

Зоологический журнал, 2021, T. 100, № 4, стр. 409-418
Фактор наследственности в выборе путей миграции и мест зимовок молодых особей орла-могильника (Aquila heliaca) из поволжской популяции
М. В. Корепов a, *, М. М. Ерохина a, **
a Ульяновский государственный педагогический университет имени И.Н. Ульянова,
Естественно-географический факультет, кафедра биологии и химии
432071 Ульяновск, Россия
* E-mail: korepov@list.ru
** E-mail: erokhina.marija@yandex.ru
Поступила в редакцию 22.04.2020
После доработки 24.08.2020
Принята к публикации 09.09.2020
Аннотация
Изучены данные о сроках и путях миграций, местах зимовок девяти молодых и одной взрослой особи орла-могильника (Aquila heliaca). Среди всех помеченных в 2017–2018 гг. спутниковыми передатчиками птиц шесть особей состояли друг с другом в родственных отношениях: два птенца из одного выводка, взрослый орел (самка) и его птенец, два птенца-погодка. Начало первой и второй осенней миграции, в целом, совпало для всех птиц и пришлось на конец сентября–начало октября. Вторая миграция молодых птиц, в сравнении с первой, оказалась менее протяженной за счет спрямления отдельных участков маршрута, а также в результате того, что места зимовки некоторых птиц изменились. Птенцы из одного выводка имели разные маршруты миграции, но зиму провели рядом, в центральной части Аравийского полуострова. Взрослая птица и ее птенец приступили к миграции в разные сроки и мигрировали порознь, но места зимовок обеих птиц совпали. Птенцы-погодки имели самые протяженные маршруты, и оба достигли южной оконечности Аравийского полуострова. При этом старший из птенцов преодолел Баб-эль-Мандебский пролив и перезимовал на Африканском континенте. Младший птенец, долетев до острова, разделяющего пролив, не решился пересечь открытое водное пространство. Совпадение мест зимовок, но использование разных путей миграции родственными птицами дает основание предполагать наследственный характер информации о местоположении зимовок. При этом маршрут миграции у молодых птиц вырабатывается в ходе их первой самостоятельной миграции и сохраняется в дальнейшем.
Изучение механизмов навигации и ориентации птиц, используемых при осуществлении сезонных миграций, – одна из ключевых тем современной орнитологии (Кишкинев, 2012; Чернецов, 2016). Особый интерес представляют первые миграции молодых птиц. Для объяснения этих миграций разработана гипотеза “компаса и часов”, которая выдвинута на основании экспериментов с воробьинообразными птицами и которая является наиболее распространенной в настоящее время (Gwinner, Wiltschko, 1978). Она предполагает существование врожденной пространственно-временной программы, отвечающей за время начала, направление и продолжительность миграции, отсутствие врожденной карты и контроля положения на трассе миграции у молодых птиц (Чернецов, 2016). Имеется достаточно убедительных экспериментов, подтверждающих, что отдельные параметры миграции (сроки, направление, длительность) передаются на генетическом уровне (Berthold et al., 1990; Helbig, 1991, 1996; Berthold et al., 1992; Berthold, Querner, 1981; Berthold, 1988; Delmore, Irwin, 2014). Для крупных парящих хищников основная роль в вопросе определения путей миграции молодых птиц из гнездового ареала к местам зимовки долгое время отводилась коммуникации со взрослыми особями (Дементьев, Гладков, 1951) по аналогии с коммуникацией у некоторых других крупных видов птиц, семейные группы у которых (например, у журавлей и белых аистов) сохраняются на протяжении первой осенней миграции и зимовки (Mueller et al., 2013; Chernetsov et al., 2004). Спутниковое мечение позволило полнее изучить особенности миграции крупных пернатых хищников, в частности была выявлена существенная роль наследственного фактора в определении сроков и путей миграций у гибридов большого и малого подорликов (Väli et al., 2018), гибридов орла-могильника и степного орла (Horváth et al., 2018). Однако навигационные способности молодых птиц и используемые ими ориентиры остаются не изучены (Fransson et al., 2001; Kullberg et al., 2003; Thorup et al., 2011; Кишкинев, 2012; Чернецов, 2016). В связи с этим точные данные, полученные современными методами телеметрии, о путях и сроках миграций, локализации районов зимовок молодых и взрослых птиц, находящихся в родственных отношениях, представляют особый интерес.
