

Зоологический журнал, 2021, T. 100, № 5, стр. 524-539
Запись зимней спячки на поверхности резцов грызунов: внутривидовая изменчивость и межвидовые различия
Г. А. Клевезаль a, *, В. А. Лобков b, **, Д. В. Щепоткин a, ***
a Институт биологии развития имени Н.К. Кольцова РАН
119991 Москва, Россия
b Одесский национальный университет имени И.И. Мечникова
65058 Одесса, Украина
* E-mail: klevezal@bk.ru
** E-mail: zoomuz2017@gmail.com
*** E-mail: dm_shchepotkin@mail.ru
Поступила в редакцию 22.03.2020
После доработки 11.05.2020
Принята к публикации 15.05.2020
Аннотация
Описаны внутри- и межвидовая изменчивость паттерна, а также механизм образования “зоны спячки” на поверхности резцов у грызунов трех групп, различающихся по характеру зимней спячки, – сурков, сусликов и хомяков. Рост их резцов продолжается и в период спячки. У сурков и сусликов, не питающихся в период спячки, нагрузка на резец до, в период и после спячки сильно меняется, прирост резца за период спячки минимальный, и на его поверхности образуется “зона спячки”. Она имеет два общих для сурков и сусликов признака – очень узкие приросты и изменение границы эмали и дентина. Межвидовые различия в характере “зоны спячки” у сурков и сусликов могут быть объяснены региональными различиями климата. У хомяков, которые могут питаться в период спячки, прирост резца за это время более существенный, нагрузка на резец меняется меньше, изменения границы эмали и дентина на поверхности резца нет и “зона спячки” отмечается только более узкими и четкими, по сравнению с суточными, приростами. Межвидовые различия записи спячки у хомяков могут быть существенными, вплоть до отсутствия записи. Эти различия, видимо, связаны не столько с климатом, сколько с характером протекания самой спячки.
На поверхности постоянно растущих резцов всех грызунов, как зимоспящих, так и не зимоспящих, видны суточные валики – приросты, четкость которых значительно варьирует. Морфология и изменчивость суточных приростов были описаны ранее (Клевезаль, Щепоткин, 2017). В настоящей работе мы анализируем данные по изменчивости структуры приростов на том участке поверхности резца, который формируется у зимоспящих грызунов в период зимней спячки11.
Запись спячки в дентине резцов (не на поверхности резца) – “зона спячки” – хорошо заметна на окрашенных срезах декальцинированного резца и у разных видов она выглядит по-разному. У мышовок рода Sicista это – узкая интенсивно окрашенная полоса (Klevezal, Mina, 1990). У сурков рода Marmota и сусликов рода Spermophilus это – сложный комплекс контрастных слоев уменьшающейся ширины с группой очень узких слоев (в несколько раз более узких, чем суточные слои) (Клевезаль, Суховская, 1995; Klevezal, 1996; Трунова, Лобков, 1997; Трунова, Клевезаль, 1999).
На поверхности резцов “зона спячки” была сначала обнаружена у сурка Marmota flaviventris (Rinaldi, 1999). Она выглядела как группы узких приростов, разделенных контрастными полосками, с утолщением или изгибом края эмали на этом участке. Затем запись периода спячки была описана у нескольких видов североамериканских и европейских сусликов Spermophilus (Goodwin et al., 2005; Goodwin, Ryckman, 2006; Клевезаль, Лобков, 2008), серого сурка (Marmota baibacina) (Клевезаль, Щепоткин, 2012), хомяков рода Mesocricetus (Клевезаль и др., 2012, 2018; Batavia et al., 2013), хомячков рода Allocricetulus (Клевезаль и др., 2015). Результаты этих работ показывали, что по числу суточных приростов, сформированных после “зоны спячки”, можно определить время пробуждения грызуна, при этом характер “зоны спячки” у разных видов был несколько различен, отмечалась и его внутривидовая изменчивость.
Таким образом, запись спячки на поверхности резцов была обнаружена у сурков, сусликов и хомяков. Эти зимоспящие грызуны существенно различаются по характеру спячки. Сурки и суслики считаются зимоспящими жиронакопителями, переживающими зиму исключительно за счет жировых запасов. Сурки не делают запасов корма на зиму. У сусликов разных видов либо накопление запасов отмечено не регулярно и не во всех районах (Чернявский, 1977; Бадмаев, 2007; Алфимов, Берман, 2009), либо запасы корма обнаружены только у самцов, да и то не у всех (Charge, 2001). Даже те суслики, которые собирают перед спячкой значительные запасы корма, по-видимому, потребляют их в норах перед спячкой или после пробуждения весной (Калабухов, 1985). Сурки, за редким исключением, зимуют в группах по несколько особей в одной зимовочной камере, а суслики – поодиночке. Хомяки не все впадают зимой в спячку, а те, что впадают (Cricetus, Mesocricetus), делают большие запасы корма, которые поедают во время периодических пробуждений (Калабухов, 1985). Так же как и суслики, хомяки зимуют поодиночке.
Мы исследовали морфологию записи спячки на поверхности резцов сурков, сусликов и хомяков, чтобы ответить на два вопроса: являются ли обнаруженные в указанных выше работах межвидовые и внутривидовые особенности рисунка “зоны спячки” отражением соответствующих особенностей протекания зимнего периода жизни особи и какова причина образования “зоны спячки” на поверхности резца.
МАТЕРИАЛ И МЕТОДИКА
Исследованный материал представлен в табл. 1. Как и в большинстве упомянутых выше работ, мы детально исследовали нижние резцы, преимущественно их медиальную стенку. Предварительно очищенные от пленок резцы фотографировали на линейке с ценой деления 0.5 мм под бинокуляром в обычном отраженном свете, а резцы от инъецированных тетрациклином особей также и в ультрафиолетовом свете. Кроме того, использовали цифровой микроскоп Keyence VHX-1000. На нем проводили съемку всего резца в высоком разрешении, фоном служил лист клетчатой бумаги с линиями через 2.5 мм.
Таблица 1.
Исследованный материал
| Вид | Колич. особей | Район сбора | Даты сбора | Примечание |
|---|---|---|---|---|
| Серый сурок (Marmota baibacina) |
112 | Тянь-Шань,
высокогорье Джеты-Огузские сырты |
18.04–5.05 и 27.05–19.07. 1967 | Резцы предоставлены зоологом Киргизской противочумной станции А.А. Михайлютой вместе с биологическими данными по этой популяции в год сбора материала |
| Длиннохвостый
суслик (Spermophilus undulatus) |
30 | Казахский Алтай. южный берег оз. Маркаколь | 21.05–24.06. 1976 | Черепа из коллекции лаборатории постнатального онтогенеза ИБР РАН |
| 5 | Юго-Восточное Прибайкалье (Бурятия, Тарбагатайский р-н) | 29.03.1989 | Черепа предоставлены Б.Б. Бадмаевым и переданы в Зоологический музей МГУ | |
| Крапчатый суслик (Spermophilus suslicus) |
74 | Северо-Западное Причерноморье | 19–28.01.2007; 7–11.03.1990; 24–26.03, 15.04 и 7.05.1987; 7.05.1985 | Резцы из коллекции Зоологического музея Одесского национального университета |
| Хомяк Брандта (Mesocricetus brandti) |
20 | Ширакское нагорье (северо-восток Армянского нагорья, окрестности Гюмри) | 17–19.04.1971 | Резцы из коллекции лаборатории постнатального онтогенеза ИБР РАН |
Измерения проводили в программе Photoshop и в программе цифрового микроскопа. Длина резца – это длина внешней дуги, измеренная по наружному краю эмали. Все промеры, вектор которых направлен вдоль продольной оси зуба, мы называем длиной. Исключение составляет промер “ширина прироста”, за которым оставлен термин “ширина”, несмотря на то что промер имеет вектор вдоль продольной оси зуба. Длину отдельных участков на поверхности резца и ширину приростов измеряли в верхней трети – середине ширины резца. Многочисленные сравнения показали, что промеры длины резца и его участков по внешней дуге и по середине стенки различаются, в зависимости от кривизны нижних резцов, на 11–13%, в среднем на 12%. При необходимости сравнений длины участков или ширины приростов с длиной резца мы вводили поправку 0.12. Ошибка измерений, как было показано ранее (Клевезаль, 2010), не превышает 6–7%.
