

Зоологический журнал, 2021, T. 100, № 2, стр. 186-193
Новый род и новые виды семейства Kinnaridae (Cicadina) из Индии и Ирана
А. Ф. Емельянов *
Зоологический институт РАН
199034 Санкт-Петербург, Россия
* E-mail: Alexandr.Emeljanov@zin.ru
Поступила в редакцию 24.10.2019
После доработки 08.12.2019
Принята к публикации 15.12.2019
Аннотация
Описаны новый род и новый вид – Ptorthus kermanicus gen. et sp. n. из Ирана (провинция Фарс) и новый вид – Adolenda marusiki sp. n. из Индии (штат Уттар Прадеш, Гималаи).
Небольшое семейство Kinnaridae было установлено довольно поздно: его описал Муир только в 1925 г. (Muir, 1925). Вычленение этого таксона, как такового, и сразу в ранге семейства, произошло значительно позднее, чем описание, например, дочернего семейства Meenoplidae, распознанного Фибером еще в 1872 г. (Fieber, 1872). Но и в каталоге Меткафа (Metcalf, 1945) роды Southia Kirkaldy 1904, Micrixia Fowler 1904 и Adolenda Distant 1911 еще по-прежнему числились в семействе Cixiidae. Внешнее сходство с Cixiidae, в которое ранее помещали представителей Kinnaridae, подводило исследователей и уже после того, как были опубликованы сообщения и выводы Муира (Dlabola, 1957, 1981). Наиболее броские и веские отличия этого семейства от Cixiidae заключается в том, что у самок Kinnaridae редуцирован яйцеклад, хорошо развитый у Cixiidae и имеются поля восковых желез на VI–VIII тергитах брюшка, которых нет у Cixiidae.
Есть основания предполагать, что пределы распространения семейства еще не вполне обозначились. Об этом говорят сравнительно недавние, можно сказать неожиданные, географические находки в Афганистане (Dlabola, 1957), Иране (Dlabola, 1981; Емельянов, 1984), на Канарских островах (Remane, 1985), в горах Средней Азии (Емельянов, 1984, 1990), Южном Китае (Liang, 2001), в ОАЭ (Wilson, 2010,) в Сомали (Van Stalle, 1986), на о-вах Реюньон (Synave, 1958), а также в Чили (Emeljanov, 2016; Campodonico, Emeljanov, 2017).
Из Индии было описано два рода киннарид – Adolenda Distant 1911 и Paramicrixia Distant 1911 (Distant, 1911). Род Adolenda известен из Гималаев и c Тянь-Шаня; возможно, и род Paramicrixia гималайский, т.к. он указан из Бенгалии без точного местонахождения, территория же исторической Бенгалии на севере доходит до Гималаев. В Иране первые находки семейства сделаны Н.А. Зарудным еще в 1901 г. но описаны гораздо позже (Емельянов, 1984) уже после публикации Длаболы (Dlabola, 1981).
Типовые экземпляры вновь описываемых в данной статье видов хранятся в коллекции Зоологического института РАН в С.-Петербурге (ZIN) и в музее насекомых Х. Мирзаянца при иранском институте защиты растений в Тегеране (Hayk Mirzayans Insect Museum, Iranian Research Institute of Plant Protection, Tehran, Iran) (HMIM). Терминология дана в соответствии с принятой в разделе “Цикадовые” в Определителе Насекомых Дальнего Востока СССР (Ануфриев, Емельянов, 1988).
Ptorthus Emeljanov gen. n.
Типовой вид Ptorthus kermanicus sp. n.
