

Зоологический журнал, 2021, T. 100, № 5, стр. 483-492
Биссусный аппарат мидии тихоокеанской (Mytilus trossulus, Bivalvia, Mytilidae) из Японского моря
Е. Е. Вехова *
Национальный научный центр морской биологии имени А.В. Жирмунского ДВО РАН
690041 Владивосток, Россия
* E-mail: evechova@gmail.com
Поступила в редакцию 18.03.2020
После доработки 25.06.2020
Принята к публикации 07.07.2020
Аннотация
У промыслового вида мидии тихоокеанской (Mytilus trossulus) из Японского моря изучали особенности морфологии биссусного аппарата, биссусных нитей и биссусной бороздки ноги. Описаны этапы процесса биссусообразования и связанные с этим процессом особенности морфологического строения биссусной бороздки у Myt. trossulus. Показано, что биссус состоит из корня, стебля и биссусных нитей, которые в поперечном сечении имеют форму эллипса. Биссусная нить состоит из рифленой проксимальной части, которая начинается сразу за манжетой и составляет 1/3 ее длины, и относительно гладкой, упругой дистальной части (2/3 длины нити), заканчивающейся на дистальном конце прикрепительным диском. На поверхности прикрепительного диска располагаются три армирующих тяжа. Наблюдаемые особенности морфологии исследуемых структур обсуждаются с точки зрения пространственного распределения митилид в прибрежной зоне моря и успешного обитания Myt. trossulus на скалистых берегах с активным воздействием волн.
Для выживания в прибойной зоне верхней сублиторали двустворчатые моллюски семейства Mytilidae на протяжении всей жизни используют способность прочно прикрепляться к субстрату посредством биссуса. Биссус возник у митилид в результате неотении (Yonge, 1962; Lee et al., 1990). Биссусное прикрепление дает митилидам ряд преимуществ перед другими двустворчатыми моллюсками с иным способом прикрепления. Наряду с прочной фиксацией, биссусное прикрепление обеспечивает более рассеянное и сглаженное воздействие волн на организм моллюсков, чем у прочно цементирующихся видов моллюсков. Помимо этого, при неблагоприятных условиях мидия может отбросить биссус и, переместившись в более пригодное местообитание, вновь прикрепиться к субстрату, образовав новый биссус с помощью желез ноги (Waite, 1983, 1997; Denny, 1987; Denny et al., 1998).
В настоящее время морфология биссусного аппарата митилид изучена методами сканирующей электронной микроскопии у многих представителей этого семейства. Среди них такие виды, как Myt. edulis, Myt. galloprovincialis, Myt. californianus, Myt. coruscus, Crenomytilus grayanus, Modiolus modiolus, Mod. metcalfei и Septifer bilocularis (Brown, 1952; Tamarin, Keller, 1972; Bairati, Vitellaro-Zuccarello, 1974; Allen et al., 1976; Banu et al., 1980; Price, 1983; Eckroat, Steel, 1993; Carrington, Gosline, 2004; Vekhova, 2019; Бергер и др., 1985; Вехова, 2007). В настоящее время методом сканирующей электронной микроскопии до сих пор остается не изученным строение биссусного аппарата, или биссуса, у мидии тихоокеанской (Mytilus trossulus Gould 1850). Многие виды рода Mytilus являются обрастателями. Они способны прочно прикрепляться к различным природным и антропогенным субстратам посредством биссуса. Исследование процесса повторного прикрепления некоторых видов митилид к разным субстратам показало, что наиболее успешно среди изученных видов митилид, таких как Myt. coruscus, Myt. trossulus, C. grayanus, Mod. modiolus, антропогенные субстраты (бетон, дерево и железо) колонизируются Myt. trossulus (Vekhova, 2006). Несмотря на то, что в настоящее время в научной литературе есть много информации о морфологической структуре биссусного аппарата различных видов митилид (Pujol et al., 1970; Price, 1983; Eckroat, Steel, 1993; Бергер и др., 1985), данные о структуре биссуса и его формировании у мидии Myt. trossulus вовсе отсутствуют в литературе.
Myt. trossulus широко представлен в донных сообществах Японского моря и играет важную роль в морских экосистемах. Этот вид является важным промысловым объектом и объектом марикультуры в Приморском крае России (Скарлато, 1981). Необходимо отметить, что Myt. trossulus как элемент обрастания пирсов, оффшорных сооружений и подводных частей кораблей наносит значительный экономический ущерб.
Цель работы – провести детальное исследование морфологии биссуса и строения биссусной бороздки ноги, а также изучить поведение при повторном прикреплении к субстрату у мидии тихоокеанской из залива Восток Японского моря.
