

Зоологический журнал, 2021, T. 100, № 2, стр. 209-219
Биология и экология чечетки (Acanthis flammea sensu lato, Passeriformes, Fringillidae) на Ямале и в Приобской лесотундре
В. Н. Рыжановский a, *, В. К. Рябицев a, **
a Институт экологии растений и животных УрО РАН
620144 Екатеринбург, Россия
* E-mail: ryzhanovskiy@yandex.ru
** E-mail: riabits@yandex.ru
Поступила в редакцию 15.09.2019
После доработки 12.01.2020
Принята к публикации 04.02.2020
Аннотация
Рассмотрены особенности биологии, экологии и годового цикла чечетки в Приобской лесотундре и субарктических тундрах Ямала: распространение, местообитания, плотность гнездования, сроки и характер весенней миграции, предгнездовой период. Дается характеристика размножения: сроки гнездования на разных широтах, гнездостроение, расположение и устройство гнезд, гнездовой материал, порядок откладки яиц, размер кладки, длительность насиживания и выкармливания птенцов, вторые и третьи кладки. Описание послегнездового периода содержит сведения о длительности пребывания в окрестностях гнезда и переход к кочевкам, ход линьки молодых и взрослых при естественном освещении и в экспериментах с искусственным фотопериодом. Исследовали миграционное состояние и ход осенней миграции, влияние на нее условий среды, переход к зимним кочевкам. Подтверждено отсутствие у чечеток привязанности к местам гнездования и рождения.
В настоящее время для пространства Северной Евразии на основе внешних признаков, прежде всего – окраски оперения, выделяют обыкновенную чечетку (Acanthis flammea L.) с подвидами A. cabaret и A. f. flammea и тундряную (пепельную) чечетку (Acanthis hornemanni Holboell) (Cramp, Perrins, 1994; Коблик и др., 2006). Другие авторы объединяют их в один вид с рядом подвидов (Портенко, 1939). В нашем районе, на севере Западной Сибири, встречаются обе формы. Мы поймали и осмотрели свыше 6000 чечеток, прилетающих в Нижнее Приобье, десятки гнездящихся пар в лесотундре Нижнего Приобья, кустарниковых тундрах Южного и Среднего Ямала. И пришли к выводу о непрерывном ряде изменчивости птиц по таким внешним признакам как размеры тела, длина крыла, окраска оперения – от крупных темных var. A. f. holboellii Brehm (Портенко, 1939) до мелких светлых A. f. exilipes/A. hornemanni (Данилов и др., 1984; Алексеева, 1988; Рябицев, 2008, 2014).
Многие полевые орнитологи на практике вышли из проблемы “неопределимости” большей или меньшей части встреченных птиц обсуждаемой группы путем объединения всех встречающихся чечеток под одним названием – обыкновенная чечетка, или просто чечетка (например, Головатин, Пасхальный, 2005; Минеев, Минеев, 2012). Другие делят их по подвидам, но осторожно: “…все встреченные птицы [Анадыря], скорее всего, принадлежали к подвиду A. f. exilipes…” (Кречмар и др., 1991, с. 248). Или, выделяя пепельную чечетку как отдельный вид, отмечают, что “…многие птицы обладали переходными признаками, и таких было много больше, чем “чистых” тундряных чечеток” (Рогачева и др., 2008, с. 636).
В предлагаемой статье не обсуждается таксономический статус чечеток. Мы называем их обыкновенными чечетками, как в нашей монографии “Птицы Ямала” (Данилов и др., 1984) и в более поздних публикациях. Мы поставили перед собой задачу обобщить результаты многолетнего изучения чечеток Acanthis flammea sensu lato в лесотундре и тундрах Ямала, чтобы познакомить коллег с характеристиками биологии и экологии этих массовых птиц севера Западной Сибири.
МАТЕРИАЛ И МЕТОДЫ
Статья основана на результатах полевых наблюдений в Нижнем Приобье и на п-ове Ямал; на материалах, полученных при обработке птиц, пойманных паутинными сетями и ловушками (свыше 6000 птиц), на результатах поиска и обследования гнезд (свыше 750), на результатах экспериментальных исследований – выкармливания птенцов и последующей передержки при разных фотопериодических условиях. Полевыми наблюдениями охвачена территория (рис. 1) от широты г. Лабытнанги (66°40′ с.ш) в Нижнем Приобье и долины р. Собь на Полярном Урале (67°00′ с.ш.) до широты фактории Тамбей (71°30′ с.ш.) на Северном Ямале в период с 1971 по 1995 и 2006 гг. Отловы птиц проводили в среднем течении р. Собь в 1976–1978 гг., в долине Нижней Оби (стационар Октябрьский 66°42′ с.ш.) в 1978–1990 гг. и 2002–2017 гг. Птиц отлавливали паутинными сетями, начиная с 1976 г. по настоящее время, а в 1979–1988 гг. на берегу протоки Выл-Посл стояла большая ловушка “рыбачинского” типа, обращенная входом на юг весной, на север – во второй половине лета. У отловленных птиц регистрировали вес, упитанность, пол, возраст, длину крыла, описывали состояние оперения по известной методике (Носков, Рымкевич, 1977).
Рис. 1.
Район исследований. Кружки с цифрами – многолетние стационары: 1 – Собь, 2– Харп, 3 – Октябрьский, 4 – Хадыта, 5 – Ласточкин берег, 6 – Хановэй, 7 – Яйбари. Буквами обозначены сезонные стационары и пункты исследований коллег: Щ – Щучья, Ба – Байдарата, Ен – Ензоръяха, Ер – Еркутаяха, П – Порсыяха, Ю – Юрибей, Се – Сеяха, Я – Ясавэйяха, Бо – Бованенково, Х – Харасавэй, С – Сабетта, оБ – о-в Белый. Штриховыми линиями обозначены границы подзон.
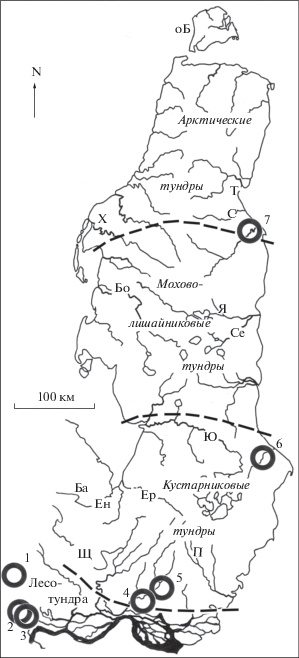
В среднем течении р. Хадытаяха в 1970–1973, 1978–1983 гг. функционировали стационары: фактория Хадыта (67°00′), Ласточкин берег (67°08′). Здесь учитывали пары на пробных площадках, ловили птиц паутинными сетями, отыскивали и прослеживали гнезда. В нижнем течении р. Нурмаяха на Среднем Ямале в 1974, 1975, 1982–1993 гг. на стационаре Хановей (68°40′ с.ш.) учитывали пары, отыскивали гнезда и регулярно их контролировали, ловили у гнезд и метили птиц цветными кольцами, подробно описывали признаки гнездящихся пар. На Северном Ямале в 1988–1995 гг. на стационаре Яйбари (71°04′ с.ш.) учитывали птиц на контрольных площадках, отыскивали гнезда.
