

Зоологический журнал, 2021, T. 100, № 11, стр. 1305-1311
Морфометрические характеристики черепа и плечевых костей восточной слепушонки (Ellobius tancrei, Rodentia, Cricetidae) из Самарканда и Бухары (Узбекистан)
Л. Б. Мардонова a, *, С. Н. Гашев a, М. Г. Митропольский a
a Тюменский государственный университет, Институт биологии
625043 Тюмень, Россия
* E-mail: mardonova.luiza@mail.ru
Поступила в редакцию 22.07.2020
После доработки 29.10.2020
Принята к публикации 31.10.2020
Аннотация
Из равнинных частей Узбекистана в начале ХХ века были описаны два подвида восточной слепушонки (Ellobius tancrei): из Самарканда E. t. fusciceps и из Бухары E. t. ognevi. Однако позже их перестали признавать. Вместе с тем наши исследования подтверждают, что эти два подвида имеют достоверные различия по морфометрии черепа, т.е. по тем признакам, по которым эти подвиды были описаны ранее. Наряду с изучением черепа, мы провели анализы морфометрических показателей плечевых костей по методике, разработанной нами. Согласно данным комплексного анализа черепов и плечевых костей, описанные ранее подвиды восточной слепушонки с равнинного Узбекистана заслуживают быть признанными.
Состав подвидовой систематики рода Ellobius Fischer 1814 продолжает обсуждаться. До выделения Топачевским (1965) восточной слепушонки в отдельный вид Ellobius tancrei Blasius 1884, внутривидовая систематика Ellobius talpinus Pallas 1770 подвергалась критике (Огнев, 1950; Corbert, 1978). Относительно внутривидового деления восточной слепушонки (Ellobius tancrei), даже с применением кариологических исследований и последовавшими за ними широкими комплексными морфологическими исследованиями (Воронцов и др., 1969; Воронцов, Якименко, 1984; Громов, Ербаева, 1995), все еще остается ряд вопросов. Среди них объединение подвидов E. t. ognevi и E. t. fusciceps, а также расширение ареала E. t. fusciceps на Приташкентские равнины без сравнения морфологических признаков с горным подвидом E. t. caenosus, с которым у экземпляров с Приташкентских равнин большие сходства по морфометрическим параметрам.
Таким образом, в начале ХХ столетия для территории равнинного Узбекистана было описано два подвида – E. t. fusciceps Thomas 1909 из окрестностей Самарканда и E. t. ognevi Dukelsky 1926 из окрестностей Бухары. В это же время Угаров (1928), изучая систематическое положение слепушонки, отнес сборы из Ташкента и его окрестностей к E. t. ognevi.
В работе мы попытались сопоставить полученные нами данные морфометрических показателей плечевых костей с морфометрическими признаками черепов, описанных в литературе и собранных авторами с территории Узбекистана, для более углубленного изучения равнинных форм восточной слепушонки.
МАТЕРИАЛ И МЕТОДИКА
Для анализа были использованы черепа и плечевые кости восточной слепушонки, собранные в погадках ушастых сов в зимний период (декабрь–апрель) из г. Бухара (парк областной больницы), из г. Самарканд (Центральный бульвар) и из г. Ташкент (Ботанический сад).
Определение видового состава жертв из погадок производилось по костям краниального и посткраниального скелетов с использованием сравнительной коллекции О. и М. Митропольских (2009).
Объемы выборок составили: по 25 черепов и нижних челюстей разной степени целостности из сборов из Самарканда и Ташкента и 11 черепов разной степени целостности и 9 нижних челюстей из сборов с территории Бухары.
Краниологические выборки не были разделены по полу и возрасту, однако анализ-костей посткраниального скелета показал, что доли взрослых особей в выборках из разных городов были примерно сопоставимы между собой: Самарканд – 72.8%, Ташкент – 73.8% и Бухара – 79.8%.
