

Зоологический журнал, 2021, T. 100, № 4, стр. 419-433
Строение, теплозащитные свойства и линька волосяного покрова обыкновенной бурозубки (Sorex araneus)
a Петрозаводский государственный университет
185910 Петрозаводск, Россия
b Карельский научный центр РАН
185035 Петрозаводск, Россия
* E-mail: Ivanter@petrsu.ru
Поступила в редакцию 10.07.2020
После доработки 04.08.2020
Принята к публикации 16.08.2020
Аннотация
На основании исследований строения волосяного покрова, его удельной теплопроводности и микроструктуры стержня волоса, выполненных на 86 шкурках обыкновенной бурозубки, выявлены приспособительные особенности ее шерсти, связанные с норно-наземным образом жизни в условиях постоянного недостатка тепла. Обитая преимущественно в лесной подстилке и ходах неглубоких нор, но нередко появляясь и на поверхности почвы, зверьки этого вида приобрели в процессе эволюции легкий, не мешающий движениям, но достаточно “теплый”, с оптимальными теплоизоляционными свойствами волосяной покров, характеризующийся умеренной густотой и длиной волос, неравномерностью опушения отдельных частей тела, булавовидной конфигурацией апикального сегмента волосяного стержня, слабой извитостью основания волоса и несколько более сильным, чем у волос из других групп млекопитающих, развитием сердцевинного канала. Несмотря на обильную жировую смазку, повышенную прочность волос и отсутствие выраженного ворса (отсюда способность шерсти сгибаться и укладываться в любом направлении), волосяной покров у землероек из-за постоянного соприкосновения с почвой довольно быстро вытирается. Его своевременному восстановлению служат так называемые дополнительные линьки и, в частности, два следующих весной друг за другом процесса смены шерсти, различных по характеру, срокам, направлению и топографии вектора.
Изучение приспособительных особенностей волосяного покрова землероек представляет большой научный интерес, и прежде всего с позиций эволюционной морфологии, экологической физиологии и таксономии мелких млекопитающих. Обитание этих животных в лесной подстилке и ходах неглубоких нор в условиях постоянной нехватки тепла, а также общее несовершенство терморегуляции способствовало выработке в процессе их эволюции целого комплекса экологических и морфофизиологических приспособлений, направленных на сохранение оптимального энергетического баланса организма со средой при низких и крайне неустойчивых окружающих температурах (Ивантер и др., 1985). Отсюда важность подробного исследования таких адаптивных механизмов физической терморегуляции зверьков, как строение и теплозащитные свойства шерсти, микроструктура волос, географическая, сезонно-возрастная и популяционная изменчивость этих показателей, выявление особенностей строения шерстного покрова, связанных с обитанием в специфических температурных условиях приземного слоя воздуха и т.п. Надо учитывать также и общую слабую изученность данного вопроса, особенно применительно к мелким насекомоядным млекопитающим таежного Севера.
Имеющиеся же в отечественной и зарубежной литературе сведения о строении волосяного покрова обыкновенной бурозубки (далее – бурозубка) не только далеко не полны, но и во многом противоречивы. Если Гудкова-Аксенова (1951) выделяет у этого вида только две категории волос, Соколов и Чернова (Соколов, 1973; Соколов и Чернова, 1998; Чернова, Целикова, 2004) – четыре, а Фогель и Кёпхен (Vogel, Köpchen, 1978) и Хуттерер и Хёртер (Hutterer, Hürter, 1981) – три, то по мнению Боровского (Borowski, 1973) и Скарена (Skaren, 1973), у землероек-бурозубок волосы одного и того же типа способны принимать разный вид в зависимости от конфигурации расширенной апикальной части стержня (концевой гранны). Отрицая дифференцировку волос у землероек-бурозубок на направляющие, остевые и пуховые, Боровский обращает внимание на различную степень их сегментации и в связи с этим выделяет “… следующие 3 типа волос: 1) летние волосы (S) с четырьмя сегментами, иногда с коротким кончиком (пятый короткий сегмент); 2) весенние волосы (Sp) с пятью сегментами, но иногда с шестым сегментом (около 5% волосков); 3) зимние волосы (W) – шестисегментные, иногда с дополнительным седьмым концевым сегментом…” (с. 249). В действительности же, как будет показано ниже, летняя, весенняя и зимняя шерсть землероек состоит из направляющих, остевых и пуховых волос, а волосы с “дополнительным коротким концевым сегментом” есть не что иное, как пуховые. Что же касается характерной для мелких насекомоядных млекопитающих сегментации волос, то, в отличие от Боровского, мы считаем это не примитивным атавистическим признаком, а важной адаптацией к специфическим условиям существования. Здесь вполне уместно вспомнить замечание, содержащееся в статье Башкирова и Жаркова (1934), которые, говоря о пережимах на стержнях остевых и пуховых волос, свойственных представителям семейств Talpidae и Soricidae, предполагают, что эти структурные особенности имеют большое приспособительное значение. Благодаря такой сегментации волосы могут сгибаться в разные стороны, не ломаясь и не свойлачиваясь, что позволяет зверькам легко передвигаться в норах не только головой вперед, но и пятиться назад. Кроме того, такая структура меха обусловливает удерживание в его толще так называемого “инертного воздуха”, придающего волосяному покрову зверьков наилучшие теплозащитные свойства.
МАТЕРИАЛ И МЕТОДИКА
Материалом для наших исследований послужили 86 шкурок землероек, отловленных в разные сезоны преимущественно в южных районах Карелии. Изучение расположения волос на шкурке, строения микроструктуры стержня отдельного волоса, его толщины, плотности и высоты, дифференцировки и топографии шерсти, а также сроков и последовательности линьки проводили по общепринятым методикам (Церевитинов, 1951; Кузнецов, 1952; Borowski, 1952; Соколов, 1973; Соколов и др., 1986; Чернова, Целикова, 2004). Микроструктуру волоса исследовали под оптическим микроскопом при увеличении 15 × 90 (иммерсионный объектив). Отпечатки кутикулярного слоя волос делали на парфюмерном лаке № 1 (Хмелевская, 1965). Определение теплоизоляционных свойств шкурок (по удельной теплопроводности) выполняли с помощью специального прибора ИТ-З, изготовленного по нашему заказу для измерения теплопроводности небольших по размеру неметаллических объектов в Киевском институте технической теплофизики АН УССР. Существенное преимущество этого прибора заключается в большой чувствительности и точности, а также в том, что все необходимые для расчета теплопроводности величины получают непосредственным измерением. Кроме того, для установления сроков и стадий линьки по состоянию мездры и меха просмотрено (в том числе в бинокулярный микроскоп) более 600 сухих коллекционных шкурок и привлечены данные о линьке, содержащиеся в картотеке лаборатории зоологии КарНЦ РАН.
При изучении хода и топографии сезонной линьки применяли метод “картирования” – зарисовки на карточках с трафаретами контура шкурки изменений окраски мездры и расположения линных пятен. В световом микроскопе определяли функциональное состояние наружных покровов, дифференцировали зрелые волосы от растущих, различали пигментированные и непигментированные участки закладки волоса у изученных объектов кожи и волос. Необходимость световой микроскопии была вызвана тем, что только с ее помощью можно, например, безошибочно отличить растущий зимний волос от окончательно выросшего летнего той же длины. Для этого достаточно рассмотреть под бинокулярным микроскопом строение волосяной луковицы: у зрелых волос она закрытого (колбовидного) типа, а у растущих – открытого (сосочкового). Кроме того, закладка волос не везде сопровождается потемнением мездры. Поскольку волосы землероек имеют зонарную окраску и их концевые сегменты на разных участках шкурки неодинаковы по цвету (на спине – темно-коричневые, на боках – бурые, а на брюхе – белые), то на боках и спине линные пятна бывают видны сразу, а на брюхе – лишь после того, как пробьются через кожу белые верхушки волос. Сегменты же стержня, лежащие ниже вершины, окрашены в черный цвет. Поэтому при микроскопии кожи с брюшка без видимых признаков линьки легко обнаруживаются изменения (в частности, функционирующие волосяные фолликулы), свидетельствующие о слабо заметной сезонной смене волос.
