

Зоологический журнал, 2021, T. 100, № 6, стр. 640-651
Насекомые-фитофаги дуба черешчатого (Quercus robur) на северо-востоке его ареала
И. В. Ермолаев a, *, В. И. Пономарев b, А. А. Васильев c, М. С. Кумаева c
a Тобольская комплексная научная станция УрО РАН
626152 Тобольск, Россия
b Ботанический сад УрО РАН
620130 Екатеринбург, Россия
c Удмуртский государственный университет
426034 Ижевск, Россия
* E-mail: ermolaev-i@yandex.ru
Поступила в редакцию 18.02.2020
После доработки 30.04.2020
Принята к публикации 25.05.2020
Аннотация
Впервые исследована значительная часть комплекса насекомых-фитофагов дуба черешчатого (Quercus robur L.) на северо-востоке его ареала. Работу провели в течение 2013−2017 гг. близ биостанции Удмуртского государственного университета (56°82′ с.ш., 53°90′ в.д.) в пойменной дубраве р. Сива (Воткинский район, Удмуртская Республика). Комплекс насекомых-фитофагов дуба включает 95 видов насекомых из 23 семейств четырех отрядов: Lepidoptera (45.3%), Coleoptera (43.1%), Hymenoptera (9.5%) и Diptera (2.1%). Листья поедает 71 вид. При этом 58 наносят повреждение на стадии личинки и 13 − при дополнительном питании на стадии имаго. Среди последних − карпофаги, ксилофаги, ризофаги дуба и насекомые, не связанные на личиночной стадии с деревом, − насчитывают 2, 4, 1 и 6 видов соответственно. Желуди, древесину и корни дуба повреждает 4, 24 и 3 вида соответственно. Комплекс сложен из типичных представителей фитофагов Q. robur, характеризуется обедненностью видового состава и стабильно низкой численностью большинства видов.
В РФ северная граница ареала дуба Quercus robur L. проходит по линии Санкт-Петербург– Вологда–Киров–Пермь. По мнению Горчаковского (1968), это обусловлено как климатическими и эдафическими условиями, так и конкурентными отношениями с другими растениями, особенно елью. На северо-востоке ареала Q. robur произрастает его ранняя феноформа (Q. robur var. praecox Czern.). Дубравы известны здесь преимущественно в долине рек Кама и Сива в виде редких, локальных, смешанных насаждений с участием липы и вяза (Добрынин, Комиссарова, 2012). Так, доля пойменных дубрав местного национального парка “Нечкинский” составляет лишь 3% (628 га) его площади. Эти растительные ассоциации значительно обеднены растениями неморального комплекса (Горчаковский, 1968).
С дубами связан один из самых богатых в таксономическом отношении комплексов насекомых-фитофагов. В фундаментальной монографии “Вредители леса” (1955) для территории бывшего СССР указано 554 вида фитофагов рода Quercus. Из них доминировали виды отряда Lepidoptera – 64.6%, представители Coleoptera, Hymenoptera, Orthoptera и Diptera составили 22.8, 9.4, 2.7 и 0.5% соответственно. Согласно данным Довнара-Запольского (1954), только на территории европейской части бывшего СССР было известно 850 видов членистоногих-фитофагов Q. robur (Довнар-Запольский, 1954). Среди них филлофаги составили 62.3%, ксилофаги, ризофаги, карпофаги и виды, косвенно связанные с дубом (питающиеся на грибах, лишайниках, в покинутых галлах), – 12.5, 10.5, 5.9 и 8.8%, соответственно. При этом 163 вида были экономически значимы (Довнар-Запольский, 1953).
Основное внимание при изучении насекомых-фитофагов Q. robur в европейской части бывшего СССР было уделено различным аспектам биологии прежде всего экономически значимых видов (например, Шевырев, 1891; Сахаров, 1928; Померанцев, 1949; Вайнштейн, 1950; Зелинская, 1964; Покозий, 1965; Прокопенко, 1975; Ивашов, 1977; Саввин, 1979; Мезенцев, 1980; Апостолов, 1981; Рубцов, Рубцова, 1984; Дубровин, 1987; Зерова и др., 1989; Уткина, 1991; Гамаюнова и др., 1999; Рубцов, Уткина, 2008).
К сожалению, работ, посвященных анализу всего комплекса насекомых-фитофагов дуба Q. robur конкретной географической территории, крайне мало. Пожалуй, одна из наиболее интересных работ – диссертация Плугаря (1964). В ходе исследования энтомофауны различных видов дубов Молдавии автор выявил 369 видов фитофагов. Чаще всего исследования сведены к анализу отдельных экологических группировок, например, филло- (Моравская, 1952; 1975) или ксилофагов (Линдеман, 1966).
Цель представленной работы – исследовать видовую структуру энтомокомплексов филло-, карпо-, ксило- и ризофагов дуба Q. robur на северо-востоке его ареала.
МАТЕРИАЛ И МЕТОДИКА
При анализе сезонной динамики повреждения листьев дуба черешчатого (Q. robur L.) провели исследование видовой структуры экологических групп насекомых филло-, карпо-, ксило- и ризофагов. Изучение равнокрылых (Homoptera) и полужесткокрылых (Hemiptera) не входило в задачи работы.
Исследования провели в течение 2013–2017 гг. близ биостанции Удмуртского государственного университета (56°82′ с.ш., 53°90′ в.д.) в пойменной дубраве р. Сива (Воткинский район Удмуртской Республики) (рис. 1). С этой целью в соответствии с ОСТ 56-69-83 были заложены две пробные площади.
Рис. 1.
Район исследования (на врезке ареал Quercus robur выделен темным цветом, место проведения исследования − точкой): 1 – пробная площадь № 1, 2 – пробная площадь № 2, 3 − с. Перевозное.