Так как черепа в погадках имеют разную степень сохранности, то основные измерения производили на наименее поврежденных частях черепа. Такие промеры, как длина и высота черепа приведены лишь для нескольких экземпляров. Использовали схему промеров черепа по Огневу (1940) и Потапову (1990). Полученные нами результаты морфометрии черепов были сопоставлены с имеющимися в литературе описаниями.
Для стандартизации и сравнения выборок по плечевым костям использованы кости с полностью приращенными головками на проксимальном эпифизе (взрослые особи) без разделения по полу, собранные в объеме по 50 костей из Самарканда, Бухары и Ташкента.
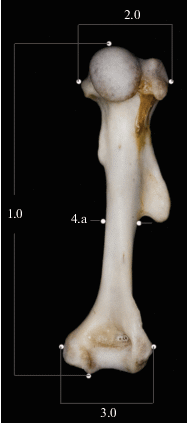
Основные промеры плечевой кости выполнены по схеме Митропольского (2005): 1.0 – общая длинна кости, 2.0 – максимальная ширина проксимального эпифиза, 3.0 – максимальная ширина дистального эпифиза, 4.а–4.в – ширина и толщина диафиза у середины длины кости (рис. 1). Далее для обработки данных мы используем два индекса: Σ 1–4.а – сумму основных промеров (1.0 + 2.0 + 3.0 + 4.а) и Σ 4.а–4.в – сумму промеров диафиза (4.а + 4.в).
Полученные морфометрические показатели плечевых костей из каждой группы восточной слепушонки были проанализированы с помощью компьютерной программы “STATISTICA (data analysis software system), version 10”, с использованием дискриминантного анализа для независимых выборок (Гашев и др., 2014).
Дискриминантный анализ проведен с целью составления уравнений дискриминации на основе “обучающих” модельных групп.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Анализ морфометрии черепа. В работе Thomas (1909) приводит данные только по одному черепу из восьми экземпляров впервые описываемого подвида E. t. fusciceps из окрестностей Самарканда (сборы в апреле 1908 г.), но и они показывают крупные размеры этого черепа (табл. 1), в том числе относительно черепов из нашей выборки (табл. 2).
Таблица 1.
Морфометрические характеристики черепов восточной слепушонки (Ellobius tancrei) по данным литературы
| Признак | Дукельская, 1926 (Бухара) |
Thomas, 1909 (Самарканд) |
Угаров, 1928 (Ташкент) |
Павленко
и др., 1974 (Фергана) |
||||
|---|---|---|---|---|---|---|---|---|
| ∆n | Среднее | min–max | Один экз. | ∆n | Среднее | min–max | Среднее | |
| Общая длина | 9 | 29.6 | 27.7–31 | 32.8 | 8 | 29.7 | 28.9–31.1 | 29.9 ± 0.18 |
| Кондилобазальная длина | 10 | 26.2 | 25.0–27.3 | 31.0 | 10 | 25.8 | 21.8–27.5 | 25.8 ± 0.21 |
| Межглазничная ширина | 10 | 5.9 | 5.4–6.3 | 5.4 | 11 | 5.5 | 4.9–5.8 | 5.4 ± 0.15 |
| Ширина черепной капсулы | 9 | 13.6 | 12.8–14.2 | – | – | – | 13.9 ± 0.17 | |
| Ширина скул | 9 | 20.5 | 19.3–21.4 | 22.3 | 10 | 21.1 | 20.4–22.0 | 19.9 ± 0.21 |
| Длина носовых костей | 11 | 7.7 | 6.7–8.1 | 8.0 | 9 | 7.6 | 6.9–8.0 | 7.5 ± 0.11 |
| Ширина их спереди | 3.4 | 10 | 2.9 | 2.8–3.3 | ||||
| Диастема | 11 | 9.7 | 9.3–10.1 | 11.0 | 11 | 9.7 | 9.2–10.6 | 10.1 ± 0.30 |
| Альвеолярная длина верхнего ряда зубов | 11 | 7.4 | 7.0–7.9 | 7.3 | 11 | 6.8 | 6.7–7.2 | 7.2 ± 0.11 |
| Длина нижней челюсти | 11 | 18.4 | 17.1–19.3 | – | 10 | 19.4 | 19.0–20.6 | – |
| Альвеолярная длина нижнего ряда зубов | 11 | 7.6 | 7.1–7.9 | – | 11 | 6.9 | 6.7–7.2 | 7.3 ± 0.10 |
Таблица 2.