Трудности в изучении линьки землероек связаны также с разновозрастным составом популяции. Продолжительность линьки одного зверька, как известно, невелика (Borowski, 1958). Но у всей популяции этот процесс растянут, и в один и тот же период в ее составе встречаются особи, находящиеся на разных стадиях линьки. Отсюда и неизбежные сложности в определении последовательности в топографии сезонной смены шерсти.
Следует особо остановиться на правомерности или, скорее, уместности использования при описании архитектоники волос землероек терминов “сегмент” и “перетяжка”. Дело в том, что хотя в световом микроскопом волосы землероек и кротов выглядят сегментированными, т.е. состоящими из более широких участков – сегментов и узких участков – перетяжек, в действительности это только кажущийся, чисто зрительный эффект. Если при изготовлении препарата волос случайно раскручивается, то хорошо видно, что он представляет собой сплошную постепенно расширяющуюся ленту, которая в нескольких местах поворачивается вдоль оси на 180°. Число таких поворотов строго определенное для каждой категории волос и изменяется посезонно. Впечатление же сегментированности создается за счет закономерно повторяющихся при поворотах расширений и сужений сердцевинного канала и неравномерности распределения в нем пигмента. Упоминание об этом мы находим и в ряде предшествующих публикаций (Keller, 1978; Hutterer, Hürter, 1981; Kapischke, Muhle, 1988; Соколов, Чернова, 1998).
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Общая характеристика волосяного покрова
Волосяной покров землероек своеобразен. Он относительно низкий, бархатистый, мягкий, ровный и густой. Волосы тонкие, растут одиночно, перпендикулярно к поверхности кожи и благодаря сегментированному строению легко укладываются в любом направлении.
Свойственная всем представителям семейств Soricidae и Talpidae сегментация волос характерна и для бурозубок. При этом сами сегменты, как правило, хорошо выражены и представляют собой утолщенные, интенсивно пигментированные участки волоса, располагающиеся под углом приблизительно 130° по отношению друг к другу. Сегментация характерна для всех типов волос, кроме направляющих, которые не играют существенной роли в образовании волосяного покрова и, видимо, в какой-то степени выполняют лишь поддерживающую и осязательную функции. Как уже указывалось, сегментное строение волос носит у землероек приспособительный характер. В местах сужения волосы согнуты под тупым углом и способны обеспечивать меху хорошие амортизирующие свойства. Благодаря этому волосяной покров не сминается и, действуя по принципу рессоры, выполняет функцию пружинящего каркаса, позволяя зверьку передвигаться в тесной лесной подстилке и в узких ходах нор.
На спине, боках и верхней стороне хвоста землероек-бурозубок шерсть насыщенного бурого или темно-шоколадного цвета с рыжеватым оттенком, а на брюшке и нижней части хвоста серовато-белого с серебристостью. Пигмент в волосах распределен неравномерно. Наиболее интенсивно окрашены расширенные части стержня волоса, а переходы между сегментами более светлые. Окраска шкурки в основном зависит от концентрации пигмента в концевых граннах остевых волос, которые несколько возвышаются над уровнем пуховых и прикрывают последние. В этих апикальных граннах располагается пигмент коричневого цвета. Такой оттенок объясняется наличием черного пигмента в нижележащих сегментах остевых и пуховых волос. На брюшке апикальные гранны остевых волос пигмента не содержат, поэтому в их окрасе нет бурых тонов, и низ тела выглядит серым. При этом клетки сердцевины волос брюшка в большей степени, чем на спине и боках, заполнены воздухом (табл. 1), что в значительной степени компенсирует относительно невысокие теплозащитные свойства редкого и короткого здесь волосяного покрова. Апикальные кончики волос брюшка бесцветны. Они не содержат пигмента и выглядят прозрачными, придавая брюшку характерный серебристый отблеск. Верхушки волос первыми снашиваются, обламываются, секутся, волосы иначе отражают свет, поэтому старая шерсть выглядит тусклой, а новая – блестящей и яркой.
Таблица 1.
Толщина сердцевинного слоя стержня волос обыкновенной бурозубки
| Категория волос | Летняя шерсть | Зимняя шерсть | ||||
|---|---|---|---|---|---|---|
| n | Толщина сердцевины, мкм | Доля от общей толщины волоса, % | n | Толщина сердцевины, мкм | Доля от общей толщины волоса, % | |
| Спина | ||||||
| Направляющий | 17 | 28.9 ± 0.5 | 57.9 | 16 | 18.7 ± 0.7 | 62.1 |
| Остевой I | 25 | 29.0 ± 0.3 | 58.0 | 20 | 19.0 ± 0.7 | 63.4 |
| Остевой II | 25 | 16.9 ± 0.3 | 62.4 | 20 | 17.9 ± 0.4 | 70.7 |
| Пуховой | 25 | 7.9 ± 0.1 | 63.0 | 24 | 5.5 ± 0.2 | 77.0 |
| Бок | ||||||
| Направляющий | 16 | 27.3 ± 0.4 | 54.6 | 15 | 18.9 ± 0.6 | 63.0 |
| Остевой I | 24 | 24.8 ± 0.2 | 49.9 | 16 | 19.6 ± 0.4 | 65.6 |
| Остевой II | 25 | 14.6 ± 0.2 | 56.1 | 18 | 17.6 ± 0.3 | 69.9 |
| Пуховой | 26 | 6.2 ± 0.1 | 63.1 | 21 | 5.8 ± 0.1 | 81.0 |
| Брюшко | ||||||
| Направляющий | 18 | 27.9 ± 0.5 | 56.1 | 17 | 19.8 ± 0.5 | 64.0 |
| Остевой I | 25 | 26.7 ± 0.4 | 53.8 | 24 | 19.5 ± 0.5 | 65.2 |
| Остевой II | 24 | 14.8 ± 0.3 | 52.9 | 26 | 17.8 ± 0.3 | 71.0 |
| Пуховой | 23 | 6.2 ± 0.2 | 61.0 | 22 | 6.1 ± 0.1 | 83.6 |
Дифференциация волос. Изучение структуры шерстного покрова и особенностей строения волос различных типов позволяет выделить у бурозубки 4 категории волос: направляющие, остевые I и II порядков и пуховые (рис. 1).
Рис. 1.
Особенности строения волосяного покрова обыкновенной бурозубки, Sorex araneus: a – дифференциация волос на категории, б – изменение высоты волос (мм) на топографически различных участках шкурки. Волос: 1 – направляющий, 2 – остевой I, 3 – остевой II, 4 – пуховой. Масштаб: а – 1 мм, б – 1 см.
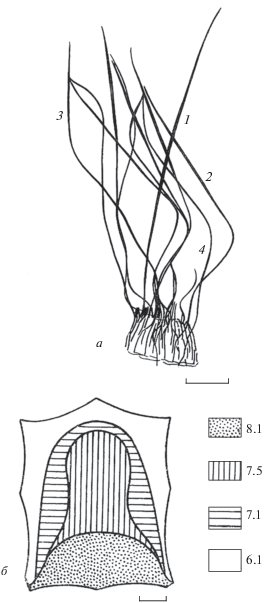
Наибольшей длиной, толщиной и упругостью отличаются направляющие волосы. Количество их незначительно (2–4 на 400–500 остевых и пуховых), но они заметно возвышаются над остальными и хорошо различимы даже невооруженным глазом. Стержень направляющего волоса упругий, почти прямой с веретенообразной, сильно растянутой гранной, без извитости, перетяжек и сегментов. Апикальный кончик постепенно утончается, он длиннее и тоньше кончика остевого волоса. Окрас направляющего волоса темно-коричневый, однотонный, а прикорневая часть и вершина бесцветны.