Терминология. Мы называем базальной часть резца, расположенную в альвеоле, где лежит ростовая зона, и апикальной – свободную часть резца, подверженную стачиванию. В поперечном сечении резец имеет форму эллипса с более плоскими боковыми стенками: медиальной (обращенной к языку) и латеральной (обращенной к щеке). Стенки резца состоят из дентина, который на внешней стенке и прилегающих частях боковых стенок покрыт толстым слоем эмали. Граница эмали хорошо видна на поверхности боковых стенок резца. Изменение границы эмали (ИГЭ) выглядит как отклонение края эмали от дентина с образованием выемки в придентиновой части эмали.
Приросты – это поперечные валики на поверхности резца, образующиеся в процессе роста в длину (прорезания) резца, их границы видны под определенным углом падения света.
Слои дентина (суточные слои) – это разнокачественные образования в толще дентина, образующиеся в результате утолщения стенок резца; они заметны на срезах (лучше – на окрашенных) резца.
Мы называем приросты четкими, если границы их легко различаются, и нечеткими, когда разграничить отдельные приросты трудно. Четкие приросты могут быть контрастными, когда рельеф поверхности хорошо выражен, и слабо контрастными, но различимыми, когда рельеф выражен плохо, хотя приросты различимы. Приросты могут быть простыми или сложными, в последнем случае четко ограниченный прирост разделен на несколько более узких.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Запись спячки у сурков и сусликов
Серый сурок (Marmota baibacina)
У исследованных особей суточные приросты, как правило, были хорошо выражены. На их фоне, на всех резцах сурков, добытых весной – в начале лета, т.е. перезимовавших хотя бы однажды, была обнаружена “зона спячки” (рис. 1 и 2). Она диагностировалась по двум основным признакам: по ИГЭ, которое апикально начинается более полого и базально заканчивается более круто, часто утолщением (“наплывом”) эмали, и по участку группирующихся очень контрастных приростов. Кроме этих, наиболее заметных, признаков, часто видны косые складки дентина в приэмалевой четверти стенки; они идут от границы с эмалью под углом к этой границе, начинаются близ апикальной части ИГЭ и оканчиваются близ “наплыва” эмали. В большинстве случаев в районе ИГЭ виден участок очень узких, обычно слабо контрастных, приростов дентина, с шириной отдельного прироста от неизмеримо малой до 70–100 мкм (по меньшей мере, в 5–9 раз ýже нормального суточного прироста). Эмаль на внешней стенке близ ИГЭ может быть утолщенной и бугристой, вплоть до образования выраженного бугорка. Наружные контуры резца в “зоне спячки” немного меняются, возникают небольшие выпуклости и впадины на большой и на малой дугах (соответственно, на внешней и внутренней стенках) резца (рис. 2). Все эти признаки подвержены индивидуальной изменчивости.
Рис. 1.
Нижний резец серого сурка. Общий вид, 1 – суточные приросты. Врезка – “зона спячки”, варианты ее деталей – в столбцах. С – косые складки дентина, ИГЭ – изменение границы эмали, Н – “наплыв” эмали, К – контрастные приросты.
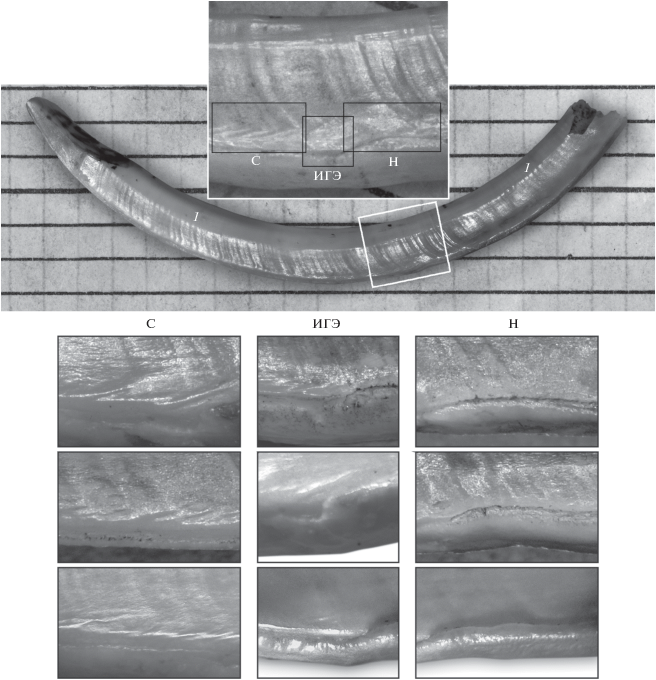
Рис. 2.
Участок резца серого сурка в районе “зоны спячки”. Стрелками указаны изменения контура. Остальные обозначения как на рис. 1.

На участке резца апикальнее “зоны спячки” приросты несколько менее четкие и менее равномерные, чем предшествовавшие им суточные. Четко определить границы такого участка крайне сложно, и ошибка измерения велика, но поскольку она не систематическая, мы измерили длину этого участка у 112 особей и сочли возможным сравнить ее у сурков разного возраста, добытых на разных территориях (табл. 2). Были выявлены две четкие тенденции: локальное и возрастное изменения длины этого участка. Выборки 1, 2, 3, территориально более или менее близкие друг к другу, имели высокую вариабельность значений признака и различия между ними статистически не значимы. Четвертая выборка, которая значимо отличалась от них и имела существенно меньшую вариабельность признака, по данным Михайлюты (1988), отдалена от них территориально, и на этой территории численность сурков меньше, чем на первых трех. Во всех выборках присутствовали как годовики, так и взрослые особи до 8 лет и старше22. Во всех выборках длина участка нечетких приростов значимо увеличивалась с возрастом (см. табл. 2). Это увеличение определялось главным образом существенно меньшей длиной участка у годовиков, которые были сеголетками перед спячкой. Так, в выборках 1–3 средняя длина участка нечетких приростов у годовиков была 10.8 мм, а у взрослых особей (старше 2 лет) – 29.2 мм. Подсчет суточных приростов (приблизительный, из-за нечеткости приростов) выявил ту же тенденцию: в выборке 1 на этом участке у трех годовиков было 15–20 суточных приростов, а у трех особей старшей возрастной группы 120–130 суточных прироста.
Таблица 2.
Длина участка менее равномерных и менее четких приростов до “зоны спячки” на резцах серого сурка
| Выборка | Колич. особей | Длина участка (средняя ± сигма), мм |
Значимость различий | Корреляция длины участка с возрастом особи |
|---|---|---|---|---|
| 1. 3-й Майнок | 30 | 26.77 ± 18.61 | t1–4 = 3.96 > t0.001 | 0.47 > r0.01 |
| 2. 4-й Майнок | 25 | 19.54 ± 16.14 | 0.57 > r0.01 | |
| 3. Тез | 23 | 25.82 ± 17.94 | 0.68 > r0.01 | |
| 4. Акташ | 34 | 13.00 ± 4.14 | 0.47 > r0.01 |
ИГЭ (см. рис. 1 и 2) сильно варьировало по степени выраженности, по длине, и по глубине. Измерить длину ИГЭ часто нелегко из-за нечетко определяемой его апикальной границы, тогда как базальная граница определяется легко по началу “наплыва”. Несмотря на большую ошибку такого измерения, выявлена четкая тенденция: у годовиков ИГЭ в среднем было мельче и короче, чем у взрослых, Так, у годовиков длина ИГЭ – 1.22 ± ± 0.37 мм (n = 12), у взрослых (старше 2 лет) – 2.98 ± 1.31 мм (n = 15), различия значимы (t = 4.96). Вероятнее всего эта тенденция определяется меньшей длиной резца у годовиков, чем у старших, т.к. корреляция длины ИГЭ и длины зуба для всей выборки существенна (линейный коэффициент корреляции r = +0.42 , n = 62), а без учета годовиков практически отсутствует (r = +0.10, n = 50).
“Наплыв” эмали в конце ИГЭ (см. рис. 1 и 2) был выражен не всегда, его длина варьировала от 1.6 до 6 мм, но никаких различий между особями разного пола, возраста или локалитета не обнаружено. Поверхность эмали (на внешней стенке) у ИГЭ была гладкой у годовиков и бугристой у старших особей.
