

Зоологический журнал, 2021, T. 100, № 3, стр. 307-316
Максимальная температура тела как параметр терморегуляции рептилий: опыт статистической оценки на примере обыкновенной гадюки (Vipera berus)
А. В. Коросов a, *, Н. Д. Ганюшина a, **
a Петрозаводский государственный университет
185000 Петрозаводск, Россия
* E-mail: korosov@mail.ru
** E-mail: ekoni@mail.ru
Поступила в редакцию 22.09.2019
После доработки 18.12.2019
Принята к публикации 04.01.2020
Аннотация
Максимальная температура тела характеризует верхнюю границу диапазона толерантности к температурному фактору. Проведенный нами статистический анализ позволил выявить пять показателей максимальной температуры тела – критическую, высшую, типичную, среднюю, добровольную. Выполнен анализ полевых замеров температуры тела ртутным термометром и данных длительного постоянного наблюдения за температурой тела с помощью логгеров. Одновременно вели видеосъемку для фиксации моментов осуществления терморегуляторного поведения. Оказалось, что выборочные “средние максимальные значения” широко варьируют в зависимости от условий наблюдений и не пригодны для сравнения внутривидовых групп. Показатели “максимальная типичная температура” (37.9 ± 0.2°С) и “максимальная добровольная температура” (33.9 ± 1.8°С) статистически более устойчивы и различаются примерно на 4°С. Для изучения терморегуляторного поведения рекомендуется применять логгеры, а в качестве устойчивого параметра терморегуляции – брать показатель “типичная максимальная температура”.
В термобиологии рептилий используются около 20 показателей (Черлин, 1991), которые можно условно разбить по происхождению на три группы: экстремальные, добровольные и статистические. Экстремальные значения температурной устойчивости (критические пороги выживания особей) достаточно легко могут быть определены в эксперименте, эти значения хорошо отражают специфические возможности вида. “Добровольные” температурные показатели оцениваются в момент свободного выбора животным той или иной температуры тела или среды и в большей мере характеризуют текущее состояние особи. Статистические температурные показатели (средние, диапазоны) призваны описать некие типичные температурные характеристики особей, обнаруженных в определенной природной обстановке. При большом разнообразии показателей сложно выявить закономерности изменения температуры тела теплолюбивых пойкилотермных животных в условиях разнонаправленных флуктуаций внешнего потока тепла. Для системного решения этой проблемы необходимо последовательно изучать смысл и работоспособность различных оценок, чтобы выбрать простые и теоретически обоснованные показатели. Одна из важных характеристик – максимальная температура; рассмотрим ее смысл относительно одного вида рептилий – обыкновенной гадюки (Vipera berus). Чаще всего в литературе приводится три варианта максимальной температуры: максимальная температура выживания, максимальная температура, замеренная в поле, и максимальная добровольная температура.
Довольно давно было определено значение верхнего предела выживания гадюки – 37.25°С (Чан–Кьен, 1967), которое кочует из публикации в публикацию (Банников и др., 1977). Тепловая смерть в природе возможна (Пестов, Маннапова, 1999), но конкретные значения критической температуры требуют уточнения; одна из последних оценок – 38.1°С (Горбунова, 2016)11.
Максимальная температура тела обыкновенной гадюки измерялась в природе многими исследователями. Ее оценки обычно лежат в диапазоне от 34 до 38.2°С (см. ниже). Понятно, что в определении этого показателя велика роль случайности, значит, необходимо выяснить, каков источник отмеченного различия в 4°С (34–38°С) и как (статистически корректно) можно точнее определить его величину.
Максимальные добровольные температуры указывают на тот уровень терпимости организма, после достижения которого включаются различные терморегуляторные механизмы, снижающие или стабилизирующие температуру тела особи. При достижении такого порога обыкновенная гадюка покидает слишком горячее место баскинга, перемещаясь в более прохладное. В литературе отражены результаты определения этого показателя, которые колеблются в диапазоне пяти градусов – 30–35°С (Черлин, 2013; Коросов, 2010).
Сравнение этих трех типов оценок у разных видов рептилий или у особей одного вида из разных точек ареала теоретически могло бы выявить региональную или межвидовую специфику физиологических особенностей групп. Однако такие различия можно обнаружить только между методически точно определенными значениями, имеющими невысокие статистические ошибки. К сожалению, в литературе значения варьируют в широких пределах. Выход из ситуации мы видим в том, чтобы, во-первых, применить технологию массового сбора данных (с помощью имплантированных логгеров) и, во-вторых, разработать методы оценки устойчивых параметров.
