

Зоологический журнал, 2021, T. 100, № 11, стр. 1312-1317
Связь полового созревания перезимовавших самок Arvicola amphibius (Rodentia, Arvicolinae) с их физическим состоянием
Г. Г. Назарова a, *, М. А. Потапов a, О. Ф. Потапова a, Е. А. Новиков a
a Институт систематики и экологии животных СО РАН
630091 Новосибирск, Россия
* E-mail: galinanazarova@mail.ru
Поступила в редакцию 10.10.2020
После доработки 10.02.2021
Принята к публикации 15.02.2021
Аннотация
В весенний период (третья декада апреля–вторая декада мая) оценивали динамику массы тела, упитанности и клеточного состава вагинального содержимого у перезимовавших самок из природной популяции водяных полевок, обитающей в подтаежной зоне Барабинской низменности. Проанализирована зависимость наступления эструса от даты отлова, массы тела и физической кондиции у отловленных животных. Результаты множественного регрессионного анализа показали, что наступление физиологической готовности самок к спариванию достоверно коррелирует с их упитанностью. Половозрелые особи имеют более высокие значения массы тела и упитанности, чем неполовозрелые.
Возникновение сезонности размножения у млекопитающих связано с регулярно повторяющимися внутригодовыми колебаниями температурного режима, обеспеченности кормом и с высокими энергетическими затратами на размножение. С сезонной динамикой абиотических и биотических параметров среды сопряжены изменения гормонального статуса и величины жировых запасов в организме животных (Штайнлехнер, Пухальский, 1999; Bartness et al., 2002; Bronson, 2009).
У грызунов умеренного пояса северного полушария период размножения приходится на период, когда состояние кормовой базы и защитные условия среды наиболее благоприятны для рождения и выращивания потомства. Такие условия складываются в весеннее-летний сезон года, хотя в теплые и мягкие зимы отмечается и подснежное размножение (Жигальский, 2012). Внешним сигналом, вовлеченным в регуляцию репродуктивной активности, выступает изменение длительности светового периода. Гормон эпифиза мелатонин выполняет центральную роль в осуществлении сезонных ритмов физиологических функций. Секреция мелатонина происходит в темное время суток, повышается осенью-зимой и снижается весной-летом. Мелатонин тормозит частоту и величину пульсирующей секреции гонадотропных гормонов гипоталамуса, подавляя высвобождение гормонов гипофиза, необходимых для инициации и поддержания репродуктивной активности (Weems et al., 2015; Vasantha, 2016; Guh et al., 2019).
Информацию о состоянии внутренней метаболической среды к центрам гипоталамуса, ответственным за комплексную регуляцию аппетита, метаболизма, репродуктивной и других функций, передает гормон жировой ткани лептин (Gueorguiev et al., 2001). В последние десятилетия выяснено, что в регуляции репродуктивной активности в зависимости от сезона года и энергетического баланса организма участвует гормон кисспептин, опосредующий действие многочисленных внешних и внутренних сигналов (Simonneaux et al., 2013; Bohlen et al., 2018).
Половое созревание – ключевая и наиболее пластичная характеристика жизненного цикла (Boonstra, 1989; Rivest, 1991). С плотностно-зависимой регуляцией возраста полового созревания связаны циклические колебания численности (Evsikov et al., 1997; Oli, Dobson, 1999). Выяснение связи между физическим состоянием организма и половым созреванием важно для понимания механизмов контроля численности популяций и путей адаптации животных к климатическим и антропогенным воздействиям.
Водяная полевка (Arvicola amphibius (Linnaeus 1758)) – подходящий модельный объект для выяснения эколого-физиологических механизмов реализации репродуктивного потенциала сезонно размножающихся млекопитающих. Водяная полевка занимает обширный ареал с характерными для разных природных регионов особенностями хода размножения. В подтаежной полосе Барабинской низменности, где численность водяной полевки подвержена тысячекратным колебаниям, сезон размножения продолжается с середины апреля до середины сентября, сокращаясь в годы спада. Половая зрелость у перезимовавших самок наступает на 2–4 недели позже, чем у самцов (Водяная полевка …, 2001). В комплексных эколого-физиологических исследованиях выяснено, что кормовая обеспеченность в зимовочных стациях резко ухудшается после прохождения пика численности, при этом индивидуальная изменчивость по эффективности переваривания и ассимиляции корма коррелирует с поведенческими и репродуктивными характеристиками особей (Евсиков и др., 2001).