Помимо этого, было организовано несколько сезонных стационаров, указанных на рис. 1: Порсыяха, Сеяха (Зеленая), Ясавэйяха. Указаны также пункты исследований коллег на Ямале: Щучья, Байдарата, Ензоръяха, Еркутаяха, Юрибей, Бованенково, Харасавэй. Не указаны на карто-схеме маршруты наших “ходовых” экспедиций разных лет, которые охватывали большую часть Ямала от его южной оконечности до окрестностей пос. Харасавэй и фактории Тамбей.
При обработке материалов применялись общепринятые методы статистики программы Statistika v. 6.0 (StatSoft Ink., 1984–2003).
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
В предгорьях Полярного Урала, в Приобской лесотундре и в тундрах Ямала чечетки гнездятся регулярно. Наиболее северная точка нахождения гнезд – стационар Яйбари. Несомненно, в качестве редкого вида чечетки гнездятся на Ямале и севернее, до предела кустарниковой растительности в подзоне арктических тундр. Но при маршрутном обследовании окрестностей Харасавэя, Сабетты и Тамбея мы чечеток не встречали, в публикациях по северному Ямалу и о-ву Белый чечетка не упоминается (Пасхальный, 1985; Морозов, Савинецкий, 1986; Сосин, Пасхальный, 1995; Дмитриев и др., 2006; 2015; Низовцев, 2017).
Биотопическое распределение. В северной тайге и лесотундре чечетки предпочитают гнездиться в пойменных смешанных лесах, в лиственничных редколесьях, зарослях ивняков по берегам озер и ручьев. В горах Полярного Урала беспокоящиеся птицы и гнезда не встречены выше 380 м над ур. м., кочующие попадались до 650 м над ур. м. (Головатин, Пасхальный, 2005). В Нижнем Приобье и на Ю. Ямале 71 из 85 найденных гнезд были в пойменном лесу, 10 – в лиственничном редколесье плакора, 4 – в ивняках и ерниках разной высоты по берегам водоемов. В кустарниковых и мохово-лишайниковых тундрах Среднего Ямала все гнезда (n = 523) были в зарослях ив (чаще – ивы сизой и ивы мохнатой) и карликовой березки на склонах балок разной экспозиции, в поймах, по берегам ручьев и озер. В лесотундре с началом послегнездовых кочевок птицы встречались повсеместно, от пойм рек до горных тундр Полярного Урала. Осенью стаи концентрируются в зарослях ольхи и в березняках, зимой небольшие стайки эпизодически встречаются в поймах.
Плотность гнездования. Чечетке свойственны значительные колебания численности гнездящихся птиц по годам, от полного отсутствия пар на контрольных участках до весьма высокой плотности. Причем в большей мере такие колебания и максимальная плотность характерны для лесотундры. На тундровом, с ивняками и отдельными лиственницами участке стационара Харп площадью 300 га (без озер), в 1982 и 2004 гг. гнездящихся пар не обнаружили, в 1970–1984, 2002, 2003 гг. было от 1 до 22 пар (0.3–7.4 пар/км2). В долине Оби на участке леса стационара Октябрьский площадью 22.4 га в 2004 г. чечеток не было, в 1978–1982, 2002–2003 гг. они гнездились с плотностью 2.1–187 пар/км2 (1–42 пары на участке). На участке пойменного леса стационара Хадыта площадью 14 га в 1978–1981 гг. гнездились 11–14 пар (78.6–107.1 пар/км2). На тундровом участке площадью 77 га гнездилось не более 5 пар (0–6.5 пар/км2). На лесном участке стационара Ласточкин берег площадью 13 га в 1971–1973 гг. гнездились 4–25 пар (30.8–192 пар/км2). Максимально высокая плотность гнездования чечеток в лесотундре приводится для долины р. Щучья: в лиственничниках с ольхой в 1973 г. учтено 270 пар/км2 (Кучерук и др., 1975).
Севернее границы лесной растительности, в подзоне кустарниковых тундр, плотность гнездования существенно ниже. В среднем течении р. Еркутаяха, впадающей в Байдарацкую губу, чечетка названа обычным гнездящимся видом (Штро и др., 2000), по данным Соколова (2006) в 2005 г. чечетки гнездились здесь с плотностью 6.3 пар/км2, в 2006 г. – 4.2 пар/км2. В верховьях р. Порсыяха в 1976 г. в пойменных ивняках на участке 41 га нами учтено 10 пар (22.4 пар/км2), на окружающем тундровом плакоре мы их не нашли. В нижнем течении р. Нурмаяха (стационар Хановей) на участке площадью 1.46 км2, включавшем пойму и плакор, в 1974, 1975, 1982–1993 гг. гнездились от 8 до 40 пар, т.е. 4.4–25.0 пар/км2. По данным Головатина и Пасхального (2008), в бассейне р. Юрибей в 2004 и 2005 гг. на всем протяжении реки на плакоре чечетки гнездились с плотностью 1.4–2.9 пар/км2, в пойме – 6.6–12.8 пар/км2. Севернее, на р. Ясавейяха в 1975 г. чечетки найдены нами только в пойме – 2 пары на площадке 50 га (4 пар/км2). Практически в том же районе, в низовьях р. Сеяха–Зеленая в 2006 г. чечетки были обычны, найдены гнезда в пойме и в овражной сети плакора (Рябицев, Примак, 2006). Несколько северо-западнее этого участка, на территории Бованенковского ГКМ чечетки в 1988–1990 гг. были обычны в пойменных ивняках (8.7–27.2 пар/км2), ивняках плакора (6.1–30.1 пар/км2) (Мониторинг …, 1997). В 2006 г. здесь же чечетка была обычным гнездящимся видом в ивняках (Слодкевич и др., 2007). На Северном Ямале, в районе стационара Яйбари чечетка зарегистрирована на гнездовании только в два года из 8 (1990 и 1993) как редкий вид, найдено по одному гнезду в пойме за пределами учетной площадки 3 км2. В другие годы на Яйбари эпизодически регистрировали одиночных птиц и пары.
Для чечеток характерно одиночное и групповое гнездование. Последнее влияет на результаты учетов, т.к. группа гнезд может быть за пределами небольшой учетной площадки или, напротив, может занимать всю площадку. Поэтому приходится говорить о локальной плотности гнездования (число пар на участке концентрации гнезд) и плотности для достаточно большой территории. В лесотундре в пойменных лесах чечетки тяготеют к колониям дроздов-рябинников (Turdus pilaris), но не всегда (Рыжановский, 1999).
