

Зоологический журнал, 2021, T. 100, № 12, стр. 1323-1327
К вопросу о статусе семейства Aididae (Lepidoptera, Zygaenoidea)
Ю. С. Волкова a, *, В. В. Золотухин b
a Ульяновский государственный университет
432063 Ульяновск, Россия
b Ульяновский государственный педагогический университет имени И.Н. Ульянова
432700 Ульяновск, Россия
* E-mail: beeme7@mail.ru
Поступила в редакцию 15.05.2019
После доработки 03.08.2019
Принята к публикации 06.08.2021
Аннотация
Исследование функциональной мускулатуры копулятивного аппарата Aididae выявило наличие у них 8 пар мышц – m1, m8(3), m5(7), m6(5), m7(6), m9, m2(10), а также наличие внутренних мышц анального конуса m20 и внутренней непарной продольной мышцы эдеагуса m21, что поддерживает предположение о принадлежности семейства Аididae в ранге подсемейства к Limacodidae. К ранее признанным аутапоморфиям айдид добавлены срастание и редукция ряда генитальных структур, асимметрия ретракторов эдеагуса, а также наличие мощных пучков мышц в транстиллярной зоне.
Aididae (Lepidoptera, Zygaenoidea) – небольшое неотропическое семейство макрочешуекрылых, относящееся к лимакоидной группе надсемейства Zygaenoidea. Согласно последним данным, семейство насчитывает всего 2 рода и 6 видов (Heppner, 1994; Nieukerken et al., 2011). Статус семейства данной группе чешуекрылых был присвоен Шаусом (Schaus, 1906), но в дальнейшем неоднократно оспаривался. В частности, высказывалась точка зрения, что Aididae – это подсемейство Megalopygidae (Fletcher, Nye, 1982; Heppner, 1994). Возвратил им статус самостоятельного семейства Эпштейн (Epstein, 1996; Epstein et al., 1998), который на основании изучения преимагинальных стадий и филогенетического анализа отметил также их близость с Limacodidae и Dalceridae. Отличия Aididae от указанных семейств, по его мнению, заключаются в наличии хетоземы у имаго, отсутствии гнатоса в гениталиях самцов, наличии продольных бороздок на мезонотуме и обширной склеротизации передней части пронотума; от Limacodidae, кроме того, они отличаются наличием крупных придаточных желез в гениталиях самок и глубоко выемчатыми анальными сосочками.
Последний признак, впрочем, нашими исследованиями не подтверждается – анальные сосочки в гениталиях самок Aididae срастаются, формируя цельную подушковидную структуру (рис. 1, 2).
Рис. 1.
Aidos amanda (Stoll 1782): 1 – жилкование крыльев (область склеротизации затемнена), 2 – гениталии самки.
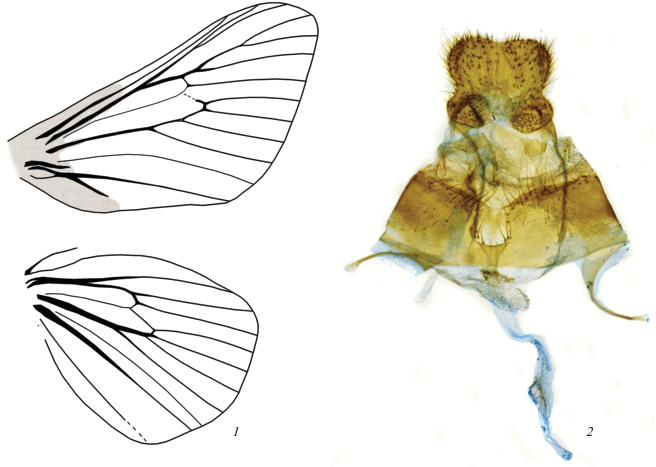
В качестве аутапоморфий семейства Эпштейном названы бесформенная вершина прядильных сосочков гусениц, наличие 1–2 отверстий в наружном слое кокона, имитирующих выходные отверстия паразитических ос, а также наличие анастомоза жилок R1 и R2 переднего крыла (рис. 1, 1) и особенности строения антенн самцов, в которых примерно две трети базального участка двугребенчатые.