Описание. Передние крылья сложены круто-крышевидно. Голова (рис. 1, 1–2) почти в 2 раза ýже переднеспинки. Корифа с резкими прямыми передним и боковыми килями, ее поверхность вогнутая, сзади к краю не приподнимается, оставаясь желобовидной, задний край глубоко прямоугольно выемчатый, с узко закругленной вершиной выемки, без ясного краевого киля. Передний край корифы сравнительно широкий, боковые края расходятся назад под острым углом, ширина корифы сзади примерно равна ее общей длине. Передний край корифы находится примерно на уровне передних краев глаз. Задняя выемка корифы занимает примерно половину ее общей длины. Средний киль не выражен. Лицо узкое, вытянутое. Метопа желобовидная без среднего киля, более чем в 3 раза длиннее своей наибольшей ширины, приходящейся на край клипеуса, нижний край метопы слабо вогнутый. Боковые кили метопы, если смотреть сбоку, почти без перелома продолжают линию края корифы, сверху изогнуты круто, ниже полого-дуговидно. Боковые кили метопы, если смотреть спереди, слабо выпуклые, от корифы к клипеусу они удаляются друг от друга и расстояние между ними в целом увеличивается более чем в 2 раза. Средний глазок крупный, лежит у самого клипеального края. Боковые кили постклипеуса высокие, составляют единое целое с боковыми килями метопы, на постклипеусе они сближаются друг с другом к границе с антеклипеусом и подходят к нему почти вплотную, средний киль резкий, но не приподнятый. Антеклипеус без боковых килей, в базальной половине (0.45–0.55) со средним килем, продолжающим киль постклипеуса, дистальнее без килей. Постклипеус примерно на треть короче метопы, антеклипеус немного короче постклипеуса. Хоботок заходит за вершины задних тазиков, он немного короче длины краниума, вершинный членик на треть короче предвершинного, вершина последнего находится на уровне вершин задних тазиков. Щеки ниже усиков с нерезким косо-поперечным килем (ребром полого-крышевидного поднятия), идущим от нижнего края усикового отверстия к нижней части боковых килей метопы, это, по-видимому, гомолог киля, свойственного многим Derbidae. Усиковое отверстие занимает примерно половину высоты щеки. Глаза сравнительно небольшие, почковидные, их нижний край полого вогнутый, здесь к нему примыкает усиковое отверстие. Усики простые, скап кольцевидный, педицелл яйцевидный, слабо вытянутый.
Рис. 1.
Ptorthus kermanicus Emeljanov gen. et sp. n.: 1 – передняя часть тела сверху, 2 – голова спереди-снизу (лицо), 3 – переднее крыло.
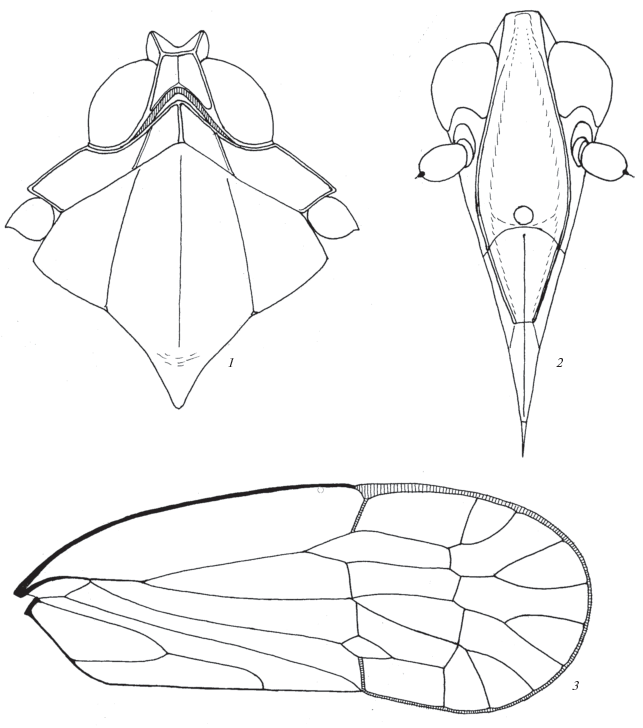
Переднеспинка сравнительно протяженная спереди назад, диск сравнительно длинный с хорошо развитыми боковыми килями. Передний край диска приблизительно прямым углом выдается вперед, на вершине закруглен, занимает заднюю выемку корифы, боковые кили расходятся назад, задний край тупоугольно вогнутый. Кили переднего края резкие, высокие, боковые кили и средний умеренно выступающие, более слабые, задний край без ясного киля. В связи с круто-крышевидным положением крыльев в покое, парадискальные поля довольно круто спускаютя вниз латерально, перелом идет по боковым килям диска. Задний край верха переднеспинки перед тегулами, плавно выпукло изгибаясь, отворачивается вперед-вбок, как обычно. Резкий киль переднего края диска, как единое целое, продолжается как передний (заглазничный) киль бочков верха переднеспинки (ее парадискальных полей) и под резким углом соединяется с таким же резким латеральным килем, коллатерального киля нет. Паранотальные лопасти, сверху ограниченные латеральным килем, ромбовидные. Тегулы без килей. Среднеспинка (щиток) обширная, ее передний край выступает тупоугольно, задний приблизительно прямоугольный, продольные кили щитка четкие, боковые слегка расходятся назад. Боковые доли щитка, так же как парадискальные доли переднеспинки, довольно круто наклонены латерально.
