задачи, из среды может быть изъято нужное (ограниченное) количество особей с установленным статусом, что позволит значительно уменьшить количество умерщвляемых особей. Однако до сих пор индивидуальное мечение, в основном, используется лишь для углубленных исследований (Карасева и др., 2008). Это связано с относительно большой трудоемкостью метода.
Трудоемкость исследований можно значительно снизить, если расположить ловушки в линию, что позволит получить репрезентативные данные при использовании 50–100 ловушек в течение 10–15 дней (Щипанов и др., 2000; Shchipanov et al., 2005; Щипанов и др., 2010 а; Калинин, 2012; Куприянова, Калинин, 2015). Однако мечение на трансектах требует унификации данных.
Для линейных учетов в живоловки были предложены способ расчета абсолютной плотности и оценка величины потока нерезидентов (Калинин, 2012). Расчетные данные соответствовали результатам, полученным при использовании учетов давилками и канавками (Калинин, 2012; Калинин, Куприянова, 2015). Вместе с тем эти расчеты довольно громоздки. В данной публикации я предлагаю более простую процедуру расчета. Валидность предлагаемого расчета была проверена при сопоставлении эмпирических результатов, полученных на площади, с расчетами для линий на той же территории, и сопоставлена с расчетами по методике Калинина (2012).
Выбор обыкновенных бурозубок (Sorex araneus (L. 1758)) в качестве объекта для проверки расчетов определяется их высокой популяционной плотностью, особенно в захламленных местообитаниях с густым травяным покровом (Churchfield, 1990; Shchipanov et al., 2019), небольшим расстоянием между крайними точками, обычно менее 30 м, и сравнительно небольшой площадью участка – 1000 м2 (Shillito, 1963; Croin Michielsen, 1966; Buckner, 1969; Churchfield, 1990; Moraleva, Telitzina, 1994; Ивантер, Макаров, 2001). Это позволяет получать хорошие выборки на относительно коротких трансектах. Реальные участки обыкновенной бурозубки разнообразны по форме и, как правило, сильно вытянуты. Поэтому сравнение эмпирической и расчетной плотности у этого вида позволяет судить о применимости метода для животных с разными конфигурациями участков.
МАТЕРИАЛ И МЕТОДЫ
Сбор данных
Мечение проходило на заброшенном участке деревни, заросшем злаками, крапивой, рудеральным разнотравьем, с отдельными старыми липами и молодым подростом ольхи и березы. На площади 0.65 га через каждые 10 м были расположены 5 параллельных линий: 4 линии по 18 ловушек длиною 135 м и одна линия, состоящая из 15 ловушек, длиною 112.5 м. Общая длина линий 652.5 м, интервал между ловушками 7.5 м. Зверьков отлавливали в специализированные ловушки (Щипанов, 1986) в соответствии с протоколом отлова (Щипанов и др., 2000; Shchipanov et al., 2005). В отличие от стандартного протокола, проверку проводили один раз в сутки, через 1.5 ч после настораживания. Остальное время суток животные перемещались свободно. При таком режиме наблюдений каждую поимку можно рассматривать как независимую, т.е. не зависящую от места предыдущей локализации особи. Ловушки настораживали в 6–7 ч утра, в период максимальной активности. Для проверки результатов расчета использованы данные за август 2014–2018 гг. Всего помечено 180 сеголеток, получено 813 поимок.
Вероятность обнаружения особи в пространстве участка
Частота встречаемости в пространстве оценена как распределение дистанций отклонений от центральной координаты регистраций (х) у каждой отдельной особи ( j):
(1)
${{х}_{j}} = {{\Sigma {{x}_{{ji}}}} \mathord{\left/ {\vphantom {{\Sigma {{x}_{{ji}}}} n}} \right. \kern-0em} n},$Дистанция (d), от центра рассчитана у каждой особи ( j) для каждой отдельной регистрации (i) как разница координат поимок:
Эти дистанции отклоняются в разные стороны, т.е. могут иметь как положительное, так и отрицательное значение, и, теоретически, не имеют пределов. Поскольку распределение отклонений включает дистанции, образовавшиеся за cчет случайного действия многих неопределенных факторов (положения трансекта на участке особи, индивидуальных особенностей, режима активности в период учета и т.д.) в соответствии с центральной предельной теоремой, можно ожидать, что распределение отклонений в выборке в целом будет стремиться к нормальному. Проверка в тесте Шапиро-Уилка показала соответствие фактических отклонений ожидаемому нормальному распределению (рис. 1а). Кумулята нормального распределения, т.е. накопленная к данному отклонению от центра распределения сумма значений нормально распределенной величины, хорошо аппроксимирует долю накопленных поимок (рис. 1б).
Рис. 1.
Дистанции отклонения (а) и накопленные частоты наблюдений (поимки) на разном удалении от центра участка особи в выборке в целом (б). 1 – фактические частоты, 2 – кривая нормального распределения (а) и кумулята частот нормального распределения (б), 3 – ожидаемые доли от максимального числа поимок (1–fd) при пересечении участка на разной дистанции от центра (пояснения в тексте). Всего 87 наблюдений (поимок) 19 сеголеток S. araneus в августе 2016 г., 3SD = 17.2 м.
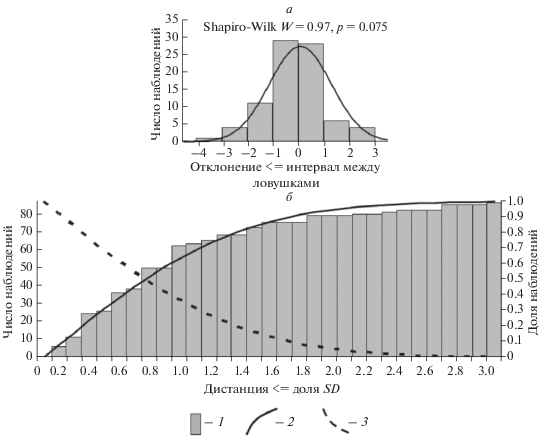
Поскольку кумулята нормального распределения аппроксимирует доли полученных к данной дистанции поимок, в соответствии с правилом трех сигм можно считать, что на дистанции 3 стандартных отклонения (SD) будет получено 99.73% всех возможных поимок особи. Если рассматривать все отклонения выборки как единый “составной” участок, можно ожидать, что при пересечении такого участка учетной линией в центре будут получены все возможные поимки, т.е. вероятность получить максимум возможных регистраций равна 1. При пересечении “составного” участка трансектом на дистанции d от центра исключается часть пространства, на которой накоплена доля поимок fd : fd = Σ fi , где fi – плотность вероятности, или ожидаемая частота события, на i-й дистанции (i от нуля до d) при нормальном распределении. Замечу, что fd соответствует высоте кумуляты на этой дистанции (сплошная линия на рис. 1). Ожидаемые доли от максимально возможного числа поимок при пересечении такого участка линией с увеличением удаления от центра убывают в соответствии с распределением 1–fd (пунктир на рис. 1).