В качестве объекта взят единственный массовый вид рептилий в Карелии, с которым авторы давно работают, – обыкновенная гадюка. Авторы рассматривают этот вид как модельный с перспективой приложения разработанных критериев к другим видам и другим экологическим ситуациям.
Цель настоящего сообщения состоит в том, чтобы проанализировать группу показателей “максимальная температура” в отношении обыкновенной гадюки и предложить технологию определения лучших характеристик.
МАТЕРИАЛЫ И МЕТОДЫ
Работы по исследованию термобиологии обыкновенной гадюки в полевых условиях проводились в 1978–1981 и в 1989–2002, 2006–2007 гг. на о-ве Кижи (62°07′ N, 35°22′ E). Измеряли температуру у всех змей, отловленных на маршруте в дневное время. Всего было выполнено 990 замеров температуры тела при разных погодных условиях. После отлова животных в течение 1–5 мин определяли следующие характеристики (Черлин, 1991): 1) температуру спинной поверхности тела измеряли с помощью дистанционного пирометра (ДТ) “Termopoint” (точность 1°С, оптическое разрешение 50 : 1), 2) внутреннюю температуру тела (ректальную или оральную) измеряли с помощью психрометрического ртутного контактного термометра (точность 0.2°С, рекомендуемое время измерения 3 мин), 3) температуру поверхности субстрата вблизи от места отлова измеряли обоими инструментами. Погода оценивалась как “ясная” при наличии отдельных облаков на небе, как “пасмурная” при сплошной облачности, “переменная” – в остальных случаях.
Исследование температуры тела в эксперименте выполнялась в мае–июне 2017–2018 гг. на биологическом стационаре Гомсельга (Республика Карелия, 60 км от г. Петрозаводск, 62°07′ N, 33°96′ E). Змеи жили в вольере (площадка 8 на 8 м, огороженная полиэтиленовым забором высотой 1 м) со скошенной травой, ночевали в типичном для Карелии убежище – каменной гряде (размером 1.5 × 1.5 × 1 м), сложенной из валунов размером 10–20 см в диаметре. Наблюдали за температурой тела и поведением 16 взрослых особей гадюки обыкновенной: 11 самок (длина тела от 52 до 60 см, 4 яловых и 7 беременных) и 5 самцов (длина тела от 51 до 56 см). Для распознавания разных змей их метили разноцветным лаком для ногтей. Помимо змей в вольеру были выпущены 10 особей травяной лягушки, часть которых съели 4 змеи, т.е. наличие логгера не препятствовало нормальной жизнедеятельности особей. Температуру регистрировали с помощью 11 логгеров ДТНЗ–28 (масса 1 г, объем 0.8 мл, точность измерения ±0.1°С, рабочий диапазон от –15 до +60°С, производство ООО “ЭМБИ”) и 5 логгеров ds1921, вживленных под кожу спины примерно посередине туловища. Логгеры фиксировали температуру тела через каждые две минуты. За 42 дня наблюдений для каждой змеи получен ряд замеров по 14 тысяч записей, общий объем базы данных для 16 змей составил около 230 000 чисел.
Поведение фиксировалось на видео web-камерой и action-камерами “Soccoo S20WS” (1080 P full HD) и “YILite”. Расшифровка видеозаписей состояла в распознавании и присвоении номера той позе, которую принимает змея (1 – расправлена в ленту, 2 – лежит свободно, тело округлое, 3 – лежит в рыхлом кольце, 4 – формирует плотный клубок) и номера типа поведения (0 – неподвижна, 2 – ползет). Номер позы и формы поведения каждой змеи вносили в базу. Заполненная база данных для каждого двухминутного периода наблюдений содержала значение температуры тела данной особи, номер ее позы и номер формы поведения. Для точности характеристик важно выполнять замеры при одной форме поведения (Черлин, 2012), мы регистрировали температуру тела гадюк только во время баскинга.
База данных организована в среде MS Excell; количественная обработка выполнялась в среде R (Коросов, Горбач, 2017).
Смысл, техника определения и расчета показателей
Анализ большого объема уникальных данных по непрерывному ряду значений температуры тела обыкновенной гадюки привел нас к необходимости ввести дополнительные показатели из серии “максимальные температуры” и переопределить показатели, использованные другими авторами. На наш взгляд, имеет смысл говорить о пяти характеристиках, которые рассчитываются по показателям вживленных термодатчиков и по результатам замеров температуры в полевых условиях.