Передние крылья почти параллельнобокие, лишь слегка расширяются к перепоночке. Костальное поле умеренно широкое, почти параллельнобокое. Перепоночка занимает около 1/3 общей длины крыла, на вершине равномерно закругленная. Объединенное основание жилок (ScRM) в области базальной ячейки и до разделения ScR и M высоко-килевидное, особенно при базальной ячейке. Базальная жилка стигмальной ячейки (Sc) ослабленная, мембранозная, светлая (рис. 1, 3). Первые разветвления стволов ScR и M лежат немного базальнее нодальных поперечных жилок. Стигмальная ячейка с одной поперечной жилкой, почти не расширена в области постнодальной поперечной жилки (ra–rp), медиальное поле здесь не сужено. Островная ячейка (замкнутая дистально вилка CuA) маленькая. Вершинные ветвления жилок RP, MA и MP лежат лишь немного отступя дистально от постнодального ряда поперечных жилок. Объединенная клавальная жилка (Pcu + A1) перед вливанием в задний край крыла довольно круто дуговидно изогнута. Поперечная жилка, лежащая у вершины клавуса, также ослаблена как базальная жилка стигмы (рис. 1, 3). Ноги средних пропорций, задние голени без боковых зубцов, на вершине с венцом зубцов по формуле 3 + 5. На задних лапках на первом членике 7 зубцов без субапикальных щетинок, боковые зубцы слабо выделяются по величине, на втором членике 6 зубцов, боковые зубцы крупные без субапикальных щетинок, 4 внутренних зубца с субапикальными щетинками.
Брюшко самки с восковыми полями на VI–VIII тергитах. Гениталии самки (не препарировались) в целом типичные для семейства. Придатки VIII сегмента (предположительно, слитые рудименты базикокситов и вальв) выглядят как цельное образование, напоминающее вершинную половину ложки. Можно различить боковые более пигментированные части (предположительно вальвиферы) и менее пигментированные и менее склеротизованные средние (пердположительно вальвы). По средней линии створки плотно сомкнуты. Впадина над первыми створками с боков ограничена краями тергита, которые сближаются дорсально к основанию анальной трубки, анальная трубка сравнительно узкая, прикрывает бóльшую часть впадины, оставляя неприкрытой щель между нею и первыми створками.
Этимология. От др. греческого πτορθος – отпрыск, побег.
Обсуждение и диагноз. Новый род может быть сближен с родами Perloma Emeljanov 1984 и Adolenda Distant 1911. Он отличается от них круто-крышевидно сложенными крыльями и соответственно более круто наклоненными латерально боковыми частями переднеспинки и щитка среднеспинки. Тупоугольный перелом между плоскостью диска переднеспинки и плоскостью ее парадискальных долей обозначен боковыми килями диска, которых нет у других представителей трибы Adolendini, у этих последних имеется единое дискально-парадискальное поле, либо плоское, либо полого поперечно выпуклое, как, например, у Adolenda marusiki sp. n. (рис. 1, 4). Кроме того, переднеспинка у нового рода более длинная, чем она напоминает переднеспинку в трибе Prosotropini, однако по степени развития и конфигурации килей сходства нет. Новый род, кроме того, несет слабо выраженный субантеннальный генальный киль, о котором уже было сказано в описании, подобный киль также имеется у представителей рода Perloma.
Ptorthus kermanicus Emeljanov sp. n. (рис. 1)
Материал. Голотип самка: Iran, Fars Province, nr. Dashte Arzhan, 2218 m, 29°33.139′ N 51°57.058′ E, 14.VI.2019, V.M. Gnezdilov leg. (ZIN). Паратип, 1 самка: Iran, Fars Province, nr. Dashte Arzhan, 2218 m, 29°33.139′ N, 51°57.058′ E, 14 VI 2019, V.M. Gnezdilov leg. (HMIM).
