

Зоологический журнал, 2021, T. 100, № 4, стр. 434-448
Флуктуирующая асимметрия и морфогенетические корреляции рисунков жевательной поверхности m1 серых полевок (Rodentia, Arvicolinae)
В. Ю. Ковалева a, *, А. А. Поздняков a, Ю. Н. Литвинов a, В. М. Ефимов a, b, c, d
a Институт систематики и экологии животных СО РАН
630091 Новосибирск, Россия
b Национальный исследовательский Томский государственный университет
634050 Томск, Россия
c Институт цитологии и генетики СО РАН
630090 Новосибирск, Россия
d Новосибирский национальный исследовательский государственный университет
630090 Новосибирск, Россия
* E-mail: v_kov_65@mail.ru
Поступила в редакцию 28.02.2020
После доработки 26.04.2020
Принята к публикации 10.05.2020
Аннотация
Исследовали 5500 пар рисунков жевательной поверхности первого нижнего щечного зуба на левой и правой ветвях нижней челюсти особей, относящихся к одиннадцати видам серых полевок Microtus s. l. Выявлено 57 различных морфотипов m1 и 229 их различных сочетаний. Редкие морфотипы (встретившиеся суммарно с обеих сторон менее 5 раз) исключены из дальнейшего анализа. Оставшиеся морфотипы (34 различных, 199 сочетаний, всего 5466 пар) сгруппировали в таблицу сопряженности и применили к ней кластерный анализ. Кластерный анализ симметризованной таблицы сопряженности позволяет интерпретировать визуальную классификацию морфотипов с онтогенетических, точнее, морфогенетических позиций. Выявилось пять морфотипических кластеров, которые различаются по степени сложности морфотипов и соответствуют определенным видам серых полевок. Расстояния между морфотипами внутри кластеров существенно ниже, чем межкластерные расстояния, что свидетельствует о наличии модульности в системе морфотипической изменчивости исследованных видов. Модули сформированы по онтогенетическому сходству, поскольку получены из данных по флуктуирующей асимметрии морфотипов. Морфотип, как отдельная морфологическая структура, также имеет свою собственную модульную структуру, состоящую из двух подмодулей, соответствующих морфогенетическому механизму его усложнения.
Жевательная поверхность первого нижнего щечного зуба (m1) некорнезубых полевок представляет собой последовательность дентиноэмалевых полей, соединенных друг с другом вдоль продольной оси зуба, и ее рисунок характеризуется высокой изменчивостью. Похожие друг на друга рисунки (морфотипы) обычно, с неизбежной долей субъективности, выделяют визуально, и совокупность таких морфотипов образует ту или иную классификацию. В классификациях традиционно учитывают количество и тип замкнутых полей, а также число входящих и выступающих углов на внутренней и внешней сторонах зуба, так называемую-складчатость (Ангерманн, 1973; Большаков и др., 1980; Поздняков, 1995; Васильев, Васильева, 2009). Морфотипы различаются по частоте встречаемости у разных видов, поэтому они могут иметь определенное диагностическое значение (Ангерманн, 1973; Малеева, 1976; Большаков и др., 1980; Поздняков, 1995; Васильев, Васильева, 2009; Лисовский и др., 2018).
У полевок с правой и левой стороны челюсти в большинстве случаев формируются одинаковые морфотипы m1. Однако достаточно часто встречаются и разные морфотипы, т.е. асимметричные сочетания (Ангерманн, 1973; Еремина, 1974; Смирнов, Бененсон, 1980; Малеева, 1982; Ковалева и др., 2002; Поздняков, Сенотрусова, 2006; Поздняков, 2007, 2010, 2011; Васильев, Васильева, 2009; Kovaleva et al., 2013). Поскольку наблюдаемая асимметрия морфотипов m1 может складываться из разных по своей природе асимметрий (направленной, флуктуирующей и антисимметрии), их следует разделять и рассматривать отдельно (Захаров, 1987; Ковалева и др., 2002; Palmer, Strobek, 2003; Kovaleva et al., 2013).
Традиционно считается, что флуктуирующая асимметрия отражает в целом несбалансированность генных регуляций внутри организма, неспособность компенсировать средовые и генетические воздействия в ходе развития и напряженность отношений в системе “организм–среда” (Palmer, Strobeck, 1986; Захаров, 1987; Parsons, 1990, 1992; Dongen, 2006; Klingenberg, 2015). Для выделения флуктуирующей асимметрии можно воспользоваться методами статистического анализа таблиц сопряженности признаков (Кульбак, 1967). В этом случае частоты парных встреч правых и левых морфотипов, представленные в виде таблицы сопряженности, усредняются относительно диагонали таблицы. Симметризованное представление изменчивости морфотипов в таблице сопряженности отражает только флуктуирующую асимметрию. Поэтому кластерный анализ симметризованной таблицы сопряженности позволяет интерпретировать визуальную классификацию морфотипов с онтогенетических, точнее, морфогенетических позиций.
Морфологами давно замечено, что признаки, связанные онтогенетически и/или функционально, коррелируют более тесно, чем любые другие (Olson, Miller, 1958). В настоящее время такие наборы интегрированных признаков называются “модулями”. Ключевым свойством модульных систем является то, что они обладают сильными и разнообразными внутренними взаимодействиями и относительно автономны от других таких же модульных единиц, с которыми они связаны в меньшей степени. Поэтому они могут не только функционировать, но и эволюционировать относительно автономно (Cheverud, 1982; Wagner, Altenberg, 1996; Magwene, 2001; Winther, 2001). Возникновение новых модулей в эволюции может происходить путем дробления одной большой группы признаков на несколько новых, более мелких, при разрыве внутримодульных взаимодействий (Wagner, 1996; Wagner, Altenberg, 1996). Новые модули могут затем изменяться независимо друг от друга, тем самым увеличивается эволюционный потенциал системы. Периодический “развал” модулей препятствует процессам канализации развития и генетической плейотропии, усиление которых с течением времени могло бы привести к ограничению возможности систем к эволюционным изменениям (Wagner, Altenberg, 1996). С другой стороны, возникновение новых модулей в течение эволюционного времени возможно, благодаря созданию новых функциональных ассоциаций признаков (Wagner, 1996; Wagner, Altenberg, 1996). Таким образом, эволюционные эффекты модульной организации живых систем необходимо рассматривать в двух аспектах: в пределах одного модуля (интеграции) и между разными модулями (автономия) (Goswami, Polly, 2010).
В многочисленных работах сформулирован ряд актуальных вопросов, касающихся роли морфологической модульности в эволюции (Raff, Sly, 2000; Schlosser, 2002; Pigliucci, 2003; Колчанов, Суслов, 2006; Wagner et al., 2007; Klingenberg, 2008, 2014; Kuratani, 2009; Murren, 2012; Goswami et al., 2014; Rasskin-Gutman, Esteve-Altava, 2014; Ковалева и др., 2019). В их числе вопросы о широте распространения модульности среди живых организмов, методах идентификации модулей, биологических гипотезах возникновения модульности и интеграции и, наконец, о наиболее типичных примерах морфологических модулей в различных организмах и морфологических системах.
В настоящее время модульная организация выявлена в самых разных морфологических структурах. В частности, показана модульная организация нижней челюсти (Ehrich et al., 2003; Renaud et al., 2012), в составе которой молярный модуль можно рассматривать в качестве полуавтономного (Labonne et al., 2014). Следуя логике иерархической “вложенности” модулей разного уровня, в настоящей статье мы рассмотрели взаимоотношения процессов модульности и интеграции в системе морфотипической изменчивости m1 серых полевок Microtus s. l. на основании данных о флуктуирующей асимметрии.
МАТЕРИАЛ И МЕТОДЫ
Исследована морфотипическая изменчивость первого нижнего щечного зуба (m1) одиннадцати видов серых полевок11 “Microtus” s. l. (номенклатура дана по: Bannikova et al., 2010; Абрамсон, Лисовский, 2012) из: Сибирского зоологического музея ИСиЭЖ СО РАН, Новосибирск; Зоологического музея МГУ, Москва; Зоологического института РАН, Санкт-Петербург.
