

Зоологический журнал, 2021, T. 100, № 6, стр. 603-617
Зоопланктон Цимлянского водохранилища и канала Волга–Дон
В. И. Лазарева a, *, Р. З. Сабитова a
a Институт биологии внутренних вод имени И.Д. Папанина РАН
152742 Ярославская обл., Некоузский р-н, пос. Борок, Россия
* E-mail: lazareva_v57@mail.ru
Поступила в редакцию 02.03.2020
После доработки 24.05.2020
Принята к публикации 03.06.2020
Аннотация
В августе–сентябре 2018 г. изучен зоопланктон Цимлянского водохранилища и водоемов Волго-Донского судоходного канала. Для Цимлянского водохранилища выявлено 15 видов, которые не были отмечены в ранее опубликованных работах. Впервые приведен список (48 видов) зоопланктона Волго-Донского канала. Установлено, что 60% численности и 70% биомассы зоопланктона Цимлянского водохранилища формируют Copepoda. Наиболее многочисленны понто-каспийская Heterocope caspia и вселенцы Calanipeda aquaedulcis, Thermocyclops taihokuensis и Acanthocyclops americanus. На основе анализа публикаций 1952–2018 гг. показано, что во второй половине 1970-х годов произошла замена большей части изначальной донской группировки доминантов зоопланктона солоноватоводными видами. К 2018 г. массовые виды Copepoda сменились полностью, среди Cladocera и Rotifera сохранились 4–6 видов, обычных в р. Дон до зарегулирования. Выявлено увеличение более чем вдвое (до 2.1 ± 0.5 г/м3) биомассы сообщества в 2015–2018 гг. по сравнению с 40-летним периодом до 2013 г. Показано, что биомасса зоопланктона в канале Волга–Дон (0.9 ± 0.4 г/м3) в 2 раза ниже, чем в Цимлянском водохранилище. Численность зоопланктона водоемов канала определяют Rotifera и Crustacea (по 50%), а биомассу – в основном Cladocera (49%). Наиболее многочисленны обычные для донского планктона виды Bosmina longirostris, Chydorus sphaericus, Bosmina cf. coregoni и Daphnia cucullata, понто-каспийская Podonevadne trigona развиваeтся в массе только в Варваровском водохранилище. Копеподы-вселенцы Thermocyclops taihokuensis, Heterocope caspia, Calanipeda aquaedulcis и Eurytemora caspica образуют 12–23% численности ракообразных. Обсуждаются динамика структуры и обилия зоопланктона Цимлянского водохранилища в 1952–2018 гг., а также расселение и значение в планктоне понто-каспийских ракообразных и вселенцев из южных регионов.
В среднем течении р. Дон около 200 км от устья (координаты центра 47°50ʹ с.ш., 42°50ʹ в.д.) существует единственное на этой реке Цимлянское водохранилище (рис. 1). Водохранилище заполнено в 1952–1953 гг., при нормальном подпорном уровне (НПУ, 36 м БС11) его площадь 2700 км2, средняя глубина 8.8 м, максимальная 30 м, скорость водобмена 1.05 год–1 (Авакян и др., 1987). Чаша водохранилища образована котловиной с тремя расширениями, приуроченными к устьевым областям рек Чир, Аксай Курмоярский и Цимла, выделяют четыре плеса: Верхний, Чирской, Потемкинский и Приплотинный (Гидрометеорологический режим…, 1977). Водохранилище характеризуется большой площадью мелководий, при НПУ участки с глубиной менее 5 м составляют 29% акватории (Вехов и др., 2014).
Рис. 1.
Цимлянское водохранилище и Волго-Донской канал на карте Яндекс. Водохранилища Волго-Донского канала: 1 – Карповское, 2 – Бреславское, 3 – Варваровское; участки Цимлянского водохранилища: I – Верхний, II – Чирской, III – Потемкинский, IV – Приплотинный.
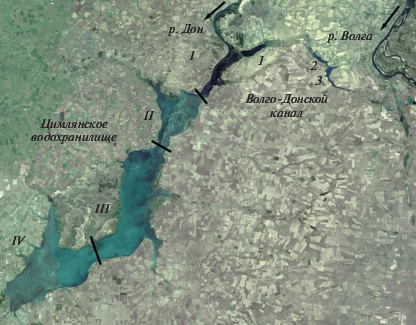
В верхнем участке Цимлянского водохранилища ниже г. Калач расположен вход в судоходный канал Волга–Дон (рис. 1), построенный в 1952 г. Со стороны Волги канал начинается ниже г. Волгоград, по долине р. Сарпа (приток р. Волга) он проходит к водоразделу рек Волги и Дона, затем выходит в долину р. Червлёная (приток р. Дон), на которой системой плотин и шлюзов образованы два небольших водохранилища Варваровское (48°34ʹ с.ш., 44°10′ в.д., 26 км2) и Бреславское (48°37′ с.ш., 44°06′ в.д., 15 км2), далее канал выходит в долину р. Карповка (приток р. Дон) и Карповское водохранилище (48°38′ с.ш., 43°40′ в.д., 42 км2) (Моруков, 2006). Канал питается донской водой, которая подается тремя насосными станциями на водораздел в Варваровское водохранилище, откуда самотеком стекает в Волгу. Карповское и Бреславское водохранилища сравнительно мелководны (средняя глубина менее 4 м), в Варваровском средняя глубина (7.8 м) близка к таковой в Цимлянском водохранилище (Авакян и др., 1987).
Цимлянское водохранилище по уровню развития фитопланктона (до 50 мг/л) характеризуют как гипертрофное (Цимлянское водохранилище…, 2011; Вехов и др., 2014; Никаноров, Хоружая, 2014). Водохранилища канала Волга–Дон также относятся к высокопродуктивным экосистемам, однако сведения об их современном трофическом статусе в литературе отсутствуют.
Зоопланктон Цимлянского водохранилища исследован с первых лет существования водоема (1952–1957 гг.) до настоящего времени (2008–2018 гг.) (Лившиц, 1954; Дзюбан, 1958; Шейнин, 1960; Кафтанникова, 1965; Гламазда, 1971, 1974, 1982; Шевлякова, 2002; Свистунова, Саяпин, 2010; Вехов и др., 2014; Предварительные материалы…, 2018; Голоколенова и др., 2019). Наиболее подробно изменения состава, структуры, динамика обилия и продуктивности сообщества водохранилища проанализированы по материалам 1950–1970-х годов (Дзюбан, 1958; Шейнин, 1960; Кафтанникова, 1965; Гламазда, 1971, 1974). Вместе с тем, тенденции в развитии зоопланктона в течение последних 40 лет изучены явно недостаточно, сведения о зоопланктоне водоемов Волго-Донского канала фактически отсутствуют.
Целью работы было более углубленно изучить состав, структуру и распределение обилия летнего зоопланктона Цимлянского водохранилища и связанных с ним водоемов Волго-Донского канала, а также проанализировать динамику сообщества за весь период существования водохранилища.
МАТЕРИАЛ И МЕТОДИКА
Материал собирали в комплексной экспедиции Института биологии внутренних вод им. И.Д. Папанина РАН в конце августа–начале сентября 2018 г. с экспедиционного судна “Академик Топчиев” и лодки. В канале Волга–Дон обследовали только участки по судовому ходу (глубина 5–12 м): два – в Варваровском и по одному – в Бреславском и Карповском водохранилищах. В Цимлянском водохранилище обследовано 11 участков в пелагиали (глубина более 5 м) и пять в прибрежье, в основном в устьевой области притоков (глубина 1.5–4 м). Одновременно с отбором проб ручными зондами YSI ProODO и YSI-85 (YSI, Inc., USA) измеряли глубину, температуру воды, электропроводность и содержание растворенного кислорода.