МАТЕРИАЛ И МЕТОДИКА
Для изучения особенности морфологии биссусного аппарата, биссусных нитей и биссусной бороздки ноги использовали мидий, собранных в 2018–2020 гг. с помощью водолазной службы Национального научного центра морской биологии им. А.В. Жирмунского ДВО РАН на глубине 1‒1.5 м в заливе Восток Японского моря (42°53′03.35″ с.ш., 132°43′41.72″ в.д.) с крупных валунов и скал. 30 экз. мидии тихоокеанской содержали в лабораторных условиях в аквариуме с проточной морской водой с нормальной соленостью 33‰ и наблюдали за их поведением при прикреплении к субстрату. С помощью штангенциркуля с точностью до ± 0.1 мм измеряли длину раковины (L, мм). У 30 экз. мидий с длиной раковины 45–50 мм изучали морфологические особенности биссусных нитей и биссуса методами световой микроскопии (СМ). У каждой особи случайным образом брали по 30 неповрежденных нитей из разных частей биссусного стебля и измеряли исследуемые параметры. Под бинокуляром при окулярах ×8 и тубусе ×7 оценивали длину нитей (l, мм), ширину нитей в проксимальной (q, мкм) и дистальной частях (t, мкм), в местах перехода проксимального участка нити в дистальный (v, мкм) и дистального участка нити в диск (s, мкм), а также долю проксимального участка нити от всей ее длины (Dp, %), измеряли диаметр прикрепительного диска вдоль наибольшей и наименьшей осей, среднее арифметическое этих измерений внесено в табл. 1 как параметр (В, мм). Кроме этого, у каждой особи считали количество функционально активных и инертных (без диска) нитей.
Особенности внешнего строения биссусных нитей и биссусной бороздки ноги изучали методами сканирующей электронной микроскопии (СЭМ). Для этого использовали особей мидии тихоокеанской одного возраста с длиной раковины 46 ± 4 мм. Индивидуальный возраст моллюсков определяли по кольцам роста на наружной поверхности раковины (Золотарев, 1989). Для СЭМ образцы биссусных нитей и ноги фиксировали в 2.5% растворе глютаральдегида, приготовленном на 0.1 М какодилатном буфере (pH 7.4) в течение 2–4 ч при температуре 4°С. После этого образцы промывали в 0.1 М какодилатном буфере в течение 15–20 мин. Далее образцы обезвоживали в спиртах возрастающей концентрацией, постепенно доводя до чистого ацетона (Миронов и др., 1994). После этого образцы окончательно высушивали в диоксиде углерода по методике сушки в критической точке, используя прибор critical point dryer 030 фирмы “BAL-TEC”, помещали на поверхность алюминиевых столиков и напыляли хромом, используя вакуумный прибор для покрытия тонких мембран Q 150T ES. После чего особенности морфологии полученных образцов биссусных нитей и ноги мидии Myt. trossulus исследовали на сканирующем электронном микроскопе Carl Zeiss, Sigma 300 VP. На полученных изображениях делали различные промеры с помощью программы SmartTiff, которая входит в приложение для пользователей СЭМ Carl Zeiss, Sigma 300 VP.
Все полученные изображения редактировали с использованием графических программ Adobe Photoshop CS6, Corel Draw Graphics Suite X6 и Corel Photo Paint X6. Статистическую обработку данных выполняли на ПК по стандартным алгоритмам, реализованным в ППП Microsoft Excel.
РЕЗУЛЬТАТЫ
Общий план организации биссусного аппарата у мидии тихоокеанской
Результаты исследований показали, что у Myt. trossulus биссус состоит из корня, стебля и биссусных нитей, которые в поперечном сечении имеют форму эллипса и которые на дистальном конце оканчиваются прикрепительными дисками (рис. 1). Корень биссуса глубоко внедрен в проксимальную часть ноги (рис. 2). Cтебель длиной 3 мм распространяется в вентральном направлении от корня и выходит из отверстия биссусной железы, расположенного в проксимальной части биссусной борозды; из раковины биссус выходит наружу через биссусное зияние вентрального края раковины. От стебля отходят биссусные нити в двух направлениях ‒ к макушке и к задней части раковины мидии. При детальном рассмотрении можно видеть, что стебель имеет ламеллярную структуру и окружен со всех сторон манжетами, которые дают начало биссусным нитям (рис. 3а). В центральной части пластинки стебля плотно упакованы в округлый сердечник и расположены под углом 90° относительно биссусной бороздки ноги. Стебель чаще всего несет лишь функционально активные нити, которые непосредственно участвуют в прикреплении моллюска (рис. 1, 3а). Оценка соотношения функционально активных и инертных нитей на стебле у особей разного размера показала, что более 90% нитей вне зависимости от размера моллюска участвуют в прикреплении моллюска к субстрату.
Рис. 1.
Внешний вид биссусного аппарата у мидии тихоокеанской (Mytilus trossulus). Длина раковины моллюска 48 мм. Шкала 1 см.
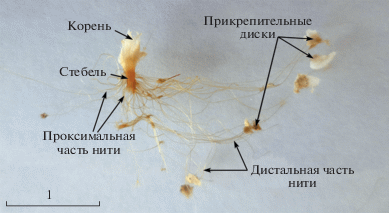
Рис. 2.
Схема продольного сечения раковины, строения биссуса мидии тихоокеанской (Mytilus trossulus) и мышечной системы, участвующей в прикреплении мидии.
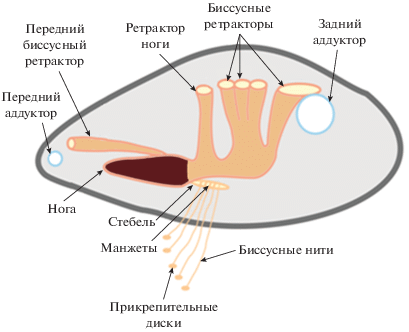
Рис. 3.