Индивидуальная высшая максимальная температура (определяется по данным логгеров) – самое большое значение, зафиксированное у отдельной особи за время наблюдений; для разных лет наблюдений и разных особей получено всего 15 таких значений. Средняя высшая максимальная температура – средняя из высших индивидуальных значений для всех особей. Высшая максимальная температура – это такое значение, при котором прекращается жизнедеятельность животного. На диаграмме распределения значений температуры эта точка ограничивает последний частотный класс (рис. 1). По этой причине показатель плохо статистически обеспечен – объем выборки для отдельной особи ограничен числом наблюдаемых солнечных дней.
Рис. 1.
Распределение значений температуры тела самки массой 115 г за весь период наблюдений: 1 – типичная максимальная температура, 2 – высшая максимальная температура, n – число наблюдений в интервалах с шагом 2°С.
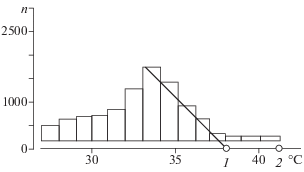
Типичная максимальная температура – значительно более репрезентативный показатель, который можно получить при анализе распределения значений (рис. 2). Его количественная оценка – точка пересечения оси температур с линией тренда падения частот в правой ветви распределения. Эта точка отсекает правый “хвост” и статистически обеспечена примерно 2000 отсчетов. Этот показатель близок к высшей температуре – среди всех наблюдений температура тела гадюки практически никогда не поднимается выше типичной максимальной. Однако у этого типичной максимальной температуры есть важное преимущество – она строго статистически обоснована и имеет очень низкую изменчивость. Это позволяет выполнять строгие статистические сравнения значений, рассчитанных для разных групп животных, различающихся по габитусу, статусу, району обитания и видовой принадлежности.
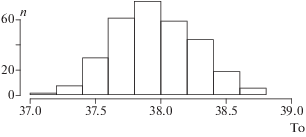
Технология оценки типичной максимальной температуры (То) состоит в построении линейной зависимости средней для класса температуры тела от частоты ее встречаемости: T' = a + b*F (где T ' – расчетная температура в данном классе, F – частота в классе). Из уравнения следует, что температура равна коэффициенту a при частоте, равной 0; это и есть точка пересечения линии регрессии с осью абсцисс; искомый параметр равен коэффициенту a. Поскольку нас интересует только резкий правый скат гистограммы, следует ограничить диапазон температур. Левая граница назначается как первый класс правее модального. Чтобы не вносить момент произвольности в назначение правой границы, ее можно не назначать, но при расчете уравнения линейного тренда нужно использовать взвешивание – тогда наиболее репрезентативные температуры из классов с большими частотами получат преимущество перед классами с экстремально высокими температурами (но низкими частотами) и “хвост” не будет влиять на изучаемый тренд. Расчет коэффициентов регрессии выполнялся в среде R (lm(t~f,weight=f )) . Для оценки устойчивости полученных оценок типичного максимума применена процедура рандомизации (Мастицкий, Шитиков, 2014), которая состоит в многократном пересчете коэффициентов регрессии на ограниченных выборках, случайно взятых из исходной выборки. Всего для одной особи гадюки были получены около 4000 отсчетов температуры тела выше 30°С. Из этой совокупности случайным образом извлекали выборки объемом по 500 отсчетов, для которых строили гистограмму, подсчитывали частоты классов, строили уравнение регрессии (Т ~ n) и оценивали То. Если повторить процедуру сотни раз, то можно построить распределение показателя (рис. 2) и рассчитать дисперсию.
Средняя пиковая максимальная температура тела (используются замеры логгерами) – показатель, рассчитанный по температурным пикам для всех змей. Показатель призван дать статистическую характеристику “обычной” максимальной температуре. Поиск пиковых значений (локальных максимумов) велся программой (на R) по следующему алгоритму. Для лага длиной от 3 до 15 соседних записей рассчитывалась скользящая линейная регрессия – положительные значения коэффициента регрессии говорят о росте температуры на анализируемом отрезке (левый склон пика), отрицательные – о снижении (правый склон); на пике происходит смена знака коэффициента регрессии, эту точку и брали в расчет. Оказалось, что при лаге длиной 9–11 отсчетов идентифицируются все визуально хорошо выраженные пики. Наборы пиковых значений служили для расчета статистического параметра – средней максимальной температуры. Всего обнаружено и включено в расчет 243 значения. Количество пиков, обнаруженных для дневного времени (9–17 ч) при ясной солнечной погоде составляет 9% от всех дневных замеров.