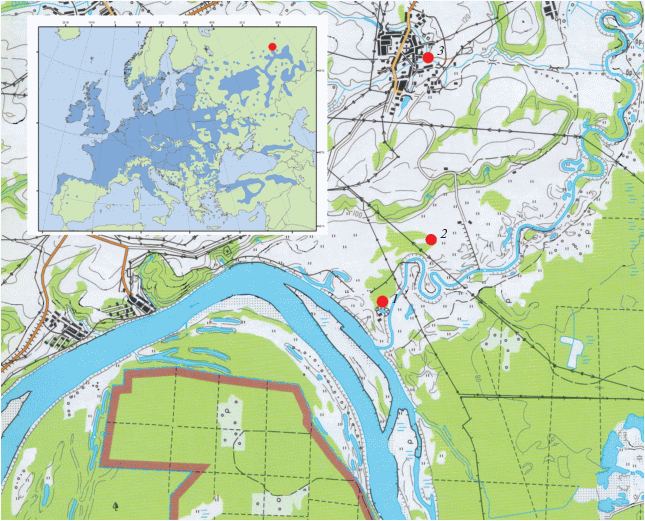
Пробная площадь № 1 располагалась в 34 квартале Костоватовского лесничества Национального парка “Нечкинский” близ биостанции “Сива”. Древостой липового дубняка был расположен в низине и представлен дубом черешчатым, липой мелколистной (Tilia cordata Mill.), вязом гладким (Ulmus laevis Pall.) и осиной (Populus tremula L.). Древостой состоял из 40% дуба, 30% липы, 20% вяза и 10% осины (4Д3Лп2В1Ос). Бонитет IV класса. Относительная полнота 0.8. Высота деревьев дуба 14.6 ± 0.2 м, диаметр ствола на высоте 130 см (на уровне груди) 34.5 ± 0.5 см (n = 100). Возраст древостоя 100 лет. Подрост высотой 5–6 м, включает липу и вяз.
Пробная площадь № 2 была заложена в 102 квартале Июльского участкового лесничества. Более высокое и сухое место. Древостой липового дубняка был представлен дубом черешчатым, липой мелколистной и осиной. Древостой состоял из 40% дуба, 30% липы и 30% осины (4Д3Лп3Ос). Бонитет IV класса. Относительная полнота 0.7. Высота деревьев дуба 16.1 ± 0.3 м, диаметр ствола на высоте 130 см – 37.2 ± 0.5 см (n = 100). Возраст древостоя 110 лет. Подрост слабо выражен, до 6 м, представлен дубом, липой и вязом.
Высоту дерева определяли с помощью электронного клинометра Haglöf EC II, диаметр на высоте 1.3 м – мерной вилкой Mantax Blue (800 мм) Haglöf.
В работе использованы карты “Удмуртская Республика. Атлас. Масштаб 1 : 100 000”, составленные ФГУП “Уралаэрогеодезия” (2009) и “Pedunculate Oak (Quercus robur) Distribution Map” электронного ресурса “European Forest Genetic Resources Programme” (2020).
Сбор филлофагов дуба осуществляли в течение вегетации обтряхиванием ветвей (не модельного) дерева внутри энтомологического сачка и на расстеленный под кроной дерева полог. Питавшихся дубом имаго усыпляли и этикетировали. Личинок свободноживущих насекомых выкармливали в полевой лаборатории до окукливания. Минеров собирали на стадии куколки. Листья с минами и галлами гербаризировали и этикетиковали. Часть редких чешуекрылых-филлофагов, специализированных на Q. robur, была выявлена с помощью световой ловушки (на основе лампы ДРЛ-250) на биостанции “Сива”. Сбор бабочек проводили периодически в течение 2013−2016 гг. Полученный список чешуекрылых был дополнен не выявленными в ходе нашего исследования и специализированными на дубе видами пядениц и совок, собранными в 1967−1988 гг. на биостанции “Сива” Е.М. Антоновой, А.В. Свиридовым и В.И. Рощиненко и отмеченными в их публикации (Антонова и др., 1989) как пойманные в ближайшем населенном пункте (с. Перевозное).
В качестве объекта для оценки суммы эффективных температур (далее СЭТ) онтогенетического развития был выбран непарный шелкопряд Lymantria dispar (L.). Из всего комплекса выявленных фитофагов Q. robur именно этот вид представляет максимальный интерес. Удмуртия входит в зону периодического интенсивного вреда L. dispar (Лямцев, 2014). Помимо этого, он имеет важное экономическое значение для соседних территорий: Татарстана (Селиховкин и др., 2014) и Башкортостана (Ханисламов и др., 1962).
Феромонный мониторинг популяции L. dispar был проведен в период в 2014−2016 гг. близ биостанции “Сива”. При мониторинге были использованы коробчатые инсектицидные ловушки типа “молочный пакет”, с диспенсерами, содержащими 500 мкг (+) – диспарлюра, и инсектицидными ловушками, пропитанными 2,2-дихлорвинилом-50 диметилфосфатом (производство США). Ежегодно использовали 10 ловушек. Ловушки располагали на ветвях деревьев на высоте 1.5−2.0 м от земли. Расстояние между ловушками 500 м.
Сумму эффективных температур онтогенетического развития L. dispar рассчитали по результатам феромонного мониторинга. Температура нижнего порога развития 7°С, верхнего – 24°С. Данные погоды в месте исследования были взяты из архива погоды ближайшей метеостанции (г. Чайковский).
Для сравнения сроков лёта и уловистости в разных регионах были использованы данные по феромонному мониторингу с помощью того же типа ловушек, полученные авторами в Нижнем Поволжье (Волго-Ахтубинская пойма, г. Краснослободск), Республике Кыргызстан (г. Бишкек, Ботанический сад), Свердловской области (г. Екатеринбург).
Сбор ксилофагов дуба осуществляли в течение 2013−2016 гг. на ослабленных и погибших в результате воздействия разных факторов (ветровал, санитарная вырубка вдоль ЛЭП, низовой пожар) стволах, под корой и в древесине дерева. Особенности заселения ствола разными видами ксилофагов были исследованы на примере 10 стволов дуба, срубленных сотрудниками электросетей при чистке ЛЭП Воткинской ГЭС.
Комплекс ризофагов дуба исследовали в 2016 г. методом почвенных раскопок (Голуб и др., 2012). В общей сложности под корнями дерева было выкопано 5 ям площадью 0.5 × 1 м.
Помимо классических “определителей насекомых европейской части СССР” использовали определитель Гусева (1984). Видовая принадлежность всех собранных фитофагов подтверждена специалистами.