В работе использована авторская методика визуального описания изменчивости жевательной поверхности m1 (Поздняков, 1993, 2011). При выделении морфотипов использовали буквенно-цифровое кодирование. На первом этапе учитывали количество и форму замкнутых дентиновых полей. В качестве критерия отчленения одного поля от другого принимали смыкание пристеночного дентина противоположных сторон зуба в перешейке между полями. На этом этапе выделено шесть групп морфотипов, обозначенных буквами. Первые четыре буквы соответствуют количеству замкнутых полей треугольной формы: H – 3, K – 4, M – 5, T – 6. В двух оставшихся группах – P и Mа − последняя пара полей (считая от задней непарной петли (ЗНП)) не разделена. В группе P слиты четвертый и пятый треугольники, в группе Ma слиты шестой и седьмой треугольники (рис. 1).
Рис. 1.
Схема классификации морфотипов m1. В рамке показаны варианты строения передней непарной петли. ПНП – передняя непарная петля, ЗНП – задняя непарная петля, I–VI – замкнутые треугольники. Остальные пояснения в тексте.
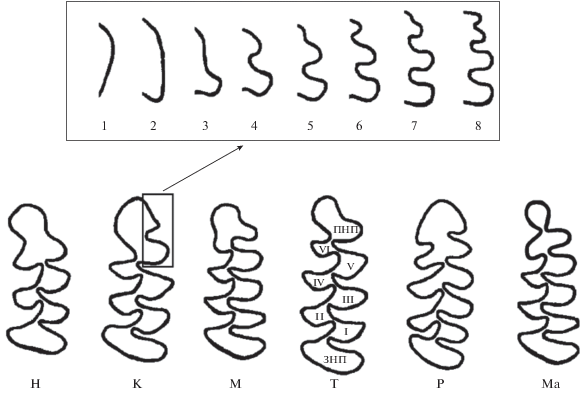
На втором этапе к буквенному обозначению морфотипа слева и справа приписывали числа, равные количеству изгибов лабиальной и лингвальной сторон передней непарной петли (ПНП), соответственно (рис. 1). Например, кодировка 3H6 обозначает морфотип с тремя замкнутыми треугольными полями, тремя изгибами лабиальной стороны ПНП и шестью – лингвальной.
Суммарную сложность морфотипов в ряду H–K–M–T рассчитывали, как сумму числа замкнутых полей (с коэффициентом 3) и числа изгибов лабиальной и лингвальной стороны ПНП. Коэффициент подобрали с учетом того, что при замыкании треугольника у основания ПНП число изгибов ПНП уменьшается на 2, при этом число замкнутых полей увеличивается на 1 (рис. 1). Следовательно, разница в сложности между морфотипами, различающимися на один замкнутый треугольник, при прочих равных обстоятельствах должна характеризоваться тремя единицами. Поэтому группам морфотипов серых полевок в качестве показателя сложности поставлены в соответствие следующие числа: Н – 15, К – 18, М – 21, Т – 24. Согласно этому алгоритму, суммарная сложность морфотипа 3H6 равна 24 (=3 + 5 × 3 + 6), а 3К6 – 27 (=3 + 6 × 3 + 6). Для групп морфотипов P и Mа, со слитыми четвертым и пятым, а также шестым и седьмым треугольниками, при разделении слитых полей образуются два дополнительных замкнутых треугольника и морфотипы группы P переходят в группу М, а морфотипы Ма – в теоретически возможную группу с показателем сложности 27, следующую за Т. Поскольку при этом количество замкнутых полей увеличивается на единицу, но не происходит никаких изменений в переднем отделе, то группе Р ставится в соответствие показатель 20 (=21-1). Аналогично, группе морфотипов Ma – 26 (=27-1).
Таким образом, в рамках данной визуальной классификации положение любого морфотипа в пространстве всех возможных морфотипов однозначно определяют три параметра: сложность лабиальной стороны, сложность замкнутых полей, сложность лингвальной стороны. Обозначим их C1ab, Ccf, Clin соответственно. Суммарная сложность морфотипа Сsum = C1ab + Ccf + Clin. В случае, когда рассматриваются пары морфотипов, встретившихся у одних и тех же особей, число признаков удваивается и они получают обозначения C1abL, CcfL, ClinL, СsumL и C1abR, CcfR, ClinR, СsumR, соответственно, для левой и правой стороны челюсти.
Особи старших возрастных групп по строению моляров существенно не различаются между собой (Voyta et al., 2013). Моляры ювенильных особей характеризуются тонкой эмалью, плохо выраженным пристеночным дентином, почти не выраженным обособлением полей. Поэтому их не включали в анализ.
Исследовали 5500 пар рисунков жевательной поверхности первого нижнего щечного зуба на левой и правой ветвях нижней челюсти особей, относящихся к одиннадцати видам серых полевок Microtus s. l. Выявлено 57 различных морфотипов m1 и 229 их различных сочетаний. Морфотипы, встретившиеся суммарно с обеих сторон менее 5 раз (3H5, 3H6, 4H4, 6H4, 1K7, 3K4, 4K7, 4K8, 2P3, 3P3, 4P4, 2M2, 3M6, 4M6, 1T3, 1T5, 3T3, 3T5, 1Ma1, 1Ma2, 1Ma3, 2Ma3, 3Ma3), исключены из дальнейшего анализа вместе со своей парой. После этого осталось 5466 пар рисунков (34 различных морфотипа, 199 сочетаний, из них 33 симметричных и 166 – асимметричных) (рис. 2). Тем не менее, симметричные пары вариантов встречаются значительно чаще, чем асимметричные. Общая по всем видам доля симметричных особей составила 72.1%, асимметричных, соответственно, 27.9%.

Асимметрия морфотипов и асимметрия сложности морфотипов − не одно и то же. Асимметрия морфотипов наблюдается, когда слева и справа есть различия хотя бы по одному показателю сложности. Под асимметрией сложности в рамках данной статьи мы рассматриваем только асимметрию суммарной сложности. Одной и той же суммарной сложности могут соответствовать несколько разных морфотипов, в том числе из разных групп. В частности, в нашем случае 34 анализируемым морфотипам соответствуют девять градаций суммарной сложности (23, 24, …, 31) (табл. 1).
Таблица 1.