Ранее экспериментально показано, что рост и наступление половой зрелости у водяной полевки контролируются фотопериодическими сигналами (Евсиков и др., 1989). Половое созревание молодых особей зависит от календарных сроков рождения и физиологического состояния матери в период беременности (Nazarova, Evsikov, 2007, 2010). До настоящего времени остается не выясненной связь между динамикой физического состояния самок в весенний период и сроками их полового созревания.
Цель работы – установить, зависит ли половое созревание перезимовавших самок от даты их отлова, массы тела и упитанности в весенний период. Для выяснения этого вопроса с третьей декады апреля по вторую декаду мая были оценены размерные характеристики самок из природной популяции и их связь с наступлением физиологической готовности к спариванию, оцениваемой по клеточному составу вагинальных мазков.
МАТЕРИАЛ И МЕТОДИКА
Исследование выполнено в подтаежной зоне Барабинской низменности в окрестностях стационара, расположенного в д. Лисьи Норки Новосибирской обл. (55°50′ с.ш., 80°00′ в.д.) с 24 апреля по 12 мая 1982 г. В период сезона размножения наблюдался резкий спад численности, начавшийся в 1981 г. (Водяная полевка …, 2001). Отлов водяных полевок проводили с помощью живоловушек в стациях размножения, по берегам рек Омь и Ангут. Всего было добыто 77 перезимовавших самок. В день отлова у самок исследовали вагинальный мазок, измеряли массу и длину тела (от ануса до кончика носа).
Упитанность определяли по отклонению массы тела от теоретически ожидаемой для наблюдаемой длины. Теоретически ожидаемую находили, используя уравнение регрессии, полученное для всей выборки (Jakob et al., 1996). Данный показатель имеет несколько преимуществ по сравнению с другими морфометрическими характеристиками и индексами (масса тела, отношение массы к длине и др.): 1) исключено влияние линейных размеров; 2) легко интерпретируется: положительный знак указывает на высокую упитанность, отрицательный знак – на низкую; 3) распределение признака соответствует нормальному. Надежность используемого индекса подтверждают результаты прямых измерений содержания жира в организме. У мышей коэффициент корреляции между индексом физического состояния и абсолютным содержанием жира равен 0.76 (Reed et al., 2003). У водяных полевок 0.63 (Назарова, 2008).
Эстральную стадию самок оценивали в баллах, отражающих приблизительную пропорцию ороговевших чешуй в вагинальном мазке (0, 25, 75, 100), поскольку у водяных полевок, как и других видов полевок с индуцированным типом овуляции (Sawrey, Dewsbury, 1985), нет регулярной смены цитологической картины мазка. К половозрелым, т.е. физиологически готовым к спариванию, относили особей, у которых ороговевшие чешуи численно доминировали над другими типами клеток (баллы 75 и 100) (Назарова и др., 2007).
Статистическую обработку результатов проводили с помощью пакета программ “Statistica 6.1”. Использовали однофакторный дисперсионный анализ, корреляционный и множественный регрессионный анализ. Уровень статистической значимости принят p < 0.05.