Распределение частот встречаемости особей с разным числом поимок
Расчет ожидаемого соотношения особей с разным числом поимок является ключевым для дальнейших вычислений. При построении распределения ожидаемых частот встречаемости особей с разным числом поимок вслед за Калининым (2012) допускается, что 1) существует равная вероятность попадания центра участка на любое удаление от учетной линии, или, иначе говоря, пересечение участка линией равновероятно на любой дистанции от центра, 2) распределение отклонений на реальном участке стремится к нормальному при накоплении числа поимок. При этом количество особей с различным числом поимок будет соответствовать распределению 1–fd. Таким образом, для разного числа предполагаемых максимально возможных поимок (M) можно рассчитать число ожидаемых поимок на каждом шаге дистанции от центра распределения, умножив M на (1–fd). В качестве шага дистанции принято значение 0.1 стандартного отклонения (SD). При расчете получаются дробные значения, но число поимок не может быть дробным, поэтому полученные значения округлены с порогом 0.5. При вероятности больше 0.5 я считал, что особь может быть поймана, при вероятности меньше 0.5 – нет. После округления получаем ряд целых величин. Эти величины соответствуют числу поимок, которые могут быть получены на определенном удалении от центра участка при разном максимальном числе поимок. В табл. 1 приведен пример такого расчета для случаев, когда особь может быть поймана максимум 2 и 3 раза.
Таблица 1.
Пример расчета числа поимок, которые могут быть получены на определенном удалении от центра участка при заданном максимальном числе поимок
| Шаги дистанции (s) | Дистанция от центра (d), SD | Кумулята (fd) | Доля от максимально возможных поимок (1– fd) | Максимально возможное число поимок | |||
|---|---|---|---|---|---|---|---|
| 2 | 3 | ||||||
| 2(1– fd) | округленное | 3(1– fd) | округленное | ||||
| 1 | 0 | 0.000 | 1.000 | 2.0 | 2 | 3.0 | 3 |
| 2 | 0.1 | 0.080 | 0.920 | 1.8 | 2 | 2.8 | 3 |
| 3 | 0.2 | 0.159 | 0.842 | 1.7 | 2 | 2.5 | 3 |
| 4 | 0.3 | 0.236 | 0.764 | 1.5 | 2 | 2.3 | 2 |
| 5 | 0.4 | 0.313 | 0.687 | 1.4 | 1 | 2.1 | 2 |
| 6 | 0.5 | 0.383 | 0.617 | 1.2 | 1 | 1.9 | 2 |
| 7 | 0.6 | 0.452 | 0.549 | 1.1 | 1 | 1.6 | 2 |
| 8 | 0.7 | 0.516 | 0.484 | 1.0 | 1 | 1.5 | 1 |
| 10 | 0.8 | 0.576 | 0.424 | 0.8 | 1 | 1.3 | 1 |
| 11 | 0.9 | 0.632 | 0.368 | 0.7 | 1 | 1.1 | 1 |
| 12 | 1 | 0.682 | 0.318 | 0.6 | 1 | 1.0 | 1 |
| 13 | 1.1 | 0.729 | 0.271 | 0.5 | 1 | 0.8 | 1 |
| 14 | 1.2 | 0.770 | 0.230 | 0.4 | 0 | 0.7 | 1 |
| 15 | 1.3 | 0.806 | 0.194 | 0.4 | 0 | 0.6 | 1 |
| 16 | 1.4 | 0.839 | 0.162 | 0.3 | 0 | 0.5 | 0 |
| 17 | 1.5 | 0.866 | 0.134 | 0.3 | 0 | 0.4 | 0 |
| 18 | 1.6 | 0.890 | 0.110 | 0.2 | 0 | 0.3 | 0 |
| 19 | 1.7 | 0.911 | 0.089 | 0.2 | 0 | 0.3 | 0 |
| 20 | 1.8 | 0.928 | 0.072 | 0.0 | 0 | 0.2 | 0 |
| 21 | 1.9 | 0.943 | 0.058 | 0.0 | 0 | 0.2 | 0 |
| 22 | 2 | 0.955 | 0.046 | 0.0 | 0 | 0.1 | 0 |
| 23 | 2.1 | 0.964 | 0.036 | 0.0 | 0 | 0.1 | 0 |
| 24 | 2.2 | 0.972 | 0.028 | 0.0 | 0 | 0.1 | 0 |
| 25 | 2.3 | 0.979 | 0.022 | 0.0 | 0 | 0.1 | 0 |
| 26 | 2.4 | 0.984 | 0.016 | 0.0 | 0 | 0.0 | 0 |
| 27 | 2.5 | 0.988 | 0.012 | 0.0 | 0 | 0.0 | 0 |
| 28 | 2.6 | 0.991 | 0.009 | 0.0 | 0 | 0.0 | 0 |
| 29 | 2.7 | 0.993 | 0.007 | 0.0 | 0 | 0.0 | 0 |
| 30 | 2.8 | 0.995 | 0.005 | 0.0 | 0 | 0.0 | 0 |
| 31 | 2.9 | 0.996 | 0.004 | 0.0 | 0 | 0.0 | 0 |
| 32 | 3 | 0.997 | 0.003 | 0.0 | 0 | 0.0 | 0 |
Для расчета долей особей с разным числом поимок на разных расстояниях от центра нужно вычислить долю расстояния (число шагов дистанций – s в табл. 1) от расстояния (числа шагов), на котором при данном максимуме возможных поимок особь будет поймана хотя бы 1 раз. Вычисление проводим для округленных значений. Пример вычисления приведен в табл. 2 (см. столбец “Доля”). В отношении оседлых особей исследователь имеет фактическое распределение только для особей с двумя и более поимками. Но не все особи с одной поимкой нерезиденты. Количество оседлых среди особей с одной поимкой в выборке предстоит вычислить. Поэтому для дальнейших расчетов потребуется узнать распределение (доли) особей, которые могут быть пойманы повторно, т.е. 2 раза и более. Для этого число шагов с количеством поимок больше 1 (>1) относим к сумме шагов дистанции с поимками >1 (см. табл. 2). Для последующего вычисления количества оседлых особей, которые пойманы 1 раз, использован коэффициент K – отношение числа шагов с одной поимкой (s1) к сумме шагов с повторными поимками (Σs>1): K= s1/ Σs>1. Пример расчета распределения ожидаемых долей и коэффициента K для 2 и 3 максимально возможных поимок приводится в табл. 2. Ожидаемое распределение долей особей с разным числом повторных поимок и коэффициент K, для максимально возможного числа поимок от 2 до 20, представлены в табл. 3.
Таблица 2.
Пример расчета коэффициента K для двух и трех максимально возможных поимок (см. табл. 1)
| Число поимок после округления | Максимально возможное число поимок | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| 2 | 3 | |||||||||
| Σ s* | доля Σ s/sT | Σ s | доля > 1 Σs/Σ s>1 | K Σs1/Σ s>1 |
Σs | доля Σs/sT | Σ s | доля > 1 Σ s/Σs>1 | K Σs1/Σ s>1 |
|
| 1 | 8 | 0.67 | 2.00 | 7 | 0.50 | 1.00 | ||||
| 2 | 4 | 0.33 | 4 | 1 | 4 | 0.29 | 4 | 0.57 | ||
| 3 | 0 | 0 | 0 | 3 | 0.21 | 3 | 0.43 | |||
| 4 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | |||
| Σs>1 | 4 | 7 | ||||||||
| Всего (sT) ΣΣs | 12 | 14 | ||||||||
* s – шаг дистанции в табл. 1.