Частота встречаемости симметричных пар морфотипов и их параметры
| Виды (n) | 4 H 5 |
5 H 4 |
5 H 5 |
5 H 6 |
6 H 5 |
1 K 4 |
1 K 5 |
1 K 6 |
2 K 4 |
2 K 5 |
2 K 6 |
2 K 7 |
3 K 5 |
3 K 6 |
4 K 5 |
4 K 6 |
3 P 4 |
1 M 3 |
2 M 3 |
3 M 3 |
4 M 3 |
2 M 5 |
1 M 4 |
2 M 4 |
3 M 4 |
1 T 4 |
3 K 7 |
3 M 5 |
4 M 4 |
4 M 5 |
5 M 4 |
5 M 5 |
2 T 4 |
2 Ma 2 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| M. agrestis (226) | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 2 | 0 | 0 | 21 | 0 | 0 | 0 | 0 | 89 | 0 | 0 | 40 | 20 | 5 | 0 | 4 | 0 | 0 |
| A. fortis (489) | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 144 | 22 | 0 | 0 | 0 | 123 | 64 | 0 | 0 | 3 | 2 | 0 | 0 | 0 | 0 | 0 |
| L. gregalis (412) | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 4 | 7 | 68 | 0 | 0 | 2 | 14 | 213 | 0 | 0 | 6 | 2 | 0 | 0 | 0 | 0 | 1 |
| B. juldaschi (120) | 2 | 3 | 86 | 2 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| A. maximowiczii (623) | 0 | 0 | 1 | 0 | 0 | 0 | 3 | 0 | 0 | 8 | 1 | 0 | 45 | 8 | 4 | 5 | 0 | 0 | 20 | 166 | 3 | 0 | 0 | 4 | 100 | 0 | 1 | 10 | 18 | 2 | 0 | 0 | 0 | 0 |
| A. middendorffii (319) | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 2 | 65 | 2 | 0 | 0 | 0 | 113 | 0 | 0 | 16 | 38 | 1 | 0 | 0 | 0 | 0 |
| A. mongolicus (215) | 0 | 0 | 0 | 0 | 0 | 0 | 2 | 0 | 0 | 6 | 3 | 0 | 6 | 9 | 0 | 0 | 0 | 0 | 4 | 15 | 0 | 0 | 0 | 14 | 67 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 |
| A. mujanensis (150) | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 3 | 0 | 0 | 2 | 0 | 0 | 0 | 13 | 4 | 0 | 0 | 0 | 33 | 1 | 0 | 1 | 34 | 3 | 0 | 0 | 2 | 0 |
| A. oeconomus (2568) | 1 | 0 | 0 | 0 | 0 | 24 | 828 | 56 | 20 | 819 | 51 | 1 | 16 | 4 | 0 | 0 | 0 | 0 | 30 | 2 | 0 | 0 | 0 | 3 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| M. paradoxus (119) | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 19 | 0 | 0 | 0 | 64 | 1 | 1 | 0 | 0 | 1 |
| M. rossiaemeridionalis (225) | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 2 | 0 | 0 | 0 | 0 | 70 | 0 | 0 | 2 | 108 | 4 | 0 | 1 | 0 | 1 |
| All | 3 | 3 | 87 | 2 | 1 | 24 | 833 | 56 | 20 | 834 | 55 | 1 | 70 | 22 | 4 | 7 | 3 | 4 | 207 | 374 | 9 | 0 | 2 | 158 | 768 | 1 | 1 | 79 | 286 | 16 | 1 | 5 | 2 | 3 |
| C1ab | 4 | 5 | 5 | 5 | 6 | 1 | 1 | 1 | 2 | 2 | 2 | 2 | 3 | 3 | 4 | 4 | 3 | 1 | 2 | 3 | 4 | 2 | 1 | 2 | 3 | 1 | 3 | 3 | 4 | 4 | 5 | 5 | 2 | 2 |
| Ccf | 15 | 15 | 15 | 15 | 15 | 18 | 18 | 18 | 18 | 18 | 18 | 18 | 18 | 18 | 18 | 18 | 20 | 21 | 21 | 21 | 21 | 21 | 21 | 21 | 21 | 24 | 18 | 21 | 21 | 21 | 21 | 21 | 24 | 26 |
| Clin | 5 | 4 | 5 | 6 | 5 | 4 | 5 | 6 | 4 | 5 | 6 | 7 | 5 | 6 | 5 | 6 | 4 | 3 | 3 | 3 | 3 | 5 | 4 | 4 | 4 | 4 | 7 | 5 | 4 | 5 | 4 | 5 | 4 | 2 |
| Сsum | 24 | 24 | 25 | 26 | 26 | 23 | 24 | 25 | 24 | 25 | 26 | 27 | 26 | 27 | 27 | 28 | 27 | 25 | 26 | 27 | 28 | 28 | 26 | 27 | 28 | 29 | 28 | 29 | 29 | 30 | 30 | 31 | 30 | 30 |
| Кластер | I | I | I | I | I | II | II | II | II | II | II | II | III | III | III | III | III | III | III | III | III | IV | IV | IV | IV | IV | V | V | V | V | V | V | V | V |
Для каждого вида абсолютная и относительная частоты (число и доля) совместной встречаемости вариантов морфотипов m1 с правой и левой стороны челюсти у одних и тех же особей представлены нами в виде квадратных таблиц сопряженности22 (Кульбак, 1967; Agresti, 2007), в которой ключами строк являются идентификаторы левых морфотипов, ключами столбцов – идентификаторы правых морфотипов. В каждой видовой таблице сопряженности сумма относительных частот равна единице.
Объемы видовых выборок сильно различаются между собой. Так, например, для A. oeconomus n = = 2568, для M. paradoxus n = 119, для B. juldaschi n = 120 (табл. 1). Если рассчитать таблицу сопряженности по всей выборке, то почти наполовину она будет состоять из таблицы сопряженности A. oeconomus. Чтобы скорректировать разницу в объемах видовых выборок, были вычислены таблицы сопряженности относительных частот по всем 11 видам полевок и средняя всех таблиц взята в качестве объединенной таблицы сопряженности. Таким образом, все видовые таблицы сопряженности вошли в объединенную таблицу с равным весом.
Относительная частота каждой пары морфотипов wij в клетке объединенной таблицы сопряженности была принята в качестве веса этой пары. Суммарный вес всех пар морфотипов, очевидно, равен 1. Между всеми восемью показателям визуальной сложности морфотипов была рассчитана матрица коэффициентов взвешенной корреляции (табл. 2) по формуле
Таблица 2.
Коэффициенты взвешенной корреляции параметров визуальной сложности морфотипов с левой и правой стороны нижней челюсти
| All Sp | ClabL | CcfL | ClinL | CsumL | ClabR | CcfR | ClinR | CsumR |
|---|---|---|---|---|---|---|---|---|
| ClabL | 1.000 | –0.290* | 0.104 | 0.320* | 0.887*10 | –0.283 | 0.095 | 0.244 |
| CcfL | –0.290* | 1.000 | –0.648*10 | 0.741*10 | –0.279 | 0.956*10 | –0.594*10 | 0.726*10 |
| ClinL | 0.104 | –0.648*10 | 1.000 | –0.250 | 0.109 | –0.585*9 | 0.784*10 | –0.275 |
| CsumL | 0.320* | 0.741*10 | –0.250 | 1.000 | 0.263 | 0.720*10 | –0.293* | 0.918*10 |
| ClabR | 0.887*10 | –0.279 | 0.109 | 0.263 | 1.000 | –0.276 | 0.095 | 0.324* |
| CcfR | –0.283 | 0.956*10 | –0.585*9 | 0.720*10 | –0.276 | 1.000 | –0.663*10 | 0.749*10 |
| ClinR | 0.095 | –0.594*10 | 0.784*10 | –0.293* | 0.095 | –0.663*10 | 1.000 | –0.276 |
| CsumR | 0.244 | 0.726*10 | –0.275 | 0.918*10 | 0.324* | 0.749*10 | –0.276 | 1.000 |
$\bar {x},\bar {y}$ – взвешенные средние показателей x и y.
Проблема уровня значимости была решена следующим образом. Минимальный объем видовой выборки, n = 119 – у M. paradoxus. Считаем его нижней оценкой для всей матрицы. Учитываем общее число коэффициентов корреляции в таблице – 28 – и делаем поправку Бонферрони, т.е., принимаем для уровня значимости p табличное значение коэффициента корреляции, рассчитанное при pB = p/28. Например, при pB = 0.05 в качестве табличного значения брали 0.286, а не 0.180, при pB = 1E-10 – 0.587, а не 0.549 и т.д. Для всех коэффициентов корреляции, превышающих по модулю 0.587, принимали pB < 1E-10. Понятно, что при таком упрощенном подходе значимость полученных коэффициентов корреляции сильно занижена, но для целей статьи этого вполне достаточно. Для сокращения записи одна звездочка в показателе степени (*) означает pB < 0.05; (*m) – pB < 1E-m, где m = 2, 3, …, 10.