В районе стационара Октябрьский в 1978–1990 гг. колонии рябинников были в течение 8 из 10 лет работы с дроздами. Поселения чечеток в колониях были в течение 4 разных лет, в остальные годы чечетки гнездились разреженно (рис. 2). Некоторые пары устраивали гнезда в нескольких метрах от гнезда дрозда и соседнего гнезда своего вида. В результате такой концентрации в 1978 г. 33 гнезда чечеток, из 42 контрольных, находились на площадке 4.5 га совместно с 7 гнездами рябинников. В 1988 г. на 26 гнезд рябинников в трех колониях было 21 гнездо чечеток по 3–7 гнезд в каждой колонии. В 1989 г – на 39 гнезд рябинников в двух больших колониях (по 10–12 гнезд) и четырех группах из 2–3-х гнезд пришлось 36 гнезд чечеток, расположенных преимущественно недалеко от гнезд дроздов (рис. 2).
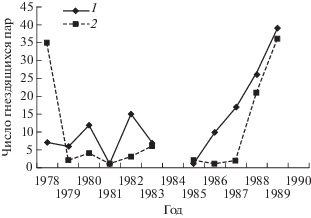
В тундрах Среднего Ямала колоний рябинников не было, но групповые поселения чечеток были обычны. Чаще они были приурочены к поросшим кустарниками склонам оврагов с удалением гнезд друг от друга на 10 и более м. На территории стационара Хановэй, на учетной площадке 1.46 км2 удалось выделить 12 участков, где в 1982–1993 гг. чечетки занимали гнездовые территории в большинстве лет наблюдений, а в трех точках чечетки гнездились ежегодно в количестве от 2 до 19 пар с локальной плотностью 3–10 пар/10 га. В подзоне мохово-лишайниковых тундр, в бассейне р. Мордыяха (район пос. Бованенково), было до 30 пар/км2 при гнездовании небольшими группами и одиночно. В окрестностях Яйбари регистрировали только одиночные гнезда.
Весенняя миграция. Обыкновенная чечетка имеет большой зимовочный ареал, выраженное стремление к стайности, благодаря чему особь может быть увлечена за сотни и тысячи километров от гнездового района. Но некоторое число чечеток, преимущественно светлой окраски, первую половину зимы проводит в лесотундре и с усилением морозов откочевывает в тайгу. В теплые зимы мы встречали чечеток в пойме Оби и долине р. Собь в январе–феврале.
Весной в Приобской лесотундре наблюдали два периода прилета чечеток: в конце марта – апреле и в конце мая – начале июня. Первый период был растянут на 2–4 недели, птицы появлялись при первом потеплении после весеннего равноденствия, иногда – раньше (14.04.1976, 28.04.1982, 7.04.1986, 18.03.1987, 2.04.1988, 5.04.2003, 28.04.2005, 26.04.2008, 14.04.2017), летели небольшими стайками и поодиночке. Появление птиц второго периода прилета наблюдали 27.05.1972, 22.15.1974, 30.05.1975, 15.05.1976, 20.05.1978, 22.05.1979, 20.05.1980, 23.05.1981, 16.05.1982, 24.05.1983, 30.05.1986, 15.05.1987, 18.05.1988, 26.05.1989, 21.05.2002, 17.05.2004. Начало второй волны обычно совпадало с ледоходом на Оби и снеготаянием; в отдельные годы это был массовый прилет, иногда его практически не наблюдали. Среднесуточная температура воздуха при начале прилета чечеток первой волны –17.8–3.2, в среднем –6.2°С (n = 8), при начале прилета второй волны –7.5–4.9, в среднем –0.7°С (n = 16). Продолжался прилет второй волны от 6 до 34 сут, в среднем 24.1 сут (n = 10).
Число пойманных мигрирующих чечеток в долине Нижней Оби из второй волны по годам различалось значительно. В 1978 г. линией из шести паутинных сетей в период с 26 мая по 10 июня поймано 1513 чечеток (2596 особей/100 м. сетей), в 1986 г. этой же линией и большой ловушкой за весь период прилета поймана 1 чечетка. Но чаще мы ловили от 10 до 100 птиц за весну. Самцы и самки прилетали одновременно, совпадали и периоды их массового отлова. В сетях и ловушке преобладали самцы, суммарно – 1180 самцов : 897 самок. Чем меньше было поймано птиц, тем ниже была доля самок: 7 : 0 (1977 г.), 14 : 3 (1987 г.), 20 : 7 (1982 г), 113 : 72 (1979 г.), 823 : 690 (1978 г.). Возможно, в годы с низкой численностью мигрантов мы ловили чечеток первой волны, где самцы при более раннем прилете активно разыскивали самок и чаще попадали в сети. В 1978 г., когда наблюдался интенсивный пролет, соотношение полов в стаях было близко к 1 : 1; в 1979 г. пролет был выражен в меньшей мере, но это также был пролет на север и соотношение полов было соответствующим.
Прилет чечеток на Средний Ямал (стационар Хановей) начинался в сроки, близкие к срокам, характерным для лесотундры, в конце мая. В начале июня чечетки были обычны все годы наблюдений (1982–1994 гг.), но прилет первых чечеток мы не регистрировали в связи с относительно поздним началом наблюдений. На Северном Ямале регистрировали начало прилета по голосам: 31.05.1989; 31.05.1990; 25.05.1991; 31.05.1993. При благоприятной погоде чечетки из лесотундры до арктических тундр долетали за 3–5 сут.
Предгнездовой период. Первое время после прилета чечетки держатся небольшими стайками, в которых происходит формирование пар, но некоторые птицы прилетали парами. Данных о гонадах птиц из первой волны нет, самцы второй волны прилетали с полностью сформированными гонадами, вес их колебался от 51 до 93 мг, в среднем 83.0 мг (n = 35). Яичники самок были слабо развиты, размеры наибольших фолликулов 0.4–1.0 мм, иногда 2.0 мм (Данилов и др., 1984).
Среди чечеток, пойманных в конце мая – начале июня, иногда встречались самки с наседными пятнами. Как правило, такие птицы попадали в сети и ловушку в составе стаи, где соотношение полов было близким к 1 : 1. Например, 1.06.1981 г. сетью поймана стайка чечеток из 8 самцов и 10 самок, где все самки имели наседные пятна стадии яйцекладки – начала насиживания; 30.05.1989 г., на третий день с начала прилета чечеток, в ловушку залетела стая из 11 самцов и 7 самок, причем 6 самок имели наседные пятна 4-й стадии (выкармливание птенцов), что позволяет предполагать утрату гнезд с птенцами где-то в тайге. Но в 1978 г. среди 760 самок особей с наседными пятнами не было.