Перечисленные признаки позволяют трактовать систематический статус Aididae двояко, как в качестве самостоятельного семейства, так и в качестве подсемейства в составе собственно Limacodidae. Впервые четкое предположение о том, что данная группа может включаться в Limacodidae, было высказано Скобле (Scoble, 1992), и доводом в его пользу стало довольно размытое заключение Холловэя – “Aidinae possibly belong to Limacodidae on the basis of forewing venation and male and female genitalia” (Holloway, 1986).
МАТЕРИАЛЫ И МЕТОДИКА
Для решения вопроса о статусе Aididae нами изучена мускулатура генитальных придатков самцов. В ходе исследования исследованы зафиксированные в спирте экземпляры представителей обоих известных родов семейства: Aidos Hübner [1820] и Brachycodilla Dyar 1898 – Aidos amanda (Stoll 1782) и Brachycodilla carmen (Schaus 1892), собранные В.В. Синяевым в Эквадоре в 2016 г. Препараты были изготовлены по стандартной методике с прокрашиванием мышц водным раствором эозина натрия, делающим их хорошо различимыми благодаря стойкой ярко-розовой окраске (Волкова и др., 2017). Номенклатура мышц дана по: Стекольников (2008).
РЕЗУЛЬТАТЫ
Общая характеристика генитального аппарата Aididae
Гениталии самцов семейства устроены однотипно у всех видов семейства. Ункус вытянутый, языковидный, базально бутылковидно расширенный, с небольшим вершинным крюком. Гнатос редуцирован и функционально замещен небольшим, обычно шиповатым склеритом, образующимся из разросшейся субанальной пластины. Соции редуцированы до небольших щетинистых латеро-вентральных участков ункуса. Вальвы широкие, с обширной мембранозной зоной, отделяющей вздутый саккулус с крюковидными вершинными придатками, всегда небольшие, уплотненные, сильно склеротизованные и часто с выраженным бугристым рельефом. Винкулум лентовидный; саккус слабо выражен. Юкста не развита, и эдеагус поддерживается срастающимися антеро-дорсальными краями саккулусов. Эдеагус небольшой, в 1.5 раза короче вальвы, узкий, кинжаловидный, со слабо разрастающимися боковыми сторонами основания, несколько асимметричный, провернутый по своей оси на 180°, из-за чего везика (здесь она простая, мешковидная, лишена корнутусов) располагается строго вентрально.
В гениталиях самок анальные сосочки сливаются в основании с образованием плоской подушки, состоящей из двух долей – более крупной каудальной и небольшой выпуклой базальной; вся их поверхность густо покрыта щетинками. Передние апофизы в полтора раза длиннее задних. Остиум слабо склеротизован, открывается в центре 8-го стернита. Вагинальные пластинки отсутствуют. Дуктус простой, мембранозный. Бурса округлая, мембранозная, без участков склеротизации.
Генерализованный тип строения гениталий самцов предполагает наличие богатого набора мышц. Нами найдено 8 парных мышц (m1, m8(3) m5(7), m6(5), m7(6), m9, m2(10)), а также мышцы анального конуса m20 и внутренние продольные мышцы эдеагуса m21. Различий в числе и топографии мышц генитального аппарата между обоими исследованными родами не найдено, поэтому описание мускулатуры приводится здесь только для Aidos amanda (Stoll 1782), являющегося типовым видом номинативного для айдид рода Aidos Hübner [1820].
Aidos amanda (Stoll 1782)
3♂♂, Эквадор (без указания точного локалитета), июнь–июль 2016, В. Синяев.
Мышцы генитального аппарата расположены симметрично (кроме фаллических мышц), и их топография в целом соответствует таковой пестрянкообразных чешуекрылых надсемейства Zygaenoidea. Однако модификация субанальной пластинки влечет за собой видоизменение топологии тергальных мышц, что отличает оба рода семейства от родственных таксонов.