Стержень волоса состоит из трех слоев ороговевших клеток: кутикулярного, коркового и сердцевинного (рис. 2). Кутикулу направляющего волоса обыкновенной бурозубки, согласно классификации Кузнецова (1952), следует отнести к некольцевидному типу. Ороговевшие бесцветные чешуйки кутикулы на протяжении волоса закономерно меняются по конфигурации и размерам. В прикорневой части волосы покрыты удлиненными чешуйками с ровным округлым краем. По мере расширения волоса в гранну клетки кутикулы становятся короче, а в месте сужения стержня они опять сильно вытягиваются вдоль волоса, а затем сменяются очень короткими и широкими, с зазубренными краями на концевой части волоса. Чем шире гранна, тем шире и короче чешуйки и ярче выражена зазубренность края. Клетки кутикулы располагаются на поверхности волоса черепицеобразно, налегая друг на друга так, что их свободные края обращены к вершине волоса (рис. 2).
Рис. 2.
Микроструктура волоса обыкновенной бурозубки (по отпечаткам на лаке): а – кутикула, б – продольная бороздка на гранне стержня волоса, в – просвечивающие сквозь кутикулу пигментированные диски сердцевины в перетяжке стержня. Микрофото. Ув. ×600.

Как считает Кузнецов (1952), такое положение клеток чешуйчатого слоя (раструбом к вершине волоса) и наличие зазубрин на их апикальном свободном краю затрудняет скатывание капелек воды, попавших на волосяной покров, к коже. Кроме того, черепицеобразное расположение чешуек на корне волоса обеспечивает прочное прикрепление его к стенке волосяного влагалища, которое выстлано такими же по форме и размерам чешуйками, но края их направлены в противоположную сторону (Соколов, 1973). К этому следует добавить, что подобное расположение кутикулярных чешуй способствует правильной укладке волос, препятствуя их свойлачиванию, а также препятствует закреплению эктопаразитов.
Под кутикулой волоса располагается корковый слой, который просматривается по всей длине стержня и который образует внутренние стенки сердцевинного канала. Толщина сердцевины и толщина коркового слоя на протяжении волоса изменяются взаимосвязано, демонстрируя отрицательную корреляцию (табл. 1). По разделяемому нами мнению Кузнецова (1952), прочность волоса зависит в основном от коркового слоя, а не от общей толщины стержня. Что же касается пигмента, то, по данным Соколова (1973), корковый слой окрашенных волос содержит его в диффузной или агрегированной форме и поэтому зрительно почти бесцветен, как и чешуйчатый слой.
У бурозубок окрас и толщина стержня волоса зависят от строения и мощности сердцевины – размера слагающих ее клеток, наличия в них пигмента и воздухоносных полостей. Толщина сердцевины меняется по мере созревания волоса и неодинакова в различных его частях. Наибольшего развития этот слой достигает в гранне волос, где клетки сердцевины располагаются в виде разделенных воздушными полостями участков. Эти участки содержат пигментные глыбки, состоящие из зерен пигмента и “дисков” (в виде монетных столбиков или таблеток в тубе) (однорядная лестничная сердцевина). В самой широкой части гранны клетки сердцевины лежат в 2–3 ряда (двух-, трехрядная лестничная сердцевина), а в узких частях гранны – в один ряд. В тонких и изогнутых и покрытых самыми длинными чешуйками частях волоса (в местах сужения и в прикорневой части растущего волоса) сердцевина очень узкая, нитевидная, ее клеточная структура не выражена и пигмент просвечивает в виде отдельных зерен, расположенных гораздо реже, чем в сердцевинных клетках гранны. В самой же вершине и прикорневой части зрелого волоса сердцевина вообще отсутствует и, наоборот, очень хорошо развита и богата пигментом в корне и прикорневой части растущего волоса, особенно в начальный период роста, когда волосы зарождаются еще в толще кожи. Луковицы растущего волоса сильно пигментированы, и это объясняет темный цвет мездры линяющих зверьков (рис. 4). Этим объясняется и темный цвет мездры линяющих зверьков (рис. 3). Таким образом, наличие сердцевины в корне волоса свидетельствует о его росте, а отсутствие сердцевины – о зрелости и окончании роста волос.
Рис. 3.
Закладка и рост волос при сезонной линьке: а – появление пигмента, б – образование луковиц новых волос, в – растущие волосы в толще кожи. Микрофото. Ув. ×600.

Рис. 4.
Последовательность появления пигментных пятен на мездре во время сезонных линек: а – осенняя линька, б – весенняя первая, в – весенняя вторая. Масштаб 1 см.

Остевой волос короче и тоньше направляющего, но отчетливо сегментирован. Наиболее выражен апикальный ланцетовидный расширенный сегмент, образующий концевую гранну стержня – он длиннее и шире остальных участков и составляет около половины или по крайней мере четверть всей длины волоса. Для данной категории волос характерны повороты стержня в местах изгиба. Именно эти повороты и обеспечивают укладку волос в любом направлении.
Остевые волосы землероек бывают двух порядков:
Ость I порядка среди сегментированных волос – самая длинная и наиболее широкая в гранне, концевой сегмент ее занимает около 50% общей длины волоса. Отгибаясь и прикрывая в виде зонтика подшерсток, апикальные сегменты ости способствуют основной механической защите подпуши и формированию воздушного теплозащитного слоя. Зимой ости шестисегментные, летом – трехсегментные (имеются в виду зрелые волосы). Отличительная черта остевых волос землероек, как уже упоминалось выше, — наличие продольных бороздок по боковым сторонам апикального (концевого) сегмента (рис. 2). В кончике волоса и в местах сгибов и продольных поворотов волоса эти бороздки замкнуты. Они характеризуются разной глубиной и конфигурацией, приобретающей на поперечном срезе форму латинской буквы H (“H-профиль”), что имеет таксономическое и адаптивное значение (Hutterer, Hürter, 1981).
Ость II порядка несколько короче и тоньше предыдущего, на концевой сегмент его приходится 25–27% от общей длины стержня. Как зимой, так и летом эти волосы содержат на один сегмент больше: зимой они семисегментные, а летом — четырехсегментные (табл. 2 и 3).
Таблица 2.
Характеристика летней шерсти обыкновенной бурозубки
| Категория волос | n | Число волос на 4 мм2 | Длина волоса, мм | Толщина волоса, мкм | Число сегментов |
|---|---|---|---|---|---|
| Спина | |||||
| Направляющий | 17 | 3.6 ± 0.2 | 5.1 ± 0.06 | 50.0 ± 0.3 | 1 |
| Остевой I | 25 | 114.0 ± 4.0 | 4.2 ± 0.01 | 50.0 ± 0.4 | 3 |
| Остевой II | 25 | 89.8 ± 5.7 | 4.0 ± 0.05 | 27.2 ± 0.4 | 4 |
| Пуховой | 25 | 235.3 ± 7.1 | 3.6 ± 0.02 | 12.6 ± 0.1 | 4 |
| Бок | |||||
| Направляющий | 16 | 2.3 ± 0.1 | 4.7 ± 0.03 | 49.9 ± 0.5 | 1 |
| Остевой I | 24 | 94.6 ± 5.7 | 4.3 ± 0.04 | 49.7 ± 0.6 | 3 |
| Остевой II | 25 | 66.3 ± 6.5 | 4.1 ± 0.09 | 26.1 ± 0.4 | 4 |
| Пуховой | 26 | 232.4 ± 5.7 | 3.4 ± 0.01 | 9.8 ± 0.2 | 4 |
| Брюшко | |||||
| Направляющий | 18 | 2.0 ± 0.1 | 4.4 ± 0.04 | 49.8 ± 0.6 | 1 |
| Остевой I | 25 | 169.2 ± 6.7 | 4.1 ± 0.04 | 49.7 ± 0.4 | 3 |
| Остевой II | 24 | 83.6 ± 4.9 | 3.6 ± 0.03 | 28.0 ± 0.3 | 4 |
| Пуховой | 23 | 224.6 ± 7.1 | 3.3 ± 0.01 | 10.2 ± 0.1 | 4 |
Таблица 3.