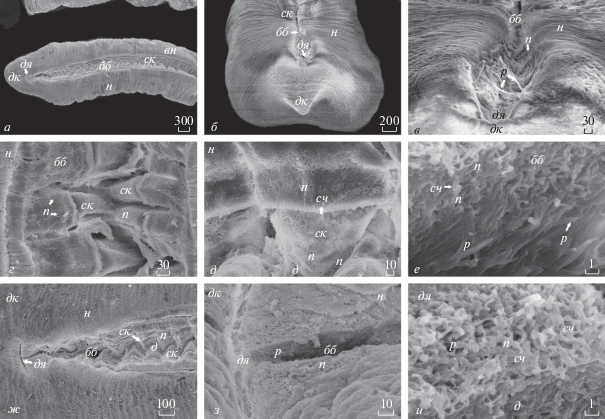
Морфология биссусной нити мидии тихоокеанской (Mytilus trossulus) (СЭМ): а – участок стебля с охватывающими его манжетами, к которым прикрепляются биссусные нити; б – наружная поверхность прикрепительного диска; в – переходный участок диска в дистальный участок биссусной нити; г – дистальный участок биссусной нити; д – переход дистального участка в проксимальный участок биссусной нити; е – проксимальный участок биссусной нити.

Корень светло-желтого цвета, стебель окрашен в светло-коричневый цвет, а биссусные нити, как правило, в проксимальной части окрашены темнее, чем в дистальной, а в дистальной части ‒ светло-бежевого цвета, при этом прикрепительные диски окрашены в темно-бежевый цвет (рис. 1). Вновь образованные биссусные нити имеют светло-желтоватый тон (рис. 4 г).
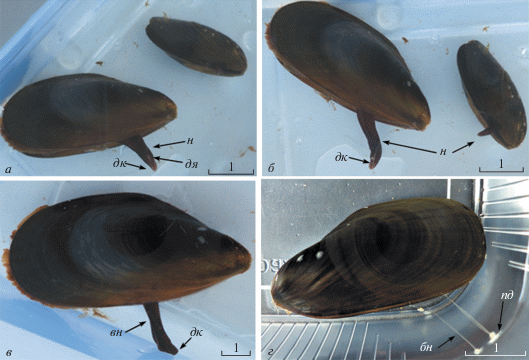
Рис. 4.
Стадии процесса биссусообразования у мидии тихоокеанской (Mytilus trossulus): а – открепившаяся мидия переползает с помощью ноги на новое место; б – с помощью дистальной ямки на кончике ноги мидия ощупывает поверхность субстрата; в – перед прикреплением мидия прижимает дистальный конец ноги к субстрату и удлиняет ее в 3 раза; г ‒ процесс формирования биссусных нитей оканчивается прикреплением мидии к субстрату и втягиванием ноги. Шкала 1 см.
Биссусный аппарат располагается в ноге моллюска. Нога обычно небольшого размера, но благодаря развитой мускулатуре может значительно менять свои размеры; в форме языка, адаптирована для перемещения по твердому субстрату; как правило, хорошо пигментирована и имеет бордовый цвет (рис. 2). Нога имеет хорошо развитую систему мышц-ретракторов, прикрепленных к стенке раковины; при сокращении ретракторов тело моллюска подтягивается к субстрату. Myt. trossulus имеет 6 пар пучков мышц-ретракторов: пара передних ретракторов биссуса, пара ретракторов ноги и четыре пары задних ретракторов биссуса. В процессе образования биссусной нити непосредственно участвует биссусная бороздка, расположенная на вентральной стороне ноги. На дистальном конце ноги с вентральной стороны располагается дистальная ямка, дно и стенки которой покрыты ресничками и булавовидными сосочками (рис. 5ж, 5и).
Рис. 5.
Морфологическое строение биссусной бороздки ноги у мидии тихоокеанской (Mytilus trossulus) (СЭМ): а – внешний вид сверху вентральной поверхности ноги и биссусной бороздки; б – фронтальный вид дистального конца ноги; в – фронтальный вид увеличенного дистального конца ноги и дистальной ямки; г – увеличенный внешний вид сверху биссусной бороздки ноги; д – увеличенный внешний вид боковой стенки биссусной бороздки ноги; е – увеличенный внешний вид ресничек, булавовидных сосочков и пор на дне боковой поверхности стенки биссусной бороздки ноги; ж – внешний вид дистальной ямки на вентральной поверхности ноги с участком биссусной бороздки ноги; з – увеличенный внешний вид дистальной ямки на дистальном конце ноги; и – увеличенный внешний вид дна дистальной ямки ноги.