Каждая пара различных морфотипов может появиться на левой и правой стороне челюсти в любом порядке, например, 2K4–2K5 и 2K5–2K4. Такой паре морфотипов соответствуют две клетки таблицы сопряженности, симметричные относительно диагонали таблицы. Частоты встречаемости морфотипов в этих клетках могут не совпадать друг с другом, характеризуя направленную асимметрию. Различия между сторонами в случае направленной асимметрии считаются генетически детерминированными (Захаров, 1987; Palmer, 1996). Направленную асимметрию в таблицах сопряженности обычно устраняют усреднением частот, симметричных относительно диагонали таблицы (Кульбак, 1967), по формуле
Флуктуирующую асимметрию характеризует суммарная доля асимметричных пар, которая не меняется после устранения направленной асимметрии. После устранения направленной асимметрии и получения симметричной матрицы каждая строка объединенной таблицы сопряженности нормировалась на сумму ее значений по формуле
Сумма значений по строкам таблицы сопряженности – это частота встречаемости каждого морфотипа. После нормировки на сумму различия между морфотипами характеризуются только их относительными частотами встречаемости в парах со всеми морфотипами, независимо от собственной частоты. Для каждого морфотипа сумма таких частот равна 1.
Между наборами полученных частот вычислена матрица евклидовых расстояний Кавалли-Сфорца–Эдвардса (Cavalli-Sforza, Edwards, 1967; Ковалева и др., 2002) по формуле
Далее к этой матрице расстояний применен кластерный анализ (Ward, 1963) (рис. 3). Для пяти полученных кластеров вычислена матрица внутри- и межкластерных средних квадратов расстояний (Миркин, 1980) (табл. 3).
Рис. 3.
Кластеризация морфотипов m1 11 видов серых полевок Microtus s. l., построенная методом Уорда (Ward, 1963) по объединенной матрице относительных частот совместной встречаемости морфотипов с правой и левой стороны нижней челюсти. I–V – классы морфотипов.

Таблица 3.
Матрица внутри- и межкластерных средних квадратов евклидовых расстояний между морфотипами в % от максимально возможных значений
| Кластер | I | II | III | IV | V |
|---|---|---|---|---|---|
| I | 34 | 97 | 99 | 100 | 100 |
| II | 97 | 52 | 84 | 88 | 97 |
| III | 99 | 84 | 64 | 78 | 85 |
| IV | 100 | 88 | 78 | 52 | 80 |
| V | 100 | 97 | 85 | 80 | 69 |
Таким образом, для данного набора морфотипов получено две классификации: параметрическая, в виде сочетания трех параметров сложности каждого морфотипа (C1ab, Ccf, Clin), и кластерная – на основе матрицы евклидовых расстояний между морфотипами. Параметры и их значения определяются систематиком на основании визуального сходства морфотипов. Визуальное восприятие достаточно субъективно. Однако при любой визуальной классификации появление любого морфотипа, а также их совместное появление у одной и той же особи объективно и определяется программой онтогенеза (морфогенеза) особи. Соответственно, чем чаще встречаются в паре разные морфотипы, тем ближе их морфогенетические программы. Таблица сопряженности морфотипов опирается не на их визуальное сходство (например, параметров сложности), а отражает только частоту совместной встречаемости морфотипов, иными словами, близость их морфогенезов. По таблице сопряженности рассчитывается матрица евклидовых расстояний между морфотипами, поэтому классификация, получаемая в результате кластерного анализа, по своей природе является морфогенетической.
Визуальная (параметрическая) и кластерная (морфогенетическая) классификации абсолютно не обязаны соответствовать друг другу. Однако, если морфотипы в полученных кластерах близки и по визуальным параметрам, это фактически означает независимое объективное подтверждение субъективной классификации систематика.
Расчеты проводились с помощью пакетов PAST (Hammer et al., 2001) и Jacobi4 (Polunin et al., 2019; jacobi4.ru).
РЕЗУЛЬТАТЫ
В большинстве исследованных нами образцов морфотипы с правой и левой стороны челюсти были симметричными (табл. 2). Однако у каждого вида от 17 до 41% особей имели различные морфотипы на разных сторонах челюсти. Кластерный анализ таблицы сопряженности, отражающей флуктуирующую асимметрию морфотипов, показал наличие пяти кластеров (рис. 3). Первый кластер характеризует изменчивость морфотипов памирской полевки (Blanfordimys juldaschi (Severtzov 1879)), в него вошли самые простые морфотипы с тремя замкнутыми треугольниками (4H5, 5H4, 5H5, 5H6, 6H5) (табл. 1, рис. 3). Второй кластер отражает изменчивость морфотипов полевки-экономки (Alexandromys oeconomus (Pallas 1776)), в него вошли морфотипы с четырьмя замкнутыми треугольниками и сложностью лабиальной стороны ПНП не выше двух (1K4, 1K5, 1K6, 2K4, 2K5, 2K6, 2K7) (табл. 1, рис. 3). Третий кластер характеризует изменчивость морфотипов A. fortis и A. maximowiczii. В него вошли морфотипы с четырьмя замкнутыми треугольниками и сложностью изгиба лабиальной стороны ПНП выше двух (3K5, 3K6, 4K5, 4K6), кроме 3K7, морфотипы с пятью замкнутыми треугольниками и сложностью изгиба лингвальной стороны ПНП, равной трем (1M3, 2M3, 3M3, 4M3), а также 3P4 (табл. 1, рис. 3). Четвертый кластер отражает изменчивость морфотипов ряда видов – узкочерепной полевки (Lasiopodomys gregalis (Pallas 1779)), темной полевки (Microtus agrestis (Linnaeus 1761)), A. middendorffii и A. mongolicus. В него вошли морфотипы, главным образом, с пятью замкнутыми треугольниками, сложностью изгиба лингвальной стороны ПНП выше трех и сложностью изгиба лабиальной и лингвальной сторон ПНП в сумме не выше семи (1M4, 2M4, 2M5, 3M4, 1T4) (табл. 1, рис. 3). Пятый кластер характеризует изменчивость морфотипов муйской полевки (A. mujanensis) копетдагской полевки (M. paradoxus Ognev et Heptner 1928) и восточноевропейской полевки (M. rossiaemeridionalis Ognev 1924). В него вошли морфотипы, главным образом, с пятью замкнутыми треугольниками и сложностью изгиба лабиальной и лингвальной сторон ПНП в сумме выше семи (3M5, 4M4, 4M5, 5M4, 5M5), а также 3K7, 2T4, 2Ma2 (табл. 1, рис. 3).
В табл. 2 приведена матрица коэффициентов взвешенной корреляции восьми билатеральных показателей визуальной сложности морфотипов. Обращает на себя внимание, что 4 из 6 самых больших корреляций концентрируются по боковым диагоналям матрицы, отражая сильные статистические связи между парными билатеральными показателями сложности (C1abL – C1abR; CcfL – CcfR; ClinL – ClinR; СsumL – СsumR). Остальные корреляции отражают положительную зависимость между сложностью замкнутых полей и суммарной сложностью морфотипа как для одной стороны (только левой или только правой CcfL – СsumL; CcfR – СsumR), так и для противоположных сторон (CcfL – СsumR; CcfR – СsumL). Кроме того, сложность замкнутых полей имеет отрицательную корреляцию со сложностью лингвальной стороны моляра (CcfL – ClinL; CcfR – ClinR; CcfL – ClinR; CcfR – ClinL), также для одной и противоположных сторон.
ОБСУЖДЕНИЕ
Идея использования флуктуирующей асимметрии для исследования закономерностей морфогенеза билатеральных морфологических структур широко известна (Астауров, 1927; Van Valen, 1962; Palmer, Strobeck, 1986, 2003; Захаров, 1987, 2001; Parsons, 1990, 1992; Graham et al., 2003; Kellner, Alford, 2003; Klingenberg, 2003; Васильев, 2005; Вершинин и др., 2007; Гилева и др., 2007; Graham et al., 2010; Ерофеева, 2014; Рахмангулов и др., 2014; Ялковская и др., 2014, 2016; Гордеева, 2016; Козлов, 2017). Щечные зубы полевок, особенно m1 и M3, ввиду широкого спектра изменчивости рисунка их жевательной поверхности предоставляют хорошую возможность для исследований в этом направлении. Известно, что разные виды полевок различаются не столько наборами вариантов жевательной поверхности моляров, сколько распределениями их частот и спецификой доминирующих морфотипов (Малеева, 1975, 1976; Васильев, 2009). Флуктуирующая асимметрия морфотипов при достаточно большом фактическом материале выявляет весь спектр их допустимых онтогенетических сочетаний.