Характеристика зимней шерсти обыкновенной бурозубки
| Категория волос | n | Число волос на 4 мм2 | Длина волоса, мм | Толщина волоса, мкм | Число сегментов |
|---|---|---|---|---|---|
| Спина | |||||
| Направляющий | 16 | 4.1 ± 0.3 | 8.5 ± 0.09 | 30.2 ± 0.2 | 1 |
| Остевой I | 20 | 119.2 ± 3.1 | 7.5 ± 0.08 | 30.1 ± 0.6 | 6 |
| Остевой II | 20 | 135.6 ± 5.0 | 7.0 ± 0.08 | 25.4 ± 0.4 | 7 |
| Пуховой | 24 | 264.2 ± 5.1 | 6.2 ± 0.03 | 7.2 ± 0.3 | 7 |
| Бок | |||||
| Направляющий | 15 | 3.0 ± 0.1 | 8.5 ± 0.06 | 30.1 ± 0.3 | 1 |
| Остевой I | 16 | 117.0 ± 4.2 | 7.1 ± 0.05 | 30.0 ± 0.5 | 6 |
| Остевой II | 18 | 104.2 ± 3.6 | 6.9 ± 0.03 | 25.2 ± 0.3 | 7 |
| Пуховой | 21 | 205.4 ± 3.9 | 5.9 ± 0.02 | 7.2 ± 0.4 | 7 |
| Брюшко | |||||
| Направляющий | 17 | 2.3 ± 0.1 | 7.9 ± 0.09 | 31.0 ± 0.4 | 1 |
| Остевой I | 24 | 122.3 ± 3.3 | 6.1 ± 0.07 | 30.2 ± 0.1 | 6 |
| Остевой II | 26 | 141.7 ± 5.1 | 5.6 ± 0.05 | 25.1 ± 0.8 | 7 |
| Пуховой | 22 | 299.1 ± 5.0 | 4.6 ± 0.02 | 7.4 ± 0.6 | 7 |
Остевым волосам свойствен зональный поперечно-полосатый окрас – чередование светлых и темных полос, обусловленный своеобразной пигментацией полостей ороговевших клеток сердцевинного слоя, особенно хорошо развитого в расширенных участках стержня (Соколов, Чернова, 1998). Тем не менее характер окраса волос на разных участках шкурки неодинаков. На брюшке концевые гранны ости белые, а на спине и боках темно-коричневые (только самые кончики их бесцветны). Остальные сегменты остевых волос на всех частях тела окрашены в интенсивно черный цвет. При этом наибольшая концентрация меланина наблюдается в клетках сердцевины расширенных участков сегментов, а в перетяжках он распределяется диффузно, и они окрашены светлее. Крупные внутри- и межклеточные воздухоносные полости, преломляя свет, придают волосу характерный блеск и вместе с зональностью окраса и сочетанием темных и светлых сегментов определяют разнообразие оттенков меха на различных участках тела.
Кутикула остевых волос бурозубки, как и других насекомоядных, относится к некольцевидному типу, она образована одним слоем вытянутых ороговевших бесцветных чешуек со свободным округлым краем, обращенным к вершине волоса (рис. 2). У корня волоса и в перетяжках чешуйки сильно удлинены, контур их ровный, без зазубрин. Относительно неплотно прилегая друг к другу, они не мешают волосу сгибаться в любом направлении и вместе с тем обеспечивают ему достаточную прочность. В гранне же клетки кутикулы превращаются в очень короткие чешуйки, зазубренные по краям и плотно охватывающие стержень.
Корковый слой остевого волоса, как и направляющего, представлен веретеновидными вакуолизированными клетками, образующими стенки полости сердцевинного канала. При этом в местах поворота стержня толщина коркового слоя заметно увеличивается, благодаря чему волосы, несмотря на постоянное сгибание, не ломаются и демонстрируют значительную прочность на разрыв. Сердцевина остевых однорядна. Отсутствуя в кончиках и прикорневой части зрелых волос, она достигает максимального развития в середине апикального сегмента в виде строго чередующихся воздушных полостей и заполненных пигментом ороговевших клеток. В перетяжках распределение пигмента диффузное, концентрация его слабая, четкой клеточной структуры не наблюдается.
Пуховые волосы – самые тонкие и короткие. Апикальная ланцетовидная гранна развита у них слабее, чем у нижележащих сегментов. По количеству на единицу площади пух превосходит волосы всех остальных типов (на различных участках шкурки они составляют 45–60% волос всех других типов) и выполняют в основном теплозащитную функцию. Пуховые волосы имеют одинаковое число сегментов, как правило, на один больше, чем остевые (4 летом и 7 зимой), но отличаются высотой стержня, длиной и шириной апикальной гранны.
По форме чешуек кутикулы пуховые волосы не отличаются от остевых, но корковый слой у них, как и весь волос, более тонкий (около 2 мкм) и составляет около 20% толщины стержня. Сердцевина же однорядная, лестничная и состоит из рыхло соединенных перемежающихся воздухоносными полостями пигментированных клеток. В расширенных граннах (за исключением апикальной) сердцевинный слой развит сильнее и совершенно отсутствует в кончике и прикорневой части зрелого волоса. Напротив, растущие пуховые волосы отличаются хорошо развитой, богато пигментированной сердцевиной и в прикорневой части, особенно в начальный период роста.
Таким образом, у бурозубок, как и у других мелких представителей отряда насекомоядных, волосяной покров достаточно четко дифференцирован на 4 категории волос, имеющих разное строение и выполняющих разную функцию. Вместе с тем по сравнению с другими млекопитающими, например грызунами или хищниками, эта дифференциация выражена в наименьшей степени (отчего пуховой волос землероек некоторые авторы склонны относить к ости III порядка), что можно объяснить их более древним происхождением, а также приспособлением к норно-подземному образу жизни.
Плотность волосяного покрова. Шерсть бурозубки довольно густая (табл. 2 и 3), зимняя шерсть гуще летней, причем плотность волос изменяется по сезонам в основном за счет пуховых волос при сравнительно одинаковым количестве остевых и направляющих. Наблюдающееся на протяжении всей жизни зверька закономерное изменение густоты волос при постоянном числе волосяных фолликулов Боровский (Borowski, 1958) связывает с эффектом Денеля – изменением массы и общей поверхности тела животных по сезонам (проявление зимней депрессии). Плотность волос на различных участках шкурки неодинакова. Общее число их на брюшке больше, чем на спине и боках. Причем пуховых волос в 2–3 раза больше, чем остевых. Направляющие волосы встречаются по всей шкурке, но их очень немного, всего 2–3 волоска на каждые 470–500 волос других категорий.
Толщина волос. Волосы разных типов значительно различаются по толщине (табл. 2, 3). При этом пуховые волосы не только гораздо тоньше остевых и направляющих, но и наиболее выражены по этому признаку. Так, если ширина ланцетовидного апикального сегмента летнего пухового волоса варьирует от 4.5 до 5.0 мкм при коэффициенте вариации (CV) 3.9%, то у остевых II порядка – от 26.0 до 28.7 мкм и CV составляет 13.6%.
В результате осенней линьки волосы всех категорий становятся гуще, тоньше и почти в 2 раза длиннее. Ширина апикальной гранны ости I не превышает зимой 29–30 мкм, ости II порядка – 24.8–26, пухового волоса – 7.2–7.4 мкм. Летом соответствующие показатели – 49.7–50.0, 26.1–28.0 и 9.8–12.6 мкм. В то же время относительная толщина сердцевины от лета к зиме заметно увеличивается: от 50–63% в бесснежный период до 62–84% зимой (табл. 1).