Mорфология биссусных нитей у мидии тихоокеанской
У исследованного вида митилид в пределах каждой биссусной нити условно можно выделить рифленую проксимальную часть, которая начинается сразу за манжетой и составляет 1/3 ее длины, и относительно гладкую, упругую дистальную часть (2/3 длины нити), заканчивающуюся на дистальном конце прикрепительным диском (рис. 1, 3, 4г). Как правило, ширина нити в проксимальной части в два раза больше, чем в дистальной части нити. Даже невооруженным глазом заметно, что ширина и характер поверхности биссусных нитей различаются на разных участках (рис. 1). Мы предприняли детальное исследование биссусных нитей в местах перехода дистальной части нити в прикрепительный диск (рис. 3в) и перехода проксимальной части нити в дистальную (рис. 3д), обозначив их соответственно зона 1 и зона 2 и (з1 и з2). Ширина перехода биссусной нити в прикрепительный диск у этого вида варьирует и составляет 38–86 мкм. Ширина перехода проксимальной части биссусной нити в дистальную часть варьирует и составляет 49‒63 мкм.
Проксимальная часть нити. Поверхность проксимального участка нити покрыта многочисленными поперечными складками и мелкими бороздами, ориентированными почти перпендикулярно продольной оси нити (рис. 3е). Эти складки делают проксимальную часть нити растяжимой. Длина проксимального участка нити варьирует у этого вида и составляет 4.4‒8.4% от общей длины биссусной нити. Ширина проксимального участка варьирует вдоль нити у каждой особи и различается в пределах вида (рис. 3е). Наибольшая ширина нити в проксимальном участке у Myt. trossulus составляет 109 мкм, а наименьшая – 54 мкм. Одна из сторон проксимального участка нити имеет помимо основных мелких поперечных складок складчатую поверхность в виде гофрированной каймы (рис. 3е). Кайма представляет собой близко расположенные одна к другой складочки более мелкого порядка и с одной стороны нити может вовсе отсутствовать у некоторых экземпляров (рис. 3е). У Myt. trossulus наиболее широкая гофрированная кайма, которая составляет в среднем 21–28 мкм (рис. 3е), находится в центральной части проксимального участка биссусной нити. Степень компактности складок изменяется вдоль проксимального участка нити. Ширина основных поперечных складок в центральной части проксимального участка нити варьирует в среднем от 0.7 до 4 мкм (рис. 3е). Например, на 20 мкм длины проксимального участка нити приходится соответственно 12 поперечных складок.
Дистальная часть нити. Морфология места перехода проксимального участка нити в дистальный участок сочетает в себе их структурные черты (рис. 3д). Ширина нити здесь варьирует в диапазоне 38–51 мкм. Дистальный участок нити начинается после переходной зоны з2 (рис. 3д) и заканчивается участком перехода дистальной части в прикрепительный диск з1, где биссусная нить постепенно становится шире (рис. 3в). В поперечном сечении биссусная нить сплющенная, имеет форму эллипса. Поверхность нити в дистальной части не складчатая, а испещрена мелкими бороздами (рис. 3в, 3г) и упругая (рис. 3г). Вдоль поверхности тянется хорошо заметный, плотный округлый тяж, занимающий латеральное положение на одной из сторон нити (рис. 3г). Участок нити с более мощным продольным тяжом и глубокими выраженными бороздками находится в центральной части дистального участка нити. Дистальная часть нити, как правило, в два раза ýже проксимального участка нити. Кроме того, ширина дистальной части нити варьирует у каждой особи: в центральной области нить самая широкая и ее ширина здесь составляет 51 мкм, а наиболее тонкая нить в области переходной зоны с проксимальной частью – 31 мкм.
Нить в зоне перехода дистального участка в прикрепительный диск (з2) слегка уплощенная, с бороздами и выраженными армирующими тяжами на поверхности (рис. 3б, 3в); ширина нити здесь 86 мкм.
Прикрепительный диск. На дистальном конце нить заканчивается прикрепительным диском, хорошо заметным невооруженным глазом (рис. 1, 3б). Он имеет вид уплощенной пластинки в форме эллипса. У Myt. trossulus продольная ось нити располагается под углом к плоскости диска (рис. 3б). Нижняя поверхность дисков повторяет контур субстрата и по внешней структуре напоминает застывшую пену.
На поверхности диска различимы округлые тяжи, которых обычно бывает от одного до трех (рис. 3б). Один мощный тяж распространяется по поверхности диска до половины длины прикрепительного диска, ориентирован вперед к макушкам раковины моллюска (рис. 3б). Наряду с ним, обычно различаются два боковых, менее выраженных тонких, коротких тяжа.
Процесс биссусообразования у мидии тихоокеанской
Лабораторные наблюдения в аквариумах за не прикрепленными к субстрату мидиями Myt. trossulus позволили отследить процессы движения и прикрепления. Мидия переползает на новое место с помощью ноги, при этом вытянутая нога высовывается из створок раковины (рис. 4а). Измерения показали, что длина ноги у мидии длиной раковины 45‒50 мм на начальном моменте биссусообразования составляет 1 см. Мидия, совершая зигзагообразные движения ногой, ощупывает поверхность субстрата дистальным кончиком ноги, где располагается дистальная ямка биссусной борозды (рис. 4б). Затем мидия прижимает к поверхности субстрата дистальную ямку ноги, при этом нога удлиняется в 3 и более раз по сравнению с длиной ноги в состоянии покоя (рис. 4в). Этот процесс занимает от 1 до 5 мин. По-видимому, это связано с процессом формирования биссусной нити. Когда биссусная нить полностью сформирована в биссусной бороздке, нога сокращается, а на субстрате остается прикрепленный диск, от которого тянется биссусная нить (рис. 4г). Процесс образования пучка биссусных нитей может занять один месяц. Myt. trossulus может образовывать новый биссус и биссусные нити на протяжении всей жизни.