Таблица 3.
Теоретическое распределение долей особей с числом поимок более 1 и коэффициент K, в зависимости от заданного максимального числа поимок
| Число поимок особи | Заданное число максимальных поимок | ||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 15 | 16 | 17 | 18 | 19 | 20 | |
| 2 | 1.00 | 0.57 | 0.44 | 0.36 | 0.25 | 0.23 | 0.21 | 0.21 | 0.20 | 0.13 | 0.19 | 0.13 | 0.18 | 0.18 | 0.12 | 0.17 | 0.17 | 0.11 | 0.11 |
| 3 | 0 | 0.43 | 0.33 | 0.27 | 0.25 | 0.23 | 0.21 | 0.14 | 0.13 | 0.13 | 0.13 | 0.13 | 0.12 | 0.12 | 0.12 | 0.11 | 0.11 | 0.11 | 0.11 |
| 4 | 0 | 0 | 0.22 | 0.18 | 0.17 | 0.15 | 0.14 | 0.14 | 0.13 | 0.13 | 0.13 | 0.13 | 0.12 | 0.06 | 0.12 | 0.06 | 0.06 | 0.11 | 0.06 |
| 5 | 0 | 0 | 0 | 0.18 | 0.17 | 0.15 | 0.14 | 0.07 | 0.13 | 0.13 | 0.06 | 0.06 | 0.06 | 0.06 | 0.06 | 0.11 | 0.06 | 0.06 | 0.11 |
| 6 | 0 | 0 | 0 | 0 | 0.17 | 0.15 | 0.07 | 0.14 | 0.07 | 0.07 | 0.06 | 0.13 | 0.06 | 0.12 | 0.06 | 0.06 | 0.06 | 0.06 | 0.06 |
| 7 | 0 | 0 | 0 | 0 | 0 | 0.08 | 0.14 | 0.07 | 0.07 | 0.07 | 0.13 | 0.06 | 0.06 | 0.06 | 0.06 | 0.06 | 0.06 | 0.06 | 0.06 |
| 8 | 0 | 0 | 0 | 0 | 0 | 0 | 0.07 | 0.14 | 0.13 | 0.13 | 0.06 | 0.06 | 0.06 | 0.06 | 0.06 | 0.06 | 0.06 | 0.06 | 0.06 |
| 9 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.07 | 0.07 | 0.07 | 0.06 | 0.06 | 0.06 | 0.06 | 0.06 | 0.06 | 0.06 | 0.06 | 0 |
| 10 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.07 | 0.07 | 0.06 | 0.06 | 0.06 | 0.06 | 0.06 | 0.06 | 0.06 | 0.06 | 0.06 |
| 11 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.07 | 0.06 | 0.06 | 0.06 | 0.06 | 0.06 | 0 | 0.06 | 0 | 0.06 |
| 12 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.06 | 0.06 | 0.06 | 0* | 0.06 | 0.06 | 0.06 | 0.06 | 0.06 |
| 13 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.06 | 0.06 | 0.06 | 0.06 | 0.06 | 0 | 0.06 | 0 |
| 14 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.06 | 0.06 | 0 | 0.06 | 0.06 | 0 | 0.06 |
| 15 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.06 | 0.06 | 0 | 0.06 | 0.06 | 0.06 |
| 16 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.06 | 0.06 | 0 | 0.06 | 0 |
| 17 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.06 | 0.06 | 0.06 | 0.06 |
| 18 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.06 | 0 | 0.06 |
| 19 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.06 | 0 |
| 20 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.06 |
| K | 2.00 | 1.00 | 0.78 | 0.55 | 0.50 | 0.46 | 0.36 | 0.43 | 0.33 | 0.40 | 0.31 | 0.31 | 0.24 | 0.29 | 0.29 | 0.22 | 0.28 | 0.28 | 0.28 |
Расчет числа оседлых и нерезидентов в выборке
Процедура расчета предусматривает с поиск такого распределения частот встречаемости особей с разным количеством поимок, которое ближе всего к фактическому. Различия фактического и симулированного распределений минимальны при наименьшей сумме квадратов разницы ожидаемого и фактического числа особей с соответствующим числом поимок. Расчет удобно рассмотреть на примере линий 1 и 5 на площадке мечения в 2018 г. В колонке “Факт” (табл. 4) приведено фактическое число особей с разным количеством поимок. После умножения общего числа особей с двумя и более поимками (в табл. 4 “Сумма > 1”) на соответствующую долю (“Множитель из табл. 3”) получаем количества особей с числом поимок, начиная с двух для разного “Максимального числа поимок”. При вычитании ожидаемого числа особей из фактического количества и возведении разницы в квадрат получаем “Квадраты отклонений”. Суммируем квадраты и выбираем колонку, для которой получена наименьшая сумма квадратов отклонений. Для линии 1 наименьшая сумма 0.91 квадратов соответствует ожидаемому распределению количества особей с разным числом поимок для пяти максимальных поимок. Для линии 5 – наименьшая сумма – 1.43, соответствует четырем максимальным поимкам. Величина K по табл. 2, для пяти поимок равна 0.55 (линия 1), а для четырех – 0.78 (линия 5). Умножив сумму зверьков с числом ловов более одного (в табл. 4 это строка “Сумма > 1) на K, получаем ожидаемое число оседлых особей, давших 1 поимку (строка “С 1”). Суммируя оседлых особей с одной поимкой и особей с повторными поимками, получаем общее число оседлых (строка “Всего/Оседлые”). На линии 1 это – 16 особей, на линии 5–12 особей. Вычитая это количество из общего числа фактически пойманных зверьков (строка “Всего/Оседлые” в колонке “Факт”), получаем количество нерезидентов для облавливаемого участка особей. В случае линии 1 это – 3, в случае линии 5 – 0. Фактическое и ожидаемые для этого примера распределения особей с разным числом поимок показаны на рис. 2.
Таблица 4.
Пример расчета количества нерезидентных особей на линиях 1 и 5 в 2018 г.