Депрессоры ункуса m1 хорошо развиты (рис. 2, 1, 2), идут двумя широкими плоскими лентами от краниального края тегумена по всей его внутренней поверхности к основанию клювовидной головчатой части ункуса, прикрепляясь вентрально в основании щетинистых полей соций. При их сокращении происходят опускание ункуса и его фиксация в согнутом состоянии в генитальном аппарате самки. Анальный конус слабо развит; изнутри он выстлан продольными пучками коротких мышц, вероятно m20, способствующими его втягиванию. Ретракторы анального конуса m2(10) сильно редуцированы, однако их присутствие – существенное отличие от представителей семейства Megalopygidae, у которых данные мышцы не найдены (Волкова и др., 2017).
Рис. 2.
Схема топографии мышц генитального аппарата самца Aidos amanda (Stoll 1782): 1 – латеральная проекция; 2 – вентральная проекция; 3 – схема расположения мышц эдеагуса, вентральная проекция.
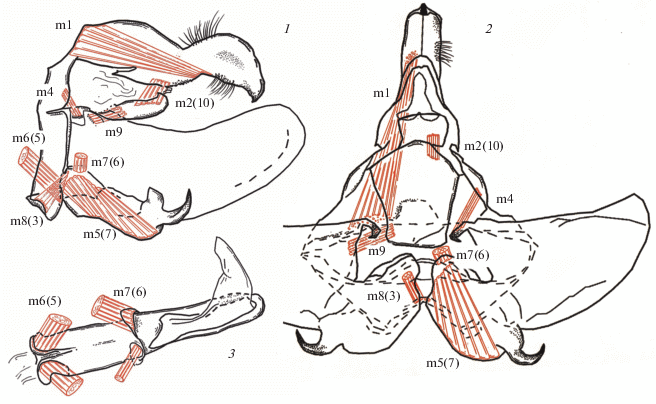
В тергальную группу мышц гениталий Aididae входят также мощная m2(10), связывающая бока каудальной части субанальной пластинки с латеральными полями тегумена, и короткая m9, соединяющая основание анальной пластинки с основанием транстиллярных отростков вальв. При этом m9 выступает, вероятно, в роли ретрактора анального конуса, а m2(10) при своем сокращении фиксирует вершину тегумена, обеспечивая ункус дополнительной опорой и делая фиксацию полов более эффективной.
Базальные отростки вальв являются местом крепления еще одной пары мышц – коротких, но мощных m4, начинающихся на основании тергита. При своем сокращении они, вероятно, обеспечивают более эффективную фиксацию при спаривании.
Протракторы эдеагуса m6(5) берут начало на расширенных и уплощенных отростках его основания с вентральной стороны (рис. 2, 1). В отличие от мышц этого комплекса у Megalopygidae, в гениталиях Aididae они демонстрируют сходство с мышцами Limacodidae – их топография симметрична и оба пучка имеют равную толщину (рис. 2, 3). Протракторы эдеагуса идут ровными, не расширяющимися лентами к каудальному краю винкулума, заходя отдельными пучками в основание наружной поверхности саккулярной зоны вальв. При своем сокращении эти мышцы поднимают вершину эдеагуса, что, вероятно, важно с учетом вентрального расположения везики. Ретракторы эдеагуса m7(6) узкие, очень короткие и четко асимметричные; они прикрепляются к основанию вентральных гребневидных отростков тела эдеагуса, которые представляют, вероятно, сильно редуцированную юксту, и далее соединяют эдеагус с краниальной зоной срастания саккулярных придатков. Именно эта зона и выполняет здесь роль юксты, поддерживая эдеагус. При этом левый пучок m7(6) практически в два раза толще правого; это, а также асимметричное расположение отверстия семяизвергательного канала и вентральное положение везики свидетельствуют о спиральном перекручивании трубки эдеагуса в ходе эволюции, что повлекло за собой морфологические изменения в строении как его мускулатуры, так и соответствующих придатков каудальной зоны.