Средняя полевая максимальная температура тела – это средняя температура, рассчитанная по 9% наибольших значений от всех замеров, выполненных в полевых условиях при ясной солнечной погоде. Объем выборки максимальных значений составил 35 значений.
Индивидуальная максимальная добровольная температура – средняя величина, рассчитанная по ряду значений температуры тела, при котором у конкретной особи включается поведенческая терморегуляция, направленная на избегание перегрева. Алгоритм был следующим: в базе данных отыскивались периоды времени (продолжительностью 26 мин или 6 соседних замеров температуры тела), в течение которых змея нагревалась, а затем начинала остывать. Далее определяли, двигалась ли гадюка в этот период. Если животное не двигалось и не меняло позу, то изменение температуры связывали с погодными эффектами (ветер, тень от облака), и такие данные не учитывали. Если же в течение этого периода змея перемещалась или меняла позу, то это поведение рассматривали как терморегуляторную реакцию и в качестве максимальной добровольной температуры брали температуру тела, наблюдаемую именно в момент осуществления поведенческого акта (Коросов, 2010). Важно отметить, что зачастую после начала перемещений температура тела продолжала подниматься и формировала температурный пик, который нужно рассматривать как “добровольный перегрев” (Черлин, 2015). Спустя 2–4 мин температура падала и стабилизировалась. Всего для 16 гадюк было обнаружено 56 значений максимальной добровольной температуры. По всем этим значениям рассчитаны средняя максимальная добровольная температура и другие статистические параметры.
Помимо этих показателей, в тексте приведены значения границ размаха и средние по выборкам.
РЕЗУЛЬТАТЫ
Общие характеристики температуры тела гадюки, полученные разными методами, различаются (табл. 1). При любой погоде логгеры дают более широкий диапазон значений (минимальные – меньше, а максимальные, они же высшие, – больше), чем измерения термометром. Логгеры показывают, что иногда и при пасмурной погоде гадюки умудряются воспользоваться тепловым потоком от скрытого солнца, чтобы довести температуру тела до максимальных значений (36.9°С), которые оказываются одинаковыми при любой погоде. Другое дело – расчеты по показаниям ртутных термометров. И высшие максимальные оценки, и средние значения в пасмурную погоду и при переменной облачности ниже, чем в ясную погоду. Причина понятна – отсутствие постоянного теплового потока, из-за чего гадюка не может поддерживать высокую температуру длительное время. Единичные и случайные по времени измерения ртутным термометром просто не попадают на моменты, когда температура тела змеи достигает наибольших значений. Таким образом, простые средние показатели ртутных термометров характеризуют не свойства вида (физиологические характеристики), но свойства среды (эколого-физиологические характеристики), а значит, рассчитывать и сопоставлять их бессмысленно. По этой причине ниже сопоставляются данные, полученные только при ясной погоде (всего за 9 дней).
Таблица 1.
Температура (°С) тела гадюки при разных погодных условиях в мае–июне
| Параметр | Ясно | Переменно | Пасмурно | |||
|---|---|---|---|---|---|---|
| ртутный термометр | логгер | ртутный термометр | логгер | ртутный термометр | логгер | |
| Минимальная температура | 14.2 | 1.4 | 14.5 | 4.6 | 17.1 | 3.7 |
| Средняя температура | 24.5 | 26.1 | 23.3 | 25.1 | 19.8 | 13.5 |
| Максимальная температура | 33 | 40.4 | 31.9 | 38.6 | 24.1 | 25.7 |
| Объем выборки | 371 | 20 160 | 304 | 23 040 | 29 | 5760 |
Максимальная переносимая температура
Исследование верхнего порога толерантности нами не проводилось. Однако температурные датчики зарегистрировали рекордно высокий уровень наружной температуры покровов живой гадюки – 40.4°С, что существенно выше общепринятого порога выживания 37°С. Очевидно, максимальная пороговая температура еще выше.
Высшая максимальная температура
Высшие значения температуры тела, измеренные логгерами, для разных гадюк составляют ряд от 36.3 до 40.4°С (рис. 3), средняя высшая максимальная температура равна 38.8 ± 0.9°С. На сегодняшний момент это наиболее репрезентативная оценка максимальной температуры тела обыкновенной гадюки в природе. Довольно высокие показатели отмечаются другими авторами (Литвинов и др., 2016) – 38.2°С, но эти наблюдения ограничены одной особью. Поскольку все животные, участвующие в опытах, выжили, такая температура не может считаться критической. Значит, названый ранее температурный предел выживания гадюки в 37°С (Чан–Кьен, 1967) таковым не является.