Морфометрические характеристики черепов восточной слепушонки (Ellobius tancrei) из крупных городов Узбекистана по нашим сборам
| Признак | Самарканд | Ташкент | Бухара | ||||||
|---|---|---|---|---|---|---|---|---|---|
| ∆n | Среднее | min–max | ∆n | Среднее | min–max | ∆n | Среднее | min–max | |
| Общая длина | 3 | 30.0 ± 0.51 | 29.3–31.0 | 3 | 30.2 ± 0.56 | 29.1–30.9 | – | – | |
| Кондилобазальная длина | 5 | 27.8 ± 0.39 | 26.6–28.6 | 6 | 27.5 ± 0.53 | 25.8–29.6 | – | – | |
| Межглазничная ширина | 25 | 5.8 ± 0.06 | 5.2–6.4 | 24 | 5.5 ± 0.082 | 4.6–6.1 | 11 | 5.7 ± 0.09 | 5.2–6.2 |
| Ширина черепной капсулы | 25 | 14.1 ± 0.06 | 13.6–14.9 | 15 | 13.8 ± 0.02 | 12.3–14.2 | 2 | 13.6 ± 0.25 | 13.4–13.9 |
| Ширина скул | 25 | 21.3 ± 0.13 | 19.4–22.6 | 24 | 20.7 ± 0.152 | 19.1–21.8 | 5 | 20.5 ± 0.212 | 20.0–21.3 |
| Длина носовых костей | 25 | 7.8 ± 0.08 | 6.9–8.6 | 24 | 7.6 ± 0.11 | 6.6–8.5 | 10 | 7.8 ± 0.06 | 7.5–8.1 |
| Ширина носовых костей спереди | 25 | 2.4 ± 0.053 | 2.4–3.4 | 24 | 2.9 ± 0.05 | 2.4–3.2 | 10 | 2.8 ± 0.07 | 2.5–3.2 |
| Диастема | 25 | 9.3 ± 0.113 | 9.3–11.6 | 25 | 10.1 ± 0.12 | 9.1–10.9 | 11 | 9.9 ± 0.15 | 9.2–10.8 |
| Альвеолярная длина верхнего ряда зубов | 25 | 6.2 ± 0.073 | 6.2–7.5 | 25 | 6.9 ± 0.06 | 6.2–7.4 | 9 | 6.9 ± 0.07 | 6.6–7.2 |
| Высота черепа | 5 | 9.4 ± 0.292 | 8.4–10.0 | 5 | 10.5 ± 0.09 | 10.2–10.7 | – | – | |
| Длина foramen palatinum | 25 | 2.3 ± 0.103 | 1.2–3.6 | 25 | 2.8 ± 0.06 | 2.1–3.4 | 11 | 2.4 ± 0.093 | 1.9–3.1 |
| Общая длина нижней челюсти | 25 | 23.9 ± 0.19 | 22.4–26.1 | 25 | 22.0 ± 0.223 | 19.7–23.9 | 9 | 22.9 ± 0.242 | 21.7–24.0 |
| Длина нижней челюсти без резцов | 25 | 20.6 ± 0.14 | 19.2–21.8 | 25 | 19.3 ± 0.153 | 17.9–20.3 | 9 | 19.7 ± 0.193 | 18.9–21.0 |
| Альвеолярная длина нижнего ряда зубов | 25 | 7.2 ± 0.08 | 6.4–8.2 | 25 | 6.7 ± 0.063 | 6–7.2 | 9 | 6.9 ± 0.091 | 6.6–7.5 |
Примечания. 1 Различия достоверны с максимальным значением при р < 0.05; 2 – при р < 0.01; 3 – при р < 0.001. Например, в строке “ширина скул” достоверность различий значений других групп указана относительно самаркандской группы (где показатель максимальный), а в строке “ширина носовых костей спереди” – относительно ташкентской и т.д. Прочерк – нет данных.