Морфология биссусной бороздки ноги у мидии тихоокеанской
Биссусные нити формируются в биссусной бороздке, расположенной вдоль центральной оси практически по всей длине вентральной стороны ноги мидии (рис. 5а, 5б). На дистальном конце ноги располагается узкая дистальная ямка (поперечная ширина 180 мкм) (рис. 5в, 5ж–5и). В этом месте нога более узкая. Биссусная бороздка ноги также сужается на дистальном конце (рис. 5ж). Ширина биссусной бороздки составляет 286‒ 300 мкм (рис. 5а, 5в, 5г). Глубина биссусной бороздки составляет 70‒250 мкм (рис. 5г, 5д). Стенки биссусной бороздки имеют складчатую поверхность и покрыты палочковидными ресничками и булавовидными выростами в виде сосочков (рис. 5г–5е). Ширина складок варьирует от 100 до 200 мкм. Поверхность складок и дна биссусной бороздки ноги покрыта многочисленными порами (рис. 5е), представляющими собой протоки желез. Поверхность дна биссусной бороздки ноги мидии несет тончайшие булавовидные сосочки (их длина 3.4 мкм) и реснички (их длина 1.5–2 мкм).
ОБСУЖДЕНИЕ
Результаты исследования и анализ литературных данных свидетельствуют, что характерные участки биссуса (рис. 1), биссусных нитей (рис. 3) и их морфология сходны у Myt. trossulus и у исследованных видов митилид Японского моря ‒ Myt. coruscus, C. grayanus, Mod. modiolus (Vekhova, 2019; Вехова, 2007). По нашим данным, размер прикрепительного диска у Myt. trossulus в среднем составляет 0.71‒0.79 мм, у Myt. coruscus ‒ 1.2, у C. grayanus ‒ 0.7, а у Mod. modiolus ‒ 0.9 мм. Сравнительный анализ свидетельствует, что у Myt. trossulus и C. grayanus эти различия носят случайный характер. Попарное сравнение свидетельствует о статистически значимых различиях в размере диска у остальных видов митилид. По длине биссусных нитей исследованные виды также различаются. Самые длинные биссусные нити имеют Myt. trossulus (23‒31 мм) и Mod. modiolus (31 мм), а самые короткие – Myt. coruscus (22 мм) и C. grayanus (23 мм). Результаты попарного сравнения показали, что первый вид статистически значимо различается длиной нитей только с M. coruscus. Результаты исследования тонкого строения биссусных нитей у этих видов митилид позволили выявить существенные различия в размере их отдельных участков. Согласно этим данным ширина биссусной нити варьирует вдоль ее длины у этих видов митилид. У Myt. trossulus ширина дистальной части нити в центральной области составляет 31‒51 мкм, у Myt. coruscus – 120‒150, у C. grayanus – 40‒70, у Mod. modiolus ‒ 20‒55 мкм. Статистически значимые различия в ширине дистальной части нити наблюдаются между Myt. coruscus и остальными видами, а также между C. grayanus и Mod. modiolus. Различия в ширине дистальной части нити между Myt. trossulus и Mod. modiolus носят случайный характер. Ширина нити в проксимальном участке составляет у Myt. trossulus 54‒109 мкм, у Myt. coruscus – 220‒250, у C. grayanus – 100‒135, у Mod. modiolus ‒ 50‒110 мкм. Попарное сравнение показало, что между Myt. trossulus, C. grayanus и Mod. modiolus различия в ширине проксимального участка носят случайный характер. Статистически значимые различия в ширине проксимального участка обнаруживаются при сравнении Myt. coruscus со всеми видами. Наиболее широкая гофрированная кайма в центральной части проксимального участка нити у Myt. trossulus составляет в среднем 21–28 мкм (рис. 3е), у Myt. coruscus – 9‒30, у C. grayanus – 5‒16, у Mod. modiolus – 2‒5 мкм. Таким образом, результаты исследования свидетельствуют о разной степени армированности биссусных нитей и разной степени ее выраженности у разных видов. Подобные различия наблюдаются и в ширине проксимальных складок у исследованных видов митилид. У Myt. trossulus ширина основных поперечных складок в центральной части проксимального участка нити составляет в среднем 0.7‒4 мкм (рис. 3е), у Myt. coruscus – 10‒34, у C. grayanus – 3‒13, у Mod. modiolus ‒ 2‒8 мкм. Самые широкие проксимальные складки наблюдаются у Myt. coruscus, самые узкие ‒ у Myt. trossulus. Различие этого параметра нити между C. grayanus и Mod. modiolus носят случайный характер.