Представление флуктуирующей асимметрии морфотипов m1 в виде таблиц сопряженности, использованное в настоящей работе, позволяет количественно оценить онтогенетические (морфогенетические) расстояния между морфотипами. Кластерный анализ матрицы евклидовых расстояний между морфотипами выявил наличие пяти кластеров, удовлетворяющих требованию фенотипической модульности (в данном случае это близкие расстояния между морфотипами внутри модуля и более далекие – между модулями) (табл. 3). Учитывая то, что исходной матрицей послужила усредненная по видам таблица сопряженности, полученная на основании флуктуирующей асимметрии (частот билатеральных встреч разных морфотипов), выделенные модули – онтогенетического происхождения. Кластеризация морфотипов произошла в порядке возрастания их сложности, каждому кластеру соответствуют определенный вид или виды полевок. Иными словами, в качестве главной тенденции эволюции жевательной поверхности m1 серых полевок следует рассматривать ее усложнение.
Первый кластер морфотипов представляет памирская полевка (Blanfordimys juldaschi). Для этого вида характерны относительно простые морфотипы, практически отсутствующие в соседних кластерах (табл. 2). Расстояния между морфотипами внутри этого кластера существенно ниже, чем между соседними кластерами (табл. 3), что свидетельствует о наличии сильных внутримодульных связей. Палеонтологические данные свидетельствуют в пользу древности этого вида (Rabeder, 1981; Chaline et al., 1999).
На втором месте по силе внутримодульных связей находятся кластеры второй и четвертый (табл. 3). Второму кластеру морфотипов соответствует A. oeconomus. Полевка-экономка появляется как уже вполне оформившийся вид в конце раннего плейстоцена с набором морфотипов, практически не изменившимся до настоящего времени (Рековец, 1994; Громов, Ербаева, 1995). Однако этот набор сильно отличается от морфотипов остальных видов современного рода Alexandromys, присутствие в соседних кластерах (I и III) незначительно (табл. 2). Четвертый кластер морфотипов соответствует видам: M. agrestis, L. gregalis, A. middendorffii, A. mongolicus. Эти виды характеризуются более широким спектром морфотипической изменчивости. Много общих морфотипов с соседними кластерами (III и V).
Наименьшую силу внутримодульных связей имеют третий и пятый кластеры (табл. 3). Наборы морфотипов третьего кластера соответствуют видам A. fortis и A. maximowiczii. Много общих морфотипов с соседним кластером (IV) (табл. 2). Ископаемые остатки A. fortis известны из позднеплейстоценовых отложений Забайкалья и раннеплейстоценовых отложений Северного Китая (Громов, Ербаева, 1995). Ископаемые остатки, предположительно принадлежащие A. maximowiczii (идентифицируемые как A. cf. maximowiczii) известны из среднеплейстоценовых отложений Забайкалья (Агаджанян, Ербаева, 1983). Пятый кластер морфотипов соответствует видам A. mujanensis, M. paradoxus, M. rossiaemeridionalis. Наблюдаемый спектр изменчивости говорит в пользу того, что эти виды по сравнению с другими видами характеризуются менее стабильным развитием моляров.
Палеонтологические данные свидетельствуют о том, что в раннем плейстоцене у представителей рода Allophaiomys резко увеличился спектр морфотипической изменчивости (Агаджанян, 2009), что свидетельствует пользу дестабилизации их фенотипа. С позиции эпигенетической теории эволюции (Шишкин, 2006, 2012) при расселении в новых условиях закрепляется один из вариантов модификационного спектра (морфоз), что в нашем случае выражается в изменении частот морфотипов и формировании модальных кластеров (Поздняков, 1996, 2007).
Разные морфологические структуры имеют свои собственные модели модульности и интеграции. Несмотря на то, что нижняя челюсть функционально, онтогенетически и топологически тесно связана с черепом, она часто изучается как отдельная морфологическая структура, так как она имеет свою собственную модель модульности независимо от остальной части черепа. В соответствии с онтогенетическими и генетическими критериями нижняя челюсть состоит из двух модулей: переднего альвеолярного и заднего мыщелкового регионов (Klingenberg et al., 2003). Это деление является также функциональным, поскольку альвеолярный модуль несет молярную зону, а мыщелковая ветвь, соединяясь с черепом, служит местом для прикрепления жевательных мышц. Тем не менее, в соответствии с функциональными и эволюционными критериями нижнюю челюсть можно дополнительно разделить на пять модулей: в альвеолярной области это резцовый и молярный модули, в мыщелковой – модули венечного (processus coronoideus), сочленовного (pr. condylaris) и углового отростков (pr. angularis) (Ehrich et al., 2003; Renaud et al., 2012). Кроме того, молярный модуль можно рассматривать в качестве полуавтономного, который далее можно разделить на модули моляров и модуль зубного ряда (Labonne et al., 2014).
В рамках другого, мерономического подхода считается, что морфотипы щечных зубов полевок являются композиционными структурами, состоящими из независимых компонентов – меронов (=модулей) (Васильев, Васильева, 2009). Для описания всего морфотипического разнообразия используется три мерона: складчатости жевательной поверхности внутренней (лингвальной) и наружной (лабиальной) сторон зуба и характер слияния дентиновых пространств. В этом случае по степени скоррелированности признаков можно судить, является ли морфотип случайным сочетанием вариаций признаков или жестко скоррелированной структурой и представляет собой “морфу”. В ходе исследования выяснилось, что между признаками есть определенные слабые корреляции, что означает невыполнение условия полностью случайного комбинирования признаков складчатости и слияния дентиновых пространств.
В нашем исследовании при описании изменчивости m1 мы применили, в общем, близкие показатели, отличия от мерономического подхода заключаются в том, что под сложностью лингвальной и лабиальной сторон зуба понимали сложность лингвальной и лабиальной сторон передней непарной петли, кроме того, дополнительно использовали один интегративный показатель – “суммарная сложность морфотипа”.
Анализ корреляций восьми билатеральных показателей выявил относительную независимость сложности лабиальной стороны зуба (C1abL, C1abR) от всех остальных для левого и правого моляра. Значимые корреляции проявились только между сторонами (C1abL – C1abR, r = 0.887*10) (табл. 2). С точки зрения блочно-модульной организации фенотипа сильные статистические связи между признаками свидетельствуют о принадлежности их к одному модулю. В данном случае параметры сложности лабиальной стороны левого и правого моляра образуют билатеральный модуль.
Сложность замкнутых полей отрицательно коррелирует со сложностью лингвальной стороны (CcfR – ClinR, r = –0.662*10; CcfL – ClinL, r = = ‒0.647*10) и положительно – с суммарной сложностью морфотипа (CcfR – СsumR, r = 0.749*10; CcfL – СsumL, r = 0.740*10). Скорее всего, эта корреляция объясняется тем, что у исследованных видов полевок, за исключением памирской полевки, с наибольшей частотой представлены морфотипы с четырьмя и пятью замкнутыми треугольниками (K и M). Соответственно, трансформация морфотипов K в M происходит путем замыкания лингвального зубца, лежащего в основании ПНП. В этом случае количество замкнутых полей увеличивается на единицу и уменьшается количество изгибов лингвальной стороны ПНП на два. Очевидно, что взаимосвязь этих признаков имеет онтогенетическую природу. Кроме того, так же, как и в случае со сложностью лабиальной стороны, сложность замкнутых полей и сложность лингвальной стороны зуба демонстрируют высокие билатеральные корреляции (CcfR – CcfL, r = 0.955*10; ClinR – ClinL, r = 0.784*10).
Следует отметить, что среди выявленных на сегодняшний день модулей млекопитающих (Esteve-Altava, 2017), билатеральных не так много. Среди них можно отметить модули дерматоглифических признаков, образованных гомологичными структурами правой и левой рук (пальцы или ладони), например, нганасан Таймырского полуострова (Galaktionov et al., 1982) или жителей Сардинии (Floris, 2015).
