

Зоологический журнал, 2021, T. 100, № 7, стр. 756-769
Жесткокрылые (Coleoptera), связанные с ксилотрофными грибами рода Trametes Fr. (Basidiomycetes, Polyporales) на Южном Урале
Б. В. Красуцкий *
Челябинский государственный университет
454001 Челябинск, Россия
* E-mail: boris_k.63@mail.ru
Поступила в редакцию 30.05.2020
После доработки 23.11.2020
Принята к публикации 25.11.2020
Аннотация
Выявлено 39 видов жуков из 25 родов и 13 семейств, связанных с 6 видами базидиальных дереворазрушающих грибов рода Trametes Fr. (T. gibbosa, T. hirsuta, T. ochracea, T. pubescens, T. suaveolens, T. versicolor) на Южном Урале. Доминирующими обитателями этих грибов являются Octothemnus glabriculus, Sulcacis nitidus, Cis hispidus, C. setiger, C. boleti (Ciidae), Tritoma subbasalis (Erotylidae) и Orchesia fusiformis (Melandryidae). Установлены закономерности заселения плодовых тел в процессе изменения их физиологического состояния и в зависимости от степени разрушенности; показано существенное различие состава комплекса обитателей живых и мертвых базидиом. Под корой деревьев, в белых гнилях древесины и в мицелиальном слое грибов рода Trametes в связи с ними и другими видами грибов (в том числе аскомицетами и дейтеромицетами) развиваются Cerylon deplanatum, C. ferrugineum, C. histeroides (Cerylonidae), Rhizophagus dispar, Rh. parvulus (Monotomidae), Cucujus haematodes (Cucujidae), Bitoma crenata (Colydiidae), Dircaea quadriguttata и Orchesia fasciata (Melandryidae).
Представленная работа является частью многолетних исследований по фауне, биологии и экологии жесткокрылых (Coleoptera), связанных с ксилотрофными базидиальными грибами (Basidiomycetes, Hymenomycetidae) Урала и Западной Сибири, главные результаты которой опубликованы ранее (Красуцкий, 1990, 1994, 1995, 1996, 1996а, 1997, 1997а, 2000, 2001, 2005, 2007, 2007а, 2010, 2013, 2013а, 2014, 2016, 2018). На территории Южного Урала эти исследования выполняли с 1990 г. по настоящее время в Ильменском заповеднике, Аршинском, Ашинском, Карагайском, Нязепетровском, Серпиевском, Троицком, Уйском, Черноборском заказниках, в окрестностях г. Челябинска (Городской и Каштакский боры), в Аргаяшском, Аргазинском, Брединском, Верхнеуральском, Каслинском, Кизильском, Красноармейском, Кунашакском, Нагайбакском, Сосновском, Катав-Ивановском и Чебаркульском р-нах (рис. 1).
В этой статье мы обсуждаем состав и структуру мицетофильного сообщества грибов рода Trametes Fr. (Polyporales, Polyporaceae) – широко распространенных и активно заселяемых насекомыми на Южном Урале и в других регионах (Компанцев, 1984; Красуцкий, 2005, 2016; Nikitsky, Schigel, 2004). Для микобиоты России указано 9 видов грибов этого рода (Бондарцева, 1998), для Южного Урала – 7–9 видов (Степанова-Картавенко, 1967; Kotiranta et al., 2005). Нами исследовано 6 наиболее обычных видов: Trametes gibbosa (Pers.) Fr., T. hirsuta (Wulf.: Fr.) Pil., T. ochracea (Pers.) Gilbn. et Ryv., T. pubescens (Schumm.: Fr.) Pil., T. suaveolens (Fr.) Fr., T. versicolor (L.: Fr.) Pil. Все представители рода трофически связаны с лиственными породами деревьев (береза, осина, липа, ольха, ясень, рябина, ильм, черемуха, ива, яблоня и др.), как исключение могут встречаться на некоторых хвойных (сосна, лиственница, ель), вызывают белую гниль (Бондарцева, 1998).