| Линия | Число поимок | Число особей | Квадраты отклонений для максимального числа поимок (E–F)2 | Множитель из табл. 3. Доля особей с числом поимок >1 (P) | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| факт – F | для максимального числа поимок – E (E = FP) | |||||||||||||
| 3 | 4 | 5 | 6 | 3 | 4 | 5 | 6 | 3 | 4 | 5 | 6 | |||
| Линия 1 | 1 | 9 | ||||||||||||
| 2 | 4 | 5.71 | 4.44 | 3.64 | 2.50 | 2.94 | 0.20 | 0.13 | 2.25 | 0.57 | 0.44 | 0.36 | 0.25 | |
| 3 | 3 | 4.29 | 3.33 | 2.73 | 2.50 | 1.65 | 0.11 | 0.07 | 0.25 | 0.43 | 0.33 | 0.27 | 0.25 | |
| 4 | 1 | 0.00 | 2.22 | 1.82 | 1.67 | 1.00 | 1.49 | 0.67 | 0.44 | 0.00 | 0.22 | 0.18 | 0.17 | |
| 5 | 2 | 0.00 | 0.00 | 1.82 | 1.67 | 4.00 | 4.00 | 0.03 | 0.11 | 0.00 | 0.00 | 0.18 | 0.17 | |
| 6 | 0 | 0.00 | 0.00 | 0.00 | 1.67 | 0.00 | 0.00 | 0.00 | 2.78 | 0.00 | 0.00 | 0.00 | 0.17 | |
| 7 | 0 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | |
| 8 | 0 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | |
| Сумма > 1 | 10 | 10 | 10 | 10 | 10 | 9.59 | 5.80 | 0.91 | 5.83 | |||||
| K (по табл. 2) | 1 | 0.78 | 0.55 | 0.5 | ||||||||||
| С 1 (=1F) | 9 | 10 | 8 | 6 | 5 | |||||||||
| Всего/Оседлые | 19 | 20 | 18 | 16 | 15 | |||||||||
| Нерезидентов (Всего F–E) | –1 | 1 | 3 | 4 | ||||||||||
| Линия 5 | 1 | 5 | ||||||||||||
| 2 | 3 | 4.00 | 3.11 | 2.55 | 1.75 | 1.00 | 0.01 | 0.21 | 1.56 | 0.57 | 0.44 | 0.36 | 0.25 | |
| 3 | 2 | 3.00 | 2.33 | 1.91 | 1.75 | 1.00 | 0.11 | 0.01 | 0.06 | 0.43 | 0.33 | 0.27 | 0.25 | |
| 4 | 1 | 0.00 | 1.56 | 1.27 | 1.17 | 1.00 | 0.31 | 0.07 | 0.03 | 0.00 | 0.22 | 0.18 | 0.17 | |
| 5 | 0 | 0.00 | 0.00 | 1.27 | 1.17 | 0.00 | 0.00 | 1.62 | 1.36 | 0.00 | 0.00 | 0.18 | 0.17 | |
| 6 | 1 | 0.00 | 0.00 | 0.00 | 1.17 | 1.00 | 1.00 | 1.00 | 0.03 | 0.00 | 0.00 | 0.00 | 0.17 | |
| 7 | 0 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | |
| 8 | 0 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | |
| Сумма > 1 | 7 | 7 | 7 | 7 | 7 | 4.00 | 1.43 | 2.91 | 3.04 | |||||
| K (по табл. 3) | 1 | 0.78 | 0.55 | 0.5 | ||||||||||
| С 1 (=1F) | 5 | 7 | 5 | 4 | 4 | |||||||||
| Всего/Оседлые | 12 | 14 | 12 | 11 | 11 | |||||||||
| Нерезидентов (Всего F-E) | –2 | 0 | 1 | 1 | ||||||||||
Рис. 2.
Распределение частот встречаемости особей с разным числом поимок на линиях для вариантов с наличием (а) и отсутствием (б) нерезидентов (2018 г., линии 1 и 5). 1 – натуральные частоты, 2 – ожидаемое распределение для 3, 4, 5 и 6 максимально возможных поимок. Белый кружок маркирует наиболее совпадающее распределение.

Для практического использования метода удобно сделать матрицу расчетов по образцу табл. 4, используя формулы, приведенные в шапках колонок, и взяв множитель и коэффициент К из табл. 3. Убедившись, что отклонения в выборке соответствуют нормальному распределению, после подстановки распределения особей по количеству поимок в колонку “Факт” остается лишь выбрать результат в колонке с ожидаемым распределением, которому соответствует минимальная сумма квадратов отклонений.
Индексы нерезидентности
На основании получаемых данных невозможно охарактеризовать абсолютное обилие нерезидентов. Однако можно получить сравнимые между собой оценки их обилия. Для этого использованы индексы.
Общую величину потока можно оценивать индексом нерезидентности, Inr (Калинин, 2012), т.е. числом обнаруженных нерезидентов (N) на единицу ловчего усилия:
где T – число проверок, а Л – число ловушек в линии.Давление иммиграции на резидентное население можно оценивать двумя индексами:
Индекс пропорции нерезидентов, Ip, к резидентному населению, числу оседлых (nR):
Индекс уловистости нерезидентов, Ic (Shchipanov et al., 2005), – отношение числа поимок нерезидентов к суммарному количеству поимок всех резидентов СR. При этом сумма поимок нерезидентов равна их количеству (N):
Подробнее оценка характеристик, получаемых с использованием этих индексов, дается в разделе “обсуждение”.
Расчет плотности популяции
Если исходить из общей идеологии нормального распределения, то облавливаемую площадь можно определить исходя из представлений о том, что 99.9% поимок среднестатистической оседлой особи укладываются в пределы ±3SD для всей выборки. На самом деле в зависимости от максимально возможного числа поимок особь можно обнаружить на меньшем пространстве, т.е. только на том пространстве, где она может дать хотя бы одну поимку. Это пространство может быть рассчитано как ± доля SD, где при данном максимуме поимок сохраняется возможность обнаружения особи. Для расчета необходимо умножить долю от максимально возможных поимок (1–fd) на дистанции d на ожидаемое максимально возможное число поимок. Дистанция d, при которой возможно получение хотя бы одной поимки, соответствует радиусу пространства (KSD), на котором возможно обнаружение оседлого зверька (табл. 5).
Таблица 5.