В ареале орла-могильника (Aquila heliaca) выделяют до 16 обособленных популяций, описанных на основе географической локализации гнездовых группировок, их биотопической приуроченности и динамики численности (Белик, Галушин, 1999). Западные популяции из Центральной Европы, Балкан, Анатолии и Кавказа считаются оседлыми или частично перелетными, восточные популяции из России и Казахстана – мигрирующими (Horváth et al., 2018). Наши исследования проведены на птицах из поволжской популяции, расположенной у северной границы гнездового ареала вида – в Среднем Поволжье. Орлы-могильники из данной популяции являются дальними мигрантами, основные районы зимовок которых сосредоточены на Ближнем Востоке. Для выяснения особенностей миграции родственных птиц были рассмотрены три модельных варианта: 1) два птенца из одного выводка (самец и самка), 2) взрослый орел (самка) и птенец (самец), 3) птенцы погодки (самцы) от одной пары. В данной работе основное внимание уделено первым осенним миграциям молодых орлов-могильников. Ранее уже было показано, что первая осенняя миграция каждой особи имеет индивидуальные особенности и играет ключевую роль в формировании миграционных маршрутов, которые в общих чертах сохраняются в дальнейшем, в том числе и в весенний период (Корепов и др., 2018).
МАТЕРИАЛ И МЕТОДИКА
Изучение путей миграции и мест зимовок орлов-могильников из поволжской популяции проведено в 2017–2018 гг. В этот период на территории Ульяновской обл. GPS/GSM-трекерами (фирма Aquila) помечено 10 орлов-могильников (по 5 особей в 2017 г. и 2018 г.), в том числе 1 взрослая самка и 9 молодых особей первого года жизни (3 самки и 6 самцов). Половую принадлежность определяли по размерам птиц. Трекеры крепили на спину по принципу “рюкзака” с помощью тефлоновых лент. Молодых особей (крупных птенцов и слетков) оснащали трекерами в конце июля–начале августа перед вылетом из гнезда. Взрослых орлов отлавливали на охотничьем участке неподалеку от гнезда с использованием приманки (молодые кролики и голуби). Вес передатчиков 35 г, элемент питания – солнечная батарея. Координаты положения птиц, оснащенных трекерами, определялись каждый час в светлое время суток и при нахождении птицы в зоне действия мобильной связи передавались на сервер несколько раз в сутки. За указанный период (1.08.2017–31.12.2018) от 10 помеченных орлов в общей сложности получено порядка 40 тыс. локаций (от 1330 до 7023 для отдельных птиц). Анализ локаций, треков и полигонов, а также метрические расчеты проведены на портале телеметрических данных производителя GPS/GSM-трекеров Aquilasystem и в программе Google Earth Pro.
В целях изучения особенностей миграции родственных птиц помечено 6 орлов на трех гнездовых участках, описанных ниже. Два птенца из одного выводка (самка Авралька и самец Филипп) помечены 28.07.2017 в гнезде среди агроландшафтов низменного левобережья Волги (Мелекесский р-н). Для них полностью выявлены пути первой (2017 г.) и второй (2018 г.) осенних миграций, первой весенней миграции (2018 г.) и районы первой зимовки (2017–2018 гг.). Взрослая самка Свияга и ее единственный птенец (самец Гуща) помечены 27 и 28.07.2018 в гнезде среди агроландшафтов в бассейне Свияги (Кузоватовский р-н). Для них полностью выявлены пути осенней миграции (2018 г.). Птенцы-погодки (единственные в выводке самцы) помечены 30.07.2017 (Шихан) и 1.08.2018 (Шиханчик) в одном гнезде, расположенном в лесостепных борах в междуречье Сызранки и Терешки на территории Богдановского заказника (Радищевский р-н). Для старшего птенца полностью выявлены пути первой осенней (2017 г.) и первой весенней (2018 г.) миграций, а также районы первой зимовки (2017–2018 гг.), для младшего птенца полностью прослежен путь первой осенней миграции (2018 г.).