Результаты исследования показали, что на основании анализа средних внутрикластерных и межкластерных квадратов расстояний в спектре морфотипов m1 выделяются пять модулей, различающихся по степени сложности входящих в них морфотипов. Морфотипы, в свою очередь, также являются интегрированными структурами, состоящими из двух модулей: модуля, включающего число замкнутых дентиновых полей плюс сложность лингвальной стороны, и модуля лабиальной стороны моляра. Таким образом, морфотипическая изменчивость m1 одиннадцати видов серых полевок имеет иерархическую модульную организацию.
ЗАКЛЮЧЕНИЕ
В настоящее время билатеральная асимметрия используется в качестве удобной природной модели для изучения эпигенетических механизмов регуляции морфогенеза (Васильев, 2005; Васильев, Васильева, 2009; Kharlamova et al., 2010; Epigenetics …, 2011). В ряде исследований проанализированы онтогенетические траектории морфологических структур среди родственных таксонов (Mitteroecker et al., 2005; Bulygina et al., 2006; Kimmel et al., 2012). Эти исследования проливают свет на процессы развития, на основе которых возникают морфологические различия между таксонами, и демонстрируют связь между онтогенетическими и эволюционными интеграциями.
Кластерный анализ матрицы евклидовых расстояний между морфотипами m1 серых полевок выявил наличие пяти кластеров. Поскольку матрица евклидовых расстояний рассчитана по таблице сопряженности, отражающей флуктуирующую асимметрию морфотипов, следовательно, выявленные кластеры по своей природе являются онтогенетическими (морфогенетическими). Одновременно кластеры морфотипов отвечают также и критерию “визуального сходства”. Связи между морфотипами внутри кластеров сильнее, по сравнению со связями морфотипов между кластерами. Таким образом, система морфотипической изменчивости m1 изученных видов полевок имеет модульную структуру. Кластеризация морфотипов отражает возрастание их сложности. Кластеры морфотипов соотносятся с определенными видами серых полевок, причем некоторые кластеры соотносятся с несколькими видами, что указывает на наличие интеграции в системе.
Коэффициенты взвешенной корреляции показателей визуальной сложности отражают модульную структуру моляра. Наиболее сильными являются билатеральные модули параметров визуальной сложности левого и правого моляра, а также модуль, отражающий морфогенетическую связь между сложностью замкнутых полей, сложностью лингвальной стороны и суммарной сложностью морфотипа. Сложность лабиальной стороны зуба продемонстрировала статистическую независимость.
В целом, полученная картина соответствует модели общих и местных факторов развития (Mitteroecker, Bookstein, 2007). Местные факторы способствуют морфологической изменчивости в пределах только одного модуля, в то время как общие факторы влияют на признаки разных модулей. В модели специально подчеркивается одновременное наличие модульности и интеграции развития: некоторые характеристики или особенности формы независимы между разными модулями, в то время как другие – являются общими для нескольких модулей, что указывает на их интеграцию.
Список литературы
Абрамсон Н.И., Лисовский А.А., 2012. Подсемейство / Subfamily Arvicolinae Gray, 1821 // Сборник трудов Зоологического музея МГУ. Т. 52. Млекопитающие России: систематико-географический справочник. М.: Товарищество научных изданий КМК. С. 220–276.
Агаджанян А.К., 2009. Мелкие млекопитающие плиоцен-плейстоцена Русской равнины // Труды Палеонтологического института РАН. Т. 289. М.: Наука. 676 с.
Агаджанян А.К., Ербаева М.А., 1983. Позднекайнозойские грызуны и зайцеобразные территории СССР. М.: Наука. 190 с.
Ангерманн Р., 1973. Гомологическая изменчивость коренных зубов у полевок (Microtinae) // Проблемы эволюции. Т. 3. Новосибирск: Наука. С. 104–118.
Астауров Б.Л., 1927. Исследование наследственного изменения галтеров у Drosophila melanogaster // Журнал экспериментальной биологии. Вып. 3. № 1/2. С. 1–61.
Баскевич М.И., Окулова Н.М., Потапов С.Г., Миронова Т.А., Сапельников С.Ф., Сапельникова И.И., Егоров С.В., Власов А.А., 2012. Новые данные о распространении видов-двойников и гибридизации 46-хромосомных форм Microtus arvalis sensu lato (Rodentia, Arvicolinae) в Центральном Черноземье // Зоологический журнал. Т. 91. № 8. С. 994–1005.
Большаков В.Н., Васильева И.А., Малеева А.Г., 1980. Морфотипическая изменчивость зубов полевок. М.: Наука. 140 с.
Васильев А.Г., 2005. Эпигенетические основы фенетики: на пути к популяционной мерономии. Екатеринбург: Академкнига. 640 с.
Васильев А.Г., 2009. Феногенетическая изменчивость и популяционная мерономия // Журнал общей биологии. Т. 70. № 3. С. 195–209.
Васильев А.Г., Васильева И.А., 2009. Гомологическая изменчивость морфологических структур и эпигенетическая дивергенция таксонов: основы популяционной мерономии. М.: Товарищество научных изданий КМК. 512 с.
Вершинин В.Л., Гилева Э.А., Глотов Н.В., 2007. Флуктуирующая асимметрия мерных признаков у остромордой лягушки: методические аспекты // Экология. № 1. С. 75–77.
Гилева Э.А., Ялковская Л.Э., Бородин А.В., Зыков С.В., Кшнясев И.А., 2007. Флуктуирующая асимметрия краниометрических признаков у грызунов (Mammalia: Rodentia): межвидовые и межпопуляционные сравнения // Журнал общей биологии. Т. 68. № 3. С. 221–230.
Гордеева И.В., 2016. Коэффициент флуктуирующей асимметрии листовой пластинки как показатель общего экологического стресса // Успехи современной науки. Т. 9. № 12. С. 105–109.
Громов И.М., Ербаева М.А., 1995. Млекопитающие фауны России и сопредельных территорий. Зайцеобразные и грызуны. СПб.: Зоологический институт РАН. 522 с.
Еремина И.В., 1974. Полиморфизм рисунка жевательной поверхности коренных зубов у обыкновенной полевки // Физиологическая и популяционная экология животных. Вып. 2 (4). Саратов: Изд-во Саратовского ун-та. С. 77–91.
Ерофеева Е.А., 2014. Влияние свинца на флуктуирующую асимметрию листа гороха посевного (Pisum sativum L.) // Вестник ННГУ. № 1. С. 162–165.
Захаров В.М., 1987. Асимметрия животных: популяционно-феногенетический подход. М.: Наука. 216 с.
Захаров В.М., 2001. Онтогенез и популяция (стабильность развития и популяционная изменчивость) // Экология. №. 3. С. 164–168.
Ковалева В.Ю., Поздняков А.А., Ефимов В.М., 2002. Изучение структуры изменчивости морфотипов коренных зубов полевки-экономки (Microtus oeconomus) через билатеральную асимметрию их проявления // Зоологический журнал. Т. 81. № 1. С. 111–117.
Ковалева В.Ю., Поздняков А.А., Литвинов Ю.Н., Ефимов В.М., 2019. Оценка сопряженности морфогенетических и молекулярно-генетических модулей изменчивости серых полевок Microtus s. l. в градиентных условиях среды // Экологическая генетика. Т. 17. № 2. С. 21–34.
Козлов М.В., 2017. Исследования флуктуирующей асимметрии растений в России: мифология и методология // Экология. № 1. С. 3–12.
Колчанов Н.А., Суслов В.В., 2006. Кодирование и эволюция сложности биологической организации // Эволюция биосферы и биоразнообразия. М.: Товарищество научных изданий КМК. С. 60–96.
Кульбак С., 1967. Теория информации и статистика. М.: Наука. 408 с.
Лисовский А.А., Кадетова А.А., Оболенская Е.В., 2018. Морфологическое определение видов восточноазиатских серых полевок Alexandromys (Rodentia, Cricetidae) России и сопредельных территорий // Зоологический журнал. Т. 97. № 1. С. 101–113.