КРАТКИЙ ОБЗОР ИССЛЕДОВАНИЙ ЭНТОМОКОМПЛЕКСОВ ГРИБОВ РОДА TRAMETES В РОССИИ
Первые обширные и систематизированные сведения о жесткокрылых, связанных с грибами, мы находим в работе Беника (1952), в которой для Средней Европы указано более 1100 мицетофильных видов жуков, для грибов рода Trametes названо 68 видов жуков из 14 семейств, а из них в качестве мицетобионтов (т.е. облигатно связанных с грибами видов по терминологии Л. Беника) – 34 вида из 6 семейств.
В 1975 г Яблоков-Хнзорян (Jablokoff-Khnzorian, 1975), проводя ревизию жуков-грибовиков Палеарктики, характеризует их связи с ксилотрофными грибами и указывает на развитие ряда видов (Aulacochilus sibiricus Rtt., A. luniferus Guer., Dacne maculata Chûjô, Tritoma niponensis (Lew.)) в грибах рода Trametes, преимущественно, T. hirsuta, T. versicolor.
Компанцев (1982) на основе оригинальных материалов с территории европейской части России, Кавказа и Дальнего Востока показал, что личинки ряда видов жуков-грибовиков (Erotylidae) являются характерными обитателями плодовых тел грибов рода Trametes. В Костромской обл. в плодовых телах T. hirsuta, T. ochracea и T. versicolor он отмечает, кроме грибовика Tritoma bipustulata (F.) (Erotylidae), 9 видов семейства Ciidae (Компанцев, 1984).
Компанцева (1987) приводит сведения о трех видах мицетофильных чернотелок (Tenebrionidae), развитие которых может быть связано с грибами рода Trametes.
В “Определителе насекомых Дальнего Востока” (1992) содержатся данные о связях некоторых жесткокрылых с конкретными грибами. Так, Криволуцкая (1992) в качестве пищевых объектов жуков-грибовиков называет некоторые грибы рода Trametes (= Coriolus). Никитский (1992) указывает, что тетратомиды Pisenus insignis (Rtt.) (Tetratomidae) развиваются в грибах Trametes (= Coriolus) spp., а тенелюб Orchesia fusiformis (Sols.) (Melandryidae) – в Trametes pubescens. Никитский и Компанцев (1995) сообщают о развитии отдельных видов грибовиков в грибах T. hirsuta, T. ochracea, T. versicolor и T. suaveolens.
В Беларуси на доминирующих видах рода Trametes (T. gibbosa, T. hirsuta, T. ochracea, T. versicolor) Цинкевич (2004) выявил 20 видов жесткокрылых из 7 семейств, 10 из которых – Ciidae.
В фундаментальной работе Рейбница (Reibnitz, 1999) по циидам Германии детально рассмотрен комплекс обитателей 5 видов рода Trametes (T. hirsuta, T. gibbosa, T. ochracea, T. suaveolens, T. versicolor). В общей сложности зарегистрировано 16 видов трутовиковых жуков из 5 родов, предложена шкала их встречаемости по 16 выделенным районам (Reibnitz, 1999). Закономерности заселения трутовиковыми жуками грибов Trametes ochracea в Финляндии изучены Комоненом (Komonen, 2008). Для многих мицетофильных видов, заселяющих в Великобритании и Ирландии разрушающуюся древесину, названы в том числе грибы, в которых происходит развитие личинок. Так, из 13 видов трутовиковых жуков этого списка в грибах рода Trametes отмечены 6 (Alexander, 2002).
На территории европейской части России (Московская, Ярославская обл., Республика Карелия, Кольский п-ов, Республика Коми, Крым), отчасти на Полярном (ст. Красный Камень) и Приполярном Урале (хребет Северные Малды) изучены ксило-мицетофильные жуки более 250 видов базидиомицетов, аскомицетов и дейтеромицетов, а также более 30 видов миксомицетов (Никитский и др., 1996; Никитский, Татаринова, 2003; Власов, Никитский, 2015; Nikitsky, Schigel, 2004; Schigel, 2004, 2005, 2006, 2009, 2011, 2011a). В итоге для грибов рода Trametes (T. cervina, T. gibbosa, T. hirsuta, T. ochracea, T. pubescens, T. suaveolens, T. velutina, T. versicolor) указано свыше 37 видов жуков из 7 семейств. В качестве типичных их обитателей названы 10 видов трутовиковых жуков: Cis boleti, C. comptus, C. fissicornis, C. jacquemarti, C. micans (F.), C. rugulosus Mel., Octotemnus glabriculus, Sulcacis nitidus, S. fronticornis, Wagaicis wagai, 2 вида грибовиков – Tritoma bipustulata, T. subbasalis, 1 вид тенелюбов – Orchesia fusiformis и несколько видов стафилинид (Staphylinidae).