Узкие приросты были видны у большинства особей. В 78% резцов они были на том же участке резца, что и ИГЭ, у остальных начинались апикальнее начала ИГЭ. При этом только у половины длина ИГЭ и длина участка узких приростов были примерно равны, у остальных длина ИГЭ была больше или меньше длины участка узких приростов. Различий между особями разного пола, возраста или локалитета не обнаружено. У взрослых особей (без годовиков), у которых ее можно было измерить, длина участка узких приростов была в среднем 2.5 ± 0.8 мм; длина резца (по внешней дуге) в среднем была 65.9 мм и, следовательно, отношение длина участка узких (с поправкой 0.12) к длине резца 0.042.
Ранее (Клевезаль, Ануфриев, 2013а) на той же выборке при использовании балльной оценки “зоны спячки”, по степени выраженности ИГЭ и узких приростов, были обнаружены статистически значимые различия между зимовавшими один раз и остальными: у молодых “зона спячки” была выражена хуже.
Косые складки (см. рис. 1), разной степени проявления, присутствовали у 90% особей. По степени выраженности (по трехбалльной шкале) не было различий между возрастными группами и между самцами и самками по частоте встречаемости разных вариантов.
Контрастные приросты (см. рис. 1 и 2) разной ширины (в 1.5–5 раза меньше ширины суточных приростов) были видны на участке резца от конца ИГЭ и над “наплывом”. Они образовывали группы разной величины и занимали участок разной длины, от 2 до 11 мм. Длина участка не коррелировала с возрастом и не различалась у самцов и самок. Иногда внутри были видны более узкие приросты. Есть косвенные свидетельства того, что эти контрастные приросты формируются после выхода сурка из спячки. Так, общее число приростов, сформированных после конца ИГЭ (т.е. контрастные приросты плюс остальные в базальной части), у добытых в апреле – первых числах мая сурков было 11–38. Если принять, что все эти приросты суточные, то сурки проснулись в период с 29 марта до 11 апреля. Это соответствует данным Михайлюты (1988), что в исследованном районе в год сбора материала первый выход на поверхность был отмечен 30 марта, а массовый выход – 3–8 апреля. Следовательно, все приросты, начиная с контрастных, сформированы после выхода сурка из спячки. Различий между особями разного пола, возраста или локалитета нами не обнаружено.
Таким образом, анализ индивидуальной изменчивости разных параметров “зоны спячки” на нашем материале выявил локальные и возрастные различия участка нечетких приростов перед “зоной спячки” и отличия зимовавших 1 раз от остальных по общей степени выраженности “зоны спячки”. Если различия отдельных параметров “зоны спячки” отражают различия каких-то особенностей зимней спячки, то они могут быть сходны у особей, зимующих вместе в одной норе. Ли с соавторами (Lee et al., 2015) обнаружили большое сходство кривых изменения температуры тела за период спячки у сурков Marmota broweri, зимовавших вместе. К сожалению, у нас не было данных о том, из каких нор были исследованные нами сурки.
Черношапочный сурок (Marmota camtschatica)
Ранее (Клевезаль, Ануфриев, 2013а) были исследованы резцы 4 особей, добытых на западном побережье Камчатки в мае 1971 г. и зимовавших не менее 2 раз. У них и суточные приросты, и “зона спячки” были выражены существенно хуже, чем у серого сурка. У 3 из 4 особей кроме базальной “зоны спячки”, которая, судя по сформированным после нее суточным приростам, являлась записью последней зимовки, была обнаружена еще одна “зона спячки” в апикальной части резца. Количество суточных приростов между этими двумя зонами варьировало, и происхождение апикальной “зоны спячки” осталось неясным.
Длиннохвостый суслик (Spermophilus undulatus)
У длиннохвостого суслика из предгорий Алтая суточные приросты были хорошо выражены. “Зона спячки” была хорошо видна у 20 и была плохо видна у 1 из 30 перезимовавших особей, у остальных, в процессе роста и стачивания резца, она, видимо, вышла в область стачивания резца. Из признаков “зоны спячки”, характерных для сурков, у сусликов было два признака: ИГЭ и участок узких приростов (по 40–80 мкм, в 5 и более раз ýже суточных) или вообще без видимых приростов (рис. 3). Поверхность медиальной стенки резца в зоне спячки была заметно изменена у 70% особей, чаще всего это было уплощение стенки под ИГЭ, реже (в 20%) уплощение или небольшое вздутие до ИГЭ. Длина “зоны”, ее рисунок и ширина приростов до и после “зоны” (более узкие, средней ширины и более широкие, в разных сочетаниях) были разными у разных особей. Ни “наплыва” эмали, ни косых складок, ни контрастных приростов, описанных у серого сурка, мы не наблюдали, но было отмечено некоторое изменение контура резца в районе “зоны спячки”.
Рис. 3.
Нижний резец длиннохвостого суслика. Общий вид и “зона спячки” (врезка). ИГЭ – изменение границы эмали, 1 – суточные приросты.

Как и в случае серого сурка, измерить длину ИГЭ было не всегда возможно из-за неопределенности границ. В тех случаях, когда это было возможно, отмечена очень слабая тенденция увеличения длины ИГЭ с возрастом, как и у сурка. У перезимовавших 1 зиму (n = 8) средняя длина была 1.78 (1.3–2.4) мм, у переживших 2–3 зимы33 (n = 9) – 2.27 (1.3–4.5) мм. Связь длины ИГЭ с длиной резца у сусликов менее очевидна, чем у сурков, поскольку длина резца у годовиков несущественно меньше (средняя 31.5 мм), чем у особей 2–3 лет (средняя 31.8 мм). В большинстве случаев узкие приросты были видны на участке с ИГЭ, но в двух случаях чуть апикальнее и в двух случаях чуть базальнее границ ИГЭ.
У 10 экз., у которых можно было измерить длину участка узких приростов, эта длина была в среднем 0.84 мм при средней длине резца 32.2 мм. Среднее отношение длины участка узких приростов (с поправкой 0.12) к длине резца – 0.029. Узкие приросты на всем этом участке у трех особей нельзя было сосчитать, у остальных можно было видеть от 10–11 до 16–18 приростов.
Таким образом, на нашем материале у длиннохвостого суслика обнаружены три признака зоны спячки: ИГЭ, уплощенная поверхность стенки резца и узкие приросты или участок без видимых приростов в районе ИГЭ.
На резцах 5 экз. длиннохвостого суслика из Бурятии суточные приросты были видны очень хорошо, “зона спячки” была представлена ИГЭ с участком узких приростов или без видимых приростов. За “зоной спячки” было сформировано 10–14 суточных приростов, что соответствовало времени пробуждения сусликов (Бадмаев, 2007). Никаких отличий длиннохвостого суслика из Бурятии от сусликов с Алтая по характеру “зоны спячки” на резцах не обнаружено.
Крапчатый суслик (Spermophilus suslicus)
У крапчатых сусликов резцы были меньшего размера, чем у длиннохвостых, суточные приросты были более вариабельны по четкости и ширине, часто они были сложные. На многих резцах были видны участки несколько деформированной стенки с неясными приростами. В большинстве случаев основной признак “зоны спячки” – ИГЭ, с узкими приростами или без них – был выражено хуже, чем у длиннохвостых сусликов; четко ИГЭ определялось лишь у отдельных особей. Особенно плохо была выражена апикальная граница ИГЭ, тогда как его базальная граница иногда подчеркивалась заметным изменением контура резца (рис. 4). Все исследованные особи были перезимовавшими, но у 4 из них четких признаков “зоны спячки” не было обнаружено. ИГЭ может быть плохо выражено, если “зона спячки” находится в самой базальной части резца, поскольку при очистке зуба мягкая базальная эмаль повреждается и ИГЭ может быть не видно. Этим можно объяснить то, что не обнаружено ИГЭ у 3 из 26 особей, добытых в январе и марте вскоре после выхода из спячки. Однако оно не было обнаружено также у одной из 48 добытых в апреле и начале мая особей, хотя сточиться эта область резца еще не могла. Обнаружению “зоны спячки” у крапчатых сусликов мешали участки деформированной стенки резца с неясными приростами (см. рис. 4). Такая деформация, выраженная в разной степени, от вздутия до уплощения, над и/или перед (апикальнее) ИГЭ, была видна у двух третей особей.
Рис. 4.
Нижний резец крапчатого суслика. Общий вид и “зона спячки” (врезка). ИГЭ – изменение границы эмали, 1 – суточные приросты, 2 – участки с деформированной стенкой.