Длина волос. Измерение длины волос проводилось в естественном их положении, без предварительного распрямления. В соответствии с полученными данными по высоте шерсти (рис. 1Б), на шкурке бурозубки можно выделить четыре зоны. Первая зона расположена на крестце, где волосы достигают максимальной длины и густоты (остевые зимнего меха составляют в среднем 8.3 мм). Вторая зона, с длиной волос 7.5 мм, занимает почти всю шкурку. Третья зона узкой полосой охватывает вторую (длина остевых 7.1 мм), а четвертая – всю вентральную сторону тела, где длина волос наименьшая (в среднем 6.1 мм). Виды, у которых, как и у обыкновенной бурозубки, наиболее длинный волос растет в области крестца, по данным Церевитинова (1958), должны относиться к группе с сакральным типом волосяного покрова, характерным в основном для наземных млекопитающих. Однако сравнительно небольшая разница в длине волос на различных участках шкурки (зонах) позволяет отнести изучаемый вид к формам с эквальным (равномерным) типом топографии волосяного покров, чаще всего встречающимся у типичных подземных животных. На наш взгляд, эти два типа топографии опушенности тела применительно к исследуемому виду вполне можно объединить. В этом случае характер распределения волос на шкурке бурозубки, а возможно и других мелких насекомоядных, ведущих норно-подземный образ жизни, следует относить к промежуточному сакро-эквальному типу.
Волосяной покров зверьков, отловленных летом, значительно ниже, чем у зимних. Пуховые волосы, входящие в состав летнего меха, – четырехсегментные, зимнего меха – семисегментные, остевые соответственно трех- и шестисегментные. Высота остевого волоса летом достигает в среднем 4.2 мм при длине эпикального сегмента 2.6 мм. Зимой – соответственно 7.5 и 2.8 мм. Таким образом, зимняя шерсть длиннее летней в 1.8–2 раза, тогда как длина концевого сегмента изменяется незначительно. Это относится и к другим категориям волос (табл. 2, 3). Причем сезонные изменения высоты волосяного покрова связаны не только с изменением числа сегментов, но и их длины. Удлинение к зиме характерно и для направляющих волос, у которых, как уже отмечалось, отсутствует сегментация. Летом длина направляющих на спине составляет в среднем 5.1 мм, а зимой – 8.5, т.е. в 1.6 раза больше, на боку: летом – 4.7 мм, зимой – 8.5, или в 1.8 раза длиннее.
Теплозащитные свойства покровов. О теплозащитных качествах шерсти зверьков мы судили по коэффициенту теплопроводности сухих коллекционных шкурок, измеренному, как уже говорилось выше, с помощью специального прибора ИТ-3 (табл. 4). Между тем известно, что теплозащитные свойства волосяного покрова млекопитающих зависят главным образом не от суммарной массы волос, а от слоя неподвижного (инертного) воздуха, находящегося между волосами. Количество же этого воздуха определяется их густотой, толщиной и длиной, а также структурой (“распушенностью”) волосяного покрова. Чем больше распушенность (несминаемость) щерсти, чем тоньше и длиннее волосы, тем больше воздушная прослойка между волосками и, следовательно, лучше сохраняется тепло. Нельзя забывать и о той роли, которую играет в этом процессе специальная кожная мускулатура, отвечающая за сохранение волос в поднятом состоянии, а значит и за теплозащитную функцию (Соколов, 1973).
Сохранение тепла зависит, правда в гораздо меньшей степени, и от инертного воздуха, заключенного в сердцевине волос (а его тем больше, чем сильнее развита сердцевина), а также в порах кожной ткани, а значит от ее толщины и плотности. Обусловленность теплозащитных свойств волосяного покрова обыкновенной бурозубки определенными морфологическими характеристиками и общим состоянием наружных покровов подтверждается нашими исследованиями теплопроводности сухих коллекционных шкурок, относящихся к трем периодам: лету, зиме и периоду осенней линьки (табл. 4).
При этом обнаруженные различия в коэффициентах теплопроводности шкурок мы объясняем указанными выше особенностями кожно-волосяного покрова. Чем длиннее, тоньше и гуще волосы и чем толще кожа, тем ниже коэффициент теплопроводности и, следовательно, выше теплоизоляционные свойства покровов. При этом, как показали результаты измерений, шкурки животных в период линьки удерживают тепло не только не хуже, но даже лучше шкурок животных, находящихся в состоянии покоя. Именно линные шкурки характеризуются наименьшими коэффициентами теплопроводности, на втором месте – шкурки зимних зверьков, на третьем – летних. Более высокие теплоизоляционные качества шкурок линяющих зверьков объясняются несколькими причинами. (1) Кожа у них приблизительно в 5 раз толще, чем в период покоя; (2) Плотность шерсти на линной шкурке возрастает за счет совмещения не выпавших еще старых волос с пробивающимися новыми; (3) К объему термоизолирующего воздуха, складывающегося из неподвижного воздуха, находящегося между волосками, и инертного – в сердцевине старых и новых волос, прибавляется инертный воздух сердцевины растущих волос, а также дополнительный воздух, скапливающийся в пазухах и просто в утолщающейся и расслаивающейся в период линьки кожной ткани. Таким образом, во время линьки механизмы физической терморегуляции не только не утрачивают своего значения, а, напротив, служат важным звеном в поддержании оптимального температурного гомеостаза зверьков и снижают непроизводительные траты энергии и потери тепла в сложные для животных переходные периоды.
Линька. Согласно данных многих авторов (Огнев, 1950; Павлова, 1951; Фадеев, 1955; Марвин, 1966, 1966а, 1969, 1974; Крыльцов, 1958, 1959, 1962, и др.), у млекопитающих первыми во время линьки развиваются обычно направляющие и остевые волосы. У бурозубки же очередность появления волос разных типов проследить затруднительно. На разных стадиях смены волос в их коже одновременно обнаруживаются крупные фолликулы будущих направляющих и остевых волос и более мелкие фолликулы – пуховых. Позднее, когда становятся заметны уже хорошо сформированные концевые сегменты волос всех категорий и они пробиваются наружу, мездра начинает постепенно светлеть и утончаться.
Как видно из приведенных в табл. 5 данных, первая линька прибылых бурозубок, покинувших гнезда, происходит только осенью – в сентябре–октябре. Первые сеголетки с темными пятнами на мездре, занимающими 20–90% площади шкурки (в среднем 46%), встречались начиная с десятых чисел сентября (11.09.1959, 12.09.1970, 14.09.1975, 11.09.2009), во второй декаде этого месяца линяло уже более 60% исследованных зверьков, а в третьей – 80%, причем интенсивность линьки (здесь и далее под термином “интенсивность” понимается относительная площадь шкурки, затронутая линькой, т.е. пигментированием) составляет в среднем 63%. В октябре линька прибылых продолжается. В первой декаде этого месяца линяет 85% исследованных обыкновенных бурозубок с интенсивностью от 8 до 100, в среднем 73%, а во второй – 50%, со средней интенсивностью 68% (3–100%). В третьей декаде октября линька прибылых заканчивается. Из 28 добытых в этот период зверьков линяло только 7 (25%), средняя интенсивность 18%. Последние линявшие молодые со шкурками, пигментированными на 3–5%, были пойманы 3 и 6.09.1969, 30.09.1975 и 1.09.2009. Волосы на шкурках прибылых зверьков, отловленных в начале ноября, были разными, т.е. на одной и той же шкурке можно было встретить и летние зрелые волосы с луковицами закрытого типа, и волосы на стадии роста (интенсивно пигментированная прикорневая часть стержня волоса, луковица открытого типа), и зрелые зимние волосы: шестисегментные остевые I, семисегментные остевые II и пуховые – все с закрытыми луковицами. Летние волосы постепенно выпадают, что подтверждается отсутствием их на шкурках зверьков из зимних и ранневесенних сборов, шерстный покров у таких землероек целиком состоял из зимних волос. Все это свидетельствует о полной смене волосяного покрова, происходящей в период осенней линьки.