Выборочная максимальная температура
Следующий наш шаг состоит в оценке пригодности полевых замеров температуры тела для статистически корректного расчета максимальной температуры. Со статистической точки зрения единственное наибольшее значение не может надежно характеризовать максимальный уровень переменной, поскольку такое значение может быть случайным. Достаточно надежной может быть только оценка, опирающаяся на выборку, для которой можно рассчитать квантили, например, для 95% (для наибольшего значения порог составит 97.5%). Условным статистически обоснованным показателем максимальной температуры тела можно считать величину, отстоящую от средней максимальной M на 1.96 S (M – средняя арифметическая из серии максимальных температур, S – их стандартное отклонение). Рассчитывать условную максимальную границу следует по выборке температур, признанных максимальными, например, по выборке таких значений, опубликованных разными авторами. Другой вариант получения выборки максимальных температур – регистрация пиковых (локальных максимальных) температур, полученных логгерами. Третий путь – использовать некоторую часть самых высоких значений из всех имеющихся температур.
Используя разные источники (Литвинов и др., 2016; Чан Кьен, 1967; Юмашев, 1995; Песков, 2003; Литвинов, 2004; Rutskina et al., 2009; Saint Girons, 1978), получаем ряд значений, которые авторы называют максимальными: 31.9, 34, 34.5, 35.5, 37.25, 38, 40°С. Эти значения не совпадают друг с другом и не соответствуют истинно максимальной (высшей) температуре тела, что говорит о невозможности статистическим путем получить корректные оценки максимальной температуры. Все значения, определенные ранее разными авторами как максимальные, – не несут этого содержания и не имеют смысла для сравнения по разным частям ареала, сезона или погоды.
Типичная максимальная температура
Показатель обладает устойчивостью и имеет очевидную биологическую интерпретацию. Он рассчитан по достаточно обширной выборке, не зависит от случайных флуктуаций исходных данных и должен коррелировать с физиологическим параметром терморегуляции. Значения температуры выше максимальной типичной имеют крайне низкую частоту встречаемости, такие значения являются случайными и относятся к “недобровольному перегреву”, поэтому показатель близок к истинной видовой максимальной температуре. Индивидуальные варьирования показателя укладываются в диапазон от 37.1 до 38.8°С и значимо не различаются; ошибка варьирована в диапазоне от 0.1 до 0.3°С. Обобщенная оценка показателя составила Т = 37.9 ± 0.24°С.
Добровольная максимальная температура
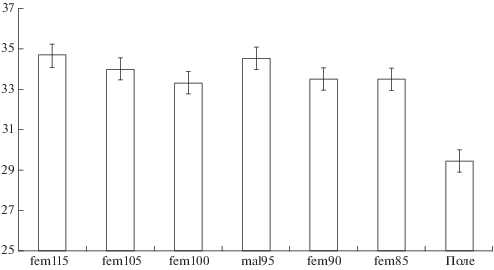
В период наблюдений при ясной погоде для разных змей было отмечено от 30 до 80 пиковых максимумов (выше 30°С), и среди них доля случаев с явным терморегуляторным поведением составила 30%. Наборы значений для всех змей давали более или менее симметричное унимодальное распределение в диапазоне 32–40°С (рис. 4). Средние значения разнились у разных особей до 1.5°С (от 33.3 до 34.8°С), но значимо по критерию Стьюдента не различались. Общая максимальная добровольная температура составляет M = 33.9°С, S = 1.8. В целом, наши данные подтверждают выводы Черлина (2012) о стабильности ключевых терморегуляторных параметров вида (рис. 5). Изучаемый нами эколого-физиологический параметр “добровольная максимальная температура” оказался независимым от пола, возраста, размеров тела и стадии созревания половых продуктов.