МАТЕРИАЛ И МЕТОДИКА
Материалом для работы послужили жуки, собранные с поверхности и из толщи 2320 базидиом грибов рода Trametes и из 38 прилежащих к ним участков субстрата (коры и древесины).
В зависимости от физиологического состояния плодовых тел мы различали следующие стадии их существования:
1 стадия – плодовые тела живые, растущие и на стадии спороношения, обычно без повреждений;
2 стадия – плодовые тела полностью закончили рост, спороношение и начинают отмирать. В конце этой стадии нередко уже заметны незначительные повреждения (до 25% объема);
3 стадия – плодовые тела мертвые, средней степени разрушенности (до 50% объема), но еще сохраняют структурные свойства;
4 стадия – плодовые тела мертвые, сильной или почти полной степени разрушенности (более 75% объема). Повреждены все части базидиом, и они утрачивают присущие им структурные свойства. В конце 4 стадии процесс деструкции заканчивается и начинается минерализация остатков плодовых тел, попавших в подстилку.
Пищевые связи имаго и личинок жуков изучали как в природе, так и в лабораторных условиях с использованием, преимущественно, адаптированных садковых методик (садками послужили завязанные сверху марлей стеклянные стаканы, на дно которых насыпали слегка увлажненные древесные опилки). Наблюдали за особенностями питания личинок, отмечали их локализацию в определенных частях базидиом (трама, гимениальный слой) и успешность развития, т.е. достижение имагинальной стадии. Взрослые жуки вывелись во всех садках. Небольшие фрагменты плодовых тел без видимых повреждений (полагали, что в них могли быть отложены яйца насекомых) помещали в пронумерованные алюминиевые стаканчики, которые ставили в темное место и в течение 3–4 месяцев периодически проверяли, выбирая появившихся жуков.
Заселенность грибов определяли отношением числа заселенных плодовых тел к общему их числу во взятой выборке. С учетом этого показателя в наших работах мы выделяли группы не заселяемых, слабо заселяемых (до 25% заселенных плодовых тел), умеренно заселяемых (от 25 до 50%), активно заселяемых (от 50 до 75%) и чрезвычайно активно заселяемых (более 75%) грибов (Красуцкий, 2005).
При описании характера взаимоотношений насекомых с грибами учитывали следующие признаки (Красуцкий, 1994, 2005, 2007, 2010; Яковлев, 1966, 1994):
1) Степень экологической специализации – критерий, учитывающий общий характер связей жуков с грибами на всех стадиях их жизненного цикла. В соответствии с этим критерием выделяли следующие две крупные группы: специализированные мицетобионты-мицетофаги – живут и развиваются только или главным образом в (на) грибах (их базидиомах) и эврибионты – могут заселять различные среды или субстраты (почва, подстилка, древесина, пни, дупла и т.д.), в том числе и плодовые тела грибов.
2) Особенности образа жизни личинок и имаго. В соответствии с этим критерием выделяли открытоживущие формы – обитают и развиваются на поверхности плодовых тел и скрытноживущие формы – заселяют толщу базидиом. При этом все найденные за время проведения наших исследований на Урале и в Западной Сибири виды являются типичными мицетофагами; полные мицетофаги и мицетосапрофаги в этой группе отсутствуют (Красуцкий, 2005, 2006, 2007, 2010, 2016).