Гнездостроение и яйцекладка. Расположение гнезд зависит от биотопа. В редколесьях плакора гнезда (n = 16) находили на ивах, лиственницах, березах, карликовой березке на высоте от 0.3 до 1.5 м. В пойменных лесах из 187 гнезд 120 были построены на елях, 20 – на березах, 19 – на ивах, 13 – на ольхе, 10 – на лиственницах, 3 – на жимолости, 2 – на пнях. Гнезда на деревьях располагались у ствола или на боковых ветвях на высоте от 0.3 до 14 м, в среднем 2.54 ± 0.37 м (n = 110). В кустарниковых тундрах единично находили гнезда на земле: в корнях ив и карликовой березки, в пучках сухой осоки, в старых наземных гнездах дроздов-белобровиков. Но подавляющая часть гнезд в тундрах была построена в развилках и переплетениях ветвей ив и карликовой березки ближе к вершине куста, на средней высоте 0.49 ± 0.02 м (n = 180). В низинах были обычны гнезда над водой. В тех оврагах стационара Хановей, где чечетки гнездились регулярно, некоторые гнезда сооружались над прошлогодними или брошенными не достроенными гнездами текущего года. На северном Ямале (Яйбари) оба найденных гнезда были устроены на земле в пойменных ивняках высотой всего около 20 см.
Строили гнезда только самки. Каркас образовывали тонкие веточки, стебли сухих трав, мох, пух ивы. Выстилка лотка была из перьев (чаще – из перьев белых куропаток) растительного пуха, шерсти, прошлогодней хвои лиственниц. Большинство гнезд являются плотной компактной постройкой со значительным количеством перьев и пуха, но встречались и рыхлые, неряшливые, просвечивающие, плохо укрепленные на ветвях. Строительство гнезда занимало от 2 до 7, обычно 3–4 сут, наиболее активно строили в первой половине дня. Защита территории у чечеток практически не выражена, что и позволяет им селиться очень плотными группировками, где гнезда располагаются всего в нескольких метрах одно от другого.
Формирование яиц в яйцеводе певчих птиц продолжается 4–5 сут (Зимин, 1988) и происходит одновременно со строительством гнезда. В лесотундре овогенез и гнездостроение начинается при среднесуточной температуре –1.0–8.1, в среднем 3.2°С (n = 15), яйцекладка в эти же годы начиналась при 1.6–13.3, в среднем 6.4°С (n = 15). Несмотря на зимовку при низких температурах и достаточно ранний прилет части птиц, чечетки имеют те же температурные пороги начала строительства гнезда и откладывания яиц, что отлетающие в низкие широты коньки, трясогузки, овсянки (Рыжановский, 2001).
В Нижнем Приобье в 1971–1989 гг. первые яйца в контрольных гнездах появлялись между 28.05 (1989) и 20.06 (1983), средняя за 16 лет наблюдений дата начала яйцекладки – 10.06. На Среднем Ямале в 1974–1994 гг. начало сезона откладывания яиц зарегистрировано между 1.06 (1991) и 23.06 (1975), средняя дата – 13.06 (n = 15). Таким образом, на пространстве от широты Полярного круга (66°30′ с.ш.) до широты стационара Хановей (68°40′ с.ш.) среднее запаздывание сроков начала откладывания яиц 1.5 сут на 1° широты, что меньше величины широтного градиента Гопкинса 4 сут (Hopkins, 1938 цит. по: Шульц, 1981). Но сопоставление дат начала яйцекладки в отдельные годы свидетельствует, что она может начаться как одновременно, так и с запаздыванием на севере до двух декад (табл. 1) в связи с поздним прилетом.
Таблица 1.
Сроки начала откладывания яиц на разных широтах в некоторые годы
| Год | Широта, град. | ||
|---|---|---|---|
| 66°40′ | 68°40′ | 70°10′ | |
| 1974 | 14.06 (5) | 18.06 (9) | – |
| 1975 | 14.06 (7) | 17.06 (5) | 20.06 (7) |
| 1983 | 20.06 (3) | 20.06 (25) | – |
| 1984 | 8.06 (13) | 16.06 (54) | – |
| 1985 | 10.06 (3) | 10.06. (49) | – |
| 1988 | 2.06 (9) | 8.06 (58) | 16.06 (5) |
| 1989 | 28.05 (27) | 18.06 (18) | 25.06 (2) |
В годы с достаточно большим числом контрольных гнезд (20 и более) период начала яйцекладки в лесотундре продолжался 10–37 сут, на Среднем Ямале – 18–31 сут. Главный период начала яйцекладки, во время которого в 80% контрольных гнезд появляется первое яйцо (Väisanen, 1977), продолжался 13–28 сут в разные годы при длительности главного периода у других северных видов 5–11 сут (Рыжановский, 2001). Значительная растянутость начала гнездового периода чечетки связана не только с повторным гнездованием после утраты гнезда, но и со вторым нормальным гнездованием небольшой части пар (Алексеева, 1986) или поздним прилетом птиц, вероятно, выкормивших птенцов первого выводка или утративших кладку в более южных районах (Hilden, 1969). В лесотундре не наблюдали прилет чечеток для второй кладки с севера, показанной для тайги окрестностей г. Архангельска (Андреев, 2016).
По объединенным данным в полной кладке чечетки было 4.79 ± 0.02 яйца (n = 757): 2 яйца было в 7 гнездах, 3 яйца – в 24 гнездах, 4 – в 146, 5 – в 522, 6 – в 58 гнездах. В лесотундре в полной кладке было 4.63 ± 0.04 (n = 228), в тундрах Среднего Ямала в кладке было 4.82 ± 0.03 (n = 525). Во втором районе было больше кладок с 5 и 6 яйцами, но увеличение величины кладки к северу не достоверно. На севере Скандинавии (Jarvinen, 1986) в гнездах было недостоверно больше яиц, чем на Ямале 4.98 ± 0.08 (n = 59), а в Северной Лапландии кладка была достоверно больше, чем в Южной Финляндии, что объясняют (Hilden, 1969) большей длиной светового дня.
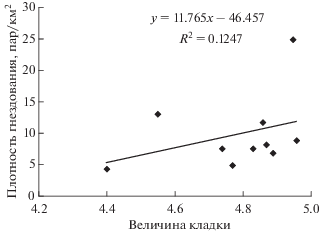
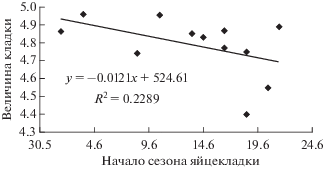
Для чечеток кустарниковых тундр Ямала отмечен положительный недостоверный тренд связи размера кладки с плотностью гнездования (рис. 3) и отрицательный недостоверный тренд связи размера кладки и сроков начала сезона гнездования (рис. 4). На участке Хановей в гнездах было больше яиц в годы с высокой плотностью пар, но чем позднее начинался сезон откладывания яиц, в основном в связи с поздним прилетом, тем меньше яиц было в гнездах.
Насиживание и выкармливание. Самки откладывают яйца в ночное время, сразу после окончания строительства, иногда в недостроенное, без пуховой выстилки, гнездо, иногда – на 2–4-е сутки после окончания строительства. В лесотундре чечетки чаще приступали к регулярному насиживанию в середине–конце периода яйцекладки, но в тундре Среднего Ямала в гнездах с 1 яйцом в дневное время самки отсутствовали в трети посещений, кладки с 2 и более яйцами насиживали обычно плотно.