Малеева А.Г., 1975. Сравнение морфотипической изменчивости зубов Arvicola terrestris L. и Lagurus lagurus Pall. // Фауна Урала и Европейского Севера. Сб. 4. Свердловск. С. 42–49.
Малеева А.Г., 1976. Об изменчивости зубов у полевок (Microtinae) // Эволюция грызунов и история формирования их современной фауны. Л.: ЗИН АН СССР. С. 48–57.
Малеева А.Г., 1982. Симметрия и асимметрия строения жевательной поверхности коренных зубов у полевок на примере степной пеструшки (Lagurus lagurus Pall.) // Морфология и систематика млекопитающих. Л.: ЗИН АН СССР. С. 23–37.
Малыгин В.М., 1983. Систематика обыкновенных полевок. М.: Наука. 207 с.
Миркин Б. Г., 1980. Анализ качественных признаков и структур. М.: Статистика. 319 с.
Моролдоев И.В., Картавцева И.В., 2017. Новые данные об инвазии восточноевропейской полевки (Microtus rossiaemeridionalis) на восток от г. Улан-Удэ // Вестник БГУ, биология, география. Вып. 3. С. 130–134.
Моролдоев И.В., Шереметьева И.Н., Картавцева И.В., 2017. Первая находка восточноевропейской полевки (Microtus rossiaemeridionalis) в Бурятии // Российский журнал биологических инвазий. Т. 10. № 2. С. 88–94.
Окулова Н.М., Сапельников С.Ф., Баскевич М.И., Власова О.П., Майорова А.Д., Егоров С.В., Миронова Т.А., Сарычев В.П., 2008. Сравнительная экология трех форм обыкновенных полевок Microtus arvalis sensu lato в Центральном Черноземье // Научные ведомости Белгородского гос. ун-та. Серия “Естественные науки”. Вып. 3. № 6. С. 128–139.
Поздняков А.А., 1993. Морфотипическая изменчивость жевательной поверхности коренных зубов серых полевок группы “maximowiczi” (Rodentia, Arvicolidae): опыт количественного статистического анализа // Зоологический журнал. Т. 72. № 11. С. 114–125.
Поздняков А.А., 1995. Таксономическая интерпретация морфологической изменчивости на примере серых полевок (Microtus s. lato, Rodentia) // Журнал общей биологии. Т. 56. № 2. С. 172–178.
Поздняков А.А., 1996. Проблемы филогении полевок подрода Alexandromys Ognev (Rodentia, Arvicolidae, Microtus). Изменчивость и палеонтологические данные // Зоологический журнал. Т. 75. № 1. С. 133–140.
Поздняков А.А., 2007. Структура морфотипической изменчивости серых полевок (Microtus: Rodentia, Arvicolidae) с точки зрения эпигенетической теории эволюции // Успехи современной биологии. Т. 127. № 4. С. 416–424.
Поздняков А.А., 2010. Морфологическое разнообразие: характеристика, структура, анализ // Сообщества и популяции животных: экологический и морфологический анализ / Труды института систематики и экологии животных СО РАН. Вып. 46. Новосибирск, М.: Товарищество научных изданий КМК. С. 133–157.
Поздняков А.А., 2011. Структура морфологической изменчивости (на примере морфотипов жевательной поверхности первого нижнего коренного зуба серых полевок) // Журнал общей биологии. Т. 72. № 2. С. 127–139.
Поздняков А.А., Сенотрусова М.М., 2006. Анализ морфотипической изменчивости степной пеструшки // Бюллетень Московское общество испытателей природы, отд. биол. Т. 111. Вып. 3. С. 18–23.
Рахмангулов Р.С., Ишбирдин А.Р., Салпагарова А.С., 2014. Флуктуирующая асимметрия – показатель дестабилизации или поиск путей адаптивного морфогенеза? // Вестник Башкирского университета. Т. 19. № 3. С. 831–834.
Рековец Л.И., 1994. Мелкие млекопитающие антропогена юга Восточной Европы. Киев: Наукова думка. 371 с.
Смирнов Н.Г., Бененсон И.Е., 1980. Изучение изменчивости рисунка жевательной поверхности М1 водяной полевки (Arvicola terrestris L.) методом канонического анализа // Внутри- и межпопуляционная изменчивость млекопитающих Урала. Свердловск: ИЭРиЖ. С. 11–17.
Шишкин М.А., 2006. Индивидуальное развитие и уроки эволюционизма // Онтогенез. Т. 37. № 3. С. 179–198.
Шишкин М.А., 2012. Системная обусловленность формообразования и ее проявления в палеонтологической летописи // Палеонтологический журнал. № 4. С. 3–15.
Юдин Б.С., Потапкина А.Ф., Галкина Л.И., Половинкина Р.А., 1977. Эколого-фаунистический анализ населения мелких млекопитающих (Micromammalia) Центрального Алтая // Фауна и систематика позвоночных Сибири / Труды биологического института. Вып. 31. Новосибирск: Наука. С. 5–31.
Якименко Л.В., Крюков А.П., 1997. Об изменчивости кариотипа восточноевропейской полевки Microtus rossiaemeridionalis (Rodentia, Cricetidae) // Зоологический журнал. Т. 76. № 3. С. 375–378.
Ялковская Л.Э., Бородин А.В., Фоминых М.А., 2014. Модульный подход к изучению флуктуирующей асимметрии комплексных морфологических структур у грызунов на примере нижней челюсти рыжей полевки (Clethrionomys glareolus, Arvicolinae, Rodentia) // Журнал общей биологии. Т. 75. № 5. С. 385–393.
Ялковская Л.Э., Фоминых М.А., Мухачёва С.В., Давыдова Ю.А., Бородин А.В., 2016. Флуктуирующая асимметрия краниальных структур грызунов в градиенте промышленного загрязнения // Экология. № 3. С. 213–220.
Agresti A., 2007. An introduction to categorical data analysis. New Jersey, Hoboken: John Wiley & Sons Inc. 372 p.
Bannikova A.A., Lebedev V.S., Lissovsky A.A. et al., 2010. Molecular phylogeny and evolution of the Asian lineage of vole genus Microtus (Rodentia: Arvicolinae) inferred from mitochondrial cytochrome b sequence // Biol. J. Linn. Soc. V. 99. № 3. P. 595–613.
Bulygina E., Mitteroecker P., Aiello L., 2006. Ontogeny of facial dimorphism and patterns of individual development within one human // Am. J. Phys. Anthropol. V. 131. № 3. P. 432–443.
Cavalli-Sforza L.L., Edwards A.W., 1967. Phylogenetic analysis. Models and estimation procedures // Am. J. Hum. Gen. V. 19. № 3. Pt 1. P. 233–257.
Chaline J., Brunet-Lecomte P., Montuire S., Viriot L., Courant F., 1999. Anatomy of the arvicoline radiation (Rodentia): palaeogeographical, palaeoecological history and evolutionary data // Ann. zool. Fennici. V. 36. P. 239–267.
Cheverud J.M., 1982. Phenotypic, genetic, and environmental morphological integration in the cranium // Evolution. V. 36. № 3. P. 499–516.
Dongen S.V., 2006. Fluctuating asymmetry and developmental instability in evolutionary biology: past, present and future // J. Evol. Biol. V. 19. № 6. P. 1727–1743.
Ehrich T.H., Vaughn T.T., Koreishi S.F. et al., 2003. Pleiotropic effects on mandibular morphology I. Developmental morphological integration and differential dominance // J. Exp. Zool. B: Mol. Dev. Evol. V. 296. № 1. P. 58–79.
Epigenetics: linking genotype and phenotype in development and evolution, 2011. Eds B. Hallgrímsson, B.K. Hall. Berkeley, Los Angeles: Univ. of California Press. 457 p.
Esteve-Altava B., 2017. In search of morphological modules: a systematic review // Biol. Rev. V. 92. № 3. P. 1332–1347.