3) Пищевые связи насекомых с грибами и друг с другом в течение всего их жизненного цикла. Выделены следующие подгруппы:
Подгруппа облигатные мицетофаги – питаются исключительно грибами независимо от того, в какой среде они обитают. Многие мицетофаги живут и развиваются в плодовых телах грибов и, таким образом, являются специализированными мицетобионтами; вместе с тем большое число насекомых живут в других субстратах (почва, подстилка, древесина, труха дупел и т.п.), но при этом питаются только грибным веществом (обычно характерно питание их мицелием и спорами) (Аксентьев, Участнова, 1986; Красуцкий, 2005).
По типу мицетофагии в подгруппе облигатных мицетофагов выделены три группировки жуков:
Типичные мицетофаги – используют только живое вещество грибов (плодовые тела, мицелий, споры);
Мицетосапрофаги – развиваются в мертвых тканях грибов (плодовых тел);
Полные мицетофаги – способны использовать в пищу как живые, так и мертвые грибные ткани.
Подгруппа миксофаги – совмещают (обязательное условие) различные типы питания, один из которых – мицетофагия. Возможны самые разнообразные сочетания типов питания: ксило-мицетофагия, сапро-мицетофагия, сапро-ксило-мицетофагия, детритофагия и мицетофагия, некрофагия и мицетофагия. В энтомокомплексах ксилотрофных грибов миксофаги чаще всего являются обитателями древесины и подкорного пространства деревьев, пораженных трутовиками и другими грибами.
Подгруппа факультативные (частичные) мицетофаги – используют грибы не как один из основных источников пищи, а лишь как дополнительный источник пищи, поэтому их связи с грибами не постоянны, часто ограничиваются кратковременными трофическими контактами, как правило, не приводящими к выраженной топической специализации. Частичные мицетофаги обычно являются членами ксилофильного сообщества или представителями комплекса герпетобионтов.
Подгруппа хищники, паразиты и некрофаги – охотятся на обитателей грибов, паразитируют на них или поедают их трупы. Проникают в плодовые тела из древесины или из почвы (подстилки). Специализированных мицетобионтов мало.
По широте трофических связей среди специализированных мицетобионтов-мицетофагов выделены полифаги, олигофаги и монофаги.
полифаги – заселяют грибы, относящиеся к разным порядкам и даже к разным классам;
олигофаги – трофически связаны с грибами одного семейства или порядка;
монофаги – связаны в питании с грибами одного вида или рода.
Еще одна небольшая группа – случайные посетители. Это жуки (преимущественно, имаго), находки которых единичны, и те виды, связи которых с грибами (их обитателями) на данный момент времени не выяснены.
Для анализа пищевых связей жуков-мицетофагов с грибами (в том числе с грибами рода Trametes) разработан простейший коэффициент предпочтения, отражающий долю участия конкретного вида гриба в общем пищевом рационе насекомых. Так, если жук N развивается за счет питания 5 видами грибов – A, B, C, D, E, то коэффициент предпочтения им гриба A будет:
где значения A, B, C, D, E соответствуют количеству плодовых тел, заселяемых жуком N.В отличие от коэффициента верности местообитанию (Ердаков и др., 1978), индекса биотопической приуроченности (Песенко, 1982), гостально-топического индекса (Богданов и др., 2006) и других близких показателей, мы делали акцент на трофике жуков без учета численности насекомых; т.е. оценивали специфику питания по факту присутствия личинок в плодовых телах конкретных видов грибов.
Количественные данные о пищевых связях даже в таком упрощенном представлении могут стать основой для более глубокого изучения структуры пищевых сетей в мицетофильном сообществе, когда мы рассматриваем, с одной стороны, множество видов насекомых-мицетофагов (и их хищников и паразитов), с другой – множество видов грибов (и обитателей грибов), с которыми они обнаруживают самые тесные связи. В зарубежной литературе уже давно проработаны вопросы детального анализа трофики (Elton, 1927) и установления неслучайного характера парных взаимодействий и взаимодействий более высокого порядка (Paine, 1988; Orledge et al., 2005).
РЕЗУЛЬТАТЫ
Заселенность грибов рода Trametes жуками высока и находится в диапазоне от 93 (T. pubescens, T. versicolor) до 100% (T. gibbosa, T. hirsuta, T. ochracea, T. suaveolens). Таким образом, эти грибы, наряду с некоторыми другими видами семейства Polyporaceae, например, Pycnoporus cinnabarinus (Jacq.: Fr.) P. Karst. и Lenzites betulina (L.: Fr.) Fr., являются чрезвычайно активно заселяемыми.