Дистанции от трансекта (KSD), на которых оседлая особь может быть обнаружена при данном максимальном числе поимок
| KSD | Доля от максимально возможных поимок (1– fd) | Максимально возможное число поимок | ||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 15 | 16 | 17 | 18 | 19 | 20 | ||
| 0 | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 15 | 16 | 17 | 18 | 19 | 20 |
| 0.1 | 0.920 | 2 | 3 | 4 | 5 | 6 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 15 | 16 | 17 | 17 | 18 |
| 0.2 | 0.842 | 2 | 3 | 3 | 4 | 5 | 6 | 7 | 8 | 8 | 9 | 10 | 11 | 12 | 13 | 13 | 14 | 15 | 16 | 17 |
| 0.3 | 0.764 | 2 | 2 | 3 | 4 | 5 | 5 | 6 | 7 | 8 | 8 | 9 | 10 | 11 | 11 | 12 | 13 | 14 | 15 | 15 |
| 0.4 | 0.687 | 1 | 2 | 3 | 3 | 4 | 5 | 5 | 6 | 7 | 8 | 8 | 9 | 10 | 10 | 11 | 12 | 12 | 13 | 14 |
| 0.5 | 0.617 | 1 | 2 | 2 | 3 | 4 | 4 | 5 | 6 | 6 | 7 | 7 | 8 | 9 | 9 | 10 | 10 | 11 | 12 | 12 |
| 0.6 | 0.549 | 1 | 2 | 2 | 3 | 3 | 4 | 4 | 5 | 5 | 6 | 7 | 7 | 8 | 8 | 9 | 9 | 10 | 10 | 11 |
| 0.7 | 0.484 | 1 | 1 | 2 | 2 | 3 | 3 | 4 | 4 | 5 | 5 | 6 | 6 | 7 | 7 | 8 | 8 | 9 | 9 | 10 |
| 0.8 | 0.424 | 1 | 1 | 2 | 2 | 3 | 3 | 3 | 4 | 4 | 5 | 5 | 6 | 6 | 6 | 7 | 7 | 8 | 8 | 8 |
| 0.9 | 0.368 | 1 | 1 | 1 | 2 | 2 | 3 | 3 | 3 | 4 | 4 | 4 | 5 | 5 | 6 | 6 | 6 | 7 | 7 | 7 |
| 1 | 0.318 | 1 | 1 | 1 | 2 | 2 | 2 | 3 | 3 | 3 | 3 | 4 | 4 | 4 | 5 | 5 | 5 | 6 | 6 | 6 |
| 1.1 | 0.271 | 1 | 1 | 1 | 1 | 2 | 2 | 2 | 2 | 3 | 3 | 3 | 4 | 4 | 4 | 4 | 5 | 5 | 5 | 5 |
| 1.2 | 0.230 | 1 | 1 | 1 | 1 | 2 | 2 | 2 | 2 | 3 | 3 | 3 | 3 | 3 | 4 | 4 | 4 | 4 | 5 | |
| 1.3 | 0.194 | 1 | 1 | 1 | 1 | 1 | 2 | 2 | 2 | 2 | 2 | 3 | 3 | 3 | 3 | 3 | 3 | 4 | 4 | |
| 1.4 | 0.162 | 1 | 1 | 1 | 1 | 1 | 1 | 2 | 2 | 2 | 2 | 2 | 2 | 3 | 3 | 3 | 3 | 3 | ||
| 1.5 | 0.134 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 3 | 3 | ||
| 1.6 | 0.110 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | |||
| 1.7 | 0.089 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 2 | 2 | 2 | 2 | ||||
| 1.8 | 0.072 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | |||||
| 1.9 | 0.058 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | |||||||
| 2 | 0.046 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | |||||||||
| 2.1 | 0.036 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | ||||||||||||
| 2.2 | 0.028 | 1 | 1 | 1 | ||||||||||||||||
| 2.3 | 0.022 | |||||||||||||||||||
| Дистанции (доли SD), на которых может быть получена хотя бы одна поимка | 1.1 | 1.3 | 1.5 | 1.6 | 1.7 | 1.8 | 1.8 | 1.9 | 1.9 | 2.0 | 2.0 | 2.0 | 2.1 | 2.1 | 2.1 | 2.1 | 2.2 | 2.2 | 2.2 | |
Соответственно площадь облова (S) рассчитана как длина линии в метрах (L), умноженная на 2KSD в метрах, поскольку отклонение возможно в обе стороны от линии (см. рис. 1): S = 2KSDL или S = 2KSDL/10 000 (га). На этом пространстве находятся центры участков всех особей, которые определены как оседлые на основе анализа распределения их по числу поимок (см. табл. 3). SD в данном случае рассчитано как корень из суммы квадратов отклонений (d 2), деленных на количество отклонений, минус 1:
(6)
$SD = {{\surd \Sigma {{d}^{2}}} \mathord{\left/ {\vphantom {{\surd \Sigma {{d}^{2}}} {n--1}}} \right. \kern-0em} {n--1}}.$Соответственно, плотность популяции (D) рассчитана как отношение общего количества оседлых (N) к площади в гектарах:
На конкретном примере этот расчет выглядит следующим образом. На линии 5 в 2018 г. 7 зверьков были пойманы более 1 раза. Отклонения от центра координат у особи можно легко получить для данных базы Access, находящейся в стандартном пакете Microsoft Office. Для этого в запросе следует затребовать среднее значение номеров ловушек (Avg), а в связанном запросе – “выражение”: Avg-номер ловушки. Отклонения (d), рассчитанные (формула 2) в номерах ловушек, составили для зверька № 1: –1.5, 1.5; № 2: –1.25, 1.25, 1.25, –1.25; № 3: –0.33, –0.33, 0.67; № 4: –1.75, 1.75; № 5: 0.33, –1.67, –0.6, 1.33, 0.33, 0.33; № 6: –0.2, 0,2; и № 7: –0.4, –0.4, 0.8. Для 22 отклонений сумма d 2 равна 23.02. Соответственно SD (формула 6) равно √23.93/21 = 1.07 дистанции между ловушками. Для перевода в метры умножаем эту величину на дистанцию между ловушками – 7.5 м и получаем SD = 8.0 м. Для линии 5 наиболее подходит модель для четырех максимально возможных поимок (см. табл. 3). Для четырех максимально возможных ловов KSD равно 1.5 или 1.5 × 7.5 = = 11.25 м. Длина 5-й линии равна 112.5 м. В итоге облавливаемая площадь составляет (2 × 11.25 × × 112.5)/10 000 = 0.25 га. На основании анализа распределения особей с разной частотой отловов (см. табл. 3) на линии в качестве оседлых определено 12 зверьков (см. строку Всего/Оседлые в табл. 4). Таким образом, плотность на линии (формула 7) – 47.4 особей/га.
Очевидно, что плотность на короткой линии характеризует лишь этот конкретный участок. Для расчета плотности на площадке в целом, для каждой из линий отдельно были рассчитаны облавливаемая площадь и количество резидентов. Подробнее эти расчеты описаны ниже.
РЕЗУЛЬТАТЫ
Плотность на площадке, как на площади без учета подразделения на линии рассчитана для зверьков, положение центра участка которых находится в пределах площади, ограниченной периметром площадки мечения. С учетом того, что разброс поимок (SD) у обыкновенных бурозубок обычно около 10 м (табл. 6), а вероятность повторной поимки, как правило, ограничена SD (в среднем распределения соответствуют примерно пяти поимкам), я считал, что у животных, пойманных в краевые ловушки, вероятность положения центра участка как внутри, так и снаружи площадки одинакова. Всех землероек, пойманных на площадке однократно, считал нерезидентами. Соответственно, из общего количества пойманных более одного раза зверьков вычтено половинное количество зверьков, пойманных на периметре (см. табл. 6).
Таблица 6.
Результаты расчетов плотности оседлых особей и индексов нерезидентности на площадке мечения
| Год | Проверок | Особи | SD, м | Поимки | Плотность особей/га |
Inr | Ip | 10 × Ic | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Всего (C) | с повторными ловами (С > 1) | 0.5 от числа оседлых на периметре | оседлые с учетом краевого эффекта | нерезиденты (C-C > 1) | всего | оседлых | |||||||
| 2014 | 23 | 52 | 28 | 3 | 25 | 24 | 9.9 | 271 | 247 | 38.3 | 1.20 | 0.96 | 0.97 |
| 2015 | 11 | 21 | 19 | 1 | 18 | 2 | 7.6 | 94 | 92 | 27.6 | 0.21 | 0.11 | 0.22 |
| 2016 | 9 | 26 | 21 | 5 | 16 | 5 | 9.1 | 92 | 87 | 24.6 | 0.64 | 0.31 | 0.57 |
| 2017 | 16 | 31 | 25 | 5.5 | 19.5 | 6 | 10.1 | 163 | 157 | 29.9 | 0.43 | 0.30 | 0.38 |
| 2018 | 14 | 50 | 35 | 8 | 27 | 15 | 9.9 | 185 | 170 | 41.4 | 1.23 | 0.56 | 0.88 |
При расчете на линиях, для каждой из линий, независимо от других линий, были рассчитаны количество оседлых, облавливаемая площадь и количество нерезидентов. При таком расчете зверьки могли одновременно быть пойманы на нескольких линиях. Поэтому суммарное количество особей, обнаруженных на линиях за период учета, значительно превышает количество особей, обнаруженное на площадке за это же время. Однако и расчетная площадь, облавливаемая линиями, в сумме превышает площадку. Количество оседлых особей и обловленные площади были суммированы и по этим суммарным значениям рассчитана плотность для всей площади в целом (табл. 7).