Период от первого яйца до вылупления первого птенца длился 10–15, в среднем 12.9 ± 0.19 сут (n = 36). При соответствии порядка вылупления порядку яйцекладки это длительность инкубации первого яйца. Сопоставление длительности развития первого яйца чечетки с развитием яиц других северных воробьиных (Рыжановский, 2001) выявило достоверно более короткую длительность инкубации первого яйца чечетки по сравнению с длительностью инкубации первого яйца краснозобого конька (Anthus cervinus), варакушки (Luscinia svecica), овсянки-крошки (Ocyris pusillus), подорожника (Calcarius lapponicus), юрка (Fringilla montifringilla). Период от откладки первого яйца до вылупления последнего птенца (суммарный период насиживания) длился 12–18, в среднем 14.23 ± 0.18 сут (n = 36) и также был короче, чем у перечисленных выше видов. Мы объясняем такие темпы развития очень плотным насиживанием кладки. Вылупление в гнезде в связи с ранним началом насиживания обычно растягивается на 2–3 сут. Потревоженные наблюдателем птенцы сидят в гнезде не менее 8 сут, на 9–10-е сутки хорошо перепархивают, улетают из гнезда и вскоре возвращаются или держатся рядом с гнездом. Не потревоженные птенцы сидят в гнезде максимально 15 сут, средняя длительность гнездового выкармливания 12.3 ± 0.39 сут (n = 23).
Общая продолжительность периода от первого яйца до вылета последнего птенца из гнезда составляет 20–31 сут, в среднем 25.6 ± 0.47 сут (n = 31). Три пары выкормили по 2 выводка за 55–60 сут, одна пара в конце августа насиживала третью кладку из 3 яиц с расчетным вылетом птенцов в первых числах сентября, через 75–80 сут после начала размножения 20.06.1993. В лесотундре в разные годы сезон размножения длился 39–62, в среднем 47.1 ± 1.6 сут (n = 8), в субарктических тундрах – 35–75 сут, в среднем 51.1 ± 3.4 сут (n = 11). Растянутость сезона до 75 сут связана с нахождением третьей кладки в 1993 г. В отловах количество чечеток поздних сроков вылупления, из вторых и даже третьих выводков, не превышало нескольких процентов: 12 из 311 в 1978 г.; 3 из 260 в 1982, 0 из 242 в 1985 г.
Послегнездовые кочевки. Два окольцованных в соседних гнездах слетка пойманы на участке в возрасте 18 и 23 сут. Два других слетка, окольцованных в возрасте 17–20 сут, пойманы повторно через 35 и 56 сут. Судя по возрасту первого отлова, это были птицы местного происхождения, т.е. чечетки могут находиться в районе вылупления свыше двух месяцев. С учетом послегнездового стайного поведения, где группы постоянно перемещаются по пойменному лесу и редколесью плакора, распадаются и объединяются, не должно быть территориальной привязанности к небольшому району. Но она есть: из 1305 первогодков, окольцованных в 1977–1981 гг., повторно поймано 68 (5.2%). Большинство повторно пойманных находились на участке не более 10 сут, но 5 молодых особей были пойманы через 16–56 сут (Рыжановский, 1997). Анализ повторных отловов и состояния линьки чечеток из долины р. Собь показал, что в июле–августе 1977 г. в районе отлова были местные, начинающие линьку, птицы. В середине августа они перестали отлавливаться, но в большом числе появились чечетки на средних этапах линьки, предположительно, прилетевшие из тайги, так как в середине–конце августа в некоторые годы в лесотундре мы неоднократно наблюдали весьма активный пролет стай на север.
Взрослые птицы, окольцованные в гнездовой период, отлавливались на контрольном участке в лесотундре повторно до середины сентября, две особи пойманы через 69 и 75 сут после кольцевания на гнездах в июне, т.е. находились на участке все лето. Большинство взрослых чечеток, перелетев совместно с молодыми птицами из тайги в лесотундру, также ограничивают подвижность. Например, в 1977 г. долине р. Собь из 427 окольцованных птиц повторно пойманы 42 (9.84%) через 2–49 сут, до снятия сетей в середине сентября (Рыжановский, 1997). На маршрутах стаи чечеток встречались до октября–ноября с постепенно убывающей численностью. В Северной Англии во время линьки взрослые чечетки живут оседло (Ewans, 1969), в Приобской лесотундре период послегнездовой оседлости чечеток также долгий.
Постювенальная линька охватывает все контурное оперение головы и туловища, большую часть кроющих крыла (рис. 5) и пуховые перья на аптериях. Карпальное перо, большие верхние кроющие первостепенных маховых, маховые крыла и крылышка, рулевые остаются юношескими. Из больших верхних кроющих второстепенных маховых обычно заменяется меньше половины перьев, но иногда не линяют все. Часть птиц не сменяет средние верхние кроющие второстепенных маховых. Из нижних кроющих маховых чаще линяют кроющие второстепенных и третьестепенных маховых, но иногда – все (Рымкевич, Рыжановский, 1987). У 11 из 260 осмотренных в 1982 г. чечеток дополнительно линяли третьестепенные маховые, но в 1977 и 1978 гг. птиц с такой линькой не ловили. Не найдено отличий полноты линьки чечеток Нижнего Приобья от чечеток Приладожья (Носков, Смирнов, 1990) при большой индивидуальной изменчивости, вызванной фотопериодическими условиями.
Рис. 5.
Полнота постювенальной линьки кроющих крыла обыкновенной чечетки в Нижнем Приобье: 1 – линяет у всех птиц, 2 – линяет у части птиц, 3 – не линяет.
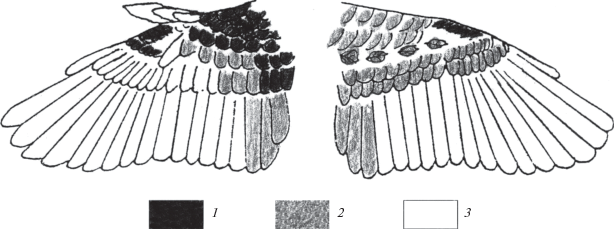
В Приладожье и Карелии, в зависимости от экспериментальных фотопериодических условий, линька начиналась в возрасте 28–70 сут (Носков, Смирнов, 1986). Чечетки, взятые из гнезд слетками в Нижнем Приобье, начинали линьку при короткодневном фотопериоде (16С : 8Т) в возрасте 25, 25, 26 сут (n = 3), при естественном фотопериоде широты Полярного круга – в возрасте 37, 44, 45 сут (n = 3), при длиннодневном фотопериоде (24С : 0Т) линька начиналась в 47–64, в среднем 54.6 ± 2.4 сут (n = 7). Таким образом, возраст начала линьки у чечеток лесотундры, как и у чечеток северной тайги Карелии, контролируется фотопериодом. Чечетки, взятые из гнезд на Среднем Ямале, выкормленные и передержанные при 24-часовом дне, начали линьку в возрасте 24, 32, 34 сут. Небольшая выборка не позволяет углубляться в последние данные, но полученные результаты позволяют сделать предположение об эндогенном контроле возраста начала линьки чечеток из субарктических тундр, как у ряда северных видов и популяций (Рыжановский, 2007).