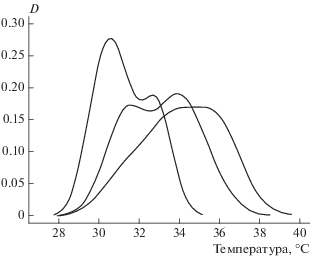
ОБСУЖДЕНИЕ
Сравним полученные нами характеристики терморегуляции обыкновенной гадюки с данными других исследователей, использующих вживляемые датчики. Обычно рассчитываются диапазон, стандартное отклонение и средняя арифметическая диапазона оптимальных или предпочитаемых температур тела (“preferred body temperature”). Естественно, нас интересуют максимальные значения указанных диапазонов, как крайнее значение, свободно выбираемое гадюкой, температура выше этого ей дискомфортна и избегается, следовательно, крайнее значение диапазона оптимальных температур и есть максимальная добровольная температура. Однако техника расчета крайних предпочитаемых температур цитированных авторов существенно отличается от оценки максимальных добровольных и дает большую ошибку. Если различие между оценками границы оптимума у разных авторов составляет 3°С (32–34.6°С), то наши оценки максимальной добровольной отличаются на 1 градус (33.5–34.6°С). Полученные авторами средние значения предпочитаемых температур примерно на 1–2 градуса ниже оцененной нами добровольной максимальной температуры (33.9°С): Lim = 30.5–33.9°С (4 экз.) (Saint–Girons, 1975), Lim = 31.7–33.8°С (9 экз.) (Herczeg et al., 2007), Lim = 32.69–34.66°С, (12 экз.) (Lourdais et al., 2013), Lim = 26–32°С (Литвинов, Ганщук, 2007). Отметим, во-первых, что некоторые из этих значений практически равны нашей оценке (33.9°С), но, во-вторых, другие существенно отличаются (32 и 34.66°С) от нашей. Причина состоит в том, что при расчете верхней границы диапазона предпочитаемых температур авторы используют большой набор значений и получают оценку путем усекновения распределения с правой стороны. Следовательно, на полученную величину влияют значения, не имеющие никакого отношения к добровольному максимуму. В расчет добровольного максимума входят и значения температуры тела действительно в момент начала перегрева, и при перегреве в процессе поиска прохладных мест, и в укрытиях от прямых лучей, и при охлаждении на прохладном субстрате, и при разном положении тела относительно солнца (различные позы). Соответственно, и диапазон и средние значения для этого диапазона будут варьировать в зависимости от условий наблюдения, региона и т.д. Т.е. эти характеристики имеют все пороки статистических показателей и сопоставлять можно только корректно оцененные добровольные максимальные температуры как физиологический параметр терморегуляции, а сравнения нескольких выборок “преферентных” температур не могут привести к содержательным выводам. Возможно, единственным выходом для сравнения наборов температурных характеристик рептилий разных видов и с разным статусом является расчет устойчивых типичных максимальных температур по частотному распределению. Их оценка не требует видеонаблюдения и снабжения ошибкой для статистических сравнений.
В нашем представлении максимальная добровольная температура тела (33.9°С) – это видоспецифический “штатный” терморегуляторный параметр, защищающий особь от избыточного перегрева. Если текущая температура тела превышает эту величину, запускаются поведенческие терморегуляторные акты – перемещение в более прохладные области, например в полутень, тень, на холодный субстрат, на обдуваемый участок, в воду и пр. При этом в процессе реализации этого поведения температура тела может еще возрасти, даже до уровня, угрожающего жизни. В наших наблюдениях типичным был случай, когда температура поверхности тела гадюки дополнительно увеличивалась до 36–40°С; в среднем до 37.5°С. Однако вскоре за этим неизменно наблюдалось падение температур ниже критических как следствие выбора змеей более благоприятных, прохладных условий среды. Это описание ясно показывает, что для оценки значений максимальной добровольной температуры как физиологического параметра нельзя брать максимальные значения температуры тела, которые оказываются не типичными и устойчивыми характеристиками, но сиюминутными, быстро проходящими и, очевидно, неприятными для животного. К сожалению, определить добровольную максимальную температуру достаточно сложно, поскольку для этого требуется расшифровка видеозаписей и синхронизация и с показаниями логгеров (возможно, это еще один из источников случайной изменчивости). Интересно было бы предложить показатель, имеющий тесную связь с максимальной добровольной температурой, но имеющий более простую технологию оценки. Таким показателем, на наш взгляд, является максимальная типичная температура.
Для расчета типичной максимальной температуры каждой особи достаточно составить распределение показателей логгеров и построить простую линейную регрессию. Показатель максимальной типичной температуры имеет высокую степень корреляции с максимальной добровольной температурой (r = 0.8), превышая добровольную на 4°С (37.9 против 33.9°С). Смысловое содержание и статистическая обоснованность типичной максимальной температуры позволяет сравнивать особей из разных частей ареала и разных видов. Возможно, максимальная типичная температура имеет тот же биологический смысл, что и предел выживания, поскольку такое значение выживаемости приводится в литературе 38.1°С. Это было бы большой удачей, поскольку оценка летальных температур не потребует острых экспериментов.