У сусликов исследованной нами популяции установлено повторное впадение в спячку весной спустя некоторое время после пробуждения (Лобков, 1999). У одной из 10 особей, добытых 24–31 марта 1987 г., на резце были видны две “зоны спячки” с нечеткими ИГЭ, между которыми было 14 приростов уменьшающейся ширины. По данным полевых наблюдений В.А. Лобкова, в ту весну суслики начали просыпаться после 23 февраля (в это время были обнаружены первые норы-веснянки, свидетельствующие об окончании зимней спячки). Потом погода ухудшилась, входы в норы пробудившихся сусликов наблюдатель забил землей, и следующих выходов из норы не было до конца марта. По-видимому, некоторые суслики, проснувшиеся первыми, еще раз впали в спячку, о чем и свидетельствует поверхность резца одной особи.
Контрастность климата как возможная причина межвидовых различий записи спячки на резцах сурков и сусликов
Характер “зоны спячки” у серого сурка высокогорья Тянь-Шаня в целом соответствовал описанию “зоны спячки” у Marmota flaviventris, которое дала Ринальди в своей пионерской работе (Rinaldi, 1999). “Зона спячки” у черношапочного сурка западного побережья Камчатки была выражена существенно хуже, чем у серых сурков Тянь-Шаня: признаки, характеризующие “зону спячки”, выражены значительно менее четко. Выборка с Камчатки слишком мала для серьезных выводов, но все же дает возможность предположить, что есть связь различий характера “зоны спячки” с условиями зимнего периода у сурков этих популяций. Так, четкий рисунок “зоны спячки” обнаружен у сурков, обитавших в резко континентальном климате высокогорья Тянь-Шаня, и существенно менее четкий у сурков, обитавших в значительно менее контрастном климате западного побережья Камчатки.
Сходная связь записи спячки с климатическими условиями прослеживается и у сусликов. Существенных различий в характере “зоны спячки” между длиннохвостыми сусликами из предгорий Алтая и из Прибайкалья не обнаружено (правда, выборка из Прибайкалья была мала). У них “зона спячки” соответствовала таковой, описанной у трех видов североамериканских сусликов, Spermophilus elegans из Монтаны, S. richardsonii из Южной Альберты и S. parryii с территории Юкона (Goodwin et al., 2005). У исследованного нами крапчатого суслика Причерноморья ИГЭ было выражено существенно менее четко, чем у длиннохвостого и чем у североамериканских сусликов (судя по описаниям, данным в работах). Это соответствует климатическим особенностям района обитания сусликов: климат Причерноморья менее континентальный, чем в местах обитания исследованных длиннохвостых сусликов (предгорий Алтая и Прибайкалья) и в местах обитания исследованных Гудвином с соавторами (Goodwin et al., 2005) североамериканских сусликов.
Влияние климатических условий на запись спячки в резцах исследовала Селькова (2003), правда, запись спячки она анализировала не на поверхности резцов, а в дентине (на окрашенных поперечных срезах резцов). Она сравнивала запись спячки в резцах крапчатых сусликов, перезимовавших в разные годы, сгруппировав характер зим в три категории: теплые, холодные и умеренные. В качестве показателей категории зимы она использовала среднюю температуру и соотношение числа дней с положительной и отрицательной среднесуточной температурой с декабря по февраль. По ее данным, ширина “зоны спячки” значимо не различалась во всех 7 попарных сравнениях, когда сравнивали зимы разных лет внутри одной категории, но статистически значимо различалась в 19 из 21 сравнениях, когда попарно сравнивали годы из разных категорий. Эти результаты показывают, что особенности зимнего периода отражаются на записи спячки в дентине, а следовательно, имеет смысл искать их и на поверхности резца.
Запись спячки у хомяков
Хомяк Брандта (Mesocricetus brandti)
Все исследованные особи были добыты в апреле и, следовательно, перезимовали. Основные приросты, которые мы, по аналогии с другими грызунами, считали суточными, были средней четкости. Ширина таких приростов варьировала, в среднем была 300–400 мкм. На их фоне выделялась группа более четких приростов, ширина которые сначала (от апикального конца) уменьшалась до очень узких, а затем увеличивалась (рис. 5), ИГЭ не отмечено. Сами узкие приросты были более широкими, чем у сусликов, и их, как правило, можно было сосчитать. Картина была сходна с той, что ранее была описана для хомяка Радде (Mesocricetus raddei) из Дагестана. Так же как было сделано при исследовании хомяка Радде (табл. 2 в статье по хомяку Радде (Клевезаль и др., 2018)), мы выделили три участка более четких приростов: апикальный участок приростов уменьшающейся ширины, центральный участок узких приростов и базальный участок приростов увеличивающейся ширины (табл. 3). Как видно из таблицы, граница между участками весьма условна и число приростов, их ширина и ее изменение сильно варьируют.
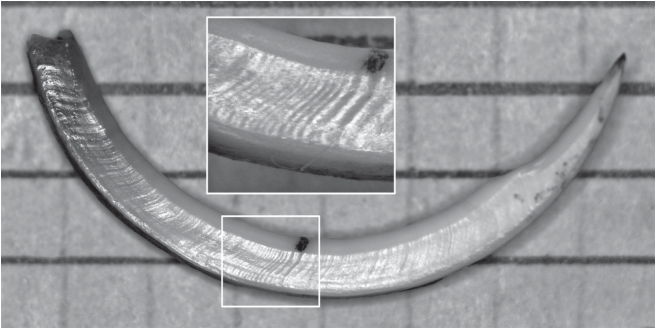
Таблица 3.
Характер зоны с относительно узкими и четкими приростами на резцах хомяка Брандта
| №, пол | Апикальная часть. Ширина приростов уменьшается |
Центральная часть. Узкие приросты | Базальная часть. Ширина приростов увеличивается | |||
|---|---|---|---|---|---|---|
| Число приростов | Ширина одного прироста, мкм | Число приростов |
Ширина одного прироста, мкм | Число приростов | Ширина одного прироста, мкм | |
| 124 ♀ | 4 | 165 | ≈6 | ≈75 | 6 + 3 | 180 + 220 |
| 125 ♀ | 3 + 4 | 170 + 90 | ? | Не более 65 | 5 + 4 | 120 + 150 |
| 126 ♀ | 5 + 3 | 150 + 110 | 11+? | От 65 до 75 | 4 + 2 | 95 + 190 |
| 127 ♀ | 6 + 4 | 180 + 100 | От 7 до 10 | От 60 до 90 | 2 + 2 + 3 + 3 | 115 + 150 + 110 + 160 |
| 134 ♂ | 9 | От 160 до 140 | 5 или 6 | От 45 до 90 | 8 или 9 | 180 |
| 136 ♂ | 6 | От 170 до 90 | 4 | От 60 до 80 | 6 или 7 | От 115 до 160 |
| 137 ♂ | 5 | От 180 до 150 | 10 | От 65 до 75 | 9 | От 90 до 170 |
| 141 ♂ | 10 | От 190 до 130 | 3 или 4 | 90 | 8 | От 180 до 300 |
| 143 ♂ | 4 | От 230 до 120 | 12 или 13 | От 90 до 50 | 2 | 135, 278 |
| 144 ♂ | 7 | От 210 до 170 | 12 | От 50 до 70 | 15 + 7 | 100 + 170 |
Для хомяка Радде на материале от особей из природной популяции и от одного экземпляра, перезимовавшего с имплантированным термодатчиком в условиях, приближенных к естественным, было показано, что участок на поверхности резца между первым приростом и последним приростом, более четким по сравнению с остальными приростами, можно считать “зоной спячки” (Клевезаль и др., 2018). У хомяков Брандта из Армении мы подсчитали количество суточных приростов, сформированных после последнего четкого, т.е. после “зоны спячки”. У 9 самок оно варьировало от 5 до 14, у 11 самцов от 21 до 35. Следовательно, хомяки проснулись в марте – начале апреля и самцы просыпались раньше самок. Последнее характерно для многих грызунов, проводящих спячку в индивидуальных норах, и может служить косвенным подтверждением того, что участок более четких приростов у хомяка Брандта является именно записью спячки.