Зоопланктон учитывали в тотальных пробах, которые отбирали сетью Джеди с диаметром входного отверстия 12 см и ситом с диагональю ячеи 105 мкм, облавливали весь столб воды от дна до поверхности водоема. Сборы фиксировали 4%-формалином и просматривали в лаборатории под стереомикроскопом StereoDiscovery-12 (Carl Zeiss, Jena). Ракообразных и коловраток идентифицировали с использованием следующих работ: Кутикова (1970); Монченко (1974); Мордухай-Болтовской, Ривьер (1987); Боруцкий и др. (1991); Определитель… (2010); Guo (1999); Monchenko (2008); Sukhikh, Alekseev (2013).
Видовое богатство зоопланктона оценивали по числу видов в пробе на каждой станции (видовая плотность) и по общему числу видов в списке каждого водоема. Доминантные виды выделяли по их относительной численности, отдельно в таксоценозах ракообразных (более 10% общего количества рачков) и коловраток (более 10% общего количества коловраток).
Сходство структуры доминантных комплексов определяли с помощью индекса Чекановского-Съеренсена для количественных данных (ICzS) (Песенко, 1982):
где pi – доля вида в общей численности зоопланктона.Прочие статистические показатели (средние число видов в пробе, численность, биомасса и ошибка среднего) рассчитывали с использованием соответствующих процедур пакета STATISTICA for Windows, v. 10 (StatSoft Russia).
РЕЗУЛЬТАТЫ
Условия обитания. Воды изученных водоемов характеризовались сравнительно высоким (более 400 мг/л) содержанием солей (по электропроводности) (табл. 1). Наибольшие значения регистрировали в канале Волга–Дон (до 730 мг/л), максимум наблюдался в Варваровском водохранилище. Прогрев воды в Цимлянском водохранилище был на 1.2°С ниже нормы, а в канале Волга–Дон – близок к норме. Для Цимлянского водохранилища среднемноголетняя (1953–1972 гг.) температура поверхности воды (норма) в августе составляет 23.2°С (Гидрометеорологический режим…, 1977). Температура придонной воды повсеместно была высокой, близкой к таковой у поверхности. Содержание растворенного кислорода (О2) в водохранилищах канала Волга–Дон и большинстве участков Цимлянского водохранилища превышало 70% насыщения во всем столбе воды и не лимитировало развитие гидробионтов. Низкие значения (ниже 5 мг/л) наблюдались на границе вода/ил локально по руслу р. Дон в Верхнем, Чирском и Потемкинском участках. Очень низкая концентрация О2 (4.4–5.2 мг/л, 49–52%) отмечена в устьевой области р. Аксеней (глубина 2.5 м), сильный запах сероводорода указывал на наличие анаэробной зоны.
Таблица 1.
Гидрофизические характеристики условий обитания зоопланктона в Цимлянском водохранилище и канале Волга–Дон в 2018 г.
| Водоем | Глубина, м | Температура воды, °С | Электропроводность, мкСм/см* | Содержание растворенного О2 | |
|---|---|---|---|---|---|
| поверхность | у дна | ||||
| Цимлянское водохранилище | 1.5–18 | 22.0 ± 0.2°С | 610–810 | $\frac{{6.4{\kern 1pt} - {\kern 1pt} 10.0}}{{75{\kern 1pt} - {\kern 1pt} 114}}$ | $\frac{{4.4{\kern 1pt} - {\kern 1pt} 9.2}}{{49{\kern 1pt} - {\kern 1pt} 84}}$ |
| Канал Волга–Дон | 5–12 | 22.5 ± 0.2°С | 750–1130 | $\frac{{7.9{\kern 1pt} - {\kern 1pt} 9.5}}{{91{\kern 1pt} - {\kern 1pt} 110}}$ | $\frac{{6.3{\kern 1pt} - {\kern 1pt} 7.6}}{{72{\kern 1pt} - {\kern 1pt} 88}}$ |
Состав и структура зоопланктона. В зоопланктоне исследованных водоемов обнаружено 64 вида (15 Cladocera, 12 Copepoda и 37 Rotifera). Из них в Цимлянском водохранилище – 56 видов (12 Cladocera, 9 Copepoda и 35 Rotifera), в канале Волга-Дон – 48 видов (14 Cladocera, 10 Copepoda и 24 Rotifera) (табл. 2). В Цимлянском водохранилище наибольшее количество видов (38–42) наблюдалось в Верхнем и Потемкинском участках (табл. 3). В среднем регистрировали от 19 до 28 видов в пробе, максимум 30–31 вид отмечали в Верхнем участке Цимлянского водохранилища ниже г. Калач, а также в Карповском и Варваровском водохранилищах. Большую часть из них (40–60%) составляли коловратки. Состав зоопланктона Цимлянского водохранилища и водоемов канала Волга–Дон отличался очень высоким сходством (79%), что можно объяснить постоянным поступлением донских видов в канал с водой водохранилища.
Таблица 2.
Состав и встречаемость зоопланктона Цимлянского водохранилища и Волго-Донского канала в августе‒сентябре 2018 г.
| Таксон | Водохранилище | Канал |
|---|---|---|
| Crustacea | ||
| Diaphanosoma orghidani Negrea 1982 | +++ | +++ |
| Daphnia (Daphnia) galeata Sars 1864 | + | ++ |
| Daphnia (D.) cucullata Sars 1862 | ‒ | ++ |
| Bosmina (Bosmina) longirostris (O.F. Müller 1785) | ++ | +++ |
| Bosmina (Eubosmina) cf. coregoni Baird 1857 | + | ++ |
| Moina micrura Kurz 1874 | ++ | ++ |
| Chydorus sphaericus (O.F. Müller 1785) | ++ | ++ |
| Chydorus gibbus Sars 1891* | + | + |
| Alona quadrangularis (O.F. Müller 1785) | ‒ | + |
| Leydigia leydigii (Schoedler 1863) | ‒ | + |
| Cornigerius maeoticus maeoticus (Pengo 1879) | + | +++ |
| Podonevadne trigona ovum (Zernov 1901) | + | ++ |
| Cercopagis pengoi (Ostroumov 1891) | + | +++ |
| Leptodora kindtii (Focke 1844) | ++ | +++ |
| Сyclops vicinus Uljanin 1875 | + | ‒ |
| Acanthocyclops americanus (Marshall 1893)* | +++ | ++ |
| Thermocyclops crassus (Fisсher 1853) | + | + |
| Thermocyclops taihokuensis (Harada 1931) | ++ | ++ |
| Mesocyclops leuckarti (Claus 1857) | + | ++ |
| Halicyclops neglectus Kiefer 1935* | + | ‒ |
| Diacyclops bicuspidatus (Claus 1857) | + | + |
| Diacyclops crassicaudis (Sars 1863) | ‒ | + |
| Eucyclops serrulatus (Fisсher 1851) | ‒ | + |
| Heterocope caspia Sars 1897 | +++ | +++ |
| Calanipeda aquaedulcis Kritschagin 1873 | +++ | +++ |
| Eurytemora caspica Sukhikh et Alekseev 2013* | +++ | +++ |
| Rotifera | ||
| Trichocerca (Trichocerca) capucina (Wierzejski et Zacharias 1893) | + | ++ |
| Trichocerca (T.) cylindrica (Imhof 1891) | ++ | ‒ |
| Trichocerca (T.) pusilla (Lauterborn 1898) | + | ‒ |
| Trichocerca (Diurella) similis (Wierzejski 1893) | ++ | + |
| Polyarthra luminosa Kutikova 1962* | +++ | +++ |
| Polyarthra major Bruckhardt 1900 | +++ | ++ |
| Polyarthra longiremis Carlin 1943* | + | ++ |
| Polyarthra minor Voigt 1904* | + | + |
| Polyarthra euryptera Wierzejski 1891 | ++ | ‒ |
| Synchaeta pectinata Ehrenberg 1832 | ++ | ++ |
| Synchaeta tremula (O.F. Müller 1786)* | + | ‒ |
| Ascomorpha ecaudis Perty 1850 | ‒ | + |
| Bipalpus hudsoni (Imhof 1891) | + | + |
| Asplanchna priodonta Gosse 1850 | ++ | ++ |
| Asplanchna henrietta Langhaus 1906* | + | + |
| Lecane (Lecane) luna (O.F. Müller 1776) | + | + |
| Lecane (L.) ungulata Gosse 1887* | + | ‒ |
| Lecane (Monostyla) lunaris Ehrenberg 1832 | + | ‒ |
| Epiphanes macroura (Barrois et Daday 1894)* | + | + |
| Euchlanis dilatata dilatata Ehrenberg 1832 | ++ | ++ |
| Euchlanis dilatata lucksiana (Hauer 1939) | ++ | ++ |
| Keratella cochlearis (Gosse 1851) | +++ | ++ |
| Keratella tropica (Apstein 1907)* | + | ‒ |
| Keratella quadrata (O.F. Müller 1786) | ++ | + |
| Brachionus calyciflorus Pallas 1776 | ++ | ++ |
| Brachionus diversicornis (Daday 1883) | ++ | ++ |
| Brachionus angularis Gosse 1851 | + | ‒ |
| Brachionus budapestinensis Daday 1885 | ‒ | + |
| Brachionus falcatus Zacharias 1898* | + | + |
| Kellicottia longispina (Kellicott 1879) | + | ‒ |
| Pompholyx sulcata Hudson 1885 | + | ‒ |
| Filinia longiseta (Ehrenberg 1834) | + | ++ |
| Filinia terminalis (Plate 1886 ) | + | ‒ |
| Collotheca pelagica (Rousselet 1893)* | ++ | + |
| Conochilus unicornis Rousselet 1892 | ++ | ‒ |
| Conochiloides coenobasis Skorikov 1914* | + | ‒ |
| Hexarthra mira (Hudson 1871) | + | + |
| Всего | 56 | 48 |
Таблица 3.