Таблица 7.
Результаты расчетов плотности оседлых особей и индексов нерезидентности на площадке мечения по линиям
| Год | Число проверок | Линия | Число особей | SD, м | Max число поимок** | KSD*** | Площадь, га**** | Число поимок | Плотность, особи/га | Inr | Ip | 10 × Ic | ||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| всего | оседлых* | пойманных повторно | нерезидентов | всего | оседлых* | |||||||||||
| 2014 | 23 | 1 | 20 | 20 | 14 | 0 | 5.1 | 10 | 1.9 | 0.3 | 80 | 80 | ||||
| 2 | 24 | 21 | 14 | 3 | 10.8 | 7 | 1.8 | 0.5 | 70 | 67 | ||||||
| 3 | 26 | 16 | 9 | 10 | 10.1 | 5 | 1.6 | 0.4 | 49 | 39 | ||||||
| 4 | 16 | 11 | 7 | 5 | 10.5 | 5 | 1.6 | 0.5 | 32 | 28 | ||||||
| 5 | 14 | 11 | 4 | 3 | 10.5 | 7 | 1.7 | 0.4 | 27 | 19 | ||||||
| Σ | 100 | 79 | 48 | 21 | 2.1 | 258 | 233 | 38 | 1.05 | 0.27 | 0.90 | |||||
| 2015 | 11 | 1 | 9 | 9 | 7 | 0 | 8.6 | 3 | 1.3 | 0.3 | 20 | 20 | ||||
| 2 | 8 | 8 | 7 | 0 | 7.1 | 4 | 1.5 | 0.3 | 18 | 18 | ||||||
| 3 | 10 | 10 | 7 | 0 | 5.7 | 4 | 1.5 | 0.2 | 23 | 23 | ||||||
| 4 | 8 | 7 | 4 | 1 | 8.6 | 4 | 1.5 | 0.3 | 16 | 15 | ||||||
| 5 | 5 | 5 | 4 | 0 | 8.6 | 8 | 1.8 | 0.3 | 17 | 17 | ||||||
| Σ | 40 | 39 | 29 | 1 | 1.5 | 103 | 102 | 26 | 0.10 | 0.03 | 0.10 | |||||
| 2016 | 11 | 1 | 11 | 11 | 8 | 0 | 7.2 | 5 | 1.6 | 0.3 | 26 | 26 | ||||
| 2 | 9 | 9 | 6 | 0 | 5.3 | 3 | 1.3 | 0.2 | 17 | 17 | ||||||
| 3 | 8 | 6 | 2 | 2 | 11.9 | 3 | 1.3 | 0.4 | 11 | 9 | ||||||
| 4 | 13 | 11 | 5 | 2 | 11.1 | 3 | 1.3 | 0.4 | 20 | 18 | ||||||
| 5 | 8 | 6 | 4 | 2 | 14.7 | 6 | 1.7 | 0.6 | 18 | 16 | ||||||
| Σ | 49 | 43 | 25 | 6 | 1.9 | 102 | 96 | 23 | 0.63 | 0.14 | 0.63 | |||||
| 2017 | 11 | 1 | 15 | 15 | 10 | 0 | 11.6 | 4 | 1.5 | 0.5 | 40 | 40 | ||||
| 2 | 11 | 11 | 7 | 0 | 7.1 | 5 | 1.6 | 0.3 | 29 | 29 | ||||||
| 3 | 16 | 16 | 8 | 0 | 11.4 | 3 | 1.3 | 0.4 | 26 | 26 | ||||||
| 4 | 10 | 5 | 4 | 5 | 6.1 | 10 | 1.9 | 0.3 | 28 | 25 | ||||||
| 5 | 7 | 7 | 6 | 0 | 9.9 | 10 | 1.9 | 0.4 | 40 | 39 | ||||||
| Σ | 59 | 54 | 35 | 5 | 1.9 | 164 | 155 | 28 | 0.52 | 0.09 | 0.32 | |||||
| 2018 | 11 | 1 | 19 | 16 | 10 | 3 | 8.3 | 5 | 1.6 | 0.4 | 39 | 36 | ||||
| 2 | 19 | 15 | 8 | 4 | 10.9 | 4 | 1.5 | 0.4 | 34 | 30 | ||||||
| 3 | 22 | 17 | 11 | 6 | 9.4 | 6 | 1.7 | 0.4 | 49 | 43 | ||||||
| 4 | 22 | 22 | 11 | 0 | 17 | 3 | 1.3 | 0.6 | 42 | 42 | ||||||
| 5 | 12 | 12 | 7 | 0 | 8 | 4 | 1.5 | 0.3 | 28 | 26 | ||||||
| Σ | 94 | 82 | 47 | 12 | 2.1 | 194 | 158 | 39 | 1.26 | 0.15 | 0.76 | |||||
По методике Калинина (2012) SD было рассчитано как среднее от среднеквадратичного отклонения координат особей с тремя и более поимками. Среднее SD рассчитано для каждой линии отдельно, и облавливаемая площадь рассчитана как длина линии в метрах, умноженная на 2SD в метрах, деленная на 10 000. Также для каждой линии отдельно рассчитано количество резидентов, попадающих в зону ± SD. Затем сумма оседлых особей отнесена к суммарной площади линий.
При расчете SD в одном случае были выбракованы выпадающие значения стандартных отклонений, как это рекомендует Калинин (2012). Во втором случае, на основе анализа распределения дистанций от центра, были выбракованы выпадающие дистанции. Для получения числа нерезидентов, обнаруженных на каждой линии, по методике Калинина (2012) рассчитан избыток особей с единичными поимками.
Плотности, полученные при расчете разными способами, практически не различались. Однако без выбраковки выпадающих дистанций плотность при расчетах по методу Калинина (2012) была существенно ниже, хотя и повторяла динамику, полученную при других расчетах (рис. 3а). Индекс нерезидентности (Inr) практически совпадал для всех способов расчета (рис. 3б). Индекс пропорции на линиях в целом повторял динамику Ip на площадке, но на линиях Ip сильно занижен, поскольку суммарное число оседлых на линиях заведомо больше, чем на площадке (рис. 3в). Индексы уловистости (Ic) нерезидентов были близки при всех вариантах расчета (рис. 3г).
Рис. 3.
Популяционная плотность (а) и индексы нерезидентности – Inr (б), пропорции нерезидентов – Ip (в) и уловистости нерезидентов – Ic (г). 1 – получены на площадке, 2 – рассчитаны для линий по новому способу, 3 – рассчитаны для линий по методу Калинина (2012) с выбраковкой выпадающих значений SD, 4 – рассчитаны для линий по методу Калинина (2012) с выбраковкой выпадающих перемещений.