Описание. Окраска. В целом, в окраске преобладают серовато-бурые тона. Ноги, а также средняя полоса вдоль тела сверху от корифы до вершины клавуса при сложенных крыльях, светлые. Корифа и верхняя часть метопы между глазами светлая, желтоватая с легким оранжево-бурым оттенком, ниже голова сероватая, переход к затемнению размытый. Глазки светлые. Над щечным килем размытое светлое пятно. Верхний край уздечек со светлым, почти белым, поперечным пятном. Диск переднеспинки белый, остальные части буровато-серые. Диск щитка среднегруди почти белый с легким буроватым оттенком, парадискальные поля буровато-серые, по бокам от светлых боковых килей окраска парадисков сгущается до черного в виде узкой полосы с размытым внешним краем. Тегулы светлые. Передние крылья в основном серовато-бурые. Кориум и перепоночка буровато-серые или серовато-бурые с неяркими мелкими светлыми пятнышками, на клавусе и в базальной половине (0.45–0.55) кубитального поля кориума крапинок нет. Жилки темные. Основание переднего крыла на уровне базальной ячейки осветлено. Впереди жилки ScR идет цепочка вытянутых светлых пятнышек, узкие темные границы между ними косые, выглядят как намечающиеся (длинные) косые линии, пересекающие все костальное поле, как у некоторых киннарид, например у Adolenda fuscofasciata Liang 2001 и других Fulgoroidea. На перепоночке между конечными жилками от края крыла идут клиновидные нечетко ограниченные светлые пятна, а на вершинах жилок затемнение усиливается. На клавусе краевое поле целиком светлое до белого, жилка A1 и продолжающая ее Pcu + A1, светлая, только на A1, немного базальнее вилки, жилка A1 на небольшом отрезке затемнена, также затемнена вершина Pcu + A1, далее до вершины клавуса располагается светлое пятно, завершающее светлую спинную полосу, описанные темные выступы на жилке A1 и вершине Pcu + A1 образуют перетяжки на этой полосе. Брюшко темно-бурое.
Длина самки 5.6–5.8 мм.
Замечание. По свидетельству В.М. Гнездилова, вид собран кошением по деревьям в разреженном лесу с дубом, боярышником, кленом и фисташкой.
Этимология. От названия иранской провинции Керман (Kerman), по местонахождению описываемого вида.
Adolenda marusiki Emeljanov sp. n. (рис. 2–3)
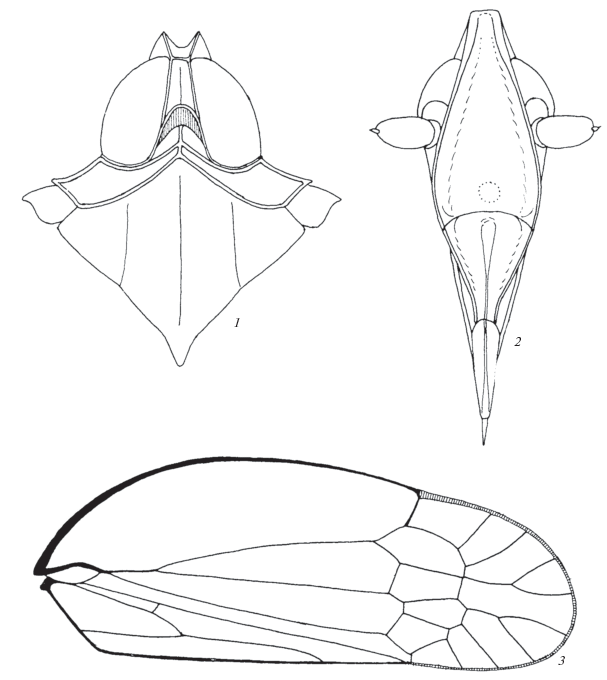
Рис. 2.
Adolenda marusiki Emeljanov sp. n.: 1 – передняя часть тела сверху, 2 – голова спереди-снизу (лицо), 3 – переднее крыло.