Таблица 5.
Интенсивность линьки обыкновенных бурозубок в Карелии по месяцам
| Месяц | Самцы | Самки | ||||
|---|---|---|---|---|---|---|
| Число исследованных зверьков, экз. | из них линяющих, % | Относительная площадь пигментированной шкурки, % от площади всей шкурки | Число исследованных зверьков | из них линяющих, % | Относительная площадь пигментированной шкурки, % от площади всей шкурки | |
| Прибылые | ||||||
| VI | 15 | 8.8* | <1 | 14 | 21.4* | <1 |
| VII | 310 | 2.2* | <1 | 249 | 5.2* | <1 |
| VIII | 328 | 0.3* | <1 | 331 | 0.9* | <1 |
| IX | 105 | 74.3 | 56 | 62 | 71.0 | 56 |
| X | 31 | 71.0 | 67 | 52 | 65.4 | 46 |
| XI | 18 | 11.1* | <1 | 22 | 4.5* | <1 |
| XII | 17 | 24.6* | <1 | 13 | 38.5* | <1 |
| I | 14 | – | – | 13 | – | – |
| II | 16 | 12.5* | <1 | 1 | – | – |
| III | 14 | – | – | 14 | – | – |
| Зимовавшие | ||||||
| IV | 15 | 6.7 | 20 | 3 | 33.3 | 30 |
| V | 7 | 47.0 | 50 | 7 | 28.5 | 90 |
| VI | 29 | 24.1 | 17 | 14 | 14.2 | 3 |
| VII | 48 | 14.6 | 6 | 61 | 16.5 | 9 |
| VIII | 19 | 31.6 | 2 | 45 | 9.7 | 22 |
| IX | 4 | 50.0 | 28 | 11 | – | – |
| X | 3 | 66.6 | 100 | 6 | 50.0 | <1 |
Следует отметить и необычайную стабильность сроков осенней линьки молодняка, которые с удивительным постоянством повторяются из года в год. Отсутствуют и географические различия. В Англии, Нидерландах, Германии, Финляндии и различных областях России (в Сибири, Московской обл., в Татарии и т.д.) осенью линька сеголеток происходит в одни и те же сроки – в сентябре–октябре (Огнев, 1928; Формозов, 1948; Dehnel, 1949; Crowcroft, 1957; Строганов, 1957; Попов, 1960; Skaren, 1964, 1973; Michielsen, 1966; Borowski, 1968).
Летней “промежуточной” линьки молодых бурозубок, о которой по наблюдениям в Германии пишет Штейн (Stein, 1954), мы не обнаружили. Из 1247 сеголеток, пойманных в июне–августе, лишь у 28 (2.2% от числа просмотренных) имелась темная “точечная” пигментация на мездре (интенсивность линьки не более 1%). Других признаков линьки, в частности роста нового волоса, у них не было, так что считать их линяющими, очевидно, нет оснований.
Имеющийся в нашем распоряжении материал позволяет установить приблизительный порядок смены волос на различных участках шкурки во время осенней линьки сеголеток (рис. 3). Судя по этим данным, начинается она обычно на спине у огузка и распространяется вперед к голове, переходя постепенно на бока и брюшко. Казалось бы, и волосы соответственно должны заканчивать рост сначала на спине, а в последнюю очередь на брюшке, однако при тщательном исследовании ноябрьских шкурок зрелые зимние волосы были обнаружены одновременно и на верхней, и на нижней стороне тела (на линии середины туловища). Возможно, это объясняется тем, что волосы всех категорий, растущие на спине, обычно превосходят по длине волосы на брюшке. Поэтому волосы на спине, хотя и пробиваются раньше, окончательной зрелости достигают одновременно с волосами брюшка (более длинные волосы дольше растут). Описанный выше порядок осенней линьки не всегда бывает достаточно четким еще и потому, что картина затушевывается неодновременным вступлением зверьков в линьку, а также многочисленными индивидуальными отклонениями.
Весенняя линька у зимовавших зверьков обыкновенной бурозубки начинается в условиях Карелии во второй половине апреля и заканчивается в первой декаде мая (табл. 5), т.е. проходит в те же сроки, что и в других частях ареала (Огнев, 1928; Формозов, 1948; Dehnel, 1949; Stein, 1954; Строганов, 1957; Попов, 1960; Skaren, 1964, 1973). Лишь Кроукрофт (Crowcroft, 1957) и Боровский (Borowski, 1968) указывают для бурозубок Англии и Польши более ранние даты начала весенней линьки – первую декаду марта.
В мае землеройки Карелии линяют с наибольшей интенсивностью. 35% добытых в этом месяце бурозубок имели темную мездру на большей части шкурки (средняя интенсивность 50–90%) и подрастающий летний волос. Между тем в апреле линял лишь один из 18 осмотренных зверьков (5%), а остальные находились в зимнем мехе. Сроки начала линьки варьируют по годам в зависимости от хода весны и состояния популяции. В годы с ранней и дружной весной (1959, 1979, 1981–1982, 1970, 1974, 1986, 1989, 1992, 1998–1999, 2005 и 2009–2010 гг.) первые линяющие особи ловились уже в 20-х числах апреля, а при поздней холодной и затяжной весне (1961–1962, 1966, 1974, 1978, 1987–1988, 1990, 2002–2003, 2006 гг.) – не ранее начала–середины мая.
О топографии (порядке смены волос) весенней линьки можно судить по характерному расположению линных пятен на мездре и подросту новых летних волос на фоне зимних. Начинается процесс в паховой области, постепенно распространяется по животу к передней части тела, а затем идет на бока и внутреннюю сторону бедер, переходит на пояснично-крестцовую область и голову и заканчивается на спинной стороне (рис. 4). Это первый этап весенней линьки, приходящийся на конец апреля–первую декаду мая. Во второй половине мая наблюдается следующая фаза весенней линьки, для которой характерен противоположный порядок смены волос. Начинается она около крестца, идет вдоль хребта к голове, затем опускается на бока и брюшко и заканчивается в паховой области, а у самцов также вокруг боковых желез. Других различий в ходе линьки у самцов и самок не отмечено. Нет половых различий и в сроках линьки.
Рассмотренная выше картина двухэтапной весенней линьки перезимовавших зверьков по срокам и направлению соответствует первой и второй весенним линькам, описанным Боровским (Borowski, 1968, 1973). Различна лишь трактовка этого явления. По Боровскому, у каждого зимовавшего зверька бывают две следующие друг за другом, но разграниченные во времени, весенние линьки I и II, при которых смена волос идет в противоположных направлениях: сначала от живота к спине, а затем от спины к животу. Мы же считаем, что каждому зверьку свойственна лишь одна обычная весенняя линька, а не совпадение ее сроков и хода у отдельных особей объясняется их принадлежностью к различным возрастным генерациям. Первыми начинают линьку зверьки прошлогодних весенних пометов, т.е. более старшие по возрасту. Они и образуют мнимую первую весеннюю линьку. Что же касается второго этапа весенней линьки (по Боровскому, это вторая весенняя линька), то она соответствует массовой линьке особей поздних (летних) генераций. Иначе говоря, зимовавшие бурозубки вступают в весеннюю линьку в зависимости от своего календарного возраста как бы поэтапно. Это и создает ложное впечатление о существовании у одного зверька двух весенних линек. К отрицанию их реальности приходят и некоторые другие исследователи (например, Hyvärinen еt al., 1971).