Запись периода спячки у хомяка Брандта в виде серии более узких и четких приростов на фоне плохо выраженных суточных приростов была подтверждена при исследовании одного экземпляра, зимовавшего с имплантированным термодатчиком в Дагестане в условиях, приближенных к естественным (Zaytseva et al., 2020), а также у хомяков Брандта из лабораторной колонии при индуцированной зимней спячке (Batavia et al., 2013).
Особенности “зоны спячки” у хомяков
Исследованные выборки хомяков рода Mesocricetus, хомяка Брандта и хомяка Радде, были взяты из горных районов со сходным континентальным климатом, и запись спячки на резцах у них сходна. У представителей этих видов, перезимовавших с вшитым термодатчиком в одинаковых, приближенных к естественным, условиях, количество более узких и четких приростов на поверхности резца соответствовало количеству эпизодов нормотермии за период спячки. Однако у хомяка Радде было соответствие динамики изменения ширины приростов динамике изменения длительности периодов нормотермии (Клевезаль и др., 2018), а у хомяка Брандта такого соответствия обнаружено не было (Zaitseva et al., 2020). От записи спячки у сусликов запись спячки хомяков Mesocricetus отличается отсутствием ИГЭ, и определяется “зона спячки” только увеличением четкости приростов при характерном уменьшении их ширины до очень узких. Сами узкие приросты были во многих случаях шире, чем узкие приросты у сусликов. Характерно также то, что у этих хомяков “зона спячки” занимает значительно бόльшую часть резца, чем у сусликов. Это четко видно при сравнении параметров “зоны спячки” у длиннохвостого суслика и хомяка Брандта: у последнего длина “зоны спячки” относительно длины резца почти в два раза больше, чем у суслика (табл. 4). Обращает на себя внимание и значительно бόльшая изменчивость этого параметра у этого хомяка, чем у суслика.
Таблица 4.
Относительная длина “зоны спячки” у 10 экз. сусликов и 10 экз. хомяков
| Вид | Длина резца, мм | Ширина всей зоны спячки, мм* | Доля зоны спячки от длины резца |
|---|---|---|---|
| Длиннохвостый суслик | 29.5–34.2 | 1.5–2.35 | 0.05–0.07 |
| Хомяк Брандта | 22.1–25.1 | 2.08–5.09 | 0.09–0.21 |
Иная картина была обнаружена у обыкновенного хомяка (Cricetus cricetus) из окрестностей г. Симферополя, где климат мягкий. Только у одной из 21 особи (включая двух, перезимовавших в естественных условиях с термодатчиком) была обнаружена запись спячки, напоминающая таковую у хомяков Брандта и Радде. Более того, не было обнаружено такой записи спячки и у двух обыкновенных хомяков из того же района, что и исследованные хомяки Радде, перезимовавшие с термодатчиками в сходных, приближенных к естественным, условиях (Клевезаль и др., 2020).
Механизм образования суточных приростов и “зоны спячки” на поверхности резцов
В экспериментальных работах, проведенных на крысах (Ohtsuka et al., 1998; Ohtsuka-Isoya et al., 2001), было показано, что суточные слои в толще дентина являются результатом околосуточных (циркадных) ритмов синтеза коллагена и секреторной активности формирующих дентин одонтобластов, и что эти ритмы регулируются супрахиазмоидальным ядром мозга. Поскольку суточные приросты на поверхности резца соответствуют суточным слоям в толще дентина по количеству и по степени четкости (Клевезаль, Лобков, 2008; Клевезаль, Ануфриев, 2013а), можно считать природу суточных слоев в толще дентина и суточных приростов на поверхности резца одинаковой. Это подтверждает и экспериментально выявленный суточный ритм роста в длину (прорезания) резца (Michaeli, Weinreb, 1968; Chiba et al., 1976). Остается вопрос, почему этот ритм роста резца фиксируется на поверхности в виде валиков – приростов.
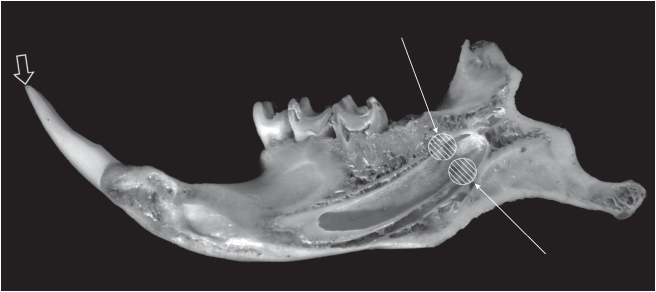
Этот волнообразный профиль суточных приростов поверхности резца отражает изгибы всей стенки резца в его базальной, ближайшей к ростовой зоне, части (Клевезаль, Щепоткин, 2017), и возникает, вероятнее всего, в силу суточных изменений механической нагрузки на зуб. Биомеханический анализ резцов грызунов (Srivastava, 1998) показал, что при функционировании резцов местом приложения механической нагрузки является апикальный конец резца, но дугообразный изгиб резца приводит к тому, что бόльшая часть нагрузки приходится на базальную часть зуба (рис. 6). В этой части и расположена ростовая зона резца, где формируется кольцо дентина, определяющее контур резца. Таким образом, контур резца формируется в самой базальной его части близ ростовой зоны в течение очень короткого периода и далее уже не меняется. В эксперименте на крысах было показано, что увеличение функциональной нагрузки на резец (длительностью 3 недели) привело к изменению формы резца – заметному округлению его поперечного сечения при сохранении объема зубных тканей (Steigman et al., 1989). Можно полагать, что внутрисуточные изменения механической нагрузки также несколько меняют форму резца, в результате чего и возникает волнообразный профиль суточных приростов.
Рис. 6.
Продольный разрез нижней челюсти серого сурка вдоль канала резца. Широкая стрелка – место приложения механической нагрузки, тонкие стрелки – область передачи большей части нагрузки в ростовой зоне.
Таким образом, формирование суточных приростов-валиков на поверхности резца определяется двумя основными процессами: суточным (околосуточным) ритмом роста тканей зуба и суточным ритмом механической нагрузки на зуб. Последний, в свою очередь, определяется суточной активностью грызуна.
Такая точка зрения дает возможность объяснить некоторые особенности суточных приростов у серого сурка, “зона спячки” которого нами изучена детально.
Во-первых, это образование менее четких и менее равномерных приростов, сформированных до “зоны спячки” (апикальнее ее). Выше было прослежено локальное и возрастное изменения длины участка таких приростов (см. табл. 2). Длина участка нечетких приростов у годовиков (которые были сеголетками в период формирования этого участка) была значимо меньше, чем у взрослых особей. Значение этого показателя на территориально близких участках было сходным и значимо отличалось от такового четвертой выборки, территориально удаленной. Это дает основание предположить, что нечеткость приростов, образованных перед спячкой, отражает период менее активного питания перед засыпанием. Этот период у сеголетков, перед первой спячкой, гораздо короче, чем у старших особей (Михайлюта, 1988), поскольку им нужно накопить необходимое для спячки количество жира. Продолжительность периода может различаться и у обитателей разных территорий.
Во-вторых, это формирование очень контрастных приростов у серых сурков после выхода из спячки. Их формирование приходится на раннюю весну, когда, как пишет Стогов (1956), до начала вегетации сурки питаются более жестким кормом: поедают оставшуюся на корню прошлогоднюю траву – ветошь, выкапывают луковицы эфемероидов и обгрызают дерновинки многолетних злаков и полыней. По-видимому, повышенная механическая нагрузка на резцы и определяет повышенную контрастность приростов.
Существенное изменение механической нагрузки на резец происходит у сурков и сусликов, которые не питаются в течение нескольких месяцев зимней спячки. Поэтому наружные контуры резца в “зоне спячки” у них немного меняются. Поскольку пролиферация и минерализация эмали и дентина в ростовой зоне резца идут не синхронно (Smith, Warshawsky, 1976; Smith, 1998; Simmer et al., 2010), при замедлении и возобновлении роста до и после спячки образуется ИГЭ, а на поверхности эмали часто возникает бугристость. Возможно, определенную роль играют и изменения в шарпеевских волокнах (прикрепляющих зуб к альвеоле), которые отмечены при изменении нагрузки на зуб (Silva, Merzel, 2004).