В лесотундре чечетки в юношеском наряде отлавливались до начала сентября (последняя регистрация 8.09.1978 г.), значительно дольше других северных воробьиных. Связано это со вторым циклом гнездования и достаточно поздним в условиях фотопериода высоких широт возрастом начала линьки. Вступающих в линьку чечеток отлавливали с третьей декады июля (20.07.1982 г. первая регистрация) до начала сентября. Период вступления в линьку в разные годы был растянут на 25–35 сут. Закончивших постювенальную линьку чечеток отлавливали, начиная с третьей декады августа: 25.8.1977, 10.09.1978, 28.08.1979, 23.08.1982 гг. Индивидуальную продолжительность линьки в клетках установить сложно, т.к. птицы постепенно ощипывали друг друга. При коротком дне три особи, жившие одиночно, перелиняли за 45, 50, 50 сут. Примерная длительность при естественном фотопериоде лесотундры – 50–70 сут, как и в опытах, проведенных Носковым и Смирновым (1986). Сезон линьки продолжался до начала октября, в частности, 5.10.1982 пойманы 2 особи на завершающих этапах регенерации оперения.
Послебрачная линька полная, т.е. заменяется все оперение. В лесотундре начинающих линьку чечеток ловили со второй декады июля, не начавшие линьку птицы встречались до конца августа. Средние, по регрессии, даты начала линьки в течение ряда лет отличаются незначительно – 6.07.1977; 1.08.1978; 1.08.1979; 3.08.1980; 26.07.1982 гг., средние даты окончания – 15.09.1977; 25.09.1978; 13.09.1979; 22.09.1980; 4.09.1982 гг. (Рыжановский, 1997). Среди четырех взятых с птенцами чечеток одна самка, кормившая 10-дневных птенцов августовского (второго) выводка, находилась на 2-й стадии линьки, две самки начали линьку через 25–27 сут после вылупления птенцов первого выводка, самец – через 29 сут. В случае моноцикличного гнездового сезона линька не совмещается с выкармливанием птенцов, при полицикличном сезоне такое совмещение должно быть обычным. Связанных с полом различий в сроках линьки у стайных птиц не выявлено, среди первых в сезон линяющих чечеток ловили самцов и самок. Среднесезонная длительность линьки в течение 5 разных лет составляла 42–56, в среднем 46.6 ± 2.6 сут.
Осенняя миграция. Южное направление полета стай чечеток преобладает со второй половины сентября, но в отдельные годы стайки летели в окрестностях Октябрьского на юг уже в середине августа, возможно, в процессе кочевок. В середине октября чечеток серой окраски в лесотундре не наблюдали, стайки светлых (пепельных) встречали зимой. Миграционное ожирение у трех первогодков в клетках начиналось в возрасте 50, 55, 65 сут. Птицы с запасами жира “больше среднего” начинали регулярно отлавливаться с третьей декады августа, на последних этапах постювенальной линьки. В середине сентября в отловах доля жирных и заканчивающих линьку первогодков достигла 67%. Увеличение массы тела взрослых птиц началось в последней пятидневке августа, продолжилось в сентябре. К середине сентября доля среднежирных птиц достигала 40%.
В низовьях р. Ензоръяха (южный “угол” Байдарацкой губы) интенсивные перемещения чечеток в южном и юго-западном направлениях в 1992 г. отмечены 31.07, 2.08 и 22.08 (Черничко и др., 1997). В междуречье Байдараты и Ензоръяхи в 1980 г. размножение было явно неудачным и основная масса чечеток отлетела на юг еще в июле, последние пролетные стайки встречены 1.09. (Калякин, 1986). На р. Еркутаяха в 2001 и 2002 гг. Соколов (2003) отмечал чечеток как обычных птиц кустарниковых зарослей до конца сентября, когда он заканчивал полевые работы.
Зимние кочевки. На зиму серые чечетки откочевывают в таежные и смешанные леса, что подтверждают два зимних возврата от птиц, окольцованных осенью в Нижнем Приобье: Рыбинский р-н Ярославской обл. и г. Нижний Тагил Свердловской обл. Пепельные чечетки зимой встречаются в пойме Нижней Оби и значительно южнее, в частности в бассейне р. Малая Сосьва (Васин и др., 2015). Местные жители юго-западного Ямала отмечали стайки чечеток в январе–феврале на р. Еркатаяха (Соколов, 2003а). Пиминов (2005) неоднократно встречал чечеток в конце ноября и в марте в период с 1977 по 1989 г. в нижнем течении р. Щучьей и предполагает, что они держались там всю зиму.
Верность месту и филопатрия. Чечетка – типичный представитель группы лабильных видов, не имеющих территориального консерватизма или привязанности взрослых птиц месту гнездования (Рябицев, 1993). Из более чем 1000 взрослых особей, окольцованных нами в гнездовое время, в последующие годы не вернулась ни одна птица. По известной закономерности (Соколов, 1991), уровень филопатрии, т.е. верности месту рождения молодых птиц, многократно ниже верности взрослых птиц месту гнездования. От чечетки как ярко выраженного номадного вида не приходится ожидать возвратов к месту рождения молодых. Тем не менее, мы зарегистрировали возвращение одной молодой птицы на стационар Октябрьский весной на следующий год после ее рождения. Это была единственная особь из 3.5 тыс. окольцованных первогодков.
Питание. Нерегулярное гнездование и инвазии чечеток в Европе связывают с состоянием растительных кормов: семян ели, березы, ольхи, трав (Peiponen, 1967; Eriksson, 1970). В лесотундре Западной Сибири в желудках взрослых птиц помимо семян трав были мелкие беспозвоночные (цикады, червецы, комары, личинки жуков Curculionidae), а в пробах корма птенцов доминировали тли, медяницы, червецы, хирономиды и семена пушицы (Данилов и др., 1984). Корм такого состава в летний период есть всегда и повсеместно, влияние его на динамику обилия чечеток не должно быть значительным. Осенью в долине Оби чечетки концентрируются в зарослях ольхи, питаясь ее семенами; зимой кормятся по краям соров с зарослями арктофилы рыжеватой, вейника Лангсдорфа, вейника незамечаемого, осоки острой. В зимнем питании чечеток существенную роль играют мелкие семена трав. Пиминов (2005) главным зимним кормом называет семена ольхи.
ЗАКЛЮЧЕНИЕ
Данилов (1966) включил обыкновенную чечетку в группу птиц, связанную происхождением с Субарктикой. Прилет растянут на две волны – сначала прилетают зимующие в лесотундре птицы, затем кочующие по средним широтам. В некоторые годы в лесотундре весенней миграции и гнездования чечеток практически не было, интенсивная миграция в другие годы сопровождалась высокой плотностью гнездования. В кустарниковых тундрах миграция мало заметна, но на контрольной территории чечетки гнездились все годы наблюдений с плотностью на порядок ниже, чем в лесотундре.