Видовое богатство зоопланктона Цимлянского водохранилища и канала Волга–Дон в сентябре 2018 г.
| Участок | Среднее число видов в пробе | Число видов в списке | |||
|---|---|---|---|---|---|
| Cladocera | Copepoda | Rotifera | общее | ||
| Верхний | 6 ± 1 | 5 ± 1 | 12 ± 2 | 23 ± 3 | 38 |
| Чирской | 5 ± 1 | 7 ± 1 | 11 ± 1 | 23 ± 1 | 31 |
| Потемкинский | 6 ± 1 | 5 ± 1 | 14 ± 1 | 25 ± 1 | 42 |
| Приплотинный | 3 ± 1 | 5 ± 1 | 11 ± 3 | 19 ± 2 | 25 |
| Все водохранилище | 5 ± 1 | 6 ± 1 | 12 ± 2 | 23 ± 2 | 56 |
| Канал | 10 ± 1 | 6 ± 1 | 12 ± 2 | 28 ± 2 | 48 |
Выявлены 4 вида ракообразных (Chydorus gibbus, Acanthocyclops americanus, Halicyclops neglectus и Eurytemora caspica) и 11 видов коловраток (см. табл. 2), не указанных ранее для Цимлянского водохранилища (см. Лившиц, 1954; Дзюбан, 1958; Шейнин, 1960; Кафтанникова, 1965; Гламазда, 1971, 1974, 1982; Шевлякова, 2002; Свистунова, Саяпин, 2010; Вехов и др., 2014; Предварительные материалы…, 2018; Голоколенова и др., 2019). Нами не найдены ракообразные Eurytemora affinis (Poppe, 1880), Diaphanosoma brachyurum (Lièvin, 1848) и коловратка Euchlanis incisa Carlin,1939, которые отмечены как массовые в 2015–2018 гг. (Предварительные материалы…, 2018; Голоколенова и др., 2019). Копеподу Eurytemora caspica ранее определяли, по-видимому, как E. affinis, а Diaphanosoma brachyurum смешивали с D. оrghidani. Коловратка Euchlanis incisa не обнаружена, вероятно, из-за небольшого числа точек отбора проб в прибрежье.
Доминантный комплекс зоопланктона Цимлянского водохранилища включал 13 видов, по 5–8 для каждого из четырех участков (табл. 4). В августе–сентябре в Верхнем и Чирском участках среди ракообразных преобладали Cyclopoida: Thermocyclops taihokuensis и Acanthocyclops americanus формировали 70–90% численности группы (Ncr). В Потемкинском и Приплотинном участках доминировали Calanoida: Heterocope caspia и Calanipeda aquaedulcis образовывали до 70% Ncr. Сходным образом изменялся состав массовых видов коловраток. В двух северных участках водохранилища были многочисленны виды рода Polyarthra (45–80% численности коловраток), в обоих южных – Keratella quadrata, Euchlanis dilatata и Conochilus unicornis (50–70%). Наиболее сходной была структура доминантного комплекса Верхнего и Чирского участков водохранилища (46% коловратки, 72% ракообразные). Остальные участки сильно отличались друг от друга (индекс сходства менее 30%), массовые виды коловраток Верхнего и Приплотинного участков были совершенно различны (индекс сходства 0%).
Таблица 4.
Доминантные виды ракообразных и коловраток различных участков Цимлянского водохранилища в августе–сентябре 2018 г.
| Таксон | Участок | |||||||
|---|---|---|---|---|---|---|---|---|
| Верхний | Чирской | Потемкинский | Приплотинный | |||||
| Nsp | %Ngr | Nsp | %Ngr | Nsp | %Ngr | Nsp | %Ngr | |
| Crustacea | ||||||||
| Moina micrura | 81 | 25 | – | – | – | – | – | – |
| Thermocyclops taihokuensis | 192 | 59 | 137 | 76 | 11 | 13 | – | – |
| Acanthocyclops americanus | 46 | 14 | 24 | 13 | – | – | 22 | 13 |
| Heterocope caspia | – | – | – | – | 50 | 57 | 20 | 12 |
| Calanipeda aquaedulcis | – | – | – | – | 14 | 16 | 47 | 28 |
| Rotifera | ||||||||
| Brachionus angularis | 11 | 16 | – | – | – | – | – | – |
| Polyarthra luminosa | 31 | 45 | 11 | 24 | – | – | – | – |
| Trichocerca similis | – | – | 6 | 13 | – | – | – | – |
| Polyarthra major | 24 | 35 | 10 | 22 | 12 | 9 | – | – |
| Keratella quadrata | – | – | 8 | 18 | 24 | 18 | 6 | 25 |
| Conochilus unicornis | – | – | – | – | 18 | 13 | 11 | 46 |
| Euchlanis dilatata | – | – | – | – | 22 | 16 | – | – |
| Asplanchna priodonta | – | – | – | – | 14 | 10 | – | – |
Примечания. Здесь и в табл. 5 Nsp – средняя численность вида, тыс. экз./м3, %Ngr – вклад (%) в численность ракообразных (Ncr) или коловраток (Nrot). Прочерк – вид немногочислен (<10% Ngr).