Отмечено (Bell, Gosline, 1996; Carrington, Gosline, 2004), что различия в биомеханических свойствах дистального и проксимального участков биссусной нити определяются различиями соответствующих участков коллагеновых волокон на молекулярном уровне. Различают упругий коллагеновый домен, который входит в состав дистального участка нити и имеет высокий предел прочности на разрыв (Bell, Gosline, 1996), а также растяжимый коллагеновый домен, который входит в состав проксимального участка биссусной нити и способен к растяжению в 15 и более раз. Благодаря этому биссусная нить может удлиняться на 120–140% от ее оригинальной величины (Allen et al., 1976; Waite, 1983). Биомеханические свойства биссусных нитей отражены в морфологических различиях структуры дистального и проксимального участков нити у митилид (Waite, 1983; Qin, Waite, 1995; Waite, 1997). Так, наиболее низкой прочностью характеризуется рыхлая, растяжимая проксимальная часть, где при создании механической нагрузки в 90‒92% случаев обычно происходит разрыв нити. Это свойство присуще многим митилидам (Allen et al., 1976; Smeathers, Vincent, 1979; Eckroat, Steel, 1993; Bell, Gosline, 1996). Сравнительный анализ литературных данных показал, что характерная для мидии Myt. trossulus морфология строения биссуса и его частей (рис. 1, 3) были отмечены также у других представителей семейства Mytilidae: Myt. galloprovincialis, Myt. edulis, Myt. californianus, Mod. metcalfei и др. (Brown, 1952; Tamarin, Keller, 1972; Bairati, Vitellaro-Zuccarello, 1974; Allen et al., 1976; Banu et al., 1980; Price, 1983; Gosling, 1992; Eckroat, Steel, 1993; Bell, Gosline, 1996; Carrington, Gosline, 2004; Бергер и др., 1985).
В целом, строение биссусной нити наглядно отражает комплекс морфологических адаптаций, направленных на улучшение физической стабильности двустворчатого моллюска Myt. trossulus на поверхности субстрата. Эллипсоидная пластинка прикрепительного диска (рис. 3б) прочно фиксирует нить к поверхности субстрата, в то время как другой конец нити прочно связывается с моллюском прикрепительной манжетой (рис. 3а), в виде обруча плотно охватывающей стебель биссуса со всех сторон. Между двумя этими “крепежами” находится участок нити, выполняющий функцию швартового каната, на дистальной части прочного и упругого (рис. 3г), а в проксимальной части чрезвычайно эластичного и растяжимого в продольном направлении (рис. 3е). Такое строение биссусной нити позволяет максимально поглощать механическое напряжение (Qin, Waite, 1995; Bell, Gosline, 1996; Gosline et al., 2002; Carrington, Gosline, 2004), вызванное воздействием волн, течений и других сил внешней среды (Denny, 1987; Denny et al., 1998). По-видимому, проксимальный участок амортизирует механические нагрузки (Bell, Gosline, 1996; Gosline et al., 2002; Carrington, Gosline, 2004), связанные с волновой активностью (Denny, 1987; Denny et al., 1998). Мощные, хорошо развитые ретракторы ноги позволяют натягивать биссусные нити, в результате чего моллюск плотнее прижимается к субстрату (рис. 2).
По данным Скарлато и др. (1967), в Японском море мидия тихоокеанская не образует промысловых скоплений в прибрежной зоне моря. Вместе с тем, этот вид, как и другой промысловый вид Myt. edulis, с успехом культивируется на искусственных субстратах (Gosling, 1992; Примаков и др., 2006). В природных условиях Myt. trossulus предпочитает селиться на скальных грунтах в прибойной зоне моря, как правило, на валунах на глубине 0.5–20 м (Скарлато, 1981). В связи с этим биссусные нити формируются на стебле биссуса мидии тихоокеанской в двух направлениях: вперед к макушке раковины и к заднему концу раковины, чтобы смягчить отраженное волновое воздействие, постоянно оказываемое на раковину (Denny, 1987; Denny et al., 1998). По-видимому, расположение переднего и боковых армирующих тяжей на прикрепительном диске и степень их выраженности (рис. 3б) соответствуют направлению волнового воздействия на прикрепленную к субстрату мидию тихоокеанскую, которая испытывает фронтальное, отраженное и более или менее выраженные боковые воздействия волн (Denny, 1987; Denny et al., 1998).
Результаты исследования показали, что при внешнем сходстве морфологии биссуса и его частей у митилид разных видов размеры биссуса и биссусных нитей существенно различаются степенью армированности дисков и размером отдельных частей биссуса (Vekhova, 2019). Как и у других видов митилид, у Myt. trossulus формирование биссуса тесно связано с процессами, протекающими в биссусной бороздке (рис. 5а, 5б) (Carter et al., 2012; Скарлато, 1960) и дистальной ямке ноги (рис. 5ж, 5з) в момент формирования нити и ее прикрепления к поверхности субстрата (Pujol et al., 1970; Price, 1983; Waite, 1983). Эти различия включают морфологические особенности биссусной бороздки ноги, ее размер и топографию поверхности (рис. 5). По-видимому, не последнюю роль здесь играют формы складок биссусной бороздки вдоль поверхности ноги и форма дистальной ямки, которые существенно различаются у разных видов митилид (рис. 5г‒5и) (Eckroat, Steel, 1993). Известно также, что в процессе формирования биссусной нити важную роль играют количество секрета, вырабатываемого каждой железой ноги, и мгновенно попадание его через поры и протоки в биссусную бороздку ноги (рис. 5а), быстрота его смешивания в биссусной бороздке ноги посредством палочковидных ресничек и булавовидных сосочков, выстилающих дно и стенки этой бороздки ноги (рис. 5е), а также постепенное задубливание фенолами и последующее воздействие сокращений мощной мускулатуры ноги мидии (Bairati, Vitellaro-Zuccarello, 1974; Allen et al., 1976; Price, 1983).