Интравальварные мышцы m5(7) представлены широкими пучками, отходящими от основания саккулуса к основанию его крючковидного отростка, и обеспечивают сведение вальв при своем сокращении. С основаниями вальв связаны короткие и широкие мышцы m8(3), берущие свое начало на внутренней стороне лентовидного саккуса и отводящие вальвы при своем сокращении.
Внутренняя пара мышц эдеагуса m21 проходит продольными лентами по всей его длине, от основания к везике, втягивая ее при своем сокращении.
ОБСУЖДЕНИЕ
Таким образом, в генитальном аппарате Aididae найдено восемь пар мышц: m1 – мышцы ункуса; m2(10), m9 – мышцы анального конуса и тергальная группа мышц; m8(3) – абдукторы вальв; m5(7) – интравальварные мышцы; m6(5) – протракторы эдеагуса; m7(6) – ретракторы эдеагуса; m20 – внутренняя мышца анального конуса, а также непарная мышца эдеагуса m21. Этот набор мышц соответствует основному плану надсемейства Zygaenoidea и вполне сопоставим с таковым собственно слизневидок – Limacodidae (Кузнецов, Стекольников, 2001; Стекольников, 2008).
Особенности топографии мышц тергального комплекса подтверждают, что гнатосоподобные образования у Aididae – на самом деле действительно лишь разрастающаяся и несколько модифицированная субанальная пластинка, которая внешне выглядит как укороченный гнатос. Таким образом, можно подтвердить редукцию гнатоса, отмеченную для Aididae (Epstein, 1996) и считать эту модификацию одной из аутапоморфий группы.
Вместе с тем ни комплекс морфологических признаков имаго, ни строение преимагинальных стадий развития не имеют сколь-нибудь существенных характеристик, четко придающих айдидам статуса семейства. Гусеницы айдид, в целом, слабо отличаются от гусениц Limacodidae и, как отмечает Эпштейн, их хетотаксия во многом совпадает с таковой слизневидок (Epstein, 1996: 77). Существует еще ряд сходных черт, а именно отсутствие бородавок субдорсальной группы и дорсальное перемещение дыхалец у гусениц Aididae, отмеченное и у некоторых родов Limacodidae (Parasa Moore 1859, Natada Walker 1855, Doratifera Duncan [&Westwood] 1841). Следует также отметить, что прикрытая голова и редукция ложноножек явно свидетельствуют о родственных связях между обеими группами.
В качестве признаков, отличающих Aididae от Megalopygidae, можно отметить строение антенн (у Megalopygidae антенны двугребенчатые или нитевидные по всей длине), полную редукцию ротового аппарата, представленного трехчлениковыми лабиальными пальпами, в то время как у Megalopygidae происходит либо редукция лабиальных пальп и всего ротового аппарата в целом, либо срастание части пальп в основании. Также стоит отметить особенности строения гениталий самцов и самок – для Megalopygidae, в отличие от Aididae, характерны пумилизация частей генитального аппарата самца на фоне гипертрофии эдеагуса, часто несущего одиночный или множественные корнутусы (Волкова и др., 2017), редукция саккулуса и значительное преобразование латеральных придатков винкулума (Волкова, 2019), а также наличие округлых анальных сосочков c оттянутой треугольной вершиной и отсутствие дополнительных областей склеротизации в гениталиях самок.
Таким образом, Aididae не имеют признаков, заметно отличающих их от Limacodidae, и должны рассматриваться лишь в качестве подсемейства последних. Этот вывод поддерживается такими признаками, как форма анальных сосочков самок, характер строения лабиальных пальп, а также расположение жилок Sc и R на заднем крыле, анастомозирующих в своем основании и отходящих общим стволом от дискальной ячейки. Данные признаки были ранее указаны в качестве синапоморфии для комплекса Aididae + Limacodidae + Dalceridae (Epstein, 1996). Аутапоморфиями собственно Aididae можно считать модификацию субанальной пластинки, редукцию юксты и гнатоса и специфическую перфорацию кокона. Еще один крайне характерный признак неясного таксономического значения отмечен нами для жилкования переднего крыла; это склеротизация основания анального поля и отхождение ослабленной A1 от внешнего края этого поля, хотя базальная часть этой жилки сохраняется в склеротизованном поле как толстая приподнятая складка, не сливающаяся с основным стволом А1.