Floris G., 2015. Palmar intertriradial ridge counts in Sardinians // Adv. Anthropol. V. 5. № 03. P. 137–143.
Galaktionov O.K., Efimov V.M., Lemza S.V., Goltsova T.V., 1982. Principal components method and nonmetric two dimensional scaling method for quantitative dermatoglyphic traits of the taimir aborigines // Am. J. Phys. Anthropol. V. 58. № 1. P. 21–26.
Goswami A., Polly P.D., 2010. Methods for studying morphological integration, modularity and covariance evolution // Quantitative methods in paleobiology / Paleontological Society Papers Series. V. 16. P. 213–243.
Goswami A., Smaers J.B., Soligo C., Polly P.D., 2014. The macroevolutionary consequences of phenotypic integration: from development to deep time // Phil. Trans. R. Soc. B: Biol. Sci. V. 369. № 1649. P. 20130254.
Graham J. H., Shimizu K., Emlen J.M., 2003. Growth models and the expected distribution of fluctuating asymmetry // Biol. J. Linn. Soc. V. 80. № 1. P. 57–65.
Graham J.H., Raz S., Hel-Or H., Nevo E., 2010. Fluctuating asymmetry: methods, theory, and applications // Symmetry. V. 2. № 2. P. 466−540.
Hammer Ø., Harper D.A., Ryan P.D., 2001. PAST: Paleontological statistics software package for education and data analysis // Palaeontol. electron. V. 4. № 1. P. 9.
Jacobi4.ru. Available at: http://www.jacobi4.ru/. Accessed July 16, 2019.
Kharlamova A.V., Trut L.N., Chase K. et al., 2010. Directional asymmetry in the limbs, skull and pelvis of the silver fox (V. vulpes) // J. Morph. V. 271. № 12. P. 1501–1508.
Kellner J.R., Alford R.A., 2003. The ontogeny of fluctuating asymmetry // Am. Nat. V. 161. № 6. P. 931–947.
Kimmel C.B., Hohenlohe P.A, Ullmann B. et al., 2012. Developmental dissociation in morphological evolution of the stickleback opercle // Evol. Dev. V. 14. № 4. P. 326–337.
Klingenberg C.P., 2003. Developmental instability as a research tool: using patterns of fluctuating asymmetry to infer the developmental origins of morphological integration // Developmental instability: causes and consequences. Ed. M. Polak. NY: Oxford University Press. V. 427. P. 442.
Klingenberg C.P., 2008. Morphological integration and developmental modularity // Annu. Rev. Ecol. Evol. Syst. V. 39. P. 115–132.
Klingenberg C.P., 2014. Studying morphological integration and modularity at multiple levels: Concepts and analysis // Phil. Trans. R. Soc. B. V. 369. № 1649. P. 20 130 249.
Klingenberg C.P., 2015. Analyzing fluctuating asymmetry with geometric morphometrics: concepts, methods, and applications // Symmetry. V. 7. № 2. P. 843–934.
Klingenberg C.P., Mebus K., Auffray J.-C., 2003. Developmental integration in a complex morphological structure: how distinct are the modules in the mouse mandible? // Evol. Dev. V. 5. № 5. P. 522–531.
Kovaleva V.Y., Efimov V.M., Litvinov Y.N., 2013. Directional asymmetry of morphological traits during postnatal ontogeny in root vole Microtus oeconomus Pall. (Rodentia, Cricetidae) // J. Siberian Fed. Univ. Biol. V. 2. № 6. P. 115–129.
Kuratani S., 2009. Modularity, comparative embryology and Evo–Devo: developmental dissection of evolving body plans // Dev. Biol. V. 332. № 1. P. 61–69.
Labonne G., Navarro N., Laffont R. et al., 2014. Developmental integration in a functional unit: deciphering processes from adult dental morphology // Evol. Dev. V. 16. № 4. P. 224–232.
Magwene P.M., 2001. New tools for studying integration and modularity // Evolution. V. 55. № 9. P. 1734–1745.
Mitteroecker P., Bookstein F.L., 2007. The conceptual and statistical relationship between modularity and morphological integration // Syst. Biol. V. 56. № 5. P. 818–836.
Mitteroecker P., Gunz P., Bookstein F.L., 2005. Heterochrony and geometric morphometrics: a comparison of cranial growth in Pan paniscus versus Pan troglodytes // Evol. Dev. V. 7. № 3. P. 244–258.
Murren C.J., 2012. The integrated phenotype // Integr. Comp. Biol. V. 52. № 1. P. 64–76.
Olson E.C., Miller R.L., 1958. Morphological Integration. Chicago: University of Chicago Press. 376 p.
Palmer A.R., 1996. From symmetry to asymmetry: phylogenetic patterns of asymmetry variation in animals and their evolutionary significance // PNAS. V. 93. № 25. P. 14 279–14 286.
Palmer A.R., Strobeck C., 1986. Fluctuating asymmetry: measurement, analysis, patterns // Ann. Rev. Ecol. Syst. V. 17. № 1. P. 391–421.
Palmer A.R., Strobek C., 2003. Fluctuating asymmetry analyses revisited // Developmental instability: Causes and consequences. Oxford: Oxford University Press. P. 279– 319.
Parsons P.A., 1990. Fluctuating asymmetry as a epigenetic measure of stress // Biol. Rev. V. 65. № 2. P. 131–145.
Parsons P.A., 1992. Fluctuating asymmetry: a biological monitor of environmental and genomic stress // Heredity. V. 68. № 4. P. 361–364.
Pigliucci M., 2003. Phenotypic integration: studying the ecology and evolution of complex phenotypes // Ecol. Lett. V. 6. № 3. P. 265–272.
Polunin D., Shtaiger I., Efimov V., 2019. JACOBI4 software for multivariate analysis of biological data. BiOrxiv, 803684.
Rabeder G., 1981. Die Arvicoliden (Rodentia, Mammalia) aus dem Pliozän und dem älteren Pleistozän von Niederösterreich // Beitr. Paläont. Österr. Bd. 8. S. 1–373.
Raff R.A., Sly B.J., 2000. Modularity and dissociation in the evolution of gene expression territories in development // Evol. Dev. V. 2. № 2. P. 102–111.
Rasskin-Gutman D., Esteve-Altava B., 2014. Connecting the dots: anatomical network analysis in morphological Evo-Devo // Biol. Theory. V. 9. № 2. P. 178–193.
Renaud S., Alibert P., Auffray J.-C., 2012. Modularity as a source of new morphological variation in the mandible of hybrid mice // BMC Evol. Biol. V. 12. № 1. P. 141.
Schlosser G., 2002. Modularity and the units of evolution // Theor. Biosci. V. 121. № 1. P. 1–80.
Van Valen L.A., 1962. Study of fluctuating asymmetry // Evolution. V. 16. № 2. P. 125–142.
Voyta L.L., Golenishchev F.N., Tiunov M.P., 2013. Analysis of shape and size variation of the first lower molar in the Far-Eastern grey voles of genus Alexandromys (Rodentia: Cricetidae) from Russian fauna using geometric morphometrics // Rus. J. Theriol. V. 12. № 1. P. 19–32.
Wagner G.P., 1996. Homologues, natural kinds and the evolution of modularity // Am. Zool. V. 36. № 1. P. 36–43.
Wagner G.P., Altenberg L., 1996. Complex adaptations and the evolution of evolvability // Evolution. V. 50. № 3. P. 967–976.
Wagner G.P., Pavlicev M., Cheverud J. et al., 2007. The road to modularity // Nat. Rev. Genet. V. 8. № 12. P. 921–931.
Ward J.H., 1963. Hierarchical grouping to optimize an objective function // J. Am. Stat. Ass. V. 58. № 301. P. 236–244.
Winther R.G., 2001. Varieties of modules: kinds, levels, origins, and behaviors // J. Exper. Zool. V. 291. № 2. P. 116–129.
Дополнительные материалы отсутствуют.
Инструменты
Зоологический журнал