Еще 4 птенца помечены на разных гнездовых участках: 27.07.2017 самец Тимоша в гнезде среди лесостепных ландшафтов в бассейне Малой Свияги (Кузоватовский р-н); 27.07.2017 самка Искорка в гнезде среди агроландшафтов в бассейне Малой Свияги (Майнский р-н); 2.08.2018 самка Терешка в гнезде среди лесостепных ландшафтов в бассейне Терешки (Старокулаткинский р-н); 6.08.2018 самец Сура в гнезде среди степных ландшафтов в долине Суры (Карсунский р-н). Для Тимоши и Искорки полностью выявлены пути первой (2017 г.) и второй (2018 г.) осенних миграций, первой весенней миграции (2018 г.) и районы первой зимовки (2017–2018 гг.). Для Терешки и Суры полностью прослежены пути первой осенней миграции (2018 г.).
За начало миграции принимали день направленного движения птиц из района гнездования/зимовки/летовки, за конец миграции принимали день пересечения ближайшего края области зимовки/летовки.
РЕЗУЛЬТАТЫ
Основные параметры осенних миграций орлов-могильников из поволжской популяции, полученные с помощью GPS/GSM-трекеров, приведены в табл. 1. Все молодые орлы начали свою первую осеннюю миграцию в третьей декаде сентября–первой декаде октября. Продолжительность первой осенней миграции варьировала от 23 до 53 дней, протяженность – от 3.7 тыс. до 6.9 тыс. км. Начало второй осенней миграции орлов оказалось более растянутым, но в целом укладывалось в тот же период, а вот продолжительность варьировала значительно выше – от 17 до 62 дней. В то же время протяженность осеннего миграционного пути на второй год у молодых орлов, сохранивших прежние районы зимовки (Филипп и Авралька), существенно уменьшилась (на 480 и 1360 км соответственно) за счет спрямления отдельных участков маршрута. Кроме того, у некоторых особей (Искорка и Тимоша) на второй год изменились и районы зимовок, расстояния до них сократилась соответственно на 780 и 2070 км по сравнению с первым годом. При этом новые районы зимовки сформировались на пути маршрута первой осенней миграции. Согласно результатам мечения орлов-могильников на Алтае, для молодых орлов также были характерны более дальняя первая осенняя миграция и более короткие последующие миграции с зимовкой в новом районе (Карякин, 2018).
Таблица 1.
Характеристика осенней миграции орлов-могильников из поволжской популяции
| Имя | Год | Начало | Конец | Продолжительность, сутки | Протяженность, км | Расстояние от гнезда до зимовки, км | Район зимовки |
|---|---|---|---|---|---|---|---|
| Тимоша | 2017 (juv) | 24 сентября | 6 ноября | 44 | 5708 | 4346 | Йемен |
| 2018 | 26 сентября | 13 ноября | 49 | 2887 | 2277 | Ирак | |
| Шихан* | 2017 (juv) | 24 сентября | 27 октября | 34 | 6851 | 4970 | Эфиопия |
| 2018 | – | – | – | – | – | – | |
| Искорка | 2017 (juv) | 29 сентября | 8 ноября | 41 | 5088 | 2257 | Саудовская Аравия |
| 2018 | 11 октября | 11 декабря | 62 | 3360 | 1477 | Азербайджан | |