Требует пояснения сравнительно небольшая доля линяющих особей в наших весенних сборах (за апрель–май – всего 21.8%). Причин здесь несколько. (1) Прежде всего, нельзя забывать о том, что сами учеты и сборы зверьков в весенний период нам удавалось проводить далеко не каждый год, так что материал за этот сезон (как и за зиму) в целом оказался немногочислен; (2) Это частично обусловлено большой индивидуальной скоростью линьки. Специальные наблюдения показали, что одна отдельно взятая землеройка может сменить весь свой волосяной покров за 2–5 дней (Wilcke, 1938; Borowski, 1964). А это вообще обусловливает достаточно низкую вероятность отлова линяющих особей; (3) Весенняя линька зверьков нередко перемежается паузами (по Боровскому, это так называемая “прерванная” линька). Мездра в этих случаях непигментированная, хотя смена волосяного покрова еще не закончилась, о чем говорит наличие зимних и летних волос на одной и той же шкурке. Такие животные тоже не попадают в категорию линяющих; (4) Относительно небольшая доля линяющих зверьков в весенних уловах может быть отнесена за счет их пониженной в этот период активности. Передвижения животных с утолщенной в результате линьки в 4–5 раз кожей, по-видимому, сравнительно более ограничены, поэтому вероятность их отлова уменьшается (Borowski, 1968).
В результате весенней линьк у бурозубок происходит замена зимних волос (шестисегментной ости I, семисегментной ости II и семисегментных пуховых) летними: трехсегментной остью I, четырехсегментной остью II и четырехсегментным пухом. Соответственно изменяется и длина волос (табл. 2, 3).
К концу мая весенняя линька у бурозубки заканчивается, и зверьки добываются уже без пигментации на мездре, с коротким летним мехом. Самая поздняя дата поимки самки с признаками начала весенней линьки 4.06.1967 г. Со второй декады июня по середину сентября мех бурозубки, по-видимому, находится в состоянии покоя, о чем свидетельствуют тонкая, светлая мездра, отсутствие линных пятен и прекращение роста волос всех категорий. На зрелость волос указывают луковицы: они закрытого типа.
Нормальной осенней линьки у зимовавших бурозубок не наблюдается, однако у них бывает так называемая старческая (сенильная – “промежуточная”, или “остаточная”) летне-осенняя линька, заключающаяся в росте нового короткого летнего волоса на некоторых частях тела, большей частью по его дорсальной стороне (Borowski, 1963). Длится эта линька с июня по октябрь, причем у одних особей она может проявиться сразу после завершения весенней линьки, а у других зверьков – после большого перерыва. В соответствии с этим намечается два пика интенсивности старческой линьки: в июне и в октябре, когда доля линяющих животных достигает 30–60%; в остальные месяцы она обычно не превышает 25% (табл. 5).
ЗАКЛЮЧЕНИЕ
Выполненное исследование посвящено обыкновенной бурозубке – одному из наиболее типичных представителей обширной экологической группы норно-наземных млекопитающих, или, как их еще называют, полунорников, мелких зверьков, обитающих в лесной подстилке и ходах неглубоких нор, но нередко появляющихся и на поверхности почвы. Этот вид землероек имеет общие, свойственные всем представителям группы мелких насекомоядных млекопитающих признак – волосяной покров образован прямыми направляющими, “шиловидными” отчетливо сегментированными остевыми двух размерных порядков и пуховыми (“зигзаговидными”). Стержень остевого волоса имеет чередующиеся расширения и продольные повороты (на 180°), наконец, вдоль стержня остевого волоса со всех четырех сторон проходят характерные именно для землероек и имеющие таксономическое значение продольные желобки разной глубины и конфигурации. Стержень состоит из трех слоев, волосы же имеют относительно небольшую длину и толщину, окраска зональная, своеобразна сезонная изменчивость. Кроме того, в строении ее наружных покровов выявлены специфические признаки, носящие явно адаптивный характер. По образу жизни, среде обитания, а значит и по строению волос, землеройковые, к которым относится и исследуемый вид, совмещают в себе многие черты, характерные как для настоящих наземных, так и для специализированных подземных млекопитающих, и по существу представляют собой переходный адаптивный тип. Обитая в умеренной и особенно холодной климатических зонах, в условиях постоянных низких температур, эти зверьки приобрели легкий, не мешающий движениям, но достаточно “теплый”, с высокими термоизоляционными свойствами волосяной покров, характеризующийся умеренной густотой и длиной волос, неравномерностью опушения отдельных частей тела, булавовидной удлиненной апикальной гранной, слабой извитостью основания волоса и несколько более сильным, чем у других групп, развитием сердцевинного канала.
Сами по себе волосы, как известно, не могут быть хорошим термоизолятором из-за относительно высокой теплопроводности их рогового вещества. Решающее значение в теплозащите, помимо “инертного” воздуха, заключенного в сердцевине волос, имеет так называемый “неподвижный” воздух, образующийся в шерстном покрове между волосами благодаря максимальному исключению конвекционных токов (Соколов, 1973). У землероек-бурозубок удержанию теплозащитного слоя воздуха способствует сама структура меха – одиночное, перпендикулярное к поверхности кожи расположение волос, их сегментация, извитость, дифференциация на категории. Разделяя пух и прикрывая его сверху, остевые и направляющие волосы не только предохраняют мех от сминания и свойлачивания, но и препятствуют выдуванию теплоизолирующего воздуха. Следовательно, обладая сравнительно слабо (в сравнении, например, с полевками) развитой сердцевиной волос, но зато гораздо большей их прочностью (за счет утолщения коркового слоя), землеройки компенсируют недостаточное количество “инертного” воздуха в сердцевине повышенным содержанием “неподвижного” воздуха в толще меха. В результате коэффициент теплопроводности их шкурок даже меньше, чем у грызунов.
Внутренняя микроструктура волос землероек также весьма своеобразна и связана со сложностью жизни в приземном слое воздуха. Сердцевина узкая или прерывистая в самых узких местах перетяжек стержня, что придает волосу особую прочность. Обширная полость стержня перегорожена равномерно распределенными относительно толстыми поперечными балками – дисками, также выполняющими важную адаптивную функцию (Соколов, Чернова, 1998).
Существенным дополнительным компонентом физической терморегуляции мелких норно-наземных млекопитающих служит и неравномерность опушения отдельных участков тела. Область спины, подвергающаяся у этих животных наибольшему охлаждению, покрыта самой высокой и густой шерстью, а на более скрытой и, следовательно, лучше защищенной от холода брюшной стороне тела волосяной покров значительно реже и ниже, чем на хребте и боках.
Наконец, у мелких насекомоядных млекопитающих волосяной покров выполняет также важную функцию защиты кожи от механических повреждений. Благодаря амортизационным свойствам он смягчает давление окружающего субстрата, которое постоянно приходится испытывать мелким зверькам при продвижении в узких ходах нор, лесной подстилке и переплетении трав. При этом функция механической защиты волосяного покрова вступает отчасти в противоречие с функцией термоизоляции (Соколов, 1973). Наибольшей прочностью обладают у землероек волосы со слабо развитой сердцевиной, однако с ее уменьшением ухудшаются их термоизоляционные свойства. Разрешается это противоречие несколькими путями. Один из них заключается в увеличении густоты таких тонкосердцевинных волос, что улучшает термозащитные качества шерсти без ущерба для его механических свойств. Второй путь – дифференциация волосяного покрова, при которой функцию механической защиты берут на себя направляющие и остевые волосы, а терморегуляционную – пуховые. Наконец, третий путь связан с различиями в строении волоса на разных участках его длины. При этом особое значение для защиты шерсти от вытирания приобретают у землероек бессердцевинные кончики волос, обладающие более высокими механическими свойствами благодаря хорошо развитому корковому слою. Большой прочностью на излом отличаются и другие зауженные участки волоса с отсутствием или слабым развитием сердцевины. В этих местах стержень, не ломаясь, легко сгибается в любую сторону, обеспечивая волосу землероек эластичность и предохраняя его от повреждений. У остевых волос полевок таких участков (их называют перетяжками) обычно два – у основания и перед гранной, а у землероек больше – от 3 до 9 в зависимости от вида зверька и сезона года.