РЕЗУЛЬТАТЫ
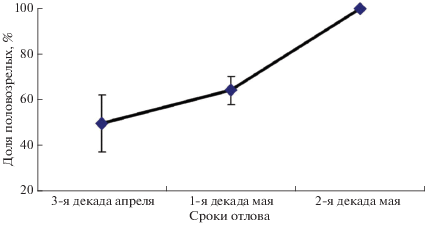
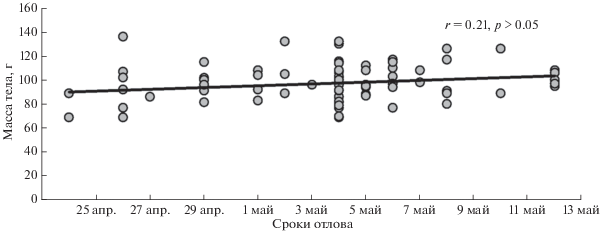
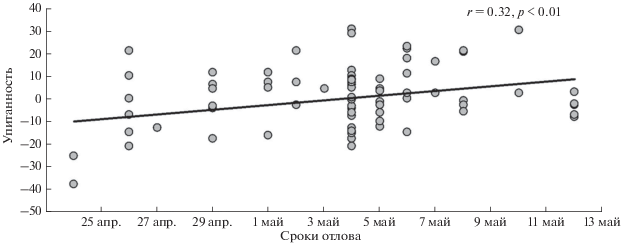
В последней декаде апреля 50% самок, судя по преобладанию ороговевших чешуй над другими клетками вагинального содержимого, были физиологически готовы к спариванию. Во второй декаде мая половой зрелости достигли все перезимовавшие самки (рис. 1). За весь период у четырех самок в мазках обнаружены сперматозоиды. Масса тела самок имела тенденцию к увеличению в течение периода исследования (рис. 2). Повышение упитанности было статистически значимым (рис. 3).
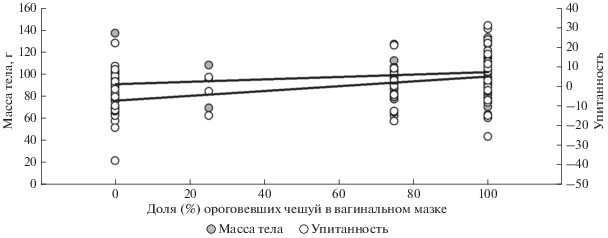
Эстральная стадия связана с физическими кондициями самок, о чем свидетельствуют достоверные корреляции процента ороговевших чешуй в вагинальном мазке с массой тела (r = 0.33, p < 0.01) и упитанностью (r = 0.39, p < 0.001), рис. 4.
Рис. 4.
Корреляция доли (%) ороговевших эпителиальных чешуй в вагинальном мазке (эстральной стадии) с массой тела и упитанностью.
Согласно результатам однофакторного дисперсионного анализа, половозрелые и неполовозрелые самки различались по массе тела (F1,75 = = 8.08, p < 0.01) и упитанности (F1,75 = 11.43, p = = 0.001) (табл. 1).
Таблица 1.
Размерные характеристики и упитанность самок разного репродуктивного статуса
| Репродуктивный статус | Число животных | Масса тела, г | Длина тела, мм | Упитанность |
|---|---|---|---|---|
| Неполовозрелые | 29 | 90.9 ± 2.8 | 147.5 ± 1.8 | –6.6 ± 2.4 |
| Половозрелые | 48 | 101.0 ± 2.1 | 146.9 ± 1.3 | 4.0 ± 1.9 |
Результаты множественного регрессионного анализа с пошаговым включением влияющих переменных – дата отлова, масса, длина тела и индекс физического состояния – показали, что процент ороговевших чешуй в вагинальном мазке зависит только от упитанности самок (F1,75 = 13.34, p < 0.001). Уравнение регрессии имеет вид:
Y = 58.8 + 1.2 * “упитанность”.
ОБСУЖДЕНИЕ
Таким образом, результаты выполненного исследования показали, что физические кондиции и состояние репродуктивной функции самок находятся под общим контролем сезонных факторов. Продолжительность светового периода и направление его изменения – достаточно надежный сигнал о происходящей внутригодовой трансформации климатических условий, трофической базы и, следовательно, наличия потенциальной возможности произвести жизнеспособное потомство (Штайнлехнер, Пухальский, 1999).
Половое созревание перезимовавших самок завершается к окончанию первой декады мая и связано с повышением их упитанности. В целом, половозрелые особи имеют более высокие значения массы тела и упитанности, чем неполовозрелые. Тесную взаимосвязь полового созревания и роста животных подтверждают результаты многих генетико-селекционных и популяционных исследований (Шварц, 1980; Bronson, 1984). Отбор мышей по скорости роста в первые 10 дней жизни приводит к однонаправленному изменению темпов полового созревания самок, причем обнаруживается пороговый эффект массы тела (Ernst et al., 1999).