| Филипп** | 2017 (juv) | 2 октября | 26 октября | 25 | 4180 | 3036 | Саудовская Аравия |
| 2018 | 20 сентября | 6 октября | 17 | 3702 | 3028 | Саудовская Аравия | |
| Авралька** | 2017 (juv) | 2 октября | 12 ноября | 42 | 5580 | 3099 | Саудовская Аравия |
| 2018 | 28сентября | 23 октября | 26 | 4221 | 3177 | Саудовская Аравия | |
| Гуща*** | 2018 (juv) | 5 октября | 12 ноября | 39 | 4148 | 2808 | Саудовская Аравия |
| Свияга*** | 2018 (ad) | 11 октября | 29 октября | 19 | 3681 | 3022 | Саудовская Аравия |
| Сура | 2018 (juv) | 5 октября | 9 ноября | 36 | 4274 | 2834 | Саудовская Аравия |
| Терешка | 2018 (juv) | 6 октября | 28 октября | 23 | 3698 | 2813 | Саудовская Аравия |
| Шиханчик* | 2018 (juv) | 28 сентября | 19 ноября | 53 | 5920 | 4174 | Йемен |
Взрослые орлы-могильники, насколько можно судить по одной помеченной птице, начинают осеннюю миграцию позже молодых птиц первого года жизни, но при этом преодолевают весь маршрут в более сжатые сроки, без длительных остановок и существенных отклонений от основного направления миграции. Так, взрослая орлица Свияга в 2018 г. приступила к осенней миграции на неделю позже своего птенца Гущи, но достигла мест зимовки, которые у Свияги и Гущи совпали, на две недели раньше (рис. 1). Исследования, проведенные раннее на больших подорликах (Aquila clanga), также показали, что все члены одной семьи (пара взрослых орлов и птенец) начали осеннюю миграцию в разные сроки, и птенец использовал иное направление миграционного пути, нежели взрослые орлы. Однако выяснить область зимовки птенца не удалось, вследствие его преждевременной гибели (Meyburg et al., 2005). Аналогичная картина предполагается для другого близкородственного вида – малого подорлика (Aquila pomarina). Две взрослые птицы (гнездящаяся пара) хоть и имели сходный маршрут миграции, приступали к ней по отдельности, в разные сроки (Meyburg, Meyburg, 2009).
Рис. 1.
Маршруты осенней миграции молодых орлов-могильников из одного выводка: Авральки в 2017 г. (а) и в 2018 г. (б), а также Филиппа в 2017 г. (в) и в 2018 г. (г).
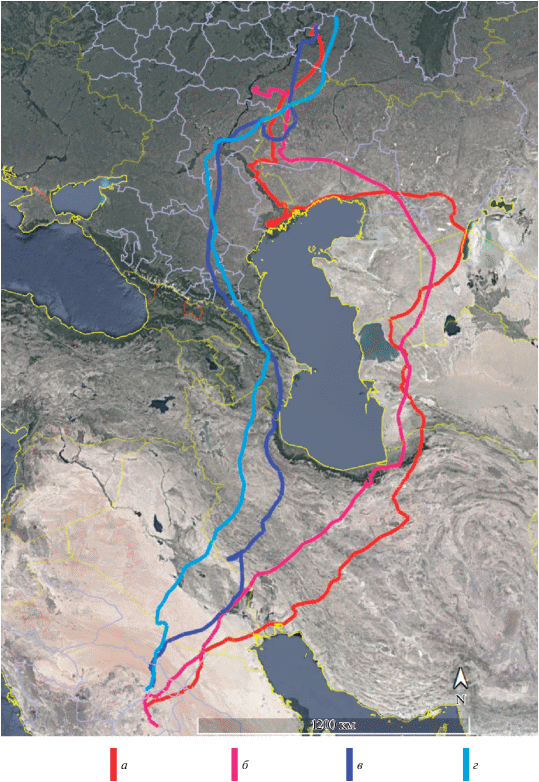
Сравнивая маршруты миграции помеченных орлов-могильников, можно выделить два основных миграционных направления: 1) к западу от Каспия – напрямую через Кавказ и 2) к востоку от Каспия – через плато Устюрт. Первое направление использует большинство орлов-могильников поволжской популяции (8 из 10 помеченных птиц). При этом выбор миграционного пути не определяется ни принадлежностью к той или иной гнездовой группировке, ни стереотипом гнездования, ни родственными связями птиц. В частности, в 2018 г. птенцы из одного выводка полетели разными путями (Филипп – через Кавказ, а Авралька – через Устюрт), однако районы их зимовки совпали (рис. 2).
Рис. 2.
Маршруты осенней миграции взрослой самки орла-могильника Свияги (а) и ее птенца Гущи (б) в 2018 г.