ОБСУЖДЕНИЕ
Соответствие результатов получено при использовании принципиально разных способов расчета. При расчете, сделанном для площадки, площадь облова фиксированная. При расчете на той же территории по линиям, площадь облова рассчитана исходя из дистанций перемещения, а количество оседлых рассчитано на основе соотношения особей с разным числом поимок. При этом по методу Калинина (2012), плотность рассчитана для особей, участки которых пересекаются линией в пределах дистанции ± среднее SD координат поимок особей с тремя и более поимками, по новому методу – для всего количества особей с повторными ловами в пределах дистанций обнаружения, рассчитываемых для выборки. Число оседлых особей на линиях, и ширина полосы учета, взятые для расчета плотности, при этом не равны. Таким образом, совпадение результатов не является следствием идентичности методов расчета.
Показательно, что во всех трех случаях значение индекса уловистости нерезидентов практически не различаются. Величина этого индекса зависит только от количества особей, определенных для данной территории как нерезиденты, так как число ловушек и количество проверок не различаются. Очевидно, что в выборке число зверьков с единичными поимками при разделении площадки на линии должно увеличиваться, однако и использование метода Калинина (2012) и новый способ расчета предусматривают процедуру исключения части особей с одной поимкой из нерезидентов. Таким образом, можно заключить, что способы определения нерезидентов на основе анализа распределения частот особей с разным количеством поимок валидны.
В своей публикации Калинин (2012) показал, что оцениваемое обилие нерезидентов хорошо соответствует их динамике, определенной другими способами. Однако до сих пор его расчеты плотности не сравнивали с эмпирическими данными, полученными при мечении на площадках. Хорошее совпадение значений расчетной и эмпирической плотности в данном случае свидетельствует о пригодности и метода Калинина (2012), и нового метода для определения локальной плотности популяции. Но при использовании методики Калинина расчетная плотность чувствительна к выпадающим дистанциям перемещения. Их нужно выбраковывать из общего ряда. Таким образом, необходимо составлять ряд распределения отклонений от центра координат для всей выборки. Однако если при расчете по методу Калинина это является лишь предварительной процедурой, то новый метод позволяет напрямую работать с этим рядом. При этом из расчетов исключается довольно громоздкое вычисление количества особей, попадающих в пределы дистанции ± SD от линии. В новой методике табл. 2 содержит все коэффициенты, позволяющие при простой подстановке ряда в матрицу распределений определить число оседлых и нерезидентных особей. Кроме того, новый метод существенно расширяет выборку за счет включения особей с двумя поимками.
Предложенный здесь способ позволяет рассчитать популяционную плотность для тех видов, у которых распределение дистанций отклонений от центральной координаты соответствует нормальному. Методика оперирует только распределением отклонений и слабо чувствительна к количеству повторных ловов. Неважно, хорошо или плохо ловится вид в живоловки, насколько ловушки заняты другим видом или насколько целевой вид чувствителен к посещению ловушек другими видами и т.п. Эти факторы не влияют на распределение особей по частоте посещения ловушек.
Принимая во внимание возможность расчета абсолютной плотности, можно рекомендовать апробированный ранее метод мечения на линиях (Щипанов и др., 2010; Куприянова, Калинин, 2015) как универсальный способ оценки плотности оседлого населения мелких млекопитающих лесной зоны. Важной составляющей экологического мониторинга является оценка величины потоков материи и энергии, связанной с отдельными видами или сообществом в целом. Способы пересчета линейных данных на площадь позволяют относительно легко нормировать показатели биомассы. Проведенная работа показала, что результаты расчета по методике Калинина (2012) и новой методике практически не различаются и выбор способа расчета зависит лишь от предпочтений исследователя.
Важной характеристикой состояния популяции является уровень миграционной активности – одной из наиболее быстрых реакций популяции на дестабилизацию экосистем (Мухачева, Лукьянов, 1997; Лукьянова, Лукьянов, 1998, 2004; Щипанов, 2002; Luk’yanov, Luk’yanova, 2002; Lukyanova et al., 1993; Григоркина, Оленев, 2013). Эта характеристика может быть получена с использованием индексов нерезидентности. Общее обилие нерезидентов в популяции характеризует индекс нерезидентности. Для популяционно-генетических исследований важна оценка величины потока мигрантов относительно обилия резидентного населения. Индекс пропорции отражает количественное соотношение нерезидентов и резидентов, но несравним для выборок, взятых на площади и на линии. В этом отношении лучше использовать индекс уловистости. Он выражен ярче и не зависит и от способа расстановки ловушек.
Однако использование индексов нерезидентности требует осторожности: эти индексы не оценивают абсолютное обилие нерезидентов. Так, у бурозубок оседлые особи хорошо ловятся в ловушки, а нерезидентные особи попадаются преимущественно в ловчие устройства типа “западней” – pitfall (Щипанов и др., 2003; Калинин, 2012). При наблюдениях, проведенных одновременно двумя способами на одной и той же территории, количество пойманных оседлых и нерезидентных особей практически не различалось, что примерно соответствует соотношению один нерезидент: один резидент в сутки (Щипанов и др., 2003). Индексы же относительного обилия показывают, что на одного оседлого в сутки приходятся сотые доли нерезидента. Вместе с тем динамика индекса соответствует динамике добычи в ловчие канавки, т.е. объективно отражает динамику потока нерезидентов (Калинин, 2012). Таким образом, индексы пригодны для межгодовых сравнений динамики обилия нерезидентов одного вида, но могут быть несравнимы для разных видов, так как значения улавливаемости нерезидентов у этих видов могут сильно различаться.
Список литературы
Бердюгин К.И., Большаков В.Н., 2005. Млекопитающие в экологическом мониторинге // Методы экологического мониторинга: большой специальный практикум: учебное пособие. Екатеринбург: Изд-во Уральского университета. С. 192–201.
Бобрецов А.В., Лукьянова Л.Е., Быховец Н.М., Петров А.Н., 2017. Влияние изменения климата на динамику популяций лесных полевок (Myodes) Северного Предуралья: роль ландшафтных эффектов // Сибирский экологический журнал. № 3. С. 245–256.
Григоркина Е.Б., Оленев Г.В., 2013. Миграции грызунов в зоне влияния Восточно-Уральского радиоактивного следа (радиобиологический аспект) // Радиационная биология. Радиоэкология. Т. 53 (1). С. 76–83.
Ивантер Э.В., Макаров А.М., 2001. Территориальная экология землероек-бурозубок (Insectivora, Sorex). Петрозаводск: Петр. ГУ. 272 с.
Ильяшенко В.Б., Лучникова Е.М., Скалон Н.В., 2015. Мелкие млекопитающие как объект биомониторинговых исследований на территории Кемеровской области // Вестник Кемеровского государственного университета. Т. 3. № 4 (64). С. 25–30.
Истомин А.В., 2014. Мелкие млекопитающие в мониторинге лесных экосистем: комплексный подход // Вестник Тверского гос. университета. Сер. Биология и экология. № 4. С. 95–113.