Все три водохранилища Волго-Донского канала характеризовались оригинальным составом доминантов, индекс сходства менее 20%. Всего выявлено 17 массовых видов по 7–8 в каждом из трех водохранилищ (табл. 5). В Варваровском водохранилище на фоне сравнительно низкой численности зоопланктона (52–58 тыс. экз./м3) более 70% обилия ракообразных формировали кладоцеры Bosmina longirostris, Podonevadne trigona и копепода Calanipeda aquaedulcis. Среди коловраток наиболее многочисленной была Asplanchna priodonta. В Бреславском водохранилище доминировали (60% Ncr) копеподы Thermocyclops taihokuensis и Heterocope caspia, а также кладоцера Bosmina cf. coregoni. Более 90% обилия коловраток формировала Keratella quadrata. В Карповском водохранилище более 60% Ncr образовывали кладоцера Chydorus sphaericus и копеподы Thermocyclops taihokuensis, Eurytemora caspica, а среди коловраток наибольшую численность (56% общей) формировали Polyarthra longiremis и Trichocerca capucina. Общность структуры доминантных комплексов зоопланктона водоемов канала Волга–Дон и участков Цимлянского водохранилища была крайне низкой: сходство состава массовых видов коловраток составляло менее 25%, ракообразных – менее 35%.
Таблица 5.
Доминантные виды ракообразных и коловраток водохранилищ Волго-Донского канала в августе–сентябре 2018 г.
| Таксон | Водохранилище | |||||
|---|---|---|---|---|---|---|
| Варваровское | Бреславское | Карповское | ||||
| Nsp | %Ngr | Nsp | %Ngr | Nsp | %Ngr | |
| Crustacea | ||||||
| Bosmina longirostris | 9 | 33 | – | – | – | – |
| Podonevadne trigona | 5 | 20 | – | – | – | – |
| Calanipeda aquaedulcis | 5 | 18 | – | – | – | – |
| Eurytemora caspica | 3 | 12 | – | – | 18 | 14 |
| Chydorus sphaericus | – | – | – | – | 37 | 29 |
| Thermocyclops taihokuensis | – | – | 24 | 17 | 29 | 23 |
| Bosmina cf. coregoni | – | – | 32 | 23 | – | – |
| Heterocope caspia | – | – | 29 | 20 | – | – |
| Daphnia cucullata | – | – | 19 | 13 | – | – |
| Diaphanosoma orghidani | – | – | 13 | 10 | – | – |
| Rotifera | ||||||
| Keratella quadrata | – | – | 138 | 92 | – | – |
| Polyarthra longiremis | – | – | – | – | 62 | 38 |
| Trichocerca capucina | – | – | – | – | 29 | 18 |
| Polyarthra major | – | – | – | – | 18 | 11 |
| Asplanchna priodonta | 7 | 25 | – | – | 17 | 10 |
| Synchaeta pectinata | 4 | 14 | – | – | 14 | 10 |
| Filinia longiseta | 4 | 13 | – | – | – | – |
В исследованных водоемах обнаружены ракообразные, происходящие из понто-каспийского (Heterocope caspia, Eurytemora caspica, Cercopagis pengoi, Cornigerius maeoticus и Podonevadne trigona), средиземноморского (Calanipeda aquaedulcis) и восточно-азиатского (Thermocyclops taihokuensis) фаунистических комплексов. Максимального обилия (более 600 тыс. экз./м3) достигал вид T. taihokuensis, в Верхнем и Чирском участках Цимлянского водохранилища он образовывал более 60% Ncr (табл. 4). В Потемкинском и Приплотинном участках водохранилища 40–70% численности и 85–95% биомассы ракообразных формировали Heterocope caspia (более 128 тыс. экз./м3), Calanipeda aquaedulcis (более 90 тыс. экз./м3) и Eurytemora caspica (до 18 тыс. экз./м3). Все четыре вида копепод обитали в водоемах Волго-Донского канала, но здесь их количество было ниже по сравнению с Цимлянским водохранилищем (табл. 5). Наибольшей численности (18–29 тыс. экз./м3) достигали Heterocope caspia, Thermocyclops taihokuensis и Eurytemora caspica.
Понто-каспийские кладоцеры Cercopagis pengoi, Cornigerius maeoticus и Podonevadne trigona были более многочисленны в водоемах канала Волга–Дон по сравнению с Цимлянским водохранилищем. В Верхнем и Потемкинском участках водохранилища они обнаружены единично (менее 100 экз./м3), в двух других – отсутствовали. Наибольшей численности (более 8 тыс. экз./м3) в Карповском и Варваровском водохранилищах достигала P. trigona, в последнем вид образовывал 20% количества ракообразных (табл. 5). Вклад этих маленьких рачков (длина тела 0.25 мм, высота раковинки 0.5 мм) в биомассу ракообразных достигал 26%. Обилие Cornigerius maeoticus было наибольшим (более 2 тыс. экз./м3) в Карповском водохранилище, а Cercopagis pengoi (300–600 экз./м3) – в Карповском и Бреславском водохранилищах.
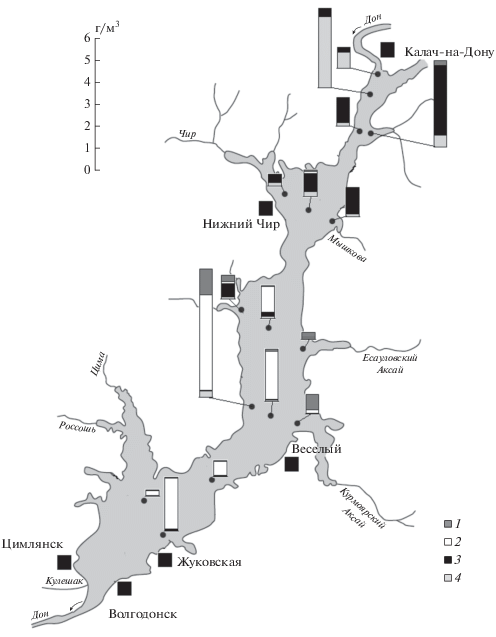
Численность и биомасса. В сроки наблюдений общее количество зоопланктона в Цимлянском водохранилище (247 ± 53 тыс. экз./м3 и 1.8 ± 0.4 г/м3) было в 1.4–2 раза выше, чем в канале Волга–Дон (173 ± 68 тыс. экз./м3 и 0.9 ± 0.4 г/м3). Вклад основных групп (Cladocera, Cyclopoida, Calanoida и Rotifera) в численность и биомассу сообщества водоемов составлял 24, 10, 13, 53% и 49, 12, 17, 24% соответственно. В Цимлянском водохранилище по численности преобладали Cyclopoida (40%) и Rotifera (32%), Cladocera были сравнительно многочисленны (22%) только в Верхнем участке водоема, а каляноидные копеподы – в Потемкинском (30%) и Приплотинном (58%) участках. Численность зоопланктона снижалась от Верхнего участка (395 ± 179 тыс. экз./м3) к Приплотинному (121 ± 32 тыс. экз./м3). Основу биомассы зоопланктона водохранилища формировали ракообразные: Calanoida – 40%, Cyclopoida – 29% и Cladocera – 20%. Максимальные ее значения регистрировали в Верхнем (до 4 г/м3) и Потемкинском (до 6 г/м3) участках, минимальные (менее 1.5 г/м3) – в Чирском участке и на большинстве станций у плотины Цимлянской ГЭС (рис. 2).
Рис. 2.
Распределение биомассы зоопланктона по акватории Цимлянского водохранилища летом 2018 г.: 1 – Rotifera, 2 – Calanoida, 3 – Cyclopoida, 4 – Cladocera.
Количество мероплактона (велигеры моллюсков рода Dreissena) в канале Волга–Дон (28 ± ± 18 тыс. экз./м3) было в 4–6 раз выше по сравнению с Цимлянским водохранилищем (6 ± 2 тыс. экз./м3). Особенно много велигеров (свыше 77 тыс. экз./м3) регистрировали в Бреславском водохранилище. На большей части акватории Цимлянского водохранилища их количество не превышало 10 тыс. экз./м3. Вклад меропланктона в суммарную (зоопланктон + + меропланктон) биомассу сообщества невелик: от 1% общей биомассы в Цимлянском водохранилище до 7% в канале Волга–Дон.