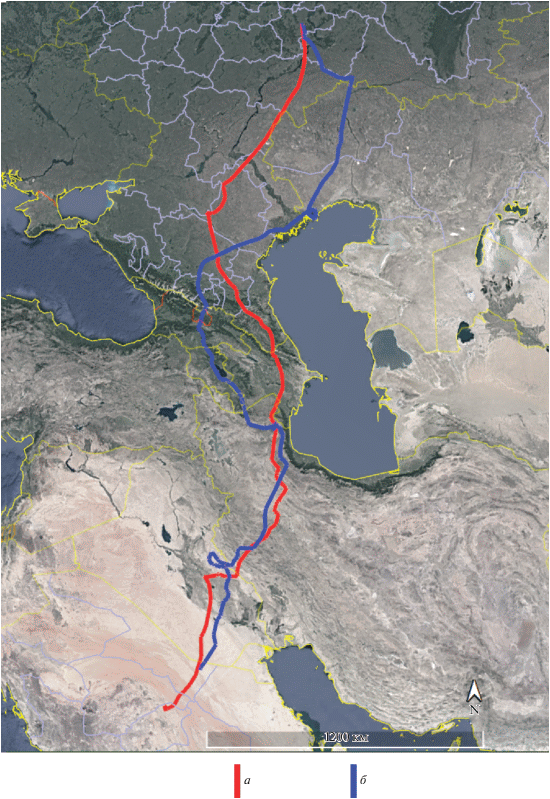
Несмотря на различия в путях миграции, область зимовок молодых орлов-могильников, совершающих первую осеннюю миграцию, в подавляющем большинстве (6 из 9 случаев) располагалась в центральной части Аравийского п-ова. Тольки три молодых орла (Тимоша, Шихан и Шиханчик) достигли южной оконечности Аравийского п-ова, а Шихан, перелетев через Баб-эль-Мандебский пролив между Красным и Аравийским морями, оказался на востоке Африки, где в Эфиопии и провел всю зиму и часть весны. Путь первой осенней миграции Шиханчика – брата-погодка Шихана – представляет особый интерес для выявления роли наследственности в выборе путей миграции и мест зимовок орлов, так как у Шихана наблюдались самый протяженный маршрут миграции и наиболее удаленная область зимовок. Отслеживание первых осенних миграций Шихана в 2017 г. и Шиханчика в 2018 г. показало, что молодые орлы выбрали различные пути перелета (Шихан – через Кавказ, Шиханчик – через Устюрт), однако обе птицы стремились попасть на Африканский континент (рис. 3). Но если Шихану удалось это сделать (23.10.2017 молодой орел со второй попытки преодолел Баб-эль-Мандебский пролив между Красным и Аравийским морями), то попытка Шиханчика 26.11.2018 преодолеть Баб-эль-Мандебский пролив не увенчалась успехом, и он остался зимовать в Йемене (рис. 4). В последнем случае решающую роль, вероятно, сыграло общее для всех парителей стремление избегать обширных морских акваторий.
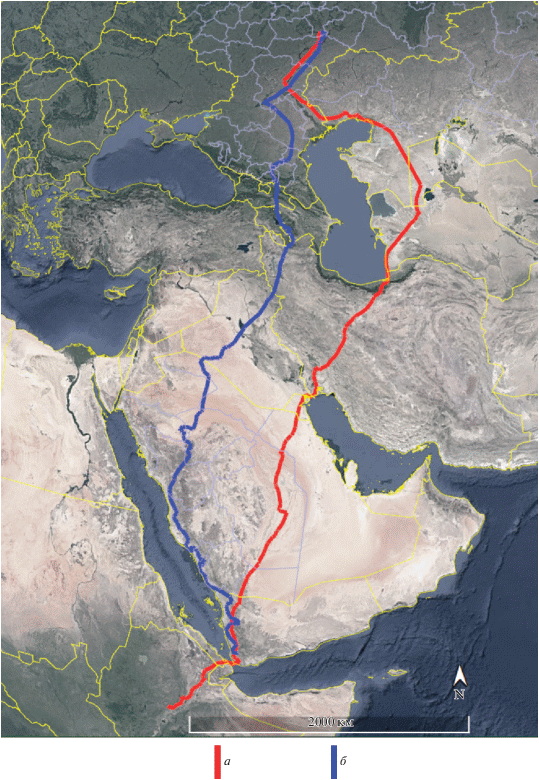
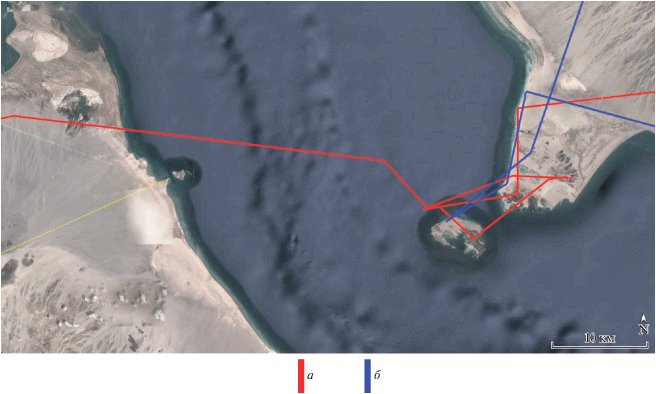
ОБСУЖДЕНИЕ
Проведенные наблюдения подтверждают, что свою первую миграцию молодые орлы совершают без сопровождения родителей и пользуются врожденными механизмами ориентации и навигации. Возможность использования неопытными мигрантами врожденного представления о магнитных и/или фотопериодических условиях, в которых они должны находиться при прохождении трассы осенней миграции, ранее уже предполагалась (Чернецов, 2016) на основе данных спутникового прослеживания перемещений молодых птиц, в частности чеглоков Элеоноры (Falco eleonorae) (Gschweng et al., 2008) и странствующих альбатросов (Diomedea exulans) (Åkesson, Weimerskirch, 2014).