Возможности оценить максимальные температуры имеют ограничения. Не могут приниматься во внимание максимальные значения температуры тела, которые регистрируются в пасмурную погоду или при переменной облачности. В условиях недостаточной инсоляции змея в принципе не может достичь такой температуры тела, которая будет означать начало перегрева. Такие “максимальные” значения характеризуют в первую очередь погодные условия, но не физиологически значимый параметр терморегуляции. В результате исследований указанных экологических явлений – недобровольный перегрев и недостаточное нагревание (а также методические погрешности) – в печати обнаруживается широкий диапазон оценок максимальной температуры, в равной мере неприемлемых для сравнения.
Важно отметить, что нами наблюдения выполнялись только в Карелии и в основном в течение мая и июня, и одна из актуальных задач состоит в оценке корректных показателей в остальные периоды теплого сезона и в других точках ареала. В литературе можно найти данные об изменении средней температуры тела гадюки в течение лета, а также о региональных различиях этого показателя (Четанов, 2010). Объективная причина этого феномена состоит, как можно догадаться, в уменьшении возможностей для баскинга гадюк из-за сезонного сокращения фотопериода и потока солнечного тепла. В этом случае средняя температура тела должна рассматриваться только как косвенная метеорологическая метрика, но не как характеристика биологических свойств вида в данный сезон или в данной местности. Как показали наши наблюдения, несмотря на ухудшение температурных условий для баскинга при сезонном и региональном изменении уровня инсоляции, ключевая физиологическая константа терморегуляции (максимальная добровольная температура) остается неизменной в течение сезона и, теоретически, сохранится неизменной на всем ареале.
Каков же источник изменчивости данного показателя в наших наблюдениях, ориентированных на поиск “истинных” физиологических параметров терморегуляции? Видимо, можно назвать следующие источники изменчивости. Техническая погрешность может возникнуть в двухминутный период между моментами регистрации температуры тела змеи. За это время змея может изменить температуру тела на 1–1.5°С, что неизбежно скажется на точности оценок. Мы также не можем полностью исключить влияние на этот параметр физиологического статуса животного (пол, беременность, сытость, процесс размножения, брачные игры), хотя наши текущие наблюдения и не выявили такой зависимости. Вне данного эксперимента мы не раз замечали изменение терморегуляторного поведения у больных и истощенных или недавно травмированных животных. Наконец, как и для любого физиологического параметра, для максимальной добровольной температуры должна быть характерна некоторая индивидуальная изменчивость.
ЗАКЛЮЧЕНИЕ
1) Для пойкилотермной рептилии понятие “максимальная температура тела” оказывается многокомпонентным, оценивается и в полевых условиях, и в лабораторных условиях. Можно говорить о пяти видах витального показателя “максимальная температура тела”.
2) Высшая максимальная температура тела определяется как самая высокая температура для конкретной особи гадюки, зафиксированная логгером на длинных рядах наблюдений в ясный летний день. Средняя для нескольких индивидуальных значений достаточно точно характеризует типичный видовой порог чувствительности к внешнему потоку тепла и составляет 38.8 ± 0.9°С.
3) Выборочные показатели максимальной температуры тела определяются как обобщение серий экстремальных значений, наблюдаемых в поле или эксперименте. Результаты широко варьируют в зависимости от выбранной методики определения и условий наблюдения (в поле или эксперименте). В целом, выборочные показатели не годятся для сравнения особенностей терморегуляции внутривидовых групп, сезонных изменений максимальной температуры тела, максимальной температуры тела особей из разных частей ареала.
4) Добровольная максимальная температура фиксируется логгерами в момент реализации терморегуляторной реакции и представляет собой стабильный физиологический видовой порог реактивности особи. Изменчивость показателя невелика, различия средних значений для разных особей не превышают 2°С и статистически не значимы. Среднее значение составляет 33.9°С (S = 1.8).
5) Типичная максимальная температура – устойчивая характеристика правой ветви частотного распределения температур. Простая технология расчета исключает появление случайных высших значений, поэтому показатель близок к истинной видовой максимальной температуре и коррелирует (r = 0.8) с максимальной добровольной температурой тела, превышая ее на 4 градуса. Простая биологическая интерпретация и низкая изменчивость делают его удобным при сравнении рептилий из разных частей ареала и разных видов.
Список литературы
Банников А.Г., Даревский И.С., Ищенко В.Г., Рустамов А.К., Щербак Н.Н., 1977. Определитель земноводных и пресмыкающихся фауны СССР. Москва: Просвещение. 415 с.
Коросов А.В., 1988. К экологии обыкновенной гадюки (Vipera berus L.) в Карелии // Фауна и экология наземных позвоночных. Петрозаводск. С. 15–23.
Коросов А.В., 2010. Экология обыкновенной гадюки (Vipera berus L.) на Севере (факты и модели). Петрозаводск: Изд-во ПетрГУ. 264 с.