Масса тела, величина жировых запасов отражают энергетический статус особей и являются показателями, скоррелированными со скоростью полового развития (Rivest, 1991). Связь между физическим состоянием и репродуктивными параметрами наиболее отчетливо прослеживается у зимоспящих видов. У сусликов Белдинга, например, низкая масса тела при выходе из спячки негативно влияет на активацию репродуктивной функции, вследствие чего многие особи не вступают в размножение (Bushberg, Holmes, 1985).
Полевые эксперименты на мелких млекопитающих, имитирующие улучшение состояния кормовой базы в зимний период, показали, что количество и качество пищи играют важную роль в инициации процессов воспроизводства и регуляции численности (McAdam, Millar, 1999; Banks, Dickman, 2000). С межпопуляционными различиями кормовой обеспеченности связана вариабельность скорости роста и полового созревания у водяной полевки (Moorhouse et al., 2008).
ЗАКЛЮЧЕНИЕ
Результаты популяционного исследования, выполненного на водяной полевке, подтверждают, что для инициации полового созревания перезимовавших самок необходим определенный запас жировых резервов в организме, который может быть создан при наличии достаточного количества трофических ресурсов во внешней среде. Половозрелые особи имеют более высокие значения массы тела и упитанности, чем неполовозрелые. Факторы, контролирующие динамику упитанности особей в весенний период, могут оказывать влияние на сроки начала размножения и его интенсивность у водяной полевки.
Список литературы
Водяная полевка: Образ вида. Сер. “Виды фауны России и сопредельных стран”, 2001. М.: Наука. 527 с.
Евсиков В.И., Скорова С.В., Назарова Г.Г., Мошкин М.П., 1989. Влияние фотопериода на рост и репродуктивную функцию водяной полевки (Arvicola terrestris L.) // Экология. № 6. С. 58–63.
Евсиков В.И., Герлинская Л.А., Мошкин М.П., Музыка В.Ю., Назарова Г.Г. и др., 2001. Генетико-физиологические основы популяционного гомеостаза // Водяная полевка. Сер. “Виды фауны России и сопредельных стран”. Пантелеев П.А. (Ред.). М.: Наука. С. 386–412.
Жигальский О.А., 2012. Динамика численности и структуры населения рыжей полевки (Myodes (Clethrionomys) glareolus) при зимнем и весеннем начале размножения // Зоологический журнал. Т. 91. № 5. С. 61–628.
Назарова Г.Г., Потапов М.А., Евсиков В.И., 2007. Вероятность наступления эструса и спаривания у водяной полевки, Arvicola terrestris L., зависят от физического состояния самок, полового опыта и поведения брачных партнеров // Зоологический журнал. Т. 86. № 12. С. 1507–1512.
Назарова Г.Г., 2008. Материнское влияние на приспособленность потомков и численность популяции. Дис. … докт. биол. наук. Новосибирск: Институт систематики и экологии животных СО РАН. 262 с.
Шварц С.С., 1980. Экологические закономерности эволюции. М.: Наука. 278 с.
Штайнлехнер С., Пухальский В., 1999. Сезонная регуляция размножения мелких млекопитающих // Сибирский экологический журнал. № 1. С. 23–35.
Banks, P.B., Dickman C.R., 2000. Effects of winter food supplementation on reproduction, body mass, and numbers of small mammals in montane Australia // Canadian Journal of Zoology. V. 78. P. 1775–1783.
Bartness T.J., Demas G.E., Song C.K., 2002. Seasonal changes in adiposity: the roles of the photoperiod, melatonin and other hormones, and sympathetic nervous system // Experimental Biology and Medicine. V. 227. P. 363–376.
Bohlen T.M., Silveira M.A., Buonfiglio D.D.C., Ferreira-Neto H.C., Cipolla-Neto J. et al., 2018. A short-day photoperiod delays the timing of puberty in female mice via changes in the Kisspeptin system // Frontiers in Endocrinology. V. 9. Article 44.