В работе Дукельской (1926) даны описания еще одного подвида E. t. ognevi из окрестностей Бухары по сборам из 11 экз. в мае–июле 1925 г. Сравнение данных промеров с нашим материалом по морфометрическим признакам черепа показывает большое сходство и общую закономерность: черепа зверьков, описанные с равнинных территорий Узбекистана, являются наиболее короткими и узкими (табл. 1 и 2).
Угаров (1928) отмечает морфометрическое сходство сборов из Бухары с образцами из Ташкента, что подтверждается и нашими данными. Также имеется сходство с выборкой из Ферганской долины (Павленко, Аллабергенов, 1974), отнесенной авторами к подвиду E. t. fusciceps. Отметим, что Thomas (1909) приводит более крупные размеры черепа слепушонок. В работе Воронцова и Якименко (1984) представлены промеры черепа для обобщенной группы E. tancrei по материалам с территории всего Южного Узбекистана, что не позволяет сравнить данные с рассматриваемыми нами локальными выборками.
Якименко (2003) обнаружила различия восточной слепушонки из Самарканда и Бухары по длине нижней челюсти, общей и кондилобазальной длине черепа, межглазничной ширине, ширине мозговой капсулы, альвеолярной длине верхнего ряда коренных зубов. В то же время отмечается сходство промеров черепов из Бухары и Ташкента.
Сопоставив морфометрические показатели черепа восточной слепушонки из разных мест ареала вида в равнинном Узбекистане, как по литературным материалам, так и по собственным сборам, мы обнаружили четкие различия у описанных в начале ХХ столетия двух форм этого вида из окрестностей Самарканда и Бухары: по ширине скул, общей длине нижней челюсти, длине нижней челюсти без резцов и альвеолярной длине нижнего ряда зубов. Животные из Ташкента достоверно отличаются от самаркандских по межглазничной ширине, ширине скул, общей длине нижней челюсти, длине нижней челюсти без резцов и альвеолярной длине нижнего ряда зубов (табл. 2).
Изменчивость плечевой кости. Нами применен метод анализа подвидовой систематики на основе морфометрических характеристик плечевой кости, который ранее показал свою эффективность (Митропольский, Митропольский, 2016).
Морфологические и морфометрические признаки костей посткраниального скелета практически не используется в таксономии. Мы согласны с замечанием Громова (1983: с. 6), который указывал на целесообразность использования в систематике грызунов не только характеристик зубной системы.
Построение систем с использованием связанных с плечевой костью признаков, с нашей точки зрения, перспективны. Проксимальный отдел передней конечности, связывающий ее с туловищем, видимо, не только крайне специфичен для всех групп, но и наиболее стабилен в эволюции. Его практически не затрагивают частные адаптации, широко представленные в черепе, зубах и задних конечностях (Митропольский, 2005).
Из видов, обитающих на территории Узбекистана, в диапазон морфологических признаков и морфометрических показателей плечевых костей восточной слепушонки попадают илийская полевка (Microtus ilaeus) Thomas 1912, краснохвостая песчанка (Meriones libycus) Lichtenstein 1823, серая крыса (Rattus norvegicus) Berkenhout 1769 и несколько видов тушканчиков семейства Dipodidae Fischer 1817.
Морфологическое описание плечевой кости. Первое короткое описание плечевой кости рода Ellobius приводит Громов (1963) в монографии “Млекопитающие фауны СССР” (с. 554). “Плечевая кость сравнительно короткая с коротким и высоким гребнем дельтовидной бугристости и слабо развитым внутренним надмыщелком”.
Ниже приведено собственное описание плечевой кости восточной слепушонки.