Калинин А.А., 2012. Оседлая и нерезидентная составляющая численности массовых видов мелких млекопитающих по данным учета на линиях живоловок // Зоологический журнал. Т. 91. Вып. 6. С. 759–768.
Калинин А.А., Куприянова И.Ф., 2015. Методика количественного учета мелких млекопитающих при миграциях через водные преграды // Зоологический журнал. Т. 94. № 3. С. 365–369.
Карасева Е.В., Телицына А.Ю., Жигальский О.А., 2008. Методы изучения грызунов в полевых условиях. М.: Изд-во ЛКИ. 416 с.
Катаев Г.Д., 2015. Мониторинг населения мелких млекопитающих северной тайги Фенноскандии // Бюллетень Московского общества испытателей природы. Отд. биол. Т. 120. В. 3. С. 3–13.
Куприянова И.Ф., Калинин А.А., 2015. Сравнение различных показателей численности мелких млекопитающих // Труды Печоро-Илычского заповедника. Вып. 17. С. 113–115.
Лукьянова Л.Е., Лукьянов О.А., 1998. Реакции сообществ и популяций мелких млекопитающих на техногенные воздействия. II. Популяции (рыжая полевка как модель) // Успехи современной биологии. Т. 118. Вып. 6. С. 694–707.
Лукьянова Л.Е., Лукьянов О.А., 2004. Экологически дестабилизированная среда: влияние на население мелких млекопитающих // Экология. № 3. С. 210–217.
Мухачева С.В., Лукьянов О.А., 1997. Миграционная подвижность населения рыжей полевки (Clethrionomys glareolus (Schreber, 1780)) в градиенте техногенных факторов // Экология. № 1. С. 34–39.
Мухачева С.В., Бердюгин К.И., Давыдова Ю.А., 2009.Опыт использования мелких млекопитающих для оценки состояния природных экосистем // Аграрный вестник Урала. № 3. С. 65–68.
Соколов В.Е., Ильенко А.И., 1977. Научное обоснование и принципы экологического мониторинга популяций позвоночных животных // Мониторинг состояния окружающей среды: Труды советско-английского симпозиума, Кардингтон (Англия). Л.: Гидрометеоиздат. С. 228–322.
Щипанов Н.А., 1986. К экологии малой белозубки (Crocidura suaveolens) // Зоологический журнал. Т. 66. № 7. С. 1051–1060.
Щипанов Н.А., 2002. Функциональная организация популяции – возможный подход к изучению популяционной устойчивости. Прикладной аспект (на примере мелких млекопитающих) // Зоологический журнал. Т. 81. № 9. С. 1048–1077.
Щипанов Н.А., Калинин А.А., Олейниченко В.Ю., Демидова Т.Б., Гончарова О.Б., Нагорнев Ф.В., 2000. К методике изучения использования пространства землеройками-бурозубками // Зоологический журнал. Т. 79. № 3. С. 362–371.
Щипанов Н.А., Куприянова И.Ф., Бобрецов А.В., Калинин А.А., Тумасян Ф.А., Щипанов А.Н., 2010. Опыт применения живоловок для мониторинга популяций мелких млекопитающих // Труды Печоро-Илычского заповедника. Вып. 16. Сыктывкар: Коми научный центр УрО РАН. С. 208–221.
Щипанов Н.А., Купцов А.В., Калинин А.А., Демидова Т.Б., Олейниченко В.Ю., Ляпина М.Г., Александров Д.Ю., Распопова А.А., Павлова С.В., Тумасьян Ф.А., 2010а. Мелкие млекопитающие юго-востока Tверской области. Cообщение 1. Фауна и биотопическое распределение // Сибирский экологический журнал. Вып. 5. С. 799–806.
Щипанов Н.А., Купцов А.В., Калинин А.А., Олейниченко В.Ю., 2003. Конуса и живоловки ловят разных землероек бурозубок (Insectivora, Soricidae) // Зоологический журнал. Т. 83. № 10. С. 1258–1265.
Bartell S.M., 2006. Biomarkers, bioindicators, and ecological risk assessment – a brief review and evaluation // Environmental Bioindicators. V. 1. P. 39–52.
Buchner C.H., 1969: Some aspects of the population ecology of the common shrew, S. araneus, near Oxford, England // Journal of Mammalogy. V. 50. P. 326–322.
Burger J., 2006. Bioindicators: types, development, and use in ecological assessment and research // Environmental Bioindicators. V. 1. P. 22–39.
Carignan V., Villard M.-A., 2002. Selecting indicator species to monitor ecological integrity: a review // Environmental Monitoring and Assessment. V. 78. P. 45–61.
Churchfield S., 1990. The natural history of shrews. London: Christopher Helm. 198 p.
Croin Michielsen N., 1966. Intraspecific and interspecific competition in the shrews Sorex araneus L. and S. minutus L. // Archives Néerlandaises de Zoologie. V. 17. P. 73–174.
Landres P.B., Verner J., Thomas J.W., 1988. Ecological uses of vertebrate indicator species: a critique // Conservation Biology. V. 2. P. 316–328.
Luk’yanov O.A., Luk’yanova L.E., 2002. Phenomenology and analysis of dispersal in populations of small mammals // Entomological Review. V. 82 (1). P. 181.
Lukyanova L., Lukyanov O., Pjastolova O., 1993. The response of the bank vole populations to technogenic patchiness // Mammalia. V. 57. № 4. P. 636.
Moraleva N.V., Telitzina A., 1994. Territoriality in juveniles of the common shrew (Sorex araneus) in prepeak and peack years of population density. Advances in the biology of shrews. Merrit J.F., Kirkland G.L. Jr., Rose R.K. (eds). Carnegie Museum Natural History Special Publication 18. P. 67–76.
Shchipanov N.A., Kalinin A.A., Demidova T.B., Oleinichenko V.Yu., Aleksandrov D.Yu., Kouptzov A.V., 2005. Population ecology of red-toothed shrews, Sorex araneus, S. caecutiens, S. minutus, and S. isodon, in Central Russia // Advances in the Biology of Shrews II. J. Merrit, S. Churchfield, R. Hutterer, B.I. Sheftel (eds). N.Y. Special publication of the international society of shrew biologists. № 1. P. 201–216.
Shchipanov N.A., Zima J., Churchfield S., 2019. Introducing the common shrew. Shrews, chromosomes and speciation. J.B. Searle, P.D. Polly, J. Zima (eds). Cambridge, N.Y.: Cambridge University Press. P. 19 – 67.
Shillito J.F., 1963. Field observation on the growth, reproduction and activity of a woodland population of the common shrew Sorex araneus L. // Proceedings of the Zoological Society of London. V. 140. P. 99–113.
Siddig A.A.H., Ellison A.M. Ochs A., Villar-Leeman C., Lau M.K., 2016. How do ecologists select and use indicator species to monitor ecological change? Insights from 14 years of publication in Ecological Indicators // Ecological Indicators. V. 60. P. 223–230.
Spellerberg I.F., 2005. Monitoring Ecological Change. Cambridge: Cambridge University Press. 324 p.
Veatch R.M., 2016. The Basics of Bioethics. 3rd Edition. N.Y.: Routledge. 224 p.
Дополнительные материалы отсутствуют.
Инструменты
Зоологический журнал