Среднюю сумму эффективных температур развития L. dispar до имагинальной стадии рассчитывали по медиане лёта самцов, определяемой как момент выхода из куколки 50% особей. Сумма эффективных температур летне-осеннего развития эмбрионов (раннеэмбриональная стадия) рассчитывали от медианы лёта самцов предыдущего года до устойчивого перехода температур осенью ниже +7°С.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
В течение вегетации листья Q. robur в долине р. Сива повреждают личинки 58 видов насекомых. Это представители 18 семейств четырех отрядов: Lepidoptera (72.4%), Hymenoptera (15.5%) Coleoptera (8.6%) и Diptera (3.5%). При этом листья объедают личинки 35 видов, минируют, скелетируют, образуют галлы и трубки – 8, 2, 7 и 4 вида соответственно (табл. 1). Среди объедателей доминировали Hypomecis roboraria, Malacosoma neustria и Phalera bucephala, среди минеров, скелетеров, галло- и трубкообразователей − Stigmella spp., Altica quercetorum, Cynips quercusfolii и Ancylis mitterbacheriana соответственно. Личинки долгоносиков Archarius pyrrhoceras и Curculio villosus проходят свое развитие, поедая ткани внутри галлов на листьях дерева. C. villosus известен на исследуемой территории по локальной находке в левобережной пойме р. Кама (Дедюхин, 2009а; 2012).
Таблица 1.
Насекомые-фитофаги дуба черешчатого Q. robur долины р. Сива
| № | Вид | Трофическая специализация | Встречаемость | |
|---|---|---|---|---|
| личинки | имаго | |||
| Scarabaeidae | ||||
| 1 | Melolontha hippocastani Fabricius 1801 | Ризофаг | Филлофаг | Обычный |
| 2 | Amphimallon solstitiale (Linnaeus 1758) | Ризофаг | Обычный | |
| Buprestidae | ||||
| 3 | Chrysobothris affinis (Fabricius 1794) | Ксилофаг | Массовый | |
| 4 | Agrilus biguttatus (Fabricius 1777) | Ксилофаг | Филлофаг | Обычный |
| 5 | A. angustulus angustulus (Illiger 1803) | Ксилофаг | Филлофаг | Обычный |
| Elateridae | ||||
| 6 | Selatosomus cruciatus (Linnaeus 1758) | Филлофаг | Обычный | |
| Cerambycidae | ||||
| 7 | Rhagium mordax (DeGeer 1775) | Ксилофаг | Обычный | |
| 8 | Stenocorus meridianus (Linnaeus 1758) | Ризофаг, Ксилофаг | Обычный | |
| 9 | Macroleptura thoracica (Creutzer 1799) | Ксилофаг | Редкий | |
| 10 | Leptura quadrifasciata quadrifasciata Linnaeus 1758 | Ксилофаг | Обычный | |
| 11 | Strangalia attenuata (Linnaeus 1758) | Ксилофаг | Обычный | |
| 12 | Necydalis major major Linnaeus 1758 | Ксилофаг | Обычный | |
| 13 | Purpuricenus kaehleri (Linnaeus 1758) | Ксилофаг | Редкий | |
| 14 | Phymatodes alni (Linnaeus 1767)* | Ксилофаг | Редкий | |
| 15 | Plagionotus detritus (Linnaeus 1758) | Ксилофаг | Редкий | |
| 16 | P. arcuatus (Linnaeus 1758) | Ксилофаг | Редкий | |
| 17 | Xylotrechus arvicola (Olivier 1795) | Ксилофаг | Редкий | |
| 18 | Chlorophorus herbstii (Brahm 1790) | Ксилофаг | Редкий | |
| 19 | Rhaphuma gracilipes (Faldermann 1835) | Ксилофаг | Редкий | |
| 20 | Mesosa myops (Dalman 1817) | Ксилофаг | Массовый | |
| 21 | Anaesthetis testacea (Fabricius 1781) | Ксилофаг | Редкий | |
| 22 | Acanthoderes clavipes (Schrank 1781) | Ксилофаг | Филлофаг | Обычный |
| 23 | Leiopus linnei Wallin, Nylander & Kvamme 2009 | Ксилофаг | Редкий | |
| 24 | Oplosia cinerea (Mulsant 1839)* | Ксилофаг | Редкий | |
| 25 | Saperda scalaris hieroglyphica (Pallas 1773) | Ксилофаг | Филлофаг | Обычный |
| Chrysomelidae | ||||
| 26 | Cryptocephalus flavipes Fabricius 1781 | Филлофаг | Обычный | |
| 27 | Luperus flavipes (Linnaeus 1758) | Филлофаг | Обычный | |
| 28 | Altica quercetorum Foudras 1860 | Филлофаг | Филлофаг | Массовый |
| Attelabidae | ||||
| 29 | Byctiscus populi (Linnaeus 1758) | Филлофаг | Филлофаг | Обычный |
| Curculionidae | ||||
| 30 | Tropideres albirostris (Schaller 1783) | Ксилофаг | Обычный | |
| 31 | Magdaliscerasi (Linnaeus 1758) | Ксилофаг | Редкий | |
| 32 | Coeliodes transversealbofasciatus (Goeze 1777)** | Карпофаг | Редкий | |
| 33 | Curculio glandium Marsham 1802 | Карпофаг | Филлофаг | Массовый |
| 34 | C. venosus (Gravenhorst 1807) | Карпофаг | Филлофаг | Редкий |
| 35 | C. villosus Fabricius 1781** | Филлофаг | Редкий | |
| 36 | Archarius pyrrhoceras (Marsham 1802) | Филлофаг | Редкий | |
| 37 | Orchestessignifer (Creutzer 1799) | Филлофаг | Обычный | |
| 38 | Phyllobius pyri (Linnaeus 1758) | Филлофаг | Массовый | |
| 39 | Polydrusus flavipes (Degeer 1775) | Филлофаг | Обычный | |
| 40 | P. undatus (Fabricius 1781) | Филлофаг | Обычный | |
| 41 | Scolytus intricatus (Ratzeburg 1837) | Ксилофаг | Массовый | |
| Tenthredinidae | ||||
| 42 | Allantus togatus (Panzer 1801) | Филлофаг | Редкий | |
| 43 | Caliroa cinxia (Klug 1816) | Филлофаг | Редкий | |
| 44 | Profenusa pygmaea (Klug 1816) | Филлофаг | Обычный | |
| 45 | Mesoneura opaca (Fabricius 1775) | Филлофаг | Редкий | |