Таким образом, ИГЭ образуется в связи с изменением нагрузки на зуб в период спячки. Возможно, базальный конец ИГЭ отмечает не выход из спячки как таковой (восстановление нормотермии), а возобновление питания после выхода из спячки. Этому есть несколько косвенных доказательств. Гудвин с соавторами (Goodwin et al., 2005) на резцах особей S. richadsonii, для которых была известна продолжительность спячки, не нашли связи продолжительности спячки и длины “зоны спячки” на резце. Об этом же косвенно свидетельствуют контрастные приросты у сурков, часто (но не всегда) начинающиеся не после ИГЭ, а в базальной его части. Об этом же говорят неопубликованные данные А.И. Ануфриева, который ввел тетрациклин четырем длиннохвостым сусликам, перезимовавшим в глубокой штольне при температуре, близкой к естественной в природе (Ануфриев, Ядрихинский, 2019), с имплантированными термодатчиками. Судя по данным термографиков, тетрациклин был введен через 2 суток после возобновления нормотермии. Лишь у одного суслика из четырех тетрациклиновая метка была видна базальнее ИГЭ, у остальных – в базальной части ИГЭ, следовательно, окончание формирования ИГЭ пришлось уже на активный период.
Особый вопрос – почему в зоне спячки сурков и сусликов формируются узкие приросты и можно ли считать, что узкие приросты регистрируют рост в период спячки (естественно, во время кратковременных пробуждений)? Возможен ли прирост резца, пусть и очень небольшой, при отсутствии питания и, казалось бы, очевидного отсутствия стачивания резца?
Для ответа на эти вопросы необходимо до и после спячки вводить витальные маркеры растущих частей резцов. Трунова и Лобков (1997) провели такое исследование времени формирования зоны спячки не на поверхности резцов (к тому времени суточные приросты и “зона спячки” на поверхности резцов еще не были обнаружены), а в толще дентина резцов у содержавшихся в неволе крапчатых сусликов, используя тетрациклин как витальный маркер. На окрашенных гематоксилином срезах резцов у сусликов из природных популяций “зона спячки” видна как группа интенсивно окрашенных контрастных слоев дентина и – как обязательный элемент зоны – группы очень узких слоев, объединенных в светлоокрашенную полосу. Авторы вводили тетрациклин от 1 до 6 раз 13 сусликам, пережившим спячку в условиях неволи, и пришли к заключению, что непосредственно в период спячки формируются контрастные, интенсивно окрашенные слои, а светлоокрашенные узкие слои – в период пробуждения от спячки. При этом исследователи определяли не только скорость аппозиции дентина, но в ряде случаев также и скорость роста в длину (прорезания) резца за период между тетрациклиновыми метками. Из приведенных этими авторами табличных данных видно, что у одной особи, не получавшей корм, в течение 22 суток в конце августа – начале сентября было 5–6 гетеротермных периодов, скорость аппозиции дентина составила 2.5 мкм/сутки и скорость прорезания резца 29.6 мкм/сутки. Это показывало, что резец продолжал прорезаться и при отсутствии корма.
Резцы некоторых особей, использованных в тех экспериментах, сохранились, метки тетрациклина были видны, и мы смогли определить прирост резцов. Один из сусликов получил две инъекции тетрациклина в период спячки. Суслик в середине сентября погрузился в спячку с периодическими пробуждениями (детали содержания и наблюдения за ходом спячки см. Трунова, Лобков, 1997). В одно из таких пробуждений, 28 сентября он получил инъекцию тетрациклина. Для введения тетрациклина еще раз 3 декабря его разбудили, перенеся в отапливаемое светлое помещение, 4 декабря вернули на место зимовки, и спячка возобновилась. За 66 суток спячки с конца сентября до начала декабря нижний резец прорезался на 2 мм, за этот период было зафиксировано два естественных пробуждения. За последующие 44 суток до гибели в середине января резец прирос на 0.5–1 мм, пробуждений зафиксировано не было44.
Сказанное выше свидетельствует, что хотя сурки и суслики не питаются в период спячки, их резцы продолжают расти, но в крайне низком темпе. Это вполне соответствует результатам многочисленных исследований характера изменений физиологических и биохимических процессов у млекопитающих в период спячки (обзор см., например, Carey et al., 2003).
Есть существенное возражение против того, что узкие приросты – это рост во время кратковременных пробуждений, когда нет потребления корма и, казалось бы, нет стачивания резцов. Если рост резца не компенсируется стачиванием, то нарушается нормальное функционирование верхних и нижних резцов. Это часто наблюдается в неволе у грызунов, которых содержат на мягком корме, и приводит к нарушению нормального смыкания верхних и нижних резцов. У исследованных нами перезимовавших грызунов никаких внешне морфологических свидетельств нарушения нормального функционирования резцов не наблюдалось. У 10 взрослых (3 года и старше) экземпляров серого сурка с длиной резца 61–74 мм ширина зоны узких приростов варьировала от 1.8 до 3.3 мм, составляя 0.032 до 0.055 длины резца. У 10 взрослых экземпляров длиннохвостого суслика при длине резца 29.5–34.2 мм ширина зоны узких приростов варьировала от 0.550 до 1.190 мм, составляя 0.017 до 0.039 длины резца. Либо такое увеличение длины резца не приводит к существенному нарушению функционирования резцов, либо в период кратковременных пробуждений в период спячки какое-то стачивание имеет место. Косвенным свидетельством последнего являются наблюдения Корзинкиной (1935), что осенью при раскопке зимовочных нор серого сурка из них вынимали 8–9 кг сена, а весной вместо сена находили мелкую перетертую труху. Следовательно, они могли перетирать резцами подстилку во время периодических пробуждений. Данные по другим видам сурков показывают, что хотя сурки и отказываются от пищи в период спячки, даже если она им предложена, атрофии пищеварительного тракта нет, он продолжает функционировать на очень низком уровне (Hume et al., 2002; Florant et al., 2013). Можно предположить, что заглатывание ничтожного количества подстилки во время кратковременных пробуждений не оказывает существенного влияния на пищеварительный тракт.
Открытым остается вопрос, являются ли узкие приросты в “зоне спячки” регистраторами кратковременных пробуждений. Ответ может дать только исследование резцов сурков или сусликов, перезимовавших с вшитым термодатчиком, которым был бы введен витальный маркер до и после спячки. Ринальди (Rinaldi, 1999) пометила тетрациклином резцы двух особей Marmota flaviventris вскоре после их выхода из спячки и не нашла соответствия числа узких контрастных приростов числу временных пробуждений за период спячки. Такое соответствие, хотя и приблизительное, было обнаружено у двух особей Spermophilus parryii, зимовавших с имплантированным термодатчиком (Клевезаль, Ануфриев, 2013). Однако эти суслики в течение трех лет были лабораторными животными, зимовали в искусственных условиях, и у них были видны некоторые аномалии роста резцов. Поэтому вопрос о том, что регистрируют узкие приросты, по нашему мнению, остается открытым.
Иначе обстоит дело с приростами в “зоне спячки” хомяков. Хомяки известны как запасатели корма. Очень мало данных о зимнем периоде хомяка Брандта в Армении. В норе этого хомяка, раскопанной 1 сентября 1971 г. в том же месте, откуда весной была взята исследованная нами выборка, гнездовая камера и отнорки были забиты зерном, ход закрыт плотной пробкой, а хомяк был найден в отнорке за камерой и был активен. По данным Магомедова и Омарова (1995), хомяк Радде питается во время кратковременных пробуждений в период спячки, но не интенсивно, и за зимний период поедает 4–7% запасов, остальное потребляет весной. Возможно, это же справедливо и для хомяка Брандта в Армении. О том, что эти хомяки питаются в период спячки, свидетельствует и длина их “зоны спячки”. Относительно длины резца она, по меньшей мере, в два раза больше, чем у сусликов (см. табл. 4), т.е. прирост резца в длину очевиден. В записи спячки на резцах хомяков рода Mesocricetus нет ИГЭ и, следовательно, у них нагрузка на резец в период спячки меняется существенно меньше, чем у сусликов. Нагрузка меняется в период спячки не столь сильно, как у сурков и сусликов, именно потому, что хомяки могут питаться в периоды пробуждения в течение спячки. Можно полагать, что количество приростов в “зоне спячки” показывает количество таких пробуждений.