Многолетняя динамика. За более 65 лет (1952–2018 гг.) существования Цимлянского водохранилища отмечены существенные изменения структуры зоопланктона и его количества. Кардинальная смена структуры сообщества произошла во второй половине 1970-х годов и наиболее четко прослеживалась в группе ракообразных (рис. 3а, 3б). В это время снизилась численность обычных для среднего течения р. Дон представителей родов Daphnia, Eudiaptomus, Acanthocyclops и Mesocyclops, многочисленными стали понто-каспийские и средиземноморские виды родов Cornigerius, Podonevadne, Heterocope, Eurytemora и Calanipeda. В группе коловраток (рис. 3в) в середине 1950-х годов появились два вида (Brachionus diversicornis и Euchlanis dilatata), многочисленные до настоящего времени.
Рис. 3.
Смены доминантов зоопланктона Цимлянского водохранилища за время его существования (1952‒2018 гг.). а – Cladocera, б – Copepoda, в – Rotifera. 1 – Diaphanosoma orghidani, 2 – Podonevadne trigona, 3 – Cornigerius maeoticus, 4 – Bythotrephes cf. longimanus и В. cederstroemii, 5 – Bosmina cf. coregoni, 6 – Daphnia galeata, 7 – D. cucullata, 8 – Acanthocyclops americanus, 9 – Thermocyclops taihokuensis, 10 – T. crassus, 11 – Eurytemora gr. affinis, 12 – Calanipeda aquaedulcis, 13 – Eurytemora velox, 14 – Heterocope caspia, 15 – Acanthocyclops vernalis, 16 – Thermocyclops oithonoides, 17 – Eudiaptomus gracilis, 18 – Mesocyclops leuckarti, 19 – Eudiaptomus graciloides, 20 – Trichocerca similis, 21 – Polyarthra luminosa, 22 – Conochilus unicornis, 23 – Euchlanis dilatata, 24 – Brachionus diversicornis, 25 – Brachionus angularis, 26 – Polyarthra major. Источники: 1952 г. – (Лившиц, 1954); 1953–1954 гг. – (Дзюбан, 1958); 1955–1962 гг. – (Кафтанникова, 1965; Шейнин, 1960); 1966–1978 гг. – (Гламазда, 1971, 1974, 1982); 1998–2001 гг. – (Шевлякова, 2002), 2008–2013 гг. – (Свистунова, Саяпин, 2010; Вехов и др., 2014), 2015‒2018 гг. – (Предварительные материалы…, 2018; Голоколенова и др., 2019; данные авторов).
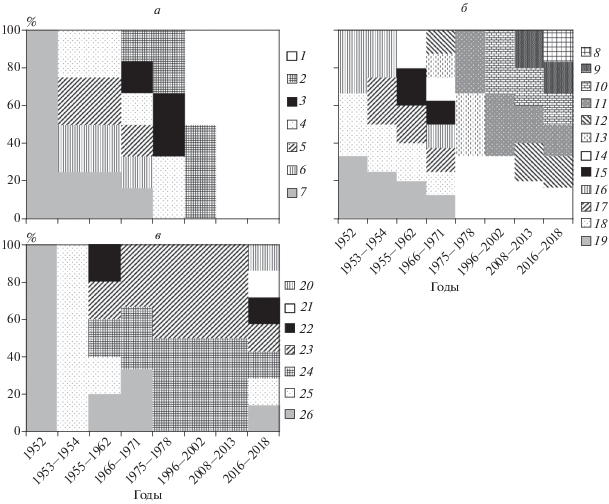
В последующем (1996–2002 гг.) происходило пополнение группировки доминантов теплолюбивыми видами (Diaphanosoma orghidani и Thermocyclops crassus), характерными для эвтрофных водоемов. В числе массовых видов перестали упоминать Bythotrephes cf. longimanus и B. cederstroemii, Cornigerius maeoticus и Eurytemora velox (Lilljeborg, 1853). В 2008–2013 гг. стала малочисленной кладоцера Podonevadne trigona. В 2012 г. в водохранилище появилась копепода Thermocyclops taihokuensis (Вехов и др., 2014), к настоящему времени ставшая самым многочисленным видом в группе Cyclopoida. В 2018 г. вторым по численности после T. taihokuensis стал вид Acanthocyclops americanus, в 1955–1970 гг. отмечали другой вид этого рода A. vernalis (Fischer, 1853). Изменился состав доминантных видов коловраток: в 2018 г. впервые отмечены как массовые Polyarthra luminosa и Trichocerca similis. Также стали многочисленны виды Conochilus unicornis, Polyarthra major (ранее P. gr. “trigla”) и Brachionus angularis, которые длительное время с 1970-х годов не входили в состав доминантов.
Отметим, что среди Cladocera и Rotifera есть виды, которые с 1950-х годов постоянно (кладоцеры Moina micrura, Bosmina longirostris) или с промежутками в несколько лет (кладоцеры Chydorus sphaericus и коловратки Asplanchna priodonta, Keratella quadrata, K. cochlearis, Brachionus calyciflorus) регистрировались как массовые. В то же время, состав доминантных видов копепод к 2018 г. сменился полностью (рис. 3б).
ОБСУЖДЕНИЕ
В зоопланктоне Цимлянского водохранилища в первые три года (1952–1954 гг.) его существования указывали 40–80 видов (Лившиц, 1954; Дзюбан, 1958). В последущий период (1955–1971 гг.) при увеличении объема наблюдений список расширился до 120–160 видов (Кафтанникова, 1965; Гламазда, 1974). Начиная с середины 1990-х годов, объем гидробиологических наблюдений на водохранилище сократился до коротких и преимущественно летних маршрутных обследований 1–2 раза за сезон. Это привело к уменьшению количества обнаруживаемых видов зоопланктона до 60–100 (Шевлякова, 2002; Голоколенова и др., 2019), если считать с подвидами – до 160 (Вехов и др., 2014). По результатам короткой (трое суток) экспедиции 2018 г. нами составлен достаточно большой список (56 видов) зоопланктона, в том числе выявлено 15 форм, отсутствующих в ранее опубликованных работах. Состав зоопланктона водохранилищ Волго-Донского канала (48 видов) представлен в настоящей работе впервые.
Со второй половины ХХ в. наблюдается увеличение общей концентрации солей, доли хлоридов и сульфатов в воде Цимлянского водохранилища, к началу 2010-х годов уровень минерализации вырос в 1.3–1.6 раза (Цимлянское водохранилище…, 2011). Максимальные значения в межень варьируют в пределах 650–1010 мг/л (Никаноров, Хоружая, 2014). Сопоставимые данные (400–530 мг/л) получены нами в конце лета 2018 г.
Повышенная минерализация воды Цимлянского водохранилища и водоемов канала Волга–Дон (более 700 мг/л) способствует распространению в них солоноватоводных ракообразных. В 1950-х годах в р. Дон ниже плотины Цимлянской ГЭС, включая дельту, стали регулярно находить Calanipeda aquaedulcis (Шейнин, 1960). С 1959 г. этот вид обитает в Цимлянском водохранилище (Гламазда, 1971). Представители рода Eurytemora (в основном E. velox) и понто-каспийская Heterocope caspia были обычны в нижнем течении р. Дон еще до зарегулирования (Шейнин, 1960). В Цимлянском водохранилище их регистрируют с начала его заполнения (с 1952 г., Лившиц, 1954), но многочисленными они стали лишь в 1960–1961 гг. С этого времени H. caspia стала вытеснять обычных для р. Дон копепод рода Eudiaptomus (Кафтанникова, 1965; Гламазда, 1971). В 1960-х годах по акватории водоема начали расселяться и достигли массового развития понто-каспийские солоноватоводные кладоцеры (Cornigerius maeoticus (1961 г.), Podonevadne trigona ovum (1966 г.) и Cercopagis pengoi (1970 г.) (Шейнин, 1964; Гламазда, 1969, 1971а). До зарегулирования р. Дон эти виды регистрировали только в восточной части Таганрогского залива Азовского моря (Шейнин, 1960). В 2018 г. все три вида понто-каспийских кладоцер в Цимлянском водохранилище были малочисленны, тогда как копеподы Heterocope caspia и Calanipeda aquaedulcis доминировали в планктоне нижней части водоема.