Список литературы
Башкиров И.С., Жарков И.В., 1934. Биология и промысел крота // Ученые записки Казанского государственного университета. Т. 34. Вып. 3. С. 18–31.
Гудкова-Аксенова Н.С., 1951. Среда обитания и ее влияние на организацию некоторых насекомоядных и грызунов // Ученые записки Горьковского государственного университета. Вып. 19. С. 135–174.
Ивантер Э.В., Ивантер Т.В., Туманов И., 1985. Адаптивные особенности мелких млекопитающих: Эколого-морфологические и физиологические аспекты. Л.: Наука. 318 с.
Крыльцов А.И., 1958. Материалы по линьке мышевидных грызунов. Сообщение I. Линька массовых видов полевок северного Казахстана // Зоологический журнал. Т. 37. Вып. 2. С. 271–286.
Крыльцов А.И., 1959. Материалы по линьке мышевидных грызунов. Сообщение II. Линька различных представителей подсемейства полевок // Зоологический журнал. Т. 38. Вып. 5. С. 745–755.
Крыльцов А.И., 1962. Топография линьки грызунов и возможность использования ее в качестве таксономического признака // Труды Института защиты растений КазССР. Алма-Ата. Т. 7. С. 418–451.
Кузнецов Б.А., 1952. Основы товароведения пушно-мехового сырья. М.: Заготиздат. 508 с.
Марвин М.Я., 1966. Строение волосяного покрова полевок рода Microtus Среднего Урала // Ученые записки Уральского государственного университета. Сер. биол. Свердловск. Вып. 3. С. 52–57.
Марвин М.Я., 1966а. Влияние внешней среды на строение меха млекопитающих // Ученые записки. Серия биол. Свердловск. Вып. 3. С. 137–143.
Марвин М.Я., 1969. Строение меха Muridae // Труды Уральского отделения Московского общества испытателей природы. Свердловск. Вып. 3. С. 137–143.
Марвин М.Я., 1974. Строение волосяного покрова грызунов Урала // Научные труды Свердловского государственного педагогического института. Сборник 235. С. 11–22.
Огнев С.И., 1928. Звери Восточной Европы и Северной Азии. Т. 1. Насекомоядные и летучие мыши. М. 631 с.
Огнев С.И., 1950. Звери СССР и прилежащих стран. М. Т. VII. 706 с.
Павлова Е.А., 1951. Сезонные изменения волосяного покрова зайца-беляка и прогноз сроков выходности меха // Труды Всесоюзного научно-исследовательского института охотничьего хозяйства. Вып. 10. С. 59–77.
Попов В.А., 1960. Млекопитающие Волжско-Камского края. Казань: Госиздат. 466 с.
Соколов В.Е., 1973. Кожный покров млекопитающих. М.: Наука. 487 с.
Соколов В.Е., Скурат Л.Н., Степанова Л.В. и др., 1986. Руководство по изучению кожного покрова млекопитающих. М.: Наука. 279 с.
Соколов В.Е., Чернова О.Ф., 1998. Новые данные об архитектонике волос землероек и кротов (Insectivora: Soricidae, Talpidae) // Доклады Академии наук. Т. 360. № 5. С. 717–720.
Строганов С.У., 1957. Звери Сибири. Насекомоядные. М. 267 с.
Фадеев Е.В., 1955. Строение меха нутрии // Труды Всесоюзного научно-исследовательского института охотничьего хозяйства. М. Вып. 15. С. 93–104.
Формозов А.Н., 1948. Мелкие грызуны и насекомоядные Шарьинского района Костромской области в период 1930-1940 гг. // Материалы по грызунам. М. Вып. 3. С. 3–110.
Хмелевская Н.В., 1965. Строение кутикулы волос грызунов, ее изменчивость и значение для систематики // Зоологический журнал. Т. 44. Вып. 7. С. 1064–1074.
Церевитинов Б.Ф., 1958. Топографические особенности волосяного покрова пушных зверей // Труды ВНИИЖП. М. Вып. 17. С. 256–307.
Церевитинов Б.Ф., 1951. Дифференцировка волосяного покрова пушных зверей // Труды Всесоюзного научно-исследовательского института охотничьего хозяйства. М. Вып. 10. С. 6–17.
Чернова О.Ф., Целикова Т.Н., 2004. Атлас волос млекопитающих. М.: Товарищество научных изданий КМК. 429 с.
Borowski S., 1952. Sezonowe zmiany u wlosienia u Soricidae. Ann. Univ. M. Curie-Sklodowska. V. 7, 2.
Borowski S., 1958. Variation in density of coat during the life cycle of Sorex araneus araneus L. // Acta theriol. V. 2. № 14. P. 286–289.
Borowski S., 1963. Old-age moult in the common shrew, Sorex araneus Linnaeus, 1758 // Acta theriol. V. 7. № 19. P. 374–375.
Borowski S., 1964. Moult of shrews (Sorex L.) under laboratory conditions // Acta theriol. V. 8. № 8. P. 125–135.
Borowski S., 1968. On the moult in the common shrew // Acta theriol. V. 13. № 30. P. 483–498.
Borowski S., 1973. Variation in coat and colour in representatives of the genera Sorex L. and Neomys Kaup // Acta theriol. V. 18. № 14. P. 247–279.
Crowcroft P., 1957. The life of the shrew. London. 106 p.
Dehnel A., 1949. Studies in the genus Sorex L. // Ann. Univ. M. Curie-Sklod. Sect. C. V. 4. № 2. S. 17–102.
Hutterer R.H., Hürter T., 1981. Adaptive Haar Structuren bei Wasserspitzmausen (Insectivora, Soricidae) // Z. Säugetierk. Bd 46. S. 1–11.
Hyvärinen H., 1969. On the seasonal changes on the skeleton of the common shrew (Sorex araneus L.) and their physiological background // Aquilo. Ser. Zool. V. 7. P. 1–32.
Hyvärinen H., Pelttari A., Saure L., 1971. Seasonal changes in the histology and alkaline phosphatase distribution of the skin of the common shrew and the bank vole as a function of hair cycle // Aquilo. Ser. Zool. V. 12. P. 43–52.
Kapischke I.J., Muhle H.-J., 1988. Zur Morphologie der Waldspitzmaus (Sorex araneus) // Staatl. Mus. Tierk Dresden. Bd. 44. № 1. S. 71–74.
Keller A., 1978. Determination des mammiferes de la Suisse par leur pelage: Talpidae et Soricidae // Revue suisse Zool. № 85. Fasc. 4. P. 758–761.
Michielsen N.C., 1966. Intraspecific and interspecific competition in the shrews Sorex araneus L., and S. minutus L. // Arch. Neerland. Zool. № 17. № 1. P. 73–174.
Skaren U., 1964. Variation in two shrews, Sorex unguiculatus Dobson and S. a. araneus L. // Ann. Zool. Fenn. V. 1. № 2. P. 94–124.
Skaren U., 1979. Variation in breeding and moulting in Sorex isodon Turov in Finland // Ann. Zool. Fenn. № 159. P. 1–30.
Skaren U., 1973. Spring moult and onset of the breeding season of the common shrew (Sorex araneus L.) in Central Finland // Acta theriol. V. 18. № 23. P. 443–458.
Stein G.H.W., 1954. Materialen zum Haawechsel deutscher Insectivoren // Mitt. Zool. Mus. Berlin. Bd 30. S. 12–34.
Vogel P., Köpchen B., 1978. Besondere Haarstructuren der Soricidae (Mammalia, Insectivora) und ihre taxonomische Deutung // Zoomorphologia. Bd 89. S. 47–56.
Wilcke G., 1938. Freilands und gefangenschaftsbeobachtungen an Sorex araneus L. // Z. Säugetierk. Bd 12. S. 332–335.
Дополнительные материалы отсутствуют.
Инструменты
Зоологический журнал