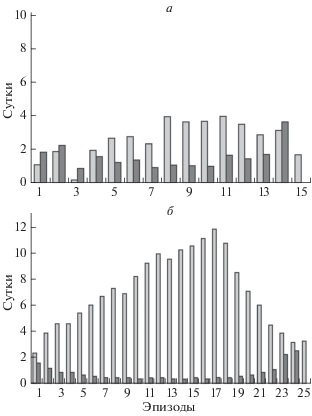
По-видимому, не только особенности питания хомяков в период спячки определяют характер записи спячки на поверхности резцов. Как было сказано выше, у обыкновенного хомяка регулярная запись спячки на резцах не была обнаружена (Клевезаль и др., 2020). Показательно сравнение динамики температуры тела в зимний период (по данным имплантированных термодатчиков) у самца обыкновенного хомяка и у самца хомяка Радде, зимовавших в одних и тех же условиях, приближенных к естественным, хотя и в разные годы (Клевезаль и др., 2020) (рис. 7). У обыкновенного хомяка продолжительность спячки была существенно меньше, длительность эпизодов нормотермии больше, а эпизодов гипотермии меньше, чем у хомяка Радде. Различалась и глубина гипотермии: до 5°С у обыкновенного хомяка и до 1°С у хомяка Радде. В отличие от хомяка Радде, на поверхности резцов этого обыкновенного хомяка вообще не было узких приростов (Клевезаль и др., 2020). Это дает основание предполагать, что на формирование узких приростов на поверхности резца могут влиять как длительность эпизодов нормотермии, так и длительность и глубина эпизодов гипотермии, которые и определяют особенности питания хомяков в это время.
ЗАКЛЮЧЕНИЕ
Рост резцов зимоспящих грызунов продолжается и в период спячки. У сурков и сусликов, не питающихся в период спячки, темп роста минимальный, нагрузка на резец до, в период и после спячки сильно меняется, в результате спячки на поверхности резца образуется “зона спячки”. У них “зона спячки” имеет два общих признака – очень узкие, по сравнению с суточными, приросты и ИГЭ – изменение границы эмали и дентина. Межвидовые различия в характере “зоны спячки” могут быть объяснены региональными климатическими условиями. У хомяков, которые могут питаться в период спячки, прирост резца за это время более существенный, нагрузка на резец меняется меньше, ИГЭ нет, и “зона спячки” отмечается только относительно узкими приростами. Межвидовые различия такой записи спячки могут быть существенными, вплоть до отсутствия записи, несмотря на наличие спячки. Эти различия, видимо, связаны не столько с климатом, сколько с характером протекания самой спячки – с продолжительностью эпизодов нормотермии и гипотермии, а также с глубиной последних.
Какую информацию об особенностях протекания спячки можно извлечь при исследовании поверхности резца? Вероятно, можно получить некоторое представление о характере климатических условий в зимний период. Зерванос с соавторами (Zervanos et al., 2010) выявили влияние климатических условий на характер спячки у зимовавших в природе сурков Marmota monax, сравнивая данные термодатчиков, имплантированных особям из разных популяций. У сурков и сусликов основным элементом записи спячки на поверхности резца является ИГЭ, степень выраженности которого широко варьирует. На нашем материале выявлена тенденция уменьшения четкости этого параметра с уменьшением сезонной контрастности климата в районе обитания выборки. Это показали сравнения четкости записи спячки на резцах в парах: сурки высокогорья Тянь-Шаня – сурки Камчатки, суслики предгорий Алтая – суслики Причерноморья. Возможно, дальнейшие исследования выявят связь характера “зоны спячки” на поверхности резцов с особенностями зимнего периода. В данное время можно только сказать, что при анализе поверхности резцов перезимовавших сурков и сусликов из районов со слабо контрастным климатом следует ожидать нечетко выраженной “зоны спячки”.
О некоторых особенностях зимнего периода сурков и сусликов может свидетельствовать наличие не одного, а двух ИГЭ на одном резце. Две “зоны спячки” на одном резце были обнаружены у сурков с западного побережья Камчатки (Клевезаль, Ануфриев, 2013а). Дополнительная (как бы несвоевременная) “зона спячка” была видна на резце одной особи Spermophilus richardsonii (Goodwin et al., 2005). Выше было сказано, что две “зоны спячки” были найдены на резце крапчатого суслика, жившего в тот период, когда, по полевым наблюдениям, спячка прерывалась периодом активности. О возможности повторного залегания в спячку серых сурков в природе писал Айзин (1979). Мало данных о характере спячки сурков этой части Камчатки, но можно предположить, что у них изменение погодных условий в зимне-весенний период в тот год нарушило нормальный ход спячки, и это отразилось на поверхности резцов. Таким образом, обнаружение двойных или дополнительных ИГЭ на резце может указывать на возможное возобновление активности в ходе зимней спячки.
Число узких приростов на участке с ИГЭ на резцах сурков и сусликов, к сожалению, нельзя считать показателем количества эпизодов нормотермии в период спячки. Во-первых, потому, что вопрос о том, что регистрируют узкие приросты, остается открытым. Во-вторых, потому, что обычно кроме узких приростов в “зоне спячки” есть участки без видимых приростов, на которых могут быть визуально не фиксируемые приросты.
Иная ситуация имеет место у хомяков. Запись спячки обнаружена у средних хомяков Mesocricetus и она существенно отличается от таковой сурков и сусликов. У средних хомяков спячка регистрируется как серия более четких и узких приростов, количество которых, судя по имеющимся данным, соответствует количеству эпизодов нормотермии за период спячки. Можно предположить, что во всех случаях, когда у особей обыкновенного хомяка на резцах видна сходная “зона спячки”, по количеству узких четких приростов на резце можно судить о количестве эпизодов нормотермии у данной особи. Однако отсутствие записи спячки на поверхности резцов обыкновенных хомяков нельзя считать свидетельством активности хомяка в течение всего зимнего периода.
По мере накопления данных исследования поверхности резцов у зимоспящих грызунов, которые зимовали в природе с имплантированными термодатчиками, объем информации наверняка будет увеличен.
Список литературы
Айзин Б.М., 1979. Грызуны и зайцеобразные Киргизии. Фрунзе: Изд-во Илим. 200 с.
Алфимов А.В., Берман Д.И., 2009. Об условиях зимовки берингийского суслика (Spermophilus parryi, Sciuridae, Rodentia) // Зоологический журнал. Т. 88. № 6. С. 731–744.
Ануфриев А.И., Ядрихинский В.Ф., 2019. Температурная регуляция процессов зимней спячки у длиннохвостого суслика Spermophilus undulatus Pallas, 1778 // Научный электронный журнал Принципы экологии. Т. 8. № 3. С. 3–12.
Бадмаев Б.Б., 2007. Длиннохвостый суслик в условиях западного Прибайкалья. Новосибирск: Наука. 107 с.
Калабухов Н.И., 1985. Спячка млекопитающих М.: Наука. 260 с.
Клевезаль Г.А., 2010. Динамика роста резцов и суточные приросты на их поверхности у трех видов мелких грызунов // Зоологический журнал. Т. 89. № 3. С. 360–370.
Клевезаль Г.А., Ануфриев А.И., 2013. О росте резцов сусликов (род Spermophilus) в период зимней спячки // Зоологический журнал. Т. 92. № 4. С. 481–489.
Клевезаль Г.А., Ануфриев А.И., 2013а. Изменчивость приростов и “зоны спячки” на поверхности резцов сурков (род Marmota) // Зоологический журнал. Т. 92. № 11. С. 1333–1348.
Клевезаль Г.А., Лобков В.А., 2008. Суточные приросты и “зона спячки” на поверхности резцов сусликов рода Spermophilus // Зоологический журнал. Т. 87. № 12. С. 495–1503.
Клевезаль Г.А., Суховская Л.И., 1995. Дентин резцов грызунов как регистрирующая структура // Зоологический журнал. Т. 74. Вып. 4. С. 932–934.
Клевезаль Г.А., Щепоткин Д.В., 2012. Скорость роста резцов грызунов и запись полного годового цикла в резцах серого сурка (Marmota baibacina centralis) // Зоологический журнал. Т. 91. № 1. С. 111–118.
Клевезаль Г.А., Щепоткин Д.В., 2017. Суточные приросты на поверхности резцов грызунов. Внутри- и межвидовая изменчивость // Зоологический журнал. Т. 96. № 7. С. 838–849.
Клевезаль Г.А., Зайцева Е.А., Щепоткин Д.В., Феоктистова Н.Ю., Чунков М.М., Суров А.В., 2020. Есть ли запись зимней спячки на поверхности резцов у обыкновенного хомяка (Cricetus cricetus, Rodentia, Cricetidae) // Зоологический журнал. Т. 99. № 1. С. 104–112.