| Cynipidae | ||||
| 46 | Neuroterus quercusbaccarum (Linnaeus 1758) | Филлофаг | Редкий | |
| 47 | Cynips quercusfolii Linnaeus 1758 | Филлофаг | Массовый | |
| 48 | Andricus curvator Hartig 1840 | Филлофаг | Редкий | |
| 49 | A. foecundatrix (Hartig 1840) | Филлофаг | Обычный | |
| 50 | A. quadrilineatus Hartig 1840 | Филлофаг | Редкий | |
| Nepticulidae | ||||
| 51 | Stigmella sp. 1 | Филлофаг | Обычный | |
| 52 | Stigmella sp. 2 | Филлофаг | Редкий | |
| Tischeriidae | ||||
| 53 | Tischeria ekebladella (Bjerkander 1795) | Филлофаг | Редкий | |
| Gracillariidae | ||||
| 54 | Acrocercops brongniardella Fabricius 1798 | Филлофаг | Редкий | |
| 55 | Phyllonorycter roboris (Zeller 1839) | Филлофаг | Обычный | |
| Coleophoridae | ||||
| 56 | Coleophora sp. | Филлофаг | Редкий | |
| Limacodidae | ||||
| 57 | Apoda limacodes (Hufnagel 1766) | Филлофаг | Редкий | |
| Tortricidae | ||||
| 58 | Tortrix viridana Linnaeus 1758 | Филлофаг | Редкий | |
| 59 | Acleris quercinana (Zeller 1849) | Филлофаг | Редкий | |
| 60 | Ancylis mitterbacheriana ([Denis & Schiffermüller] 1775) | Филлофаг | Обычный | |
| 61 | Cydia triangulella (Goeze 1783) | Карпофаг | Редкий | |
| Geometridae | ||||
| 62 | Ennomos autumnaria (Werneburg 1859) | Филлофаг | Редкий | |
| 63 | E. erosaria ([Denis & Schiffermüller] 1775) | Филлофаг | Редкий | |
| 64 | Selenia lunularia (Hübner [1788])*** | Филлофаг | Редкий | |
| 65 | Plagodis dolabraria (Linnaeus 1767)*** | Филлофаг | Редкий | |
| 66 | P. pulveraria (Linnaeus 1758)*** | Филлофаг | Редкий | |
| 67 | Hypomecispunctinalis (Scopoli 1763) | Филлофаг | Обычный | |
| 68 | H. roboraria ([Denis & Schiffermüller] 1775) | Филлофаг | Массовый | |
| 69 | Parectropis similaria (Hufnagel 1767)*** | Филлофаг | Редкий | |
| 70 | Cyclophora porata (Linnaeus 1767) | Филлофаг | Редкий | |
| 71 | C. punctaria (Linnaeus 1758) | Филлофаг | Обычный | |
| 72 | C. quercimontaria (Bastelberger 1897) | Филлофаг | Редкий | |
| Lasiocampidae | ||||
| 73 | Malacosoma neustria (Linnaeus 1758) | Филлофаг | Массовый | |
| 74 | Gastropacha quercifolia (Linnaeus 1758) | Филлофаг | Редкий | |
| Shingidae | ||||
| 75 | Mimas tiliae (Linnaeus 1758) | Филлофаг | Редкий | |
| Notodontidae | ||||
| 76 | Drymonia dodonaea ([Denis & Schiffermüller] 1775) | Филлофаг | Редкий | |
| 77 | Ptilodon capucina (Linnaeus 1758) | Филлофаг | Обычный | |
| 78 | Phalera bucephala (Linnaeus 1758) | Филлофаг | Массовый | |
| Lymantriidae | ||||
| 79 | Calliteara pudibunda (Linnaeus 1758) | Филлофаг | Обычный | |
| 80 | Euproctis similis (Fuessly 1775) | Филлофаг | Обычный | |
| 81 | Lymantria dispar (Linnaeus 1758) | Филлофаг | Обычный | |
| Noctuidae | ||||
| 82 | Meganola strigula ([Denis & Schiffermüller] 1775) | Филлофаг | Редкий | |
| 83 | Pseudoips prasinana (Linnaeus 1758) | Филлофаг | Обычный | |
| 84 | Trisateles emortualis ([Denis & Schiffermüller] 1775)*** | Филлофаг | Редкий | |
| 85 | Catephia alchymista ([Denis & Schiffermüller] 1775)*** | Филлофаг | Редкий | |
| 86 | Catocala promissa ([Denis & Schiffermüller] 1775) | Филлофаг | Редкий | |
| 87 | C. sponsa (Linnaeus 1767) | Филлофаг | Редкий | |
| 88 | Colocasia coryli (Linnaeus 1758) | Филлофаг | Редкий | |
| 89 | Moma alpium (Osbeck 1778) | Филлофаг | Редкий | |
| 90 | Acronicta aceris (Linnaeus 1758) | Филлофаг | Редкий | |
| 91 | A. auricoma ([Denis & Schiffermüller] 1775) | Филлофаг | Обычный | |
| 92 | Conistra vaccinii (Linnaeus 1761) | Филлофаг | Редкий | |
| Lycaenidae | ||||
| 93 | Nordmannia w-album (Knoch 1782) | Филлофаг | Редкий | |
| Cecidomyiidae | ||||
| 94 | Macrodiplosis dryobiae (Low 1877) | Филлофаг | Редкий | |
| 95 | M. volvens Kieffer 1895 | Филлофаг | Редкий | |
В процессе своего развития личинки ряда видов наносили по два вида повреждений. Так, гусеницы младших возрастов объедателей M. neustria, Ph. bucephala, Calliteara pudibunda, Euproctis similis, Meganola strigula скелетируют листья. Личинки трубкообразователей скелетируют и объедают свернутый лист.
Дополнительное питание листьями дуба проходят имаго шести видов жуков: Selatosomus cruciatus (Linnaeus 1758), Cryptocephalus flavipes Fabricius,1781, Luperus flavipes (Linnaeus 1758), Phyllobius pyri (Linnaeus 1758), Polydrusus flavipes (Degeer 1775) и P. undatus (Fabricius 1781). При этом нет достоверных данных о питании личинок указанных видов какими-либо органами дуба.
Виды, потенциально способные питаться на дубе, представлены в табл. 2. Все указанные полифаги были пойманы близ биостанции энтомологическим сачком (жуки) или с помощью световой ловушки (бабочки). Питание этих видов на Q. robur известно в более южных географических точках, однако в районе исследований этого не наблюдали.
Таблица 2.