Во всех трех экспериментах у родственных особей орлов-могильников районы зимовок совпали, при этом в двух из них маршруты первой осенней миграции у родственных птенцов отличались максимально возможным способом. Таким образом, можно предположить, что генетически (наследственно) детерминированными у орлов являются не маршруты миграции, а районы первой зимовки, к которой молодые орлы самостоятельно прокладывают свой индивидуальный маршрут миграции. Как правило, в дальнейшем первый осенний маршрут в общих чертах повторяется в ходе последующей весенней миграции и становится основным для сезонных перемещений конкретной особи. На второй год маршрут осенней миграции в целом остается прежним, но у отдельных особей значительно сокращается, в результате чего формируются новые места зимовок, расположенные ближе к районам летовки и гнездования. Подтверждением наследственной обусловленности первой осенней миграции орлов-могильников является и то, что в оседлых западных популяциях вида небольшая часть молодых птиц по-прежнему демонстрирует в первую зиму четкую миграцию на юг (Horváth et al., 2018).
Список литературы
Белик В.П., Галушин В.М., 1999. Популяционная структура ареала орла-могильника в Северной Евразии // Королевский орел. Распространение, состояние популяций и перспективы охраны орла-могильника. С. 129–139.
Деменьтев Г.П., Гладков Н.А., 1951. Птицы Советского Союза. Т. 1. М.: Советская наука. С. 275–279.
Карякин И.В., Николенко Э.Г., Шнайдер Е.П., Хорват M., Проммер М., Юхаш Т., Паженков А.С., Зиневич Л.С., 2018. Направление, характер и сроки миграции орлов-могильников из Волго-Уральского региона и Русского Алтая (Россия) по данным GSM/GPS и Argos/GPS-телеметрии // Пернатые хищники и их охрана. Спецвып. 1. С. 140–143.
Кишкинев Д.А., 2012. О последних достижениях в области изучения навигации у мигрирующих птиц // Бутурлинский сборник: Материалы IV Международных Бутурлинских чтений. Ульяновск: Корпорация технологий продвижений. С. 131–142.
Корепов М.В., Ковалев В.В., Ерохина М.М., Адамов С.Г., Корепова Д.А., и др., 2018. Миграции, зимовки и летние кочёвки молодых орлов-могильников из Поволжья, Россия // Пернатые хищники и их охрана. № 38. С. 83–92.
Чернецов Н.С., 2016. Ориентация и навигация мигрирующих птиц // Зоологический журнал. Т. 95. № 2. С. 128–146.
Åkesson S., Weimerskirch H., 2014. Evidence for sex-segregated ocean distributions of first-winter Wandering Albatrosses at Crozet Islands // PloS ONE. V. 9. № 2. P. e86779.
Berthold P., 1988. Evolutionary aspects of migratory behavior in European warblers // Journal of Evolutionary Biology. V. 1. № 3. P. 195–209.