Прилет в лесотундру чечеток с развитыми наседными пятнами свидетельствует о попытке гнездования в тайге. Успешно гнездившиеся в тайге птицы в лесотундру для второго цикла гнездования, вероятно, прилетали редко, поэтому не отлавливались. Возвращение для второго гнездования из тундр в лесотундру также не наблюдали, т.е. гнездовое население чечеток региона относительно стабильно. Групповое гнездование чечеток в лесотундре привязано к колониям рябинников, защищающих свою территорию от врановых, что повышает успех размножения. Но в тундрах, где рябинников нет, чечетки также тяготеют к формированию групп, проецируя стремление к стайности на гнездовой период.
На пространстве Нижнего Приобья и Ямала широтной изменчивости числа яиц в гнезде не выявлено, кладка была несколько больше при раннем ее начале и высокой плотности пар. Для чечеток Субарктики характерны высокие темпы роста–развития эмбрионов и птенцов, что позволяет выкормить два, иногда – три выводка. После распадения выводков взрослые птицы в связи с началом линьки частью остаются в гнездовом районе, частью перелетают в другой район, где останавливаются на период линьки. Молодые птицы в связи с линькой значительной полноты не должны совершать больших перелетов, но в некоторые годы в августе линяющие чечетки в массе появлялись в лесотундре, предположительно, из тайги.
Вид адаптирован к фотопериодическим условиям высоких широт. Выражается это в созревании гонад на подлете к широте Полярного круга, высоких темпах линьки при 24-часовом дне, в частичном совмещении послебрачной линьки с выкармливанием птенцов второго выводка, относительно раннем начале формирования миграционного состояния у первогодков. Фотопериодический контроль сроков начала постювенальной линьки чечеток, родившихся в лесотундре, и предполагаемый эндогенный контроль линьки чечеток, взятых из гнезд в субарктических тундрах, требует дополнительных исследований. Зимой на широте Полярного круга чечетки начинали кормовую активность при освещенности менее 1 люкс и минимальной длине кормового дня 5 час (Рыжановский, Пасхальный, 2008), что достаточно для обеспечения энергией на ночь, но, несомненно, ограничивает зимовку севернее, в зоне полярных ночей субарктических тундр Ямала.
Спектр питания и места сбора корма, вероятно, не ограничивают распространение вида. Птенцам требуется частично животная пища, взрослые питаются семенами, которых достаточно в тундрах и тем более в средней и южной тайге. Различия в величине клюва между морфами – чем светлее особь, тем короче клюв – вероятно, связаны с разной величиной объектов питания. Клюв пепельной чечетки слишком мал для обработки шишек ольхи в тайге и лесотундре, но удобен для шелушения зерновок и колосков тундровых трав. Помимо этого, маленький клюв снижает теплоотдачу в соответствии с правилом Дж. Аллена.
Список литературы
Алексеева Н.С., 1986. Вторая кладка у обыкновенной чечетки на Ямале // Орнитология. М.: Изд-во Московского ун-та. Вып. 21. С. 145.
Алексеева Н.С., 1988. Морфологическая изменчивость чечетки на Ямале // Изучение и охрана птиц в экосистемах Севера. Владивосток. С. 5–7.
Андреев В.А., 2016. Чечетка Acanthis flammea в Архангельске и его пригородной зоне // Русский орнитологический журнал. Т. 25. № 1274. С. 1359–1367.
Васин А.М., Лыхварь В.П., Буйдалина Ф.Р., Загузов А.В., Сыжко В.В., 2015. Позвоночные животные заповедника “Малая Сосьва” (Северное Зауралье). Аннотированный список и краткий очерк. Ижевск: Изд-во Пермяков. 136 с.
Головатин М.Г., Пасхальный С.П., 2005. Птицы Полярного Урала. Екатеринбург: Изд-во Уральского ун-та. 560 с.
Головатин М.Г., Пасхальный С.П., 2008. Современное состояние орнитофауны долины р. Юрибей (Южный Ямал) и перспективы создания в бассейне реки природного парка // Научный вестник ЯНАО. № 8. С. 81–102.
Данилов Н.Н., 1966. Пути приспособления наземных позвоночных животных к условиям существования в Субарктике. Т. 2. Птицы // Труды Ин-та биологии. Вып. 56. Свердловск: Изд-во Уральский рабочий. 140 с.
Данилов Н.Н., Рыжановский В.Н., Рябицев В.К., 1984. Птицы Ямала. М.: Наука. 334 с.
Дмитриев А.Е., Емельченко Н.Н., Слодкевич В.Я., 2006. Птицы острова Белого // Материалы к распространению птиц на Урале, в Приуралье и Западной Сибири. Вып. 11. С. 57–67.
Дмитриев А.Е., Низовцев Д.С., Харитонов С.П., 2015. Птицы острова Белый (Ямало-Ненецкий автономный округ) – результаты исследований 2014 года // Фауна Урала и Сибири. № 2. С. 61–71.
Зимин В.Б., 1988. Экология воробьиных птиц северо-запада СССР. Л.: Наука. 184 с.
Калякин В.Н., 1986. Птицы междуречья Байдараты и Ензор-яхи // Фауна позвоночных Урала и сопредельных территорий. Свердловск: Изд-во Уральского ун-та. С. 96–102.
Коблик Е.Ф., Редькин Я.А., Архипов В.Ю., 2006. Список птиц Российской Федерации М.: Товарищество научных изданий КМК. 281 с.
Кречмар А.В., Андреев А.В., Кондратьев А.Я., 1991. Птицы северных равнин. СПб.: Наука. 288 с.
Кучерук В.В., Ковалевский Ю.В., Сурбанос А.Г., 1975. Изменения населения фауны птиц Южного Ямала за последние 100 лет // Бюллетень Московского общества испытателей природы. Отд-ние биол. Т. 80. № 1. С. 141–148.
Минеев Ю.Н., Минеев О.Ю., 2012. Птицы Большеземельской тундры и Югорского полуострова. СПб.: Наука. 384 с.
Мониторинг биоты полуострова Ямал в связи с развитием объектов добычи и транспорта газа, 1997. Под ред. Добринского Л.Н. Екатеринбург: Изд-во Аэрокосмоэкология. 191 с.
Морозов В.В., Савинецкий А.Б.,1986. К авифауне С. Ямала // Орнитология. М.: Изд-во Московского ун-та. Вып. 21. С. 139–140.
Низовцев Д.С., 2017. Дополнения к орнитофауне острова Белый (Ямало-Ненецкий автономный округ) // Фауна Урала и Сибири. № 2. С. 171–172.