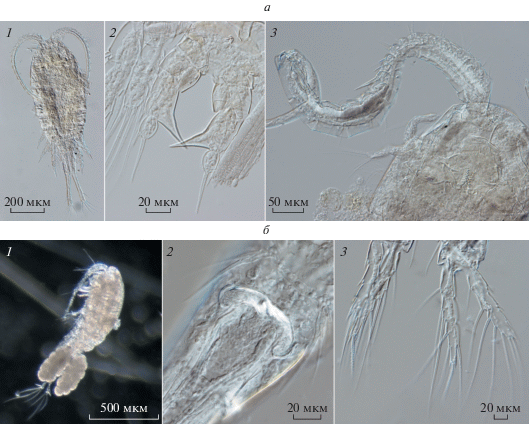
Для Цимлянского водохранилища с середины 1970-х годов как массовый вид указывали Eurytemora cf. affinis (Гламазда, 1982; Шевлякова, 2002; Свистунова, Саяпин, 2010; Вехов и др., 2014), который был многочислен также в Азовском море и р. Дон ниже Цимлянского водохранилища (Селифонова, 2013; Свистунова и др., 2014). Представителя этого рода, вселившегося в 1980-х годах в водохранилища Средней и Нижней Волги, также идентифицировали как E. cf. affinis (Тимохина, 2000). По материалам из дельты Волги и Северного Каспия в 2013 г. из комплекса криптических видов E. affinis s. l. выделен новый вид E. caspica (Sukhikh, Alekseev, 2013). На основании морфологических и молекулярных методов было показано (Сухих и др., 2020), что в водохранилищах Волги массовым видом является E. caspica, тогда как типичная E. affinis не обнаружена. В 2018 г. в зоопланктоне Цимлянского водохранилища и канала Волга–Дон повсеместно обитала только E. caspica. Важными определительными признаками E. caspica служат форма выростов последнего торакального сегмента и строение ног пятой пары самки, а у самца – строение геникулирующей антеннулы и форма мандибул (рис. 4а). Определительный ключ видов этого рода приведен в статье (Сухих и др., 2020).
Рис. 4.
Определительные признаки видов Eurytemora caspica Sukhikh et Alekseev 2013 (а) и Thermocyclops taihokuensis (Harada 1931) (б). На рис. а: 1 – общий вид самки, 2 – пятая пара ног самки, 3 – геникулирующая (правая) антеннула и мандибулы самца; на рис. б: 1 – общий вид, 2 – семяприемник, 3 – четвертая пара ног самки.
Согласно представлениям Мордухай-Болтовского (1960), каспийская автохтонная фауна и формы, производные от нее, населяют многие участки понто-каспийского бассейна, в том числе Азовское море и устьевые области впадающих в него рек. Если предположить, что E. caspica представляет собой древний каспийский вид, то находки этого вида в среднем течении р. Дон вполне вписываются в ареал автохтонной каспийской фауны. Необходимо перепроверить определения видов рода Eurytemora в Азовском море и дельте р. Дон, поскольку, возможно, там тоже обитает E. caspica.
Недавний вселенец Thermocyclops taihokuensis (Harada, 1931) (syn. T. asiatiсus (Kiefer, 1932) обычен в Китае, Центральной и Восточной Азии, в том числе в Казахстане (Guo, 1999; Monchenko, 2008, Калымбетова, 2017). В последние годы (2005–2018 гг.) он быстро расселяется на запад. В начале 2000-х годов этот вид обнаружен в Северо-Восточном Каспии (Шарапова, 2014), в 2010-х годах – в Волго-Ахтубинской пойме (Нечаев, 2016), р. Волга ниже г. Волгограда (Лазарева и др., 2018) и даже в устьевой области р. Сура (Чебоксарское водохранилище) (Жихарев и др., 2019). В тот же период (с 2012 г.) он зарегистрирован в Цимлянском водохранилище (Вехов и др., 2014). В 2018 г. авторами установлено, что T. taihokuensis – формирует высокую численность не только в Цимлянском, но также в Бреславском и Карповском водохранилищах канала Волга-Дон. По вооружению 4–5-й пары ног и фуркальных ветвей вид сходен с T. oithonoides (Sars, 1863), отличительными признаками служат форма семяприемника самки, S-образно изогнутые апикальные щетинки фурки и мощный внутренний шип дистального членика эндоподита четвертой пары ног (рис. 4б).
Два других вида копепод Acanthocyclops americanus и Halicyclops neglectus, впервые обнаруженные в Цимлянском водохранилище в 2018 г., обычны на юге европейской части России (Монченко, 1974). Оба вида зарегистрированы в дельте р. Дон (Свистунова и др., 2014). Кроме того, Acanthocyclops americanus широко распространен в водохранилищах Волги и Камы (Лазарева и др., 2018).
Среди массовых видов ракообразных Цимлянского водохранилища только два (Calanipeda aquaedulcis и Moina micrura) относятся к фильтраторам фито-детритофагам (Гунько, Алдакимова, 1963; Монаков, 1998). Основу сообщества (до 90% обилия ракообразных) формируют всеядные копеподы-хвататели Thermocyclops taihokuensis, Acanthocyclops americanus, а также Heterocope caspia, наряду с захватом крупной пищи способная к фильтрации мелких частиц (табл. 4). Взрослые особи этих трех видов и их копеподиты с третьей стадии развития могут потреблять животную пищу (коловратки, инфузории, мелкие ракообразные и собственная молодь) (Монаков, 1998; Монченко, 1974). Как результат, в конце лета уровень хищничества в сообществе был очень высоким, соотношение биомассы хищников и не хищных животных (индекс Вхищ/Вмир) составляло 1.1. Для сравнения, в водохранилищах канала Волга–Дон этот индекс был в четыре раза ниже (Вхищ/Вмир 0.25). Заметим, что расчетный уровень хищничества не всегда отражает реальные трофические взаимодействия в сообществе с доминированием всеядных видов. При недостатке животной пищи они переходят на питание водорослевым и бактериальным детритом (Монаков, 1998). Облигатные хищники в Цимлянском водохранилище представлены двумя малочисленными видами Cercopagis pengoi (менее 50 экз./м3) и Leptodora kindtii (10–500 экз./м3).
В водохранилищах канала Волга–Дон сравнительно многочисленны (в сумме до 12 тыс. экз./м3) четыре вида хищных кладоцер (Leptodora kindtii, Cercopagis pengoi, Cornigerius maeoticus и Podonevadne trigona). Среди них преобладают (70–90% общего количества) мелкие (длина тела менее 0.5 мм) виды, спектр доступной пищи которых ограничен простейшими, коловратками и науплиусами копепод. В большинстве водоемов канала доминируют (до 40% численности ракообразных) всеядные копеподы Thermocyclops taihokuensis, Heterocope caspia и Eurytemora caspica (табл. 5), которым доступен весь спектр пищевых ресурсов. Однако их вклад в сообщество много меньше, чем в Цимлянском водохранилище. Фильтраторы здесь представлены шестью видами, среди которых преобладают мелкие (длина тела менее 1 мм) кладоцеры (табл. 5). Наряду с низким индексом Вхищ/Вмир, это указывает на меньший пресс планктонных хищников.