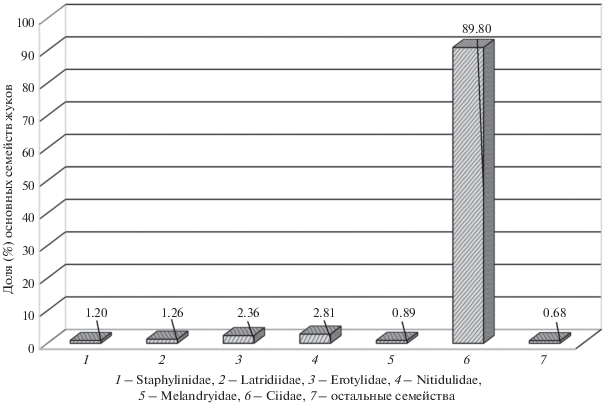
С грибами рода Trametes на Южном Урале связаны 39 видов жесткокрылых из 25 родов и 13 семейств (табл. 1). Доминирующими обитателями их плодовых тел являются Ciidae, из них наиболее обычны и многочисленны Sulcacis nitidus и Octothemnus glabriculus, несколько реже встречается Cis boleti и ряд других видов рода Cis (Latr.) (рис. 2). Встречаемость представителей остальных семейств относительно низка; из их числа следует назвать грибовиков Dacne bipustulata, Tritoma subbasalis (Erotylidae), блестянок Epuraea variegata (Nitidulidae), некоторых жуков-скрытников (Latridiidae), стафилинид рода Scaphisoma Leach. (Staphylinidae) и тенелюбов Orchesia fusiformis (Melandryidae). Очень редки в (на) плодовых телах коротконадкрылые Acrulia inflata, Cilea silphoides. Oxyporus maxillosus, Lordithon lunulatus, L. trimaculatus, Sepedophilus bipustulatus (Staphylinidae), Anisotoma axillaris (Leiodidae), Ampedus pomonae (Elateridae), грибоеды Mycetophagus piceus, M. quadripustulatus (Mycetophagidae), обнаруженные только на стадии имаго. В мицелиальном слое грибов под корой березы и осины иногда развиваются гладкотелы рода Cerylon Latr. (Cerylonidae), монотомиды Rhizophagus dispar, Rh. parvulus (Monotomidae), узкотелка Bitoma crenata (Colydiidae) и тенелюбы Dircaea quadriguttata и Orchesia fasciata (Melandryidae).
Таблица 1.
Жесткокрылые (Coleoptera), связанные с грибами рода Trametes (Polyporales), на Южном Урале
| Семейства и виды жуков | Встречаемость жуков в (на) грибах рода Trametes (% заселенных плодовых тел от всех исследованных) |
|||||
|---|---|---|---|---|---|---|
| gibb | hir | ochr | pub | suav | ver | |
| I. Staphylinidae | ||||||
| 1. Scaphisoma agaricinum (L.)* | ед. | + | 2.27 | + | – | 1.74 |
| 2. Scaphisoma inopinatum (Lobl)* | + | 1.42 | 2.72 | + | + | 3.04 |
| 3. Acrulia inflata (Gyll.)* | ед. | – | – | – | – | ед. |
| 4. Cilea silphoides (L.)* | – | – | – | – | – | ед. |
| 5. Oxyporus maxillosus (F.)* | ед. | – | + | – | – | + |
| 6. Lordithon lunulatus (L.)* | + | – | + | – | – | + |
| 7. Lordithon trimaculatus (F.)* | ед. | – | ед. | – | – | – |
| 8. Sepedophilus bipustulatus (Oliv.)* | + | + | + | ед. | – | + |
| II. Leiodidae | ||||||
| 9. Anisotoma axillaris Gyll.* | – | + | – | – | – | – |
| III. Elateridae | ||||||
| 10. Ampedus pomonae (Steph.)* | – | – | – | – | – | ед. |
| IV. Cerylonidae | ||||||
| 11. Cerylon deplanatum Gyll.** | – | – | ед. | – | – | ед. |
| 12. Cerylon ferrugineum Steph.** | – | + | + | – | – | + |