Подсемейство Aidinae Schaus 1906 stat. n. по целому ряду признаков представляется достаточно продвинутым. Поскольку у собственно Limacodidae имеется, как правило, развитый непарный гнатос, а также ряд других генитальных структур, отсутствующих у Aidinae, можно предположить, что их редукция у последних вторична и может рассматриваться как аутапоморфия. Наличие мощных пучков мышц в транстиллярной зоне – также характерная особенность подсемейства.
В ходе дальнейших исследований нами планируется детальное изучение мускулатуры генитальных придатков самцов представителей семейств Dalceridae и Limacodidae Нового Света с целью уточнения списка аутапоморфий и таксономического статуса семейств, входящих в состав лимакоидного комплекса.
Список литературы
Волкова Ю.С., 2019. Wittinia gen. n. (Lepidoptera, Megalopygidae) – новый род чешуекрылых Неотропики // Зоологический журнал. Т. 98. № 1. С. 43–47.
Волкова Ю.С., Золотухин В.В., Куршаков П.А., 2017. Мускулатура генитальных придатков мегалопигид и ее значение в систематике семейства (Lepidoptera, Megalopygidae) // Зоологический журнал. Т. 96. № 10. С. 1213–1220.
Кузнецов В.И., Стекольников А.А., 2001. Новые подходы к системе чешуекрылых мировой фауны (на основе функциональной морфологии брюшка) // Труды Зоологического ин-та РАН. Т. 282. 462 с.
Стекольников А.А., 2008. Морфологические принципы эволюции мускулатуры насекомых. Санкт-Петербург: изд-во СПбГУ. 180 с.
Epstein M.E., 1996. Revision and phylogeny of the limacodid-group families, with evolutionary studies on slug caterpillars (Lepidoptera: Zygaenoidea) // Smithsonian contributions to zoology. № 582. P. 1–102.
Epstein M.E., Geertsema H., Naumann C.M., Tarmann G.M., 1998. The Zygaenoidea // Handbook of Zoology. Kristensen P. (ed.). V. IV. Arthropoda: Insecta. Part 35. P. 159–175.
Fletcher D.S., Nye I.W.B., 1982. Generic Names of Moths of the World: Bombycoidea, Mimallonoidea, Castnioidea, Sesioidea, Cossoidea, Sphingoidea, Zygaenoidea. V. 4. Rudolph William Sabbott Natural History Books. 206 p.
Heppner J.B., 1994. Atlas of Neotropical Lepidoptera, Checklist. Part 4B. Drepanoidea–Bombycoidea–Sphingoidea. Association for Tropical Lepidoptera. IV. Florida: Scientific Publishers Gainesville. 87 p.
Holloway J.D., 1986. The Moths of Borneo: Key to Families: Families Cossidae, Metarbelidae, Ratardidae, Dudgeonidae, Epipyropidae and Limacodidae // The Malayan Nature Journal. V. 40. P. 1–166.
Nieukerken E.J., Kaila L., Kitching I.J., Kristensen N.P., Lees D.C., et al., 2011. Order Lepidoptera Linnaeus, 1758. Zhang, Z.-Q. (Ed.) Animal biodiversity: An outline of higher-level classification and survey of taxonomic richness // Zootaxa. V. 3148. № 1. P. 212–221.
Schaus W., 1906. Descriptions of the new South American moths // Proceedings of the United States National Museum. V. 29. P. 179–346.
Scoble M.J., 1992. The Lepidoptera: Form, Function and Diversity. Oxford: Oxford University Press. 404 p.
Дополнительные материалы отсутствуют.
Инструменты
Зоологический журнал