До начала 1970-х годов выделяют три периода развития зоопланктона Цимлянского водохранилища (Гламазда, 1971, 1974). В первый период (1952–1954 гг.) происходило перераспределение по акватории речного сообщества и его трансформация в озерное, массовое развитие эвритопных видов и достижение наибольшей за время существования водоема биомассы (4–5 г/м3) (Дзюбан, 1958; Шейнин, 1960). Во второй период (1955–1958 гг.) отмечены снижение летней биомассы зоопланктона менее 3 г/м3 и вытеснение эвритопных форм озерными (Кафтанникова, 1965). Третий период (1959–1971 гг.) характеризуется массовым развитием понто-каспийских видов кладоцер и особенно солоноватоводных копепод, при этом биомасса зоопланктона снижается до менее чем 2 г/м3, в отдельные годы ниже 1 г/м3 (Кафтанникова, 1965; Гламазда, 1971, 1974).
Важным в эволюции зоопланктона водохранилища стал четвертый период, начавшийся в середине 1970-х и, вероятно, продолжавшийся в 1980-е годы. В это время на фоне низкого количества зоопланктона (летняя биомасса менее 1.5 г/м3) отмечено вытеснение солоноватоводными видами большинства представителей ветвистоусых и веслоногих ракообразных, ранее обычных для среднего течения р. Дон. Изменение состава и структуры зоопланктона, в том числе вселение в водоем новых южных теплолюбивых видов, происходит до настоящего времени.
Максимальную летнюю биомассу (3.8–4.6 г/м3) зоопланктона регистрировали в первые три года существования Цимлянского водохранилища (Лившиц, 1954; Дзюбан, 1958). В конце 1950-х годов она снизилась до менее чем 1.5 г/м3 (Кафтанникова, 1965; Шейнин, 1960), в 2000-х годах иногда регистрировали менее 0.5 г/м3 (Шевлякова, 2002; Свистунова, Саяпин, 2010; Вехов и др., 2014). Отличительной чертой современного периода (2015–2018 гг.) стало увеличение биомассы до уровня конца 1950-х годов преимущественно за счет крупных (длина тела 1.1–1.6 мм) ракообразных Heterocope caspia, Calanipeda aquaedulcis и Acanthocyclops americanus. Современная биомасса зоопланктона (2.1 ± 0.5 г/м3) в Цимлянском водохранилище сопоставима с наблюдаемой в эвтрофных водохранилищах Верхней Волги и Камы (1–2 г/м3) (Лазарева и др., 2018). Она существенно выше по сравнению с мезотрофными водохранилищами Нижней Волги (0.2–1.2 г/м3) (Малинина и др., 2016; Лазарева и др., 2018).
ЗАКЛЮЧЕНИЕ
В августе–сентябре 2018 г. в составе зоопланктона исследованных водоемов обнаружено 64 вида, среди которых преобладали Rotifera (60%). В Цимлянском водохранилище зарегистрировано 56 видов, в том числе выявлено 15 форм, которые не упоминаются в ранее опубликованных работах. Впервые представлен состав зоопланктона водохранилищ Волго-Донского канала (48 видов). Установлено, что основу (60%) численности зоопланктона Цимлянского водохранилища формируют Copepoda, они же доминируют (70%) по биомассе. Наиболее многочисленны понто-каспийская Heterocope caspia и вселенцы Calanipeda aquaedulcis, Thermocyclops taihokuensis и Acanthocyclops americanus. Обилие зоопланктона снижается от верхнего участка водохранилища к плотине Цимлянской ГЭС. На основе анализа литературных данных за 1952–2018 гг. показано, что смена донского комплекса доминантов зоопланктона водохранилища на солноватоводные виды произошла во второй половине 1970-х годов. К 2018 г. в таксоценозах Cladocera и Rotifera сохранились небольшие группировки исходных доминантов р. Дон, тогда как массовые виды Copepoda сменились полностью. Выявлено увеличение более чем вдвое (до 2.1 ± 0.5 г/м3) биомассы сообщества в последние четыре года (2015–2018 гг.) по сравнению с 40-летним периодом до 2013 г. (0.8 ± 0.1 г/м3). Количество зоопланктона в Цимлянском водохранилище существенно выше по сравнению с другими водохранилищами аридной зоны. Численность зоопланктона водоемов Волго-Донского канала определяется в равной мере Rotifera и Crustacea, биомассу определяют в основном Cladocera (49%). Наиболее многочисленны Bosmina longirostris и Podonevadne trigona (Варваровское водохранилище), Chydorus sphaericus (Карповское водохранилище), Bosmina cf. coregoni и Daphnia cucullata (Бреславское водохранилище). В числе массовых видов (12–23% численности ракообразных) присутствуют вселенцы Thermocyclops taihokuensis, Calanipeda aquaedulcis и понто-каспийские Heterocope caspia, Eurytemora caspica. Средняя биомасса зоопланктона в канале Волга-Дон (0.9 ± ± 0.4 г/м3) вдвое ниже, чем в Цимлянском водохранилище.
Список литературы
Авакян А.Б., Салтанкин В.П., Шарапов В.А., 1987. Водохранилища. М.: Мысль. 325 с.
Боруцкий Е.С., Степанова Л.А., Кос М.С., 1991. Определитель Calanoida пресных вод СССР. Л.: Наука. 504 с.
Вехов Д.А., Науменко А.Н., Горелов В.П., Голоколенова Т.Б., Шевлякова Т.П., 2014. Современное состояние и использование водных биоресурсов Цимлянского водохранилища (2009–2013 гг.) // Рыбохозяйственные исследования на водных объектах Европейской части России. СПб.: ГосНИОРХ. С. 116–145.
Гидрометеорологический режим озер и водохранилищ СССР. Цимлянское, водораздельные и Манычские водохранилища, 1977. Л.: Гидрометеоиздат. 204 с.
Гламазда В.В., 1969. О полифемидах Цимлянского водохранилища // Гидробиологический журнал. Т. 5. № 5. С. 97–98.
Гламазда В.В., 1971. Динамика зоопланктона Цимлянского водохранилища (по материалам 1966–1968 гг. // Труды Волгоградского отделения Гос. науч.-исслед. ин-та озерного и речного рыбного хоз-ва (ГосНИОРХ). Т. 5. С. 25–44.
Гламазда В.В., 1971а. О нахождении Cercopagis pengoi (Ostr.) в Цимлянском водохранилище // Гидробиологический журнал. Т. 7. № 4. С. 70–71.
Гламазда В.В., 1974. Зоопланктон Цимлянского водохранилища в 1969–1971 гг. // Труды Волгоградского отделения Гос. науч.-исслед. ин-та озерного и речного рыбного хоз-ва (ГосНИОРХ). Т. 8. С. 29–38.
Гламазда В.В., 1982. Современное состояние зоопланктона Цимлянского водохранилища и его продукция // Труды Гос. науч.-исслед. ин-та озерного и речного рыбного хоз-ва (ГосНИОРХ). Вып. 184. С. 30–34.
Голоколенова Т.Б., Горелов В.П., Шевлякова Т.П., Басько Ю.В., 2019. Кормовая база Цимлянского водохранилища в 2016–2018 гг. // Проблемы устойчивого развития и экономической безопасности региона. Матер. XIV Междунар. науч.-практ. конф. Волгоград–Волжский 10–11 апреля 2019 г. Волгоград: Изд-во ВолГУ. С. 115–121.
Гунько А.Ф., Алдакимова А.Я., 1963. Материалы о питании Calanipeda aquae-dulcis Kritsch. (Crustacea, Calanoida) в Азовском море // Рыбохозяйственные исследования в Азовском бассейне по запасам рыб и условиям их обитания. М.: Рыбное хозяйство. С. 3–5.