Виды, потенциально способные питаться на дубе Q. robur
| № | Вид | Встречаемость |
|---|---|---|
| Chrysomelidae | ||
| 1 | Labidostomis tridentata (Linnaeus 1758)* | Редкий |
| 2 | Clytra quadripunctata (Linnaeus 1758) | Обычный |
| 3 | Cryptocephaluscordiger (Linnaeus 1758)* | Редкий |
| 4 | C. bipunctatus (Linnaeus 1758) | Обычный |
| 5 | C. flavipes Fabricius 1781 | Обычный |
| 6 | C. labiatus (Linnaeus 1761) | Обычный |
| 7 | C. pusillus Fabricius 1777* | Редкий |
| 8 | C. quinquepunctatus (Scopoli 1763)* | Редкий |
| 9 | Altica brevicollis Foudras 1860* | Редкий |
| Tortricidae | ||
| 10 | Acleris hastiana (Linnaeus 1758) | Редкий |
| 11 | Choristoneura hebenstreitella (Müller 1764) | Редкий |
| 12 | Ptycholoma lecheana (Linnaeus 1758) | Редкий |
| 13 | Notocelia cynosbatella (Linnaeus 1758) | Обычный |
| Geometridae | ||
| 14 | Cabera pusaria (Linnaeus 1758) | Обычный |
| 15 | Ascotis selenaria ([Denis & Schiffermüller] 1775) | Обычный |
| 16 | Ectropis crepuscularia ([Denis & Schiffermüller] 1775)** | Редкий |
| 17 | Idaea biselata (Hufnagel 1767) | Редкий |
| Lymantriidae | ||
| 18 | Lymantria monacha (Linnaeus 1758) | Обычный |
| Noctuidae | ||
| 19 | Catocala fraxini (Linnaeus 1758) | Обычный |
| 20 | Acronicta leporina (Linnaeus 1758)** | Редкий |
| 21 | Amphipyra pyramidea (Linnaeus 1758)** | Редкий |
| 22 | Cosmia pyralina ([Denis & Schiffermüller] 1775)** | Редкий |
| 23 | C. trapezina (Linnaeus 1758)** | Редкий |
| 24 | Euplexia lucipara (Linnaeus 1758) | Обычный |
| 25 | Lacanobia w-latinum (Hufnagel 1766) | Редкий |
| 26 | Agrotis exclamationis (Linnaeus 1758) | Обычный |
Сумму эффективных температур онтогенетического развития L. dispar оценили по результатам феромонного мониторинга. Согласно литературным данным (Ильинский, 1965), его онтогенез включает четыре периода: 1) раннеэмбриональное развитие перед зимовкой (до 300 градусо-дней при пороге развития 7°С), 2) весеннее доразвитие эмбрионов (до 110 градусо-дней при пороге развития 6°С), 3) развитие гусениц – самцов (650 градусо-дней при пороге развития 6°С) и самок (740 градусо-дней при пороге развития 6 °С), 4) развитие куколок – самцов (170 градусо-дней при пороге развития 9°С) и самок (140 градусо-дней при пороге развития 9°С). Таким образом, при пороге развития 7°С для полного развития яиц (весной), гусениц и куколок требуется сумма эффективных температур равная 930 (для самцов) − 990 градусо-дням (для самок) (Ильинский и др., 1965). Показатели порога развития L. dispar могут значительно варьировать в разных популяциях (Мешкова, 2009).
Результаты исследования, проведенного посредством феромонного мониторинга, показали, что время и медиана лёта самцов непарного шелкопряда в долине р. Сива могут варьировать в разные годы (рис. 2). В годы учета фенологический сдвиг начала лёта в пойме р. Сива достигал 10 дней: 5 июля в 2015 г. и 15 июля 2014 г. Тоже справедливо для окончания лёта: 1 августа 2016 г. и 9 августа 2014 г. В общей сложности продолжительность лёта составляет около месяца (от 26 до 32 дней). В то же время, согласно данным, приведенным в табл. 3, календарные сроки лёта в популяциях разного широтного происхождения различаются не существенно. Медиана лёта, как правило, во всех популяциях приходится на середину − конец июля. Однако в отдельные годы в северных популяциях (Удмуртская Республика, Свердловская область) отмечен сдвиг медианы лёта на начало августа.
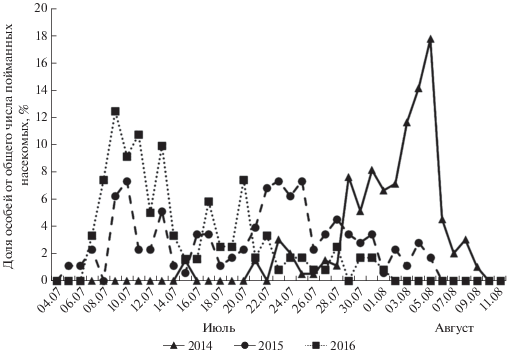
Таблица 3.
Продолжительность и даты лёта самцов непарного шелкопряда в популяциях разных широтных регионов
| Год | Дата | Продолжительность лёта, дни |
||
|---|---|---|---|---|
| начала лёта | конца лёта | медиана | ||
| Удмуртская Республика, Воткинский р-н, биостанция “Сива” | ||||
| 2014 | 15 июля | 9 августа | 2 августа | 26 |
| 2015 | 5 июля | 5 августа | 22 июля | 32 |
| 2016 | 7 июля | 1 августа | 12 июля | 26 |
| Свердловская обл., г. Екатеринбург | ||||
| 2014 | 31 июля | 19 августа | 9 августа | 20 |
| 2015 | 16 июля | 24 июля | 23 июля | 9 |
| 2016 | 8 июля | 31 июля | 20 июля | 24 |
| Волгоградская обл., Волго-Ахтубинская пойма, Лещевское лесничество | ||||
| 2011 | 08 июля | 28 июля | 15 июля | 21 |
| 2012 | 07 июля | 27 июля | 18 июля | 21 |
| Республика Кыргызстан, г. Бишкек, Ботанический сад | ||||
| 2012 | 25 июня | 30 июля | 24 июля | 36 |
| 2013 | 24 июня | 05 августа | 18 июля | 43 |
Незначительные календарные сдвиги медианы лёта самцов в северных популяциях связаны с суммой эффективных температур развития этого вида до имаго. Значения этого показателя для развития L. dispar близ биостанции “Сива” в 2014−2016 гг. представлены в табл. 4. Среднее значение, необходимое для развития особей филлофага до имаго, составило 847.1 градусо-дней, что совпадает с показателями развития непарного шелкопряда до имаго в северных популяциях Урала и Западной Сибири (от 750 до 920 градусо-дней) и существенно меньше суммы эффективных температур развития этого вида до имаго в южных популяциях. Так, в Нижнем Поволжье (Волго-Ахтубинская пойма) показатель составлял в разные годы от 1010 до 1580 градусо-дней, в Кыргызстане (г. Бишкек) – от 980 до 1208 градусо-дней (Ponomarev et al., 2019).