Материал. Голотип самец: India, Uttar Pradesh, Babind Ghat village, 30°37.5′ N, 79°33.5′ E, 1750–1900 m, 17–23. V 1999, Yu.M. Marusik leg. (ZIN). Паратипы: 1самец, 3 самки, India: Uttar Pradesh, Babind Ghat village, 30°37.5′ N, 79°33.5′ E, 1750–1900 m, 17–23. V 1999, Yu. M. Marusik leg. (ZIN).
Описание. Сходен с типовым видом рода – Adolenda typica Distant 1911, к которому, по-видимому, близок (гениталии A. typica не изучены). Передний край корифы узкий, корифа желобовидная, спереди отгорожена ее передним килем, сзади не замкнутая, желоб не ограничен перегородкой. Задний край глубоко остроугольно выемчатый, вершина выемки узко закруглена, выемка доходит примерно до уровня середин глаз (если смотреть сверху). Корифа и метопа в профиль соединяются под тупым углом, близким к прямому. Метопа без среднего киля, также желобовидная, ее боковые кили листовидные, в поперечном сечении она снизу трапециевидная, сверху почти клиновидная. Боковые кили в верхней половине (0.45–0.55) слегка вогнутые, в нижней – более отчетливо выпуклые, плавно, без перерыва переходят в боковые кили постклипеуса, которые сближаясь и понижаясь, подходят к антеклипеусу; перед самым антеклипеусом кили слегка отгибаются наружу и становятся параллельными. Граница метопы и клипеуса – прямая желобовидно вдавленная. Средний глазок крупный, примерно на треть своего диаметра отодвинут от шва. Хоботок умеренно длинный, у самцов почти доходит до переднего края пигофора, его основание приходится на вершины задних тазиков. Переднеспинка короткая, особенно в средней части, где передний и задний кили сближены почти на собственную толщину или на 1.5 ее. Задние и передние кили резкие, соединены по бокам боковыми килями верха переднеспинки, столь же резкими. Средний киль диска переднеспинки выходит вперед от места соединения половинок переднего киля. Щиток крупный, его видимый передний край, обозначенный задним краем переднеспинки, слабо тупоугольный, задний примерно прямоугольный, боковые кили щитка расходятся назад слабее, чем у A. typica на рисунке у Дистанта (Distant, 1911, 1916). Передние крылья A. marusiki отличаются от таковых у A. typica более широким костальным полем и более круто изогнутой костальной жилкой. Первая постстигмальная жилка RP1 отходит как вторая стигмальная жилка. Интеррадиальная жилка присоединяется сзади не к общему стволу RP, а к его перовой ветви RP1 вблизи основания ствола, апикальная интеррадиальная ячейка при этом выглядит, как часть стигмы, однако, в отличии от собственно стигмы, она не уплотнена. У A. typica, судя по рисунку Дистанта (l. c.), стигма также одноклеточная и уплотненная, но первая постстигмальная жилка отходит обособленно от ствола от ствола RP или в точке соединения жилки ir с RP. На вершине задней голени 7 зубцов (3 + 4). На первом членике задних лапок 7 зубцов без субапикальных щетинок, на втором членике – 6 зубцов, из них 4 внутренних с субапикальными щетинками.
Основной тон окраски светло-бурый, кили светлые, почти белые. Боковые листовидные кили головы буроватые со светлыми гребнями. Глазки светлые. Хоботок темно-бурый. Тегулы буроватые с широко осветленными краями. Передние крылья прозрачные с буроватыми жилками. На перепоночке буровато затемнены уплотненная стигма и постклавальная краевая ячейка. Более интенсивно затемнены поперечные жилки нодального ряда и соединяющие их участки продольных жилок, в постнодальном ряду затемнены жилки rm и im. На перепоночке окрашены белым жилки MA1 и MA2, окаймляющие ячейку между ними в антеапикальном ряду и жилка, отходящая от вершины этой ячейки. Все апикальные жилки перепоночки затемнены, затемнение усиливается к краю крыла. Жилка MP между постнодальным рядом и основанием апикальной вилки также светлая. На клавусе два темных пятна, одно на его вершине и второе в вилке между жилкой Pcu + A1 и краем крыла. Бедра передних и средних ног бурые со светлыми килями, голени более равномерно бурые, лапки немного темнее голеней, иногда бедра и голени почти целиком светлые. Задние бедра буроватые со светлыми килями, задние голени светло-бурые с черными зубцами на вершине, лапки также светло-бурые с черными зубцами. Брюшко темно-бурое, задние края сегментов осветлены. Гениталии у обоих полов более светлые с размытыми более темными пятнами, как бы линялые.