Дзюбан Н.А., 1958. Зоопланктон Цимлянского водохранилища // Известия Всесоюзного науч.-исслед. ин-та озерного и речного рыбного хоз-ва (ВНИРО). Т. 45. С. 51–74.
Жихарев В.С., Гаврилко Д.Е., Шурганова Г.В., 2019. Находка тропического вида Thermocyclops taihokuensis (Harada, 1931) (Copepoda: Cyclopoida) в Европейской части России // Поволжский экологический журнал. № 2. С. 264–270.
Калымбетова М.Т., 2017. Современное состояние зоопланктона Шардаринского водохранилища // Вестник сельскохозяйственной науки Казахстана. № 5–6. С. 80–83.
Кафтанникова О.Г., 1965. Зоопланктон Цимлянского водохранилища по материалам 1955–1962 гг. // Труды Волгоградского отделения Гос. науч.-исслед. ин-та озерного и речного рыбного хоз-ва (ГосНИОРХ). Т. 1. С. 71–91.
Кутикова Л.А., 1970. Коловратки фауны СССР. Л.: Наука. 744 с.
Лазарева В.И., Сабитова Р.З., Быкова С.В., Жданова С.М., Соколова Е.А., 2018. Распределение летнего зоопланктона в каскаде водохранилищ Волги и Камы // Труды Ин-та биологии внутренних вод РАН. Вып. 83(86). С. 62‒84. https://doi.org/10.24411/0320-3557-2018-10030
Лившиц Н.М., 1954. Планктон Цимлянского водохранилища по летним сборам 1952 г. // Известия Всесоюзного науч.-исслед. ин-та озерного и речного рыбного хоз-ва (ВНИРО). Т. 34. С. 61–77.
Малинина Ю.А., Джаяни Е.А., Филинова Е.И., Колозин В.А., Шашуловский В.А., 2016. Оценка темпов многолетних изменений качественных и количественных параметров экосистемы Саратовского водохранилища // Современное состояние биоресурсов внутренних водоемов и пути их рационального использования. Материалы Всерос. конф. Казань: Гос. ин-т речн. рыб. хоз-ва (ГосНИОРХ). С. 618–625.
Монаков А.В., 1998. Питание пресноводных беспозвоночных. М.: Ин-т проблем экологии и эволюции. 321 с.
Монченко В.I., 1974. Щелепноротi циклопоподiбнi. Циклопи (Cyclopidae). Киев: Наук. думка. 452 с. (Фауна України; Т. 27, вип. 3).
Мордухай-Болтовской Ф.Д., 1960. Каспийская фауна в Азово-Черноморском бассейне / ред. Кузин Б.С. М.-Л.: Изд-во АН СССР. 288 с.
Мордухай-Болтовской Ф.Д., Ривьер И.К., 1987. Хищные ветвистоусые Podonidae, Polyphemidae, Cercopagidae и Leptodoridae фауны мира. Л.: Наука. 182 с.
Моруков Ю.Н., 2006. Волго-Донской канал // Большая Российская Энциклопедия. Т. 5. С. 625–626.
Нечаев Д.Ю., 2016. Фаунистическое разнообразие планктонных беспозвоночных Волго-Ахтубинской поймы // Материалы Всерос. молодежной гидробиол. конф. “Перспективы и проблемы современной гидробиологии”, пос. Борок, Ин-т биологии внутренних вод им. И.Д. Папанина РАН, 10–13 ноября 2016 г. Ред. Турбанов И.С., Климова Я.С, Синельников С.Ю. Ярославль: Филигрань. С. 117–119.
Никаноров А.М., Хоружая Т.А., 2014. Внутриводоемные процессы в крупных водохранилищах юга России (загрязнение, эвтрофирование, токсификация) // География и природные ресурсы. № 2. С. 35–43.
Определитель зоопланктона и зообентоса пресных вод Европейской России, 2010. Т. 1. Зоопланктон. Ред. Алексеев В.Р. М.: Товарищество научных изданий КМК. 495 с.
Песенко Ю.А., 1982. Принципы и методы количественного анализа в фаунистических исследованиях. М.: Наука. 286 с.
Предварительные материалы оценки воздействия на окружающую среду эксплуатации энергоблока № 1 Ростовской АЭС в 18-месячном топливном цикле на мощности реакторной установки 104% от номинальной, 2018. Книга 2. Гидротехпроект, 504 c. [Электронный ресурс] URL: http://old.volgodonskgorod.ru/all/public/pages/dosc/kniga_2_ovos_3_12.18.pdf Дата обновления: 26.02.2020.
Селифонова Ж.П., 2013. Состояние таксоцена веслоногих раков (Copepoda) в Азовском море // Vestnik zoologii. Т. 47. № 5 С. 421–430.
Свистунова Л.Д., Саяпин В.В., 2010. Проблема эвтрофирования Цимлянского водохранилища (Ростовская область) и новые данные о его зоопланктоне // Вестник Южного научного центра РАН. Т. 6. № 4. С. 61–67.
Свистунова Л.Д., Брынько В.А., Набоженко М.В., 2014. Современное состояние летнего зоопланктона дельты реки Дон // Вестник Южного научного центра РАН. Т. 10. № 3. С. 75–82.
Сухих Н.М., Лазарева В.И., Алексеев В.Р., 2020. Копепода Eurytemora caspica Sukhikh et Alekseev, 2013 (Crustacea, Calanoida) в водохранилищах рек Волга и Кама // Биология внутренних вод. № 2. С. 129‒ 136. https://doi.org/10.31857/S0320965220020163
Тимохина А.Ф., 2000. Зоопланктон как компонент экосистемы Куйбышевского водохранилища. Тольятти: Ин-т экологии волжского бассейна РАН. 193 с.
Цимлянское водохранилище: состояние водных и прибрежных экосистем, проблемы и пути их решения, 2011. Ред. акад. Матишов Г.Г. Ростов-на-Дону: Изд-во ЮНЦ РАН. 216 с.
Шарапова Л.И., 2014. Зоопланктон Северо-Восточного Каспия. Приложение 2. // Мониторинг окружающей природной среды Северо-Восточного Каспия при освоении нефтяных месторождений. Алматы: АджипККО. С. 228–229.
Шевлякова Т.П., 2002. Современное состояние зоопланктона Цимлянского водохранилища (по материалам 1996–2001 гг.) // Рыбохозяйственные исследования в бассейне Волго-Донского междуречья на современном этапе (к 50-летию Волгоградского отделения ГосНИОРХ). СПб.: Квинта Северо-Запад. С. 46–52.
Шейнин М.С., 1960. Зоопланктон Нижнего Дона, его водохранилищ и восточной части Таганрогского залива в годы зарегулированного стока // Труды Азовского науч.-исслед. ин-та рыбного хоз-ва (АзНИИРХ). Т. 1. Вып. 1. Ростовское книжное издательство. С. 231–258.
Guo X., 1999. The genus Thermocyclops Kiefer, 1927 (Copepoda: Cyclopidae) in China // Hydrobiologia. V. 403. P. 87–95.
Monchenko V.I., 2008. Redescription of the Oriental Thermocyclops taihokuensis (Harada, 1931) (Copepoda: Cyclopoida) from its westernmost population // Zoology in the Middle East. V. 43. № 1. P. 99–104. https://doi.org/10.1080/09397140.2008.10638274
Sukhikh N.M., Alekseev V.R., 2013. Eurytemora caspica sp. nov. from the Caspian Sea – one more new species within the E. affinis complex (Copepoda: Calanoida, Temoridae) // Proceedings of the Zoological Institute RAS. V. 317. № 1. P. 85–100.
Дополнительные материалы отсутствуют.
Инструменты
Зоологический журнал