Общая длина кости от 15 до 17 мм, с относительно широкими проксимальным (3–4 мм) и дистальным (3–4 мм) эпифизами. Отличительная особенность – отсутствие надмыщелкового отверстия foramen supracondyloideus, которое присутствует у крыс, песчанок и полевок (Фомин, Лобачев, 2000). Также гребень большого бугорка crista tuberculi majoris расположен на диафизе ниже блока проксимального эпифиза и имеет округлые формы. У тех же песчанок и крыс он спускается сразу из-под суставной головки кости caput humeri и имеет вид прямоугольного треугольника. У тушканчиков кончик гребня заметно вздернут.
Морфометрия плечевых костей. Нам удалось выделить два типа плечевых костей, которые отражают закономерности географического распределения восточной слепушонки.
Удлиненность костей характерна для особей из популяций, совершающих активные перемещения. Такие перемещения на значительные расстояния могут быть обусловлены общим увеличением численности в отдельных местах либо неблагоприятными изменениями условий существования (Шубин, 1978). Для популяций из Самарканда это возможность осваивать большие пригодные для обитания территории – например, долину р. Зеравшан в Центральном Узбекистане (Мекленбурцев, 1935).
Укороченные и массивные кости характерны для популяций, которые живут в изолированных водными, песчаными и горными преградами оазисах Бухары и Ташкента.
Морфометрическое сходство частей краниального и посткраниального скелета восточной слепушонки из Бухары и Ташкента мы объясняем не принадлежностью зверьков к одному подвиду, а особенностями приспособления вида к условиям среды обитания.
Наиболее вариабельными признаками являются: общая длина кости и размеры диафиза. Самые крупные и длинные кости описаны для слепушонок из Самарканда; самые короткие кости – для животных из Ташкента и Бухары. Массивность диафиза слабо, но коррелирует с длиной кости (коэффициент корреляции составляет в Самарканде 0.36 в Ташкенте 0.44). Так, кости экземпляров из окрестностей Бухары имеют небольшую длину и самый утолщенный диафиз. Самая неизменная часть плечевой кости – проксимальный эпифиз.
Диапазоны промеров плечевых костей приведены в табл. 3 для сборов из Самарканда, Бухары и Ташкента. Видно, что по всем промерам вариабельность не превышает 10%, но наибольшей является для промеров 4а и 4в, особенно в Ташкенте.
Таблица 3.
Морфометрические характеристики плечевой кости восточной слепушонки (Ellobius tancrei) из крупных городов Узбекистана
| Промеры | Переменная | Среднее | min–max | Коэффициент вариации |
|---|---|---|---|---|
| Самарканд | ||||
| 1.0 | х1 | 16.1 ± 0.06 | 14.9–17.0 | 2.534766 |
| 2.0 | х2 | 3.9 ± 0.03 | 3.3–4.4 | 6.103496 |
| 3.0 | х3 | 4.3 ± 0.05 | 3.0–4.7 | 8.404725 |
| 4.а | х4 | 1.5 ± 0.02 | 1.1–1.8 | 8.599683 |
| 4.в | х5 | 1.5 ± 0.02 | 1.3–1.7 | 7.907782 |
| ∑ 1.0-4.а | х6 | 25.9 ± 0.12 | 22.7–27.1 | 3.336754 |
| ∑ 4.а-4.в | х7 | 2.9 ± 0.03 | 2.4–3.5 | 6.861932 |
| Бухара | ||||
| 1.0 | х1 | 15.4 ± 0.08 | 13.7–16.4 | 3.736198 |
| 2.0 | х2 | 3.8 ± 0.02 | 3.5–4.2 | 4.430291 |
| 3.0 | х3 | 4.1 ± 0.03 | 3.6–4.5 | 5.174419 |
| 4.а | х4 | 1.2 ± 0.01 | 1.1–1.6 | 8.978960 |
| 4.в | х5 | 1.4 ± 0.01 | 1.3–1.8 | 6.874117 |
| ∑ 1.0-4.а | х6 | 24.6 ± 0.13 | 21.9–26.0 | 3.669799 |
| ∑ 4.а-4.в | х7 | 2.7 ± 0.03 | 2.4–3.3 | 6.797188 |