Таблица 4.
Показатели развития суммы эффективных температур (градусо-дни) L. dispar близ биостанции “Сива” в 2014−2016 гг.
| Год | Сумма эффективных температур развития до стадии имаго | Сумма эффективных температур | |||
|---|---|---|---|---|---|
| начала лёта | конца лёта | медиана | летне-осеннего развития эмбрионов | вегетационного сезона | |
| 2014 | 756.1 | 1013.9 | 909.5 | 461.2 | 1370.7 |
| 2015 | 698.1 | 1003.9 | 850.4 | 550.6 | 1401.2 |
| 2016 | 686.0 | 1066.3 | 781.5 | 904.6 | 1686.1 |
Общая теплообеспеченность сезона достаточна для устойчивого существования местной популяции L. dispar. Тем не менее, уловистость на ловушку была очень небольшая, от 7 до 43 самцов, что значительно меньше уловистости для данного типа ловушек по результатам мониторинга в других регионах (Свердловская область, Нижнее Поволжье, Республика Кыргызстан) даже при небольшой плотности по яйцекладкам (150–500 самцов на ловушку при плотности кладок 0.05–0.1 на дерево (Пономарев и др., 2014). Эти результаты, а также учет кладок в районе исследования (при осмотре до 200−300 деревьев кладки обнаружены не были) свидетельствуют о крайне низкой плотности популяции непарного шелкопряда в пойменных дубравах р. Сива в период исследований.
Помимо классических филлофагов листья дуба при дополнительном питании повреждали представители имаго карпо-, ксило- и ризофагов.
Карпофаги дуба долины р. Сива представлены 4 видами. В женских соцветиях дерева проходит развитие личинки долгоносика Coeliodes transversealbofasciatus (Goeze 1777) (Дедюхин, 2012). Среди карпофагов выявлены Curculio glandium Marsham 1802, C. venosus (Gravenhorst 1807) и Cydia triangulella (Goeze 1783). Доминировал C. glandium. В начале вегетации имаго Curculio spp. проходят дополнительное питание на листьях дуба.
В комплекс ксилофагов Q. robur входят 24 вида из трех семейств Coleoptera. На ослабленные дубы нападают Chrysobothris affinis (Fabricius 1794), Agrilus biguttatus (Fabricius 1777), A. angustulus angustulus (Illiger 1803), Plagionotus arcuatus (Linnaeus 1758), P. detritus (Linnaeus 1758), Mesosa myops (Dalman 1817) и Scolytus intricatus (Ratzeburg 1837). Под корой погибших дубов проходят развитие личинки 15 видов усачей и 2 видов долгоносиков (табл. 1). Особый интерес представляет находка усача Leiopus linnei Wallin, Nylander & Kvamme 2009, описанного десять лет назад из Скандинавии (Wallin et al., 2010). Дополнительное питание на листьях дуба проходят имаго A. biguttatus, A. angustulus angustulus, A. clavipes и S. scalaris hieroglyphica.
Комплекс ризофагов был представлен тремя видами: Melolontha hippocastani Fabricius 1801, Amphimallon solstitiale (Linnaeus 1758) и Stenocorus meridianus (Linnaeus 1758). Имаго M. hippocastani проходит дополнительное питание, в том числе и на листьях дуба. Среди щелкунов в дубраве обычны следующие виды: Agrypnus murinus (Linnaeus 1758), Agriotes obscurus (Linnaeus 1758), A. lineatus (Linnaeus 1767), A. sputator (Linnaeus 1758), Dalopius marginatus (Linnaeus 1758), Melanotus castanipes (Paykull 1800), M. villosus (Fourcroy 1785), Hemicrepidius niger (Linnaeus 1758), Athous haemorrhoidalis (Fabricius 1801), Denticollis linearis (Linnaeus 1758), Ctenicera pectinicornis (Linnaeus 1758), Paraphotistus nigricornis (Panzer 1799), Selatosomus aeneus (Linnaeus 1758). Однако прямая трофическая связь личинок этих жуков с дубом нами не установлена.
Таким образом, комплекс насекомых-фитофагов дуба черешчатого в долине р. Сива включает 95 видов насекомых из 23 семейств четырех отрядов: Lepidoptera (45.3%), Coleoptera (43.1%), Hymenoptera (9.5%) и Diptera (2.1%). Листья поедает 71 вид насекомых. При этом 58 наносят повреждение на стадии личинки и 13 − при дополнительном питании на стадии имаго. Среди последних видов − карпофаги, ксилофаги, ризофаги дуба и насекомые, не связанные на личиночной стадии с деревом, насчитывают 2, 4, 1 и 6 видов соответственно. Монофагия наблюдается у галлообразователей (100%), минеров (75%), карпофагов (25%) и ксилофагов (4%). Желуди, древесину и корни дуба повреждают 4, 24 и 3 вида соответственно. Комплекс сложен из типичных представителей фитофагов Q. robur, однако имеет две особенности.