| 13. Cerylon histeroides (F.)** | ед. | – | – | – | – | ед. |
| V. Cucujidae | ||||||
| 14. Cucujus haematodes (Er.)** | – | – | – | – | – | ед. |
| VI. Erotylidae | ||||||
| 15. Dacne bipustulata (Thunbg.) | + | 8.57 | 2.72 | + | – | 2.90 |
| 16. Tritoma subbasalis (Rtt.) | – | – | – | – | – | 1.30 |
| VII. Latridiidae | ||||||
| 17. Enicmus fungicola Thoms.* | ед. | ед. | ед. | – | – | ед. |
| 18. Enicmus rugosus (Herbst.)* | 2.50 | 2.85 | 1.36 | 1.70 | – | 2.60 |
| 19. Latridius consimilis (Mannh.) | ед. | 5.00 | 1.36 | 1.70 | – | 1.30 |
| 20. Stephostethus pandellei (Bris.)* | – | – | – | – | 2.40 | ед. |
| VIII. Nitidulidae | ||||||
| 21. Epuraea variegata (Herbst.) | 2.50 | 3.33 | 4.18 | 2.85 | 1.42 | 2.60 |
| IX. Monotomidae | ||||||
| 22. Rhizophagus dispar (Pk.)** | – | – | – | – | – | + |
| 23. Rhizophagus parvulus (Pk.)** | – | + | + | + | – | 1.20 |
| X. Ciidae | ||||||
| 24. Cis boleti (Scop.) | 41.25 | 28.50 | 46.36 | 52.85 | 37.14 | 33.89 |
| 25. Cis comptus (Gyll.) | 17.50 | 15.00 | 22.27 | 20.00 | 17.14 | 20.33 |
| 26. Cis fissicornis (Mel.) | 12.50 | 12.85 | 28.78 | 8.33 | 21.42 | 14.50 |
| 27. Cis hispidus (Gyll.) | 26.25 | 8.57 | 11.36 | 15.00 | 30.00 | 26.44 |
| 28. Cis jacquemarti (Mel.) | – | 6.42 | – | – | – | 8.64 |
| 29. Cis setiger (Oliv.) | 18.75 | 10.71 | 10.75 | 2.85 | 42.85 | 26.77 |
| 30. Octothemnus glabriculus (Gyll.) | 76.25 | 71.43 | 67.27 | 68.33 | 21.42 | 55.93 |
| 31. Rhopalodontus strandi (Lonse)* | – | – | + | + | + | + |
| 32. Sulcacis nitidus (F.) | 100.00 | 90.00 | 77.72 | 91.67 | 94.28 | 82.60 |
| 33. Sulcacis fronticornis (Pz.) | 21.25 | 9.28 | 10.75 | 3.33 | – | 10.51 |
| XI. Colydiidae | ||||||
| 34. Bitoma crenata (F.)** | ед. | ед. | ед. | – | – | ед. |
| XII. Mycetophagidae | ||||||
| 35. Mycetophagus quadripustulatus (L.)* | – | ед. | – | – | – | + |
| 36. Mycetophagus piceus L.* | ед. | ед. | ед. | – | + | + |
| XIII. Melandryidae | ||||||
| 37. Dircaea quadriguttata (Pz.)** | – | – | ед. | – | – | – |
| 38. Orchesia fasciata (Oliv.)** | – | – | + | – | – | – |
| 39. Orchesia fusiformis (Solsky) | – | – | 2.27 | ед. | – | 3.05 |
Примечания. * Жуки найдены только в фазе имаго. ** Жуки, личинки которых развиваются, преимущественно, в мицелиальном слое грибов, их имаго иногда встречаются на плодовых телах (встречаемость этих видов указана по имаго на грибах). ед. – единичные находки на (в) плодовых телах, “+” – встречаемость менее 0.50. Виды грибов рода Trametes: gibb – gibbosa, hir – hirsuta, ochr – ochracea, pub – pubescens, suav – suaveolens, ver – versicolor.