Boonstra R., 1989. Life history variation in maturation in fluctuating meadow vole populations (Microtus pennsylvanicus) // Oikos. V. 54. P. 265–274.
Bronson F.H., 1984. Energy allocation and reproduction development in wild and domestic house mouse // Biology of Reproduction. V. 31. P. 83–88.
Bronson F.H., 2009. Climate change and seasonal reproduction in mammals // Philosophical Transactions of the Royal Society. B. V. 364. P. 3331–3340.
Bushberg D.M., Holmes W.G., 1985. Sexual maturation in male Belding’s ground squirrels. Influence of body weight // Biology of Reproduction. V. 33. P. 302–308.
Ernst C.A., Crenshaw P.D., Atchley W.R., 1999. Effect of selection for development rate on reproductive onset in female mice // Genetics Research. V. 74. P. 54–64.
Evsikov V.I., Nazarova G.G., Potapov M.A., 1997. Genetic-ecological monitoring of a cyclic population of water voles Arvicola terrestris L. in the South of Western Siberia // Russian Journal of Genetics. V. 33. № 8. P. 963–972.
Gueorguiev M., Gyth M.I., Korbonits M., 2001. Leptin and puberty: a review // Pituitary. V. 4. P. 79–86.
Guh Y.-J., Tamai T.K., Yoshimura T., 2019. The underlying mechanisms of vertebrate seasonal reproduction // The Proceedings of the Japan Academy, Series B. V. 95. № 7. P. 343–357.
Jakob E.M., Marshall S.D., Uetz G.W., 1996. Estimating fitness: a comparison of body condition indices // Oikos. V. 77. P. 61–67.
McAdam A.G., Millar J.S., 1999. Dietary protein constraint on age at maturity: an experimental test with wild deer mice // Journal of Animal Ecology. V. 68. P. 733–740.
Moorhouse T.P., Gelling M., Macdonald D.W., 2008. Effects of forage availability on growth and maturation rates in water voles // Journal of Animal Ecology. V. 77. P. 1288–1295.
Nazarova G.G., Evsikov V.I., 2007. Sexual maturation of daughters depends on mother’s body condition during pregnancy: an example of the water vole, Arvicola terrestris L. // Doclady Biological Science. V. 42. P. 53–55.
Nazarova G.G., Evsikov V.I., 2010. Physical state of mother at the period of pregnancy and sexual maturation of descendants-sons of the water vole (Arvicola terrestris L.) // Journal of Evolutionary Biochemistry and Physiology. V. 46. № 2. P. 173–178.
Oli M.K., Dobson F.S., 1999. Population cycles in small mammals: the role of age at sexual maturity // Oikos. V. 86. № 3. P. 557–565.
Reed D.R., Li X., McDaniel A.H., Lu K., Li S. et al., 2003. Loci on Chromosomes 2, 4, 9, and 16 for body weight, body length, and adiposity identified in a genome scan of an F2 intercross between the 129P3/J and C57BL/ 6ByJ mouse strains // Mammalian Genome. V. 14. P. 302–313.
Rivest R.W., 1991. Sexual maturation in female rats: hereditary, devepopmental nd environmental aspect // Experimenta. V. 47. P. 1026–1039.
Simonneaux V., Ancel C., Poirel V.J., Gauer F., 2013. Kisspeptins and RFRP-3 act in concert to synchronize rodent reproduction with seasons // Frontiers in Neuroscience. V. 7. Article 22.
Sawrey D.K., Dewsbury D.D., 1985. Control of ovulation, vaginal estrus, and behavioral receptivity in voles (Microtus) // Neuroscience & Biobehavioral Reviews. V. 9. P. 563–571.
Vasantha I., 2016. Physiology of seasonal breeding: a review // The Journal of Veterinary Science & Technology. V. 7. № 3. P. 1–4.
Weems P.W., Goodman R.L., Lehman M.N., 2015. Neural mechanisms controlling seasonal reproduction: principles derived from the sheep model and its comparison with hamsters // Frontiers in Neuroendocrinology. V. 37. P. 43–51.
Дополнительные материалы отсутствуют.
Инструменты
Зоологический журнал