Клевезаль Г.А., Ушакова М.В., Чунков М.М., Феоктистова Н.Ю., Суров А.В., 2012. Запись зимней спячки на поверхности резцов хомяка Радде (Mesocricetus raddei) // Зоологический журнал. Т. 91. № 6. С. 714–720.
Клевезаль Г.А., Феоктистова Н.Ю., Щепоткин Д.В., Суров А.В., 2015. Особенности записи зимней спячки на поверхности резцов хомячков рода Allocricetulus // Зоологический журнал. Т. 94. № 2. С. 259–272.
Клевезаль Г.А., Чунков М.М., Омаров К.З., Щепоткин Д.В., 2018. Запись зимней спячки на поверхности резцов хомяка Радде (Mesocricetus raddei, Rodentia, Cricetidae) из Дагестана // Зоологический журнал. Т. 97. № 5. С. 591–598.
Корзинкина Е.М., 1935. Биология и экология сурка и сурочий промысел в Кош-Агачском аймаке (Ойротия) // Экология сурка и сурочий промысел. М.-Л.: Внешторгиздат. С. 30–62.
Лобков В.А., 1999. Крапчатый суслик Северо-Западного Причерноморья: биология, функционирование популяций. Одесса: Астропринт. 272 с.
Магомедов М.-Р., Омаров К.З., 1995. Особенности питания и состояния природной популяции хомяка Радде (Mesocricetus raddei avaricus) в агроландшафтах горного Дагестана // Зоологический журнал. Т. 74. № 3. С. 123–133.
Михайлюта А.А., 1988. Структура популяции серого сурка в высокогорье Тянь-Шаня. Дис. … канд. биол. наук. М.: МГУ. 176 с.
Селькова Ю.Е., 2003. Дентин резцов зимоспящих грызунов как индикатор климатических факторов // Доклады Академии Наук. Т. 393. № 1. С. 138–141.
Стогов И.И. 1956, Материалы по питанию серого сурка (Marmota baibacina Kastsh.) // Зоологический журнал. Т. 35. Вып. 9. С. 1390–394.
Трунова Ю.Е., Клевезаль Г.А., 1999. Межвидовые различия в регистрации зимней спячки дентином резцов грызунов // Зоологический журнал. Т. 78. Вып. 12. С. 1455–1464.
Трунова Ю.Е., Лобков В.А., 1997. Регистрация особенностей спячки дентином резцов крапчатого суслика Spermophilus suslicus (Sciuridae, Rodentia) // Зоологический журнал. Т. 76. Вып. 8. С. 940–947.
Чернявский Ф.Б., 1977. Некоторые экологические особенности арктического длиннохвостого суслика (Citellus parryi Rich 1927) на крайнем северо-востоке Сибири // Механизмы зимней спячки млекопитающих. Владивосток. С. 5–17.
Batavia M., Nguyen G., Zucker I., 2013. The effects of day length, hibernation, and ambient temperature on incisor dentin in the Turkish hamster (Mesocricetus brandti) // Journal of Comparative Physiology. P. B. V. 183. P. 557–566.
Carey H.V., Andrews M.T., Martin S.L., 2003. Mammalian hibernation: Cellular and molecular responses to depressed metabolism and low temperature // Physiological Reviews. V. 83. P. 1153–1181.
Charge T.D., 2001. Hibernation biology of Richardson’s ground squirrels: Hibernaculum systems and energy utilization // Thesis Master of Science Lethbridge, Alberta, Canada. 139 p.
Chiba M., Tashiro T., Tsuruta M., Eto K., 1976. Acceleration and circadian rhythm of eruption rates in the rat incisor // Archives of Oral Biology. V. 21. P. 269–271.
Florant G.L., Fenn A.M., Healy J.E., Gregory K. Wilkerson G.K., Handa R.J., 2013. To eat or not to eat: the effect of AICAR on food intake regulation in yellow-bellied marmots (Marmota flaviventris) // The Journal of Experimental Biology. V. 213 P. 2031–2037.
Goodwin H.T., Michener G.R., Gonzalez D., Rinaldi C.E., 2005. Hibernation is recorded in lower incisors of recent and fossil ground squirrels (Spermophilus) // Journal of Mammalogy. V. 86. P. 323–332.
Goodwin H.T., Ryckman E.M., 2006. Lower incisors of prairie dogs (Cynomys) as biorecorders of hibernation and season of death // Journal of Mammalogy. V. 87. P. 1002–1012.
Hume I.D., Beiglböck C., Ruf T., Frey-Roos F., Bruns U., Arnold W., 2002. Seasonal changes in morphology and function of the gastrointestinal tract of free-living alpine marmots (Marmota marmota) // Journal of Comparative Physiology B. V. 172. P. 197–207.
Klevezal G.A., 1996. Recording structures of mammals. Determination of age and reconstruction of life history. Rotterdam: A.A. Balkema. 274 p.
Klevezal G.A., Mina M.V., 1990. Daily layers and hibernation marks in incisor dentin of Sicista pseudonapaea and some biological remarks // Acta Theriologica. V. 35. P. 345–356.
Lee T.N., Kohl F., Buck C.L., Barnes B.M., 2015. Hibernation strategies and patterns in sympatric arctic species, the Alaska marmot and the arctic ground squirrel // Journal of Mammalogy. V. 97. P. 135–144.
Michaeli Y., Weinreb M.M., 1968. Role of attrition in the physiology of the rat incisor. III. Prevention of attrition and occlusal contact in the non articulating incisor // Journal of Dental Research. V. 47. P. 633–640.
Ohtsuka M, Saekil S., Igarashil K., Shinoda H., 1998. Circadian rhythms in the incorporation and secretion of 3H-proline by odontoblasts in relation to incremental lines in rat dentin // Journal of Dental Research. V. 77. P. 1889–1895.
Ohtsuka-Isoya M., Hayashi H., Shinoda H., 2001. Effect of suprachiasmatic nucleus lesion on circadian dentin increment in rats // American Journal of Physiology. Regulatory, Integrative and Comparative Physiology. V. 280. P. R1364–R1370.
Rinaldi C., 1999. A record of hibernation in the incisor teeth of recent and fossil marmots (Marmota flaviventris) // Proc. 11th International Symposium on Dental Morphology. Oulu, Finland: Oulu University Press. P. 112–119.
Silva M.A.J., Merzel J., 2004. Alveolar bone Sharpey fibers of the rat incisor in normal and altered functional conditions examined by scanning electron microscopy // Anatomical Record. Part A. V. 279. P. 792–797.
Simmer J.P., Papagerakis P., Smith C.E, Fisher D.C., Rountrey A.N. et al., 2010. Regulation of dental enamel shape and hardness // Journal of Dental Research. V. 89. P. 1024–1038.
Smith C.E., 1998. Cellular and chemical events during enamel maturation // Critical Reviews in Oral Biology and Medicine. V. 9. P. 128–161.
Smith C.E., Warshawsky H., 1976. Movement of entire cell population during renewal of the rat incisor as shown by radiography after labeling with 3H-thymidine: The concept of a continuously differentiating cross-sectional segment // American Journal of Anatomy. V. 145. P. 225–260.
Srivastava R., 1998. Biomechanical analyses of rodent incisors: A morphological and microstructural adaptation // Mitteilungen der Bayerischen Staatssammlung für Paläontologie und Histor. Geologie. Heft 38. S. 209–225.
Steigman S., Michaeli Y., Yitzhaki M., Weinreb M., 1989. A three-dimensional evaluation of the effects of functional occlusal forces on the morphology of dental and periodontal tissues of the rat incisor // Journal of Dental Research. V. 68. P. 1269–1274.
Zaytseva E.A., Chunkov M.M., Omarov K.Z., 2020. Hibernation records on the incisor surface in the Turkish hamster (Mesocricetus brandti, Rodentia, Cricetidae) // Povolzhskiy Journal of Ecology. V. 1. P. 44–51.
Zervanos S.M., Maher C.R., Waldvogel J.A., Florant G.L., 2010. Latitudinal differences in the hibernation characteristics of woodchucks (Marmota monax) // Physiological and Biochemical Zoology. V. 83. P. 135–141.
Дополнительные материалы отсутствуют.
Инструменты
Зоологический журнал