Согласно литературным данным в ноге Myt. edulis локализовано 5 желез, участвующих в процессе биссусообразования (Pujol et al., 1970; Allen et al., 1976; Price, 1983; Waite, 1983). Можно предположить, что они участвуют в этом процессе и у Myt. trossulus. На дистальном конце ноги выше дистальной ямки находятся мукоциты, выстилающие поверхность ноги и вырабатывающие слизь, которая выходит на поверхность через поры при фиксации прикрепительного диска. Это мукоидная железа, которая секретирует мукополисахариды. Дистальную ямку окружает фенольная (или пурпурная) железа. Она секретирует небольшие глобулы, которые затем постепенно выводятся через протоки и примешиваются в дистальной части биссусной борозды к продукту секреции белой железы. Гранулы фенольной железы содержат полифенольный протеин и орто-дифеноламин, представленный 3,4-дигидроксифенолаланином (ДОФА), который участвует в процессе задубливания белка. От фенольной железы к основанию ноги, примыкая к биссусной бороздке и дистальной ямке, располагается энзимная (дополнительная) железа. Секреторные клетки этой фенольной железы содержат гранулы и большое количество орто-дифенолов. Они выделяются в биссусную бороздку и проявляют там фенолоксидазную активность. Как и в клетках фенольной железы, ортодифенольный компонент представлен протеином, содержащим α-ДОФА. Этот белок является предшественником β-коллагена, который используется при формировании наружной кутикулы каждой отдельной биссусной нити, прикрепительных дисков и манжет стебля. Белая (или коллагеновая) железа образована клетками, синтезирующими коллаген. Она самая большая по размеру и простирается от дистальной ямки до корня биссуса. Ее клетки содержат секреторные гранулы, которые проходят по нитчатым отросткам секреторных клеток прямо к дистальной ямке. Протоки имеют такой вид, чтобы выкачивать продукты секреции желез в биссусную бороздку и дистальную ямку. В проксимальной части ноги, в ее основании расположена биссусная железа, клетки которой синтезируют коллагеновые гранулы.
Таким образом, биссусные нити образуются в биссусной бороздке ноги по принципу отливки в фильерах. Эпителий, выстилающий каждую внутреннюю стенку фильера, образует небольшое количество складок ‒ железистые островки. Они разделены мышечными пучками, которые играют важную роль в поддержке железы. Этот способ формирования нити заключается в выделении полимера через фильеры в виде тонких непрерывных нитей, которые затем вытягиваются в воздухе (Pujol et al., 1970).
Буквенные обозначения на рис. 3, 4 и 5: бб – биссусная бороздка ноги, бм ‒ борозды мелкого порядка, бн – биссусные нити, бт – боковой тяж, вн – вентральная поверхность ноги, гк – гофрированная кайма, д ‒ дно биссусной бороздки, дк – дистальный конец, дя – дистальная ямка, з1 – переходная зона из диска в дистальный участок нити, з2 – переходная зона из дистального в проксимальный участок нити, м – манжеты, н – нога мидии, ос – оппозитная сторона нити, п ‒ поры в бороздке, пб – поперечная бороздка, пд – прикрепительный диск, пк – проксимальный конец, плд – пластинка диска, прт ‒ продольный тяж, пт – передний тяж, пск – поперечная складка, р ‒ реснички в биссусной бороздке, с – стебель биссуса, ск – складки биссусной борозды, сч ‒ булавовидный сосочек биссусной бороздки ноги.
Список литературы
Бергер В.Я., Летунов В.Н., Вшевцов Г.В., Саранчова О.Л., 1985. Морфофункциональные и экологические аспекты биссусообразования у мидии (Mytilus edulis L.) // Экология обрастания в Белом море. Л.: ЗИН АН СССР. С. 67‒75.
Вехова Е.Е., 2007. Сравнительная морфология биссусных нитей трех представителей семейства Mytilidae (Bivalvia) из Японского моря // Зоологический журнал. Т. 86. № 2. С. 154‒162.
Золотарев В.Н., 1989. Склерохронология морских двустворчатых моллюсков. Киев: Наукова думка. 112 с.
Миронов А.А., Комиссарчик Я.Ю., Миронов В.А., 1994. Методы электронной микроскопии в биологии и медицине: методическое руководство. СПб.: Наука. 400 с.
Примаков И.М., Лезин П.А., Иванов М.В., Кулаковский., 2006. Пути оптимизации марикультуры мидий в Белом море. М.: Товарищество научных изданий КМК. 72 с.
Скарлато О.А., 1960. Двустворчатые моллюски дальневосточных морей СССР (Dysodonta). Определители по фауне СССР, издаваемые Зоологическим институтом Академии наук СССР. Т. 71. 150 с.
Скарлато O.A., 1981. Двустворчатые моллюски умеренных широт западной части Тихого океана. Л.: Наука. 479 с.