| Ташкент | ||||
| 1.0 | х1 | 15.4 ± 0.07 | 14.0–16.6 | 3.397377 |
| 2.0 | х2 | 3.9 ± 0.02 | 3.6–4.4 | 4.549153 |
| 3.0 | х3 | 4.2 ± 0.03 | 3.6–4.6 | 4.931609 |
| 4.а | х4 | 1.4 ± 0.02 | 1.1–1.8 | 9.772689 |
| 4.в | х5 | 1.4 ± 0.02 | 1.2–1.7 | 9.663935 |
| ∑ 1.0-4.а | х6 | 24.9 ± 0.12 | 23.1–27.1 | 3.412259 |
| ∑ 4.а-4.в | х7 | 2.8 ± 0.03 | 2.4–3.3 | 7.829045 |
Для дискриминантного анализа использовались все 7 параметров плечевой кости (табл. 3), однако в дискриминантное уравнение в качестве переменных включены лишь те параметры, вес которых в уравнении (по значению коэффициента вариации) был наибольшим: это переменные х6 – сумма промеров диафиза 4.а и 4.в, х7 – сумма промеров 1.0-4.а, х3 – ширина дистального эпифиза и х2 – ширина проксимального эпифиза (табл. 4). Таким образом, для “обучающей выборки” плечевых костей восточной слепушонки из Бухары и Самарканда были составлены дискриминантные функции:
Таблица 4.
Статистическая характеристика основных промеров плечевых костей восточной слепушонки (Ellobius tancrei) из Самарканда и Бухары, включенных в дискриминантные уравнения в качестве переменных
| Переменная | Лямбда Уилкса |
Частная лямбда |
F-исключения (1.93) | Уровень значимости | Толерантность значений | Квадрат толерантности |
|---|---|---|---|---|---|---|
| х6 | 0.684385 | 0.793161 | 24.77384 | 0.000003 | 0.270620 | 0.729380 |
| х7 | 0.600372 | 0.904152 | 10.07087 | 0.002030 | 0.748697 | 0.251303 |
| х3 | 0.594819 | 0.912593 | 9.09899 | 0.003281 | 0.352169 | 0.647832 |
| х2 | 0.553948 | 0.979926 | 1.94612 | 0.166258 | 0.574914 | 0.425086 |
Значение дискриминирующих функций для экземпляров из Бухары – DF (Б) = 792.09, для образцов из Самарканда – DF(С) = 545.40.
Приведенные уравнения позволяют уверенно дифференцировать особей “бухарской” (76% из смешанной выборки) и “самаркандской” (84%) группировок из смешанной “обучающей выборки”, что предполагает диагностическую ценность использованных промеров. Общая успешность дискриминации составляет 80%. В будущем эффективность дискриминации будет проверена на независимой выборке.
Статистическая достоверность различий между двумя подвидами по морфометрическим характеристикам плечевой кости подтверждается значениями Р-уровня значимости (табл. 4), где такие признаки, как ширина проксимального эпифиза и промеры диафиза являются слабо варьирующими.
Таким образом, по нашим данным, два подвида восточной слепушонки E. t. ognevi из Бухары и E. t. fusciceps из Самарканда, описанные ранее, имеют достоверные различия как по промерам черепа, в соответствии с их описаниями, так и по характеристикам плечевой кости.
Морфометрические данные необходимо дополнить данными о локализации хромосомных наборов, а также данными по метагеномной и ядерной ДНК для дальнейшего обоснования таксономической ценности тех или иных морфологических показателей.
Список литературы
Воронцов Н.Н., Ляпунова Е.А., Захарян Г.Г., Иванов В.Г., 1969. Кариология и систематика рода Ellobius (Microtinae. Rodentia) // Млекопитающие: эволюция, систематика, кариология, фаунистика. Новосибирск: Наука. С. 127–130.
Воронцов Н.Н., Якименко Л.В., 1984. Морфометрический анализ изменчивости черепа слепушонок (Rodentia, Ellobius) // Зоологический журнал. Т. 63. № 12. С. 1859–1871.