Носков Г.А., Рымкевич Т.А., 1977. Методика изучения внутривидовой изменчивости линьки у птиц // Методика исследования продуктивности и структуры видов в пределах их ареалов. Вильнюс: Изд-во Мокслас. Т. 1. С. 37–48.
Носков Г.А., Смирнов Е.Н., 1986. Некоторые особенности фотопериодического контроля сроков и полноты постювенальной линьки обыкновенной чечетки // Актуальные проблемы орнитологии. М.: Наука. С. 69–75.
Носков Г.А., Смирнов Е.Н., 1990. Чечетка – Acanthis flammea (L) // Линька воробьиных птиц Северо-Запада СССР. Л.: Изд-во Ленинградского ун-та. С. 131–135.
Пасхальный С.П., 1985. К фауне куликов и воробьиных арктической тундры Ямала // Распределение и численность наземных позвоночных полуострова Ямал. Свердловск: УНЦ АН СССР. С. 34–38.
Пиминов В.Н., 2005. К зимней орнитофауне Южного Ямала // Материалы к распространению птиц на Урале, в Приуралье и Западной Сибири. Екатеринбург: Изд-во Уральского ун-та. С. 225–228.
Портенко Л.А., 1939. Фауна Анадырского края. Птицы. Ч. 1. Л.: Изд-во Главсевморпути. 209 с.
Рогачева Э.В., Сыроечковский Е.Е., Черников О.А., 2008. Птицы Эвенкии и сопредельных территорий. М.: Товарищество научных изданий КМК. 754 с.
Рыжановский В.Н., 1997. Экология послегнездового периода жизни воробьиных птиц Субарктики. Екатеринбург: Изд-во Уральского ун-та. 288 с.
Рыжановский В.Н., 1999. Взаимоотношение чечеток Acanthis flammea и дроздов-рябинников Turdus pilaris в Нижнем Приобье // Русский орнитологический журнал. Т. 8. № 58. С. 9–14.
Рыжановский В.Н., 2001. Гнездовой сезон как часть годового цикла воробьиных Субарктики // Гнездовая жизнь птиц. Пермь: Пермский гос. пед. ин-т. С. 3–22.
Рыжановский В.Н., 2006. Доказательства существования и границы распространения на п-ве Ямал высокоширотной популяции белой трясогузки // Экология. № 2. С. 134–139.
Рыжановский В.Н., 2007. Роль фотопериодических условий высоких широт в дифференциации популяций воробьиных птиц // Русский орнитологический журнал. Т. 16. № 380. С. 307–314.
Рыжановский В.Н., Пасхальный С.П., 2008. Фотопериод в жизни воробьиных птиц Субарктики // Сибирский экологический журнал. № 1. С. 145–159.
Рымкевич Т.А., Рыжановский В.Н., 1987. Линька птиц на Полярном Урале // Орнитология. М.: Изд-во Московского ун-та. Вып. 22. С. 84–95.
Рябицев В.К., 1993. Территориальные отношения и динамика сообществ птиц в Субарктике. Екатеринбург: Наука. 297 с.
Рябицев В.К., 2008. Птицы Урала, Приуралья, и Западной Сибири: справочник – определитель. 3-е изд., испр. и доп. Екатеринбург: Изд-во Уральского ун-та. 634 с.
Рябицев В.К., 2014. Птицы Сибири: справочник-определитель в 2 т. М.– Екатеринбург: Кабинетный ученый. Т. 1. 438 с. Т. 2. 452 с.
Рябицев В.К., Примак И.В., 2006. К фауне птиц Среднего Ямала // Материалы к распространению птиц на Урале, в Приуралье и Западной Сибири. Екатеринбург: Изд-во Уральского ун-та. С. 184–191.
Слодкевич В.Я., Пилипенко Д.В., Яковлев А.А., 2007. Материалы по орнитофауне долины реки Мордыяха // Материалы к распространению птиц на Урале, в Приуралье и Западной Сибири. Екатеринбург: Изд-во Уральского ун-та. С. 221–234.
Соколов В.А., 2003. Осенний аспект населения птиц на юго-западном Ямале // Материалы к распространению птиц на Урале, в Приуралье и Западной Сибири. Екатеринбург: Изд-во Уральского ун-та. С. 170–175.
Соколов В.А., 2003а. К орнитофауне юго-западного Ямала // Материалы к распространению птиц на Урале, в Приуралье и Западной Сибири. Екатеринбург: Изд-во Уральского ун-та. С. 167–169.
Соколов В.А., 2006. Население птиц на юго-западном Ямале и его динамика. Дис. … канд. биол. наук. Екатеринбург. 16 с.
Соколов Л.В., 1991. Филопатрия и дисперсия птиц. Л.: Труды Зоологического ин-та. Т. 230. 232 с.
Сосин В.Ф., Пасхальный С.П., 1995. Материалы по фауне и экологии наземных позвоночных о. Белый // Современное состояние растительного и животного мира полуострова Ямал. Екатеринбург: Наука. С. 100–140.
Черничко И.И., Громадзкий М., Дядичева Е.А., Гринченко А.Б., 1997. Летне-осенний состав птиц восточного побережья Байдарацкой губы // Материалы к распространению птиц на Урале, в Приуралье и Западной Сибири. Екатеринбург: Изд-во Уральского ун-та. С. 149–155.
Штро В.Г., Соколов А.А., Соколов В.А., 2000. Орнитофауна реки Еркатаяха // Материалы к распространению птиц на Урале, в Приуралье и Западной Сибири. Екатеринбург: Изд-во Уральского ун-та С. 183–187.
Шульц Г.Э., 1981. Общая фенология. Л.: Наука. 188 с.
Cramp S., Perrins C.M. (Eds)., 1994. Handbook of the birds of Europe, the Middle East and North Africa. The birds of the Western Palearctic. V. 8. Crows to finches, Oxford: Oxford Univ. Press. 899 p.
Eriksson K., 1970. Ecology of the irruption and wintering of Fennoscandian redpolls (Carduelis flammea coll) // Annali Zoolgici Fennici. V. 7. № 3. P. 273–282.
Ewans P.R., 1969. Ecological aspects of migration and pre-migratory fat deposition in the Lesser Redpoll Carduelis flammea carabet // Condor. № 3. P. 316–330.
Hilden O., 1969. Ueber Vorkommen und Brutbiologie des Birkenzeisigs (Carduelis flammea) in Finisch-Lappland im Sommer 1968 // Ornis fennica. V. 46. № 3. P. 93–112.
Jarvinen A., 1986. Clutch size of passerines in harsh environment // Oikos. № 46. P. 365–371.
Peiponen V.A., 1967. Südliche Fortpflanzung und Zug von Carduelis flammea (L.) in Jahre 1965 // Annales Zoologici Fennici. V. 4. № 4. P. 547–559.
Väisanen R.A., 1977. Geographic variation in timing of breeding and egg sizes eight European specials of waders // Annales Zoologici Fennici. V. 14. P. 1–25.
Дополнительные материалы отсутствуют.
Инструменты
Зоологический журнал