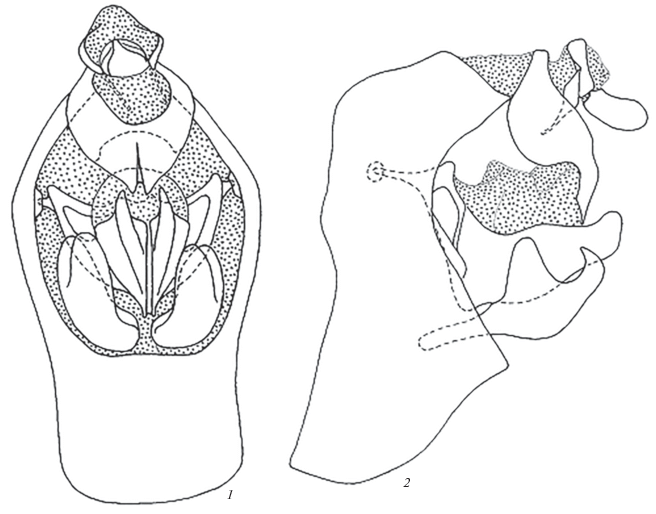
Гениталии самца. Пигофор короткий и высокий, его высота более чем в 2 раза превосходит наибольшую ширину. Боковая часть заднего края полого дуговидно вогнутая, верхняя часть над сочленением с анальной трубкой также полого дуговидно вогнутая, выступ для сочленения с анальной трубкой прямоугольный с резко выраженной вершиной. Нижняя стенка пигофора продольно и поперечно выпуклая, обращенная в целом дорсо-каудально, соответственно, заднее отверстие в склеротизации пигофора меньше по высоте, чем переднее. Анальная трубка с короткой дорсальной стенкой, боковые стенки широкие с дуговидно выступающим вентрально нижним краем, перед вершинными отростками нижний край круто сужается, так что отростки служат продолжением дорсальной поверхности трубки. Отростки широкие дорсо-вентрально уплощенные, их медиальные края тесно сближенные, сросшиеся почти до самых вершин, разделены дорсальным желобком, свободные вершины заостренные. Заднее отверстие в склеротизации пигофора закругленно-пятиугольное, примерно в полтора раза выше своей ширины. На внутренней поверхности боковых стенок пигофора в его верхней трети располагается пара чашечек сочленения с фаллобазой. Фаллобаза с треугольным в очертаниях основанием, вершиной обращена вниз, вблизи ее боковых углов расположены мыщелки, входящие в вышеописанные чашечки на стенках пигофора. В верхней части бока фаллобазы имеют вид рамки благодаря утолщенным краям. Ближе к средней линии фаллобазы от нее отходит назад пара вытянутых латерально сдавленных лопастей с закругленными вершинами. Между лопастями и над ними располагается аморфная мембранозная часть пениса, имеющая вид комка. Под фаллобазой, занимая все пространство до нижнего края заднего отверстия пигофора, помещаются стилусы, они, как обычно в семействе, в целом дуговидные, толстые, отогнутые вершинами дорсально и несущие также толстые суббазальные выступы на своей дорсальной стороне.
Длина самца 5.2, самки – 5.4–5.6 мм.
Сравнительные заметки. От типового вида, A. typica, новый вид отличается (1) более узкой слабо расширяющейся назад корифой, (2) более широкой метопой с выпуклыми, а не прямыми боковыми килями, почти параллельными, а не в значительной степени расходящимися назад боковыми килями щитка, (3) более широким костальным полем с более круто изогнутой костальной жилкой и (4) тем, что первая постстигмальная жилка RP1 отходит, как вторая стигмальная, т.к. поперечная жилка ir присоединена к RP1, а не к общему стволу RP перед его первым разветвлением, как это имеет место у A. typica.
Этимология. Вид назван в честь сборщика, известного арахнолога Ю.М. Марусика.