Гашев С.Н., Бетляева Ф.Х., Лупинос М.Ю., 2014. Математические методы в биологии: анализ биологических данных в системе Statistica. Тюмень: изд-во Тюменского государственного университета. 208 с.
Громов И.М., 1963. Род Ellobius Fischer (1814) – Слепушонки // Млекопитающие фауны СССР. Л. С. 553– 562.
Громов И.М., 1983. Введение // История и эволюция современной фауны грызунов (неоген–современность). М.: Наука. С. 3–8.
Громов И.М., Ербаева М.А., 1995. Млекопитающие фауны России и сопредельных территорий. Зайцеобразные и грызуны. СПб. 522 с.
Дукельская Н.М., 1926. Материалы к познанию фауны млекопитающих Средней Азии // Бюллетень Средне-Азиатского гос. ун-та. Ташкент. № 15. С. 67–89.
Мекленбурцев Р.Н., 1935. К экологии и распространению некоторых грызунов равнинной части Зеравшанской долины. Труды Средне-Азиатского гос. ун-та. Ташкент. Вып. 17. Серия VIII-a. Зоология. 30 с.
Митропольский. О.В., 2005. Проведение экологического мониторинга по плечевым костям. Методическое руководство. Бишкек: изд-во ОсОО “Бисмарк”. 43 с.
Митропольский М.Г., Митропольский О.В., 2009. Млекопитающие в питании зимующих ушастых сов в крупных городах Узбекистана // Совы Северной Евразии: экология, пространственное и биотопическое распределение. М. С. 66–69.
Митропольский М.Г., Митропольский О.В., 2016. Использование морфометрических данных плечевых костей млекопитающих в различных аспектах изучения видов // Териофауна России и сопредельных стран. Материалы междунар. сов. Х Съезд Териологического общества при РАН. М.: Товарищество научных изданий КМК. С. 269.
Огнев С.И., 1940. Звери СССР и прилежащих стран. Грызуны. М.-Л.: изд-во Акад. наук СССР. Т. IV. 615 с.
Огнев С.И., 1950. Род Ellobius Fischer (1814) // Звери СССР. М.: изд-во Акад. наук СССР. Т. VII. С. 658–706.
Павленко Т.А., Аллабергенов К., 1974. Грызуны – Rodentia // Позвоночные животные Ферганской долины. Ташкент. С. 142–185.
Потапов Е.Р., 1990. Использование погадок для изучения питания хищных птиц // Методы изучения и охраны хищных птиц. М. С. 103–118.
Топачевский В.А., 1965. Насекомоядные и грызуны Ногайской позднеплиоценовой фауны. Киев: Наукова Думка. С. 97–100.
Угаров А.А., 1928. К систематике рода Ellobius Fischer. Труды Средне-азиатского государственного университета. Ташкент. Серия VIII-a. Зоология. Вып. 4. 20 с.
Фомин С.В., Лобачев В.С., 2000. Надмыщелковое отверстие плечевой кости и его использование в систематике мелких млекопитающих // Систематика и филогения грызунов и зайцеобразных. М. С. 167–170.
Шубин И.Г., 1978. Род Слепушонки – Ellobius Fischer, 1814 // Млекопитающие Казахстана. Алма-Ата. Т. 1. Ч. 3. С. 188–208.
Якименко Л.В., 2003. Цитогенетическая и морфологическая дифференциация в процессе эволюции слепушонок и домовых мышей. Дис. … докт. биол. наук. Владивосток. 309 с.
Corbet G.B., 1978. The Mammals of the Palacarctic region: a taxonomic review. British Museum (Nat. Hist.). London–Ithaka: Cornell Univ. Press. 314 p.
Thomas O., 1909. On Mammals collected in Turkestan by Mr. Douglas Carruthers. The Annals magazine of natural history: zoology, botany and geology. London. V. III. P. 257–266.
Дополнительные материалы отсутствуют.
Инструменты
Зоологический журнал