Berthold P., Querner U., 1981. Genetic basis of migratory behavior in European warblers // Science. V. 212. № 4490. P. 77–79.
Berthold P., Helbig A.J., Mohr G., Querner U., 1992. Rapid microevolution of migratory behaviour in a wild bird species // Nature. V. 360. № 6405. P. 668–670.
Berthold P., Wiltschko W., Miltenberger H., Querner U., 1990. Genetic transmission of migratory behavior into a nonmigratory bird population // Experienti. V. 46. P. 107–108.
Chernetsov N., Berthold P., Querner U., 2004. Migratory orientation of first-year white storks (Ciconia ciconia): inherited information and social interactions // Journal of Experimental Biology. V. 207. № 6. P. 937–943.
Delmore K.E., Irwin D.E., 2014. Hybrid songbirds employ intermediate routes in a migratory divide // Ecology Letters. V. 17. № 10. P. 1211–1218.
Gschweng M., Kalko E.K.V., Querner U., Fiedler W., Berthold P., 2008. All across Africa: highly individual migration routes of Eleonora’s falcon // Proceedings of the Royal Society B. V. 275. № 1653. P. 2887–2896.
Gwinner E., Wiltschko W., 1978. Endogenously controlled changes in migratory direction of the garden warbler, Sylvia borin // Journal of Comparative Physiology A. V. 125. № 3. P. 267–273.
Fransson T., Jakobsson S., Johansson P., Kullberg C., Lind J., Vallin A., 2001. Magnetic cues trigger extensive refuelling // Nature. V. 414. № 6859. P. 35–36.
Helbig A.J., 1991. Inheritance of a migratory direction in a bird species: a cross-breeding experiment with SE- and SW-migrating blackcaps (Sylvia atricapilla) // Behavioral Ecology and Sociobiology. V. 28. № 1. P. 9–12.
Helbig A.J., 1996. Genetic basis, mode of inheritance and evolutionary changes of migratory directions in Palearctic warblers (Aves: Sylviidae) // Journal of Experimental Biology. V. 199. № 1. P. 49–55.
Horváth M., Özcan C., Juhász T., Kovács A., Tatar B., Karyakin I., Schmidt M., Tavares J., 2018. Breeding Population Surveys of Eastern Imperial Eagles and Steppe Eagles in Central Anatolia, Turkey // Journal on raptors of the East Europe and North Asia. Suppl. 1. P. 113–114.
Kramer G., 1953. Die Sonnenorientierung der Vögel // Verhandlungen der DeutschenZoologischen Gesellschaft, ZoologischerAnzeiger. Supplementband. 16. P. 72–84.
Kullberg C., Lind J., Fransson T., Jakobsson S., Vallin A., 2003. Magnetic cues and time of season affect fuel deposition in migratory thrush nightingales (Luscinia luscinia) // Proceedings of the Royal Society B. V. 270. № 1513. P. 373–378.
Mueller T., O’Hara R.B., Converse S.J., Urbanek R.P., Fagan W.F., 2013 Social learning of migratory performance // Science. V. 341. № 6149. P. 999–1002.
Meyburg B.-U., Meyburg C., 2009. Annual cycle, timing and speed of migration of a pair of Lesser Spotted Eagles (Aquila pomarina) – a study by means of satellite telemetry // Populationsökologie Greifvogel und Eulenarten. V. 6. P. 63–85.
Meyburg B.-U., Meyburg C., Mizera T., Maciorowski G., 2005. Family break up, departure, and autumn migration in Europe of a family of Greater Spotted Eagles (Aquila clanga) as reported by satellite telemetry // Journal of Raptor Research. V. 39. № 4. P. 462–466.
Thorup K., Ortvard T.E., Rabøl J., Holland R.A., Tøttrup A.P., Wikelski M., 2011. Juvenile songbirds compensate for displscement to oceanic islands during autumn migration. Public Library of Science One. V. 25. P. e17903
Väli Ü., Mirski P., Selis U., Dagys M., Maciorowski G., 2018. Genetic determination of migration strategies in large soaring birds: evidence from hybrid eagles // Proceedings of the Royal Society B. V. 285. № 1884. P. 20 180 855.
Дополнительные материалы отсутствуют.
Инструменты
Зоологический журнал