Первая особенность связана с обедненностью видового состава комплекса. Низкая теплообеспеченность на северо-востоке ареала дуба ограничивает возможность существования значительного числа видов фитофагов дерева. Пойменная дубрава р. Сивы, являясь самой северной в Удмуртии, не имеет в составе энтомокомплексов такие виды как Lucanus cervus (Linnaeus 1758) (Lucanidae), Attelabus nitens (Scopoli 1763), Coeliodes trifasciatus Bach 1854, Orchestes quercus (Linnaeus 1758) (Curculionoidea), Rhagium sycophanta (Schrank 1781), Anoplodera sexguttata (Fabricius 1775), Phymatodes testaceus (Linnaeus 1758), Clytus arietis (Linnaeus 1758) и Cyrtoclytus capra (Germar 1824) (Cerambycidae). Между тем в южных районах Удмуртии эти виды хорошо известны (Дедюхин, 2005; 2012; Красная книга Удмуртской Республики, 2012). Кроме того, в комплексе фитофагов исследованной территории отсутствуют такие заметные и экономически значимые виды как Aleimma loeflingiana (Linnaeus 1758) (Tortricidae), Lycia hirtaria (Clerck 1759), Erannis defoliaria Leach 1815, Operophthera fagata (Scharfenberg 1805) (Geometridae), Eriogaster lanestris (Linnaeus 1758) (Lasiocampidae), Peridea anceps (Goeze 1781) (Notodontidae), Euproctis chrysorrhoea Linnaeus 1758, Parocneria detrita (Esper 1785) (Lymantriidae). В итоге, если в нашем случае был выявлен 71 вид насекомых, повреждающих листья Q. robur (табл. 1), то по югу ареала дерева, например, в Молдавии, Воронежской и Волгоградской областях РФ − 243 (Плугарь, 1963), 184 (Моравская, 1954; 1957; 1975) и 140 (Моравская, 1952) видов филлофагов соответственно. Схожая картина была показана при исследовании зонально-поясной фауны филлофагов лиственницы (Плешанов, 1982). Если в районах с низкой теплообеспеченностью при сумме биологически активных температур 800−1200° на лиственнице обитает от 6 до 12 видов филлофагов, то в древостоях с умеренной (1200−1600°) и высокой теплообеспеченностью (1600−2000°) на дереве встречается от 23 до 31 и от 48 до 60 видов соответственно (Плешанов, 1982). В условиях низкой теплообеспеченности конкурентное преимущество получают виды фитофагов, для развития которых необходима относительно небольшая сумма эффективных температур.
Вторая особенность выражается в стабильно низкой численности большинства членов комплекса. За более чем 35-летний период функционирования биостанции “Сива” не было отмечено ни одного случая реализации эруптивного типа динамики численности фитофагов. Это относится в том числе и к популяциям таких экономически значимых видов, как Altica quercetorum, Acrocercops brongniardella, Tortrix viridana, Cydia triangulella, Malacosoma neustria, Phalera bucephala, Calliteara pudibunda и Lymantria dispar. На примере ареала последнего вида было показано, что при продвижении с юга на север в популяциях филлофага происходит смена типов динамики численности с эруптивного на продромальный и далее на стабильный (Эпова, Плешанов, 1995, стр. 12). На наш взгляд, трансформация типов динамики численности в популяциях насекомых может быть связана с зональным изменением их стаций.
Список литературы
Антонова Е.М., Свиридов А.В., Рощиненко В.И., 1989. Фауна пядениц и совок (Geometridae, Noctuidae) Удмуртии // Фауна и экология животных УАССР и прилежащих районов. Ижевск: УдГУ. С. 108−121.
Апостолов Л.Г., 1981. Вредная энтомофауна лесных биогеоценозов Центрального Приднепровья. Киев: Вища школа. 281 с.
Вайнштейн Б.А., 1950. Листогрызущие вредители дуба и их сезонная динамика // Зоологический журнал. Т. 29. Вып. 2. С. 107−112.
Вредители леса, 1955. М.; Л.: Издательство АН СССР. Т I., Т II. 1097 с.
Гамаюнова С.Г., Новак Л.В., Войтенко Ю.В., Харченко А.Е., 1999. Массовые хвое- и листогрызущие вредители леса. Харьков: Украинский научно-исследовательский институт лесного хозяйства и агролесомелиорации им. Г.Н. Высоцского. 172 с.
Голуб В.Б., Цуриков М.Н., Прокин А.А., 2012. Коллекции насекомых: сбор, обработка и хранение материала. М.: Товарищество научных изданий КМК. 339 с.
Горчаковский П.Л., 1968. Растения европейских широколиственных лесов на восточном пределе их ареала. Свердловск: Институт экологии и животных. Вып. 59. 207 с.
Гусев В.И., 1984. Определитель повреждений лесных, декоративных и плодовых деревьев и кустарников. М.: Лесная промышленность. 472 с.
Дедюхин С.В., 2005. Жуки-усачи (Coleoptera, Cerambycidae) национального парка “Нечкинский” (с обзором фауны семейства в Удмуртии) // Вестник Удмуртского Университета. Биология. № 10. С. 81−96.
Дедюхин С.В., 2009. Жуки-листоеды (Coleoptera, Chrysomelidae) национального парка “Нечкинский” // Вестник Удмуртского Университета. Биология. Науки о Земле. Вып. 1. С. 101−116.
Дедюхин С.В., 2009а. Материалы к фауне долгоносикообразных жесткокрылых (Coleoptera, Curculionoidea) национального парка “Нечкинский” // Вестник Удмуртского университета. Биология. Науки о Земле. Вып. 2. С. 34−48.
Дедюхин С.В., 2012. Долгоносикообразные жесткокрылые (Coleoptera, Curculionoidea) Вятско-Камского междуречья: фауна, распространение, экология. Ижевск: “Издательство Удмуртский университет”. 340 с.
Добрынин А.П., Коммисарова М.Г., 2012. Самые северные дубравы России. Вологда: “Полиграф-книга”. 188 с.
Довнар-Запольский Д.П., 1953. Энтомофауна дуба в пределах европейской части Советского Союза. Автореф. дис. … канд. сельскохоз. наук. Воронеж. 12 с.
Довнар-Запольский Д.П., 1954. Очерк энтомофауны черешчатого дуба (Quercus robur) в европейской части СССР // Зоологический журнал. Т. 33. Вып. 4. С. 794−806.
Дубровин В.В., 1987. Экология и динамика численности зимней пяденицы в дубравах лесостепи. Автореф. дис. … канд. биол. наук. Воронеж: Воронежский лесотехнический институт. 19 с.
Зелинская Л.М., 1964. Непарный шелкопряд и зеленая дубовая листовертка в лесах Южного Преднепровья. Автореф. дис. … канд. биол. наук. Киев: Институт зоологии. 19 с.
Зерова М.Д., Котенко А.Г., Сергина Л.Я., Толканиц В.И., 1989. Энтомофаги дубовой листовертки и непарного шелкопряда юго-запада Европейской части СССР. Киев: Наукова Думка. 200 с.
Ивашов А.В., 1977. Физиолого-биохимические основы взаимодействия дуба черешчатого и дубовой зеленой листовертки в лесных биогеоценозах Присамарья. Автореф. дис. … канд. биол. наук. Днепропетровск: Днепропетровский государственный университет. 24 с.
Ильинский А.И., Евлахова А.А., Сиротина М.И., Швецова О.И., Андреева Г.И., Кондаков Ю.П., Звоскова Г.А., Распопов П.М., Черная Г.С., 1965. Надзор, учет и прогноз массовых размножений хвое- и листогрызущих насекомых в лесах СССР. М.: Изд-во Лесная промышленность. 525 с.