Скарлато О.А., Голиков А.Н., Василенко С.В. и др., 1967. Состав, структура и распределение донных биоценозов в прибрежных водах залива Посьет (Японское море) // Исследования фауны морей. Т. 5. № 13. P. 5‒61.
Allen J.A., Cook M., Jackson D.J. et al., 1976. Observations on the rate of production and mechanical properties of the byssus threads of Mytilus edulis L. // Journal of Molluscan Studies. V. 42. № 2. P. 279‒289.
Brown C.H., 1952. Some structural proteins of Mytilus edulis // The Quarterly Journal of Microscopical Science. V. 93. P. 487‒502.
Bairati A., Vitellaro-Zuccarello L., 1974. The ultrastructure of the byssal apparatus of Mytilus galloprovincialis. II. Observations by microdissection and scanning electron microscopy // Marine Biology. V. 28. P. 145‒158.
Banu A., Shymasundari K., Hanumantha Rao K., 1980. The organization and chemistry of the byssus of some bivalves of the Waltair coast, India // The Veliger. V. 23. № 1. P. 77‒82.
Bell E.C., Gosline J.M., 1996. Mechanical design of mussel byssus material yield enhances attachment strength // The Journal of Experimental Biology. V. 199. № 4. P. 1005‒1017.
Carrington E., Gosline J.M., 2004. Mechanical design of mussel byssus: Load cycle and strain rate dependence // American Malacological Bulletin. V. 18. P. 135‒142.
Carter J.G., Harries P.J., Malchus N., Sartori A.F., Anderson L.C., Bieler R., Bogan A.E., Coan E.V., Cope J.C.W., Cragg S.M., Cartἰa-March J.R., Hylleberg J., Kelley P., Kleemann K., Kříž J., McRoberts C., Mikkelsen P.M., Pojeta J., Jr., Tëmkin I., Yancey T., Zieritz A., 2012. Illustrated Glossary of the Bivalvia. Treatise online № 48. V. 1. P. 15.
Denny M.W., 1987. Lift as a mechanism of patch initiation in mussel beds // Journal of Experimental Marine Biology and Ecology. V. 113. P. 231‒245.
Denny M., Gaylord B., Helmuth B., Daniel T., 1998. The menace of momentum: Dynamic forces of flexible organisms // Limnology and Oceanography. V. 43. № 5. P. 955‒968.
Eckroat L.R., Steel L.M., 1993. Comparative morphology of the byssi of Dreissena polymorpha and Mytilus edulis // American Malacological Bulletin. V. 10. P. 103‒108.
Gosline J., Lillie M., Carrington E., Guerette P., Ortlepp C., Savage K., 2002. Elastic proteins: biological roles and mechanical properties // Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences. V. 357. № 1418. P. 121‒132.
Gosling E.M., 1992. The mussel Mytilus: ecology, physiology, genetics and culture. Amsterdam: New York: Elsevier. Developments in aquaculture and fisheries science. V. 25. 590 p.
Lee C.Y., Lim S.S.L., Owen M.D., 1990. The rate and strength of byssal reattachment by blue mussels (Mytilus edulis L.) // Canadian Journal of Zoology. V. 68. P. 2005‒2009.
Price H.A., 1983. Structure and formation of the byssus complex in Mytilus (Mollusca, Bivalvia) // Journal of Molluscan Studies. V. 49. № 1. P. 9−17.
Pujol J.P., Rolland M., Lasry S., Vinet S., 1970. Comparative study of the amino acid composition of the byssus in some common bivalve mollusks // Comparative Biochemistry and Physiology. V. 34. P. 193‒201.
Qin X., Waite J.H., 1995. Exotic collagen gradients in the byssus of mussel M. edulis // Journal of Experimental Biology. V. 198. P. 633–644.
Smeathers J.E., Vincent J.F.V., 1979. Mechanical properties of mussel byssus threads // Journal of Molluscan Studies. V. 45. P. 219‒230.
Tamarin A., Keller P.J., 1972. An ultrastructural study of the byssal thread forming system in Mytilus // Journal of Ultrastructure Research. V. 40. P. 401–416.
Vekhova E.E., 2006. Reattachment of certain species of mytilid bivalves to various substrates // Russian Journal of Marine Biology. V. 32. № 5. P. 308‒311.
Vekhova E.E., 2019. The adaptive morphology of byssus in Mytilus coruscus, Crenomytilus grayanus, and Modiolus modiolus (Mytilidae, Bivalvia) from the Sea of Japan // Biology Bulletin. V. 46. № 9. P. 1030–1044.
Waite J., 1983. Adhesion in byssally attached bivalves // Biological Reviews of the Cambridge Philosophical Society. V. 58. № 2. P. 209–231.
Waite J.H., 1997. Marine bioadhesion: unraveling the chemistry // Journal of The Adhesion Society of Japan. V. 33. № 5. P. 186–194.
Yonge C.M., 1962. On the primitive significance of the byssus in the Bivalvia and its effects in evolution // Journal of the Marine Biological Association of the United Kingdom. V. 42. № 1. P. 113–125.
Дополнительные материалы отсутствуют.
Инструменты
Зоологический журнал