Коросов А.В., Горбач В.В., 2017. Компьютерная обработка биологических данных. Петрозаводск: Изд-во ПетрГУ. 96 с.
Литвинов Н.А.,Ганщук С.В., 2007. Термобиология обыкновенной гадюки (Vipera berus, Reptilia, Serpentes) в Волжском бассейне // Известия Самарского научного центра Российской академии наук. Т. 9. № 4. С. 950–957.
Литвинов Н.А., Панова М.К., Окулов Г.А., 2016. Изучение терморегулирующего поведения рептилий методом имплантации регистраторов температуры // Вестник СПбГУ. Сер. 3. Вып. 3. С. 69–76.
Литвинов Н.А., 2004. Термобиологические исследования // Бакиев А.Г., Гаранин В.И., Литвинов Н.А., Павлов А.В., Ратников В.Ю. Змеи Волжско-Камского края. Самара: Изд-во Самарского научного центра РАН. С. 109–146.
Мастицкий С.Э., Шитиков В.К., 2014. Статистический анализ и визуализация данных с помощью R. Электронная книга, адрес доступа: http://r–analytics.blogspot.com
Песков А.Н., 2003. Гадюки (Serpentes, Viperidae, Vipera) Волжского бассейна: фауна, экология, охрана и прикладное значение. Дис. … канд. биол. наук. Тольятти: ИЭВБ РАН. 169 с.
Пестов М.В., Маннапова Е.И., 1999. Тепловой шок как причина гибели обыкновенных гадюк в естественных условиях // Актуальные проблемы герпетологии и токсинологии. Сб. науч. трудов. Вып. 3. Тольятти. С. 87–89.
Чан Кьен, 1967. Систематика и экология обыкновенной гадюки Vipera berus (Linné, 1758). Автореф. дис. … канд. биол. наук. Л.: ЛГУ. 14 с.
Черлин В.А., 1991. Методы исследований по термобиологии пресмыкающихся // Экология животных Узбекистана. Ташкент. С. 70–97.
Черлин В.А.,2012 Термобиология рептилий. Общая концепция. СПб.: Изд-во “Русско-Балтийский информационный центр “Блиц”. 362 с.
Черлин В.А., 2013. Сложности и возможные ошибки при полевых исследованиях по термобиологии рептилий // Современная герпетология: проблемы и пути их решения. Статьи по материалам докладов Первой международной молодежной конференции герпетологов России и сопредельных стран (Санкт-Петербург, Россия, 25–27 ноября 2013 г.). Зоологический институт РАН. СПб. 2013. С. 32–39.
Черлин В.А., 2015. Тепловые адаптации рептилий и механизмы их формирования // Принципы экологии. № 1. С. 17–76.
Четанов Н.А., 2010. Статистический анализ влияния некоторых абиотических факторов на температуру тела пресмыкающихся Пермского края. Автореф. дис. … канд. биол. наук. Тольятти. 20 с.
Юмашев И.Ю., 1995. Термобиологические показатели обыкновенной гадюки в бассейне Верхней Волги // Первая конференция герпетологов Поволжья. Тез. докл. Тольятти. С. 63–65.
Herczeg G., Saarikivi J., Gonda A., Perälä J., Tuomola A., Merilä J., 2007. Suboptimal thermoregulation in male adders (Vipera berus) after hibernation imposed by spermiogenesis // Biological Journal of the Linnean Society. № 92. P. 19–27.
Lourdais O.,Guillon M., DeNardo D., Blouin–Demers G., 2013. Cold climate specialization: Adaptive covariation between metabolic rate and thermoregulation in pregnant vipers // Physiology & Behavior. № 119. P. 149–155.
Rutskina I.M., Litvinov N.A., Roshchevskaya I.M., Roshchevskii M.P., 2009. Temperature Adaptation of the Heart in the Grass Snake (Natrix natrix L.), Common European Viper (Vipera berus L.), and Steppe Viper (Vipera renardi Christoph) (Reptilia: Squamata: Serpentes) // Russion Journal of Ecology. V. 40. № 5. P. 314–319.
Saint–Girons H., 1975. Observations preliminaires sur la thermoregulation des Viperes d’Europe // Vie et Milieu. V. 25. Fasc. 1. Ser. C. P. 137.
Saint–Girons H., 1978. Thermoregulation comparee des viperes d’Europe. Etude biotelemetriaue // Terre et vie. № 3. P. 417–440.
Дополнительные материалы отсутствуют.
Инструменты
Зоологический журнал