Красная книга Удмуртской Республики, 2012. Чебоксары: Перфектум. 458 с.
Линдеман Г.В., 1966. Заселение дуба стволовыми вредителями в связи с его ослаблением и отмиранием в дубравах лесостепи (на примере Теллермановского леса) // Влияние животных на продуктивность лесных биогеоценозов. М.: Наука. С. 75−96.
Лямцев Н.И., 2014. Очаги размножения непарного шелкопряда в европейской России // Лесной вестник. № 6. С. 78−85.
Мезенцев А.И., 1980. Вредители стволов и ветвей, их значение и борьба с ними в очагах усыхания дуба в лесостепной части Харьковской области. Автореф. дис. … канд. биол. наук. Харьков: Харьковский сельскохозяйственный институт. 17 с.
Мешкова В.Л., 2009. Сезонное развитие хвоелистогрызущих насекомых. Харьков: Планета-принт. 396 с.
Моравская А.С., 1952. Насекомые-вредители дуба в условиях степного лесоразведения засушливого Юго-Востока. Автореф. дис. … канд. биол. наук. Москва: Московский государственный университет. 15 с.
Моравская А.С., 1954. Вредители листвы основных древесных пород Теллермановского леса (предварительное сообщение) // Сообщения института леса АН СССР. Вып. 3. С. 30−38.
Моравская А.С., 1957. Повреждаемость насекомыми рано- и позднораспускающихся форм дуба и ильмовых пород // Сообщения института леса АН СССР. Вып. 8. С. 44−63.
Моравская А.С., 1975. Насекомые-фитофаги дубравных лесов // Дубравы лесостепи в биогеоценологическом освещении. М.: Наука. С. 199−210.
Плешанов А.С., 1982. Насекомые-дефолианты лиственничных лесов Восточной Сибири. Новосибирск: Наука. 209 с.
Плугарь С.Г., 1963. Некоторые результаты изучения энтомофауны дуба в лесах Молдавии // Вредная энтомофауна Молдавии и меры борьбы с ней. Кишинев: Издательство “Картя Молдовеняскэ”. С. 7−38.
Плугарь С.Г., 1964. Вредная энтомофауна дуба в лесах Молдавии. Автореф. дис. … канд. биол. наук. Кишинев: Академия наук Молдавской ССР. 22 с.
Покозий И.Т., 1965. Важнейшие листогрызущие и почвообитающие вредители дуба в Придонецкой части Украины и борьба с ними. Автореф. дис. … докт. биол. наук. Харьков: Харьковский сельскохозяйственный институт. 40 с.
Померанцев Д.В., 1949. Вредные насекомые и борьба с ними в лесах и лесных полосах юго-востока европейской части СССР. М., Л.: Гослесбумиздат. 212 с.
Пономарев В.И., Клобуков Г.И., Орозумбеков А.А., Серый Г.А., 2014. Влияние погодных факторов на результативность феромонного мониторинга непарного шелкопряда Lymantria dispar (L.) // Известия Санкт-Петербургской лесотехнической академии. Вып. 207. С. 202−211.
Прокопенко Н.И., 1975. Дубовая зеленая листовертка (Tortrix viridana L.) в насаждениях Харьковской и Ворошиловской областей в 1961−1971 гг. и мероприятия по борьбе с ней. Автореф. дис. … канд. биол. наук. Харьков: Харьковский сельскохозяйственный институт. 18 с.
Рубцов В.В., Рубцова Н.Н., 1984. Анализ взаимодействия листогрызущих насекомых с дубом. М.: Наука. 183 с.
Рубцов В.В., Уткина И.А., 2008. Адаптационные реакции дуба на дефолиацию. М.: Институт лесоведения РАН. 302 с.
Саввин И.М., 1979. Вредные лесные насекомые и их роль в отмирании дубрав Чувашской АССР. Автореф. дис. … канд. сельскохоз. наук. Л.: Лесотехническая академия. 17 с.
Сахаров Н., 1928. Вредные для лесных и садовых насаждений бабочки. Аткарск: Типография № 1 Промкомбината Аткарского УИКа. 51 с.
Селиховкин А.В., Магдеев Н.Г., Поповичев Б.Г., Ахматович Н.А., 2014. Доминирующие вредители леса в системе контроля плотности популяций в республике Татарстан // Лесной вестник. № 6. С. 93−101.
Удмуртская Республика. Атлас. Масштаб 1 : 100 000, 2009. ФГУП “Уралаэрогеодезия”. 124 с.
Уткина И.А., 1991. Восстановление листвы дуба черешчатого после дефолиации крон листогрызущими насекомыми. Автореф. дис. … канд. биол. наук. М.: Лаборатория лесоведения АН СССР. 24 с.
Шевырев И.Я., 1891. О вредных насекомых степных лесничеств в 1889 г. СПб.: Типография В. Демакова. 139 с.
Ханисламов М.Г., Гирфанова Л.Н., Яфаева З.Ш., Степанова Р.К., 1962. Условия формирования резерваций и нарастания численности непарного шелкопряда в Башкирии // Исследование очагов вредителей леса в Башкирии. Уфа: БФ АН СССР. С. 32−66.
Эпова В.И., Плешанов А.С., 1995. Зоны вредоносности насекомых-филлофагов Азиатской России. Новосибирск: Наука. Сибирская издательская фирма РАН. 47 с.
Pedunculate Oak (Quercus robur) Distribution Map., 2020. European Forest Genetic Resources Programme [Электронный ресурс]. Режим доступа: http:// www.euforgen.org/distribution-maps. Дата обновления: 5.02.2020
Ponomarev V.I., Napalkova V.V., Klobukov G.I., 2019. Forecasting the phenology of gypsy moth development and the time frames for the pest control based on sum of effective temperatures according to latitudinal origination of population // Modern Synthetic Methodologies for Creating Drugs and Functional Materials (MOSM2018). Proceedings of the II International Conference. C. 040042.
Wallin H., Nylander U., Kvamme T., 2010. Two sibling species of Leiopus Audinet-Serville, 1835 (Coleoptera: Cerambycidae) from Europe: L. nebulosus (Linnaeus, 1758) and L. linnei sp. nov. // Zootaxa. P. 31−45.
Дополнительные материалы отсутствуют.
Инструменты
Зоологический журнал