Список литературы
Ануфриев Г.А., Емельянов А.Ф., 1988. Подотряд Cicadinea (Auchenorrhyncha) – Цикадовые // Определитель насекомых Дальнего Востока СССР. Т. II. Равнокрылые и Полужесткокрылые. Л.: Наука. С. 12–485.
Емельянов А.Ф., 1984. К познанию семейств Kinnaridae и Meenoplidae (Homoptera, Fulgoroidea) // Энтомологическое обозрение. Т. 63. Вып. 3. С. 468–483.
Емельянов А.Ф., 1990. Новый вид рода Bashgultala (Homoptera, Kinnaridae) из Таджикистана // Зоологический журнал. Т. 69. Вып. 4. С. 137–139.
Campodonico J.F.,Emeljanov A.F., 2017. A new species of Apocathema Emeljanov, 2016 (Hemiptera, Fulgoroidea: Kinnaridae) from Coastal Central Chile // Entomological Review. V. 97. № 8. C. 1080–1084.
Distant W.L., 1911. Description of new genera and species of Oriental Homoptera // Annals and Magazine of Natural History. Ser. 8. V. 8. P. 735–747.
Distant W.L., 1916. Rynchota. The Fauna of British India, including Ceylon and Burma. London. V. VI. Homoptera: Appendix. Vii + 248 p. 177 figs.
Dlabola J., 1957. Die Zikaden Afghanistans (Homopt. – Auchenorrhyncha) // Mitteilungen der Münchner Entomologischen Gesellschaft. Bd. 47. S. 265–303.
Dlabola J., 1981. Ergebnisse der Tschechoslovakisch-Iranischen Entomologischen Expeditionen nach dem Iran (1970 und 1973). (Mit Angaben über einigen Sammelresultate in Anatolien). Homoptera: Auchenorrhyncha (II. Teil) // Acta Entomologica Musei Nationalis Pragae. V. 40. P. 127–311.
Emeljanov A.F., 2016. First record of the planthopper family Kinnaridae (Homoptera, Fulgoroidea) in Chile // Entomological Review. V. 96. № 9. P. 1203–1208.
Fieber F.X., 1872. Katalog der europäischen Cicadinen, nach Originalien mit Benützung der neuesten Literatur. i–iv, 1–19.
Fowler W.W., 1904. Order Rhynchona. Sudorder Hemiptera-Homoptera // Biologia Centrali Americana. V. 1. P. 21–139. Tabl. 4–13.
Kirkaldy G.W., 1904. Bibliographical and nomenclatural notes on the Hemiptera. № 3 // Entomologist. V. 37. P. 279–283.
Liang Ai-Ping, 2001. First record of the genus Adolenda Distant (Hemiptera: Fulgoroidea: Kinnaridae) from China, with description of one new species // Zoological Studies. V. 40. № 4. P. 365–370.
Metcalf Z.P., 1945. Kinnaridae. General catalogue of the Homoptera. Publ. by Smith College, Northampton, Fasc. 4. Pt. 7. P. 219–249.
Muir F., 1925. On the genera of Cixiidae, Meenoplidae and Kinnaridae // Pan-Pacific Entomologist. V. 1. № 3. P. 97–110.
Remane R., 1985. Kinnaridae in der S-W Paläarktis: zwei neue Taxa von den Kanaren (Homoptera Fulgoromorpha) // Marburger Entomologische Publikationen. Bd. 10. Ht. 1. S. 241–264.
Synave H., 1958. Une famille nouvelle pour la faune des îles de la Réunion et Maurice: les Kinnaridae (Homoptera-Fulgoroidea) // Bulletin et annales de la Société royale belge d’entomologie. V. 94. № 3–4. P. 118–121.
Van Stalle J., 1986. Propleroma atrifasciata sp. n., first record of a Kinnarid on the African continent (Homoptera, Kinnaridae) // Annali del Museo Civico di Storia Naturale “G. Doria” Genova. Doriana. Suppl. V. 6. № 258. P. 1–4.
Wilson M.R., 2010. Order Hemiptera, families Meenoplidae and Kinnaridae // Arthropod Fauna of the UAE. V. 3. P. 126–131.
Дополнительные материалы отсутствуют.
Инструменты
Зоологический журнал