Динамику энтомокомплекса при изменении состояния и степени разрушения плодовых тел (микросукцессии) отражает схема на рис. 3.
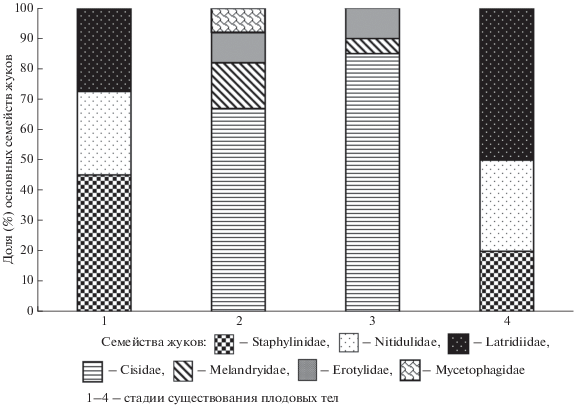
На растущих, живых, спороносящих грибах жуки редки и встречаются только во взрослой стадии. Доминируют стафилиниды Scaphisoma agaricinum, S. inopinatum, S. bipustulatus, довольно обычны скрытники Enicmus rugosus, Stephostethus pandellei, иногда Enicmus fungicola, могут встречаться лейодиды A. axillaris, гладкотелы Cerylon ferrugineum, C. histeroides и монотомиды Rh. parvulus.
Формирование комплекса разрушителей плодовых тел наиболее активно происходит на 2 стадии, когда в массе появляются трутовиковые жуки Cis boleti, C. comptus, C. fissicornis, C. hispidus, C. setiger, O. glabriculus, S. affinis, несколько реже C. jacquemarti, S. fronticornis. В это же время поселяются грибовик D. bipustulata и тенелюб O. fusiformis. Жуки-грибоеды M. piceus, очень редко M. quadripustulatus, на стадии имаго проходят лишь дополнительное питание, но не развиваются в этих грибах.
Основной комплекс обитателей приурочен к мертвым плодовым телам 3 стадии, когда в большом числе встречаются Ciidae, проходящие весь жизненный цикл в грибах. Лимитирующим фактором для развития подавляющего большинства циид являются высокая влажность субстрата, ослизнение и плесневение плодовых тел. Кроме вышеперечисленных жуков семейства Ciidae, здесь развиваются уже упомянутые Erotylidae и Melandryidae, которые совместно с трутовиковыми жуками доводят плодовые тела почти до полного разрушения.
В мертвых, сильно разрушенных, влажных и обычно заплесневелых базидиомах (4 стадия) могут поселяться скрытники Latridius consimilis, S. pandellei, стафилиниды A. inflata, C. silphoides и блестянки E. variegata.
Насекомые, характерные для 1 стадии, не участвуют в деструкции плодовых тел; многих из них привлекает обилие пищи в виде грибных спор, являющихся богатым источником азота. Попутно они разносят споры, способствуя расселению грибов в новые субстраты, что, возможно, является неотъемлемой частью их поведения (Инголд, 1957; Красуцкий, 2005). Вторая группа жуков – активных деструкторов плодовых тел (энтомокомплексы 2 и особенно 3, 4 стадий) – играет важную роль в утилизации значительной части грибной органики, обеспечивая поступление переработанного вещества грибов в детритную пищевую цепь.
ОБСУЖДЕНИЕ
С учетом экологической специализации и трофики насекомых сообщество жесткокрылых, связанных с грибами рода Trametes, может быть представлено следующим образом.
Группа специализированных мицетобионтов включает 22 вида жуков из 5 семейств. Открытоживущими обитателями грибов, типичными мицетофагами с широким кругом пищевых объектов, личинки которых развиваются, как правило, на спороносном слое (гимении) за счет питания трубочками гименофора и спорами, являются стафилиниды S. agaricinum, S. inopinatum и S. bipustulatus. Но грибы рода Trametes для имаго этих жуков служат дополнительным пищевым ресурсом, обычно стафилиниды указанных видов предпочитают живые, обильно спороносящие плодовые тела Fomes fomentarius (L.: Fr.) Fr., Fomitopsis pinicola (Sw.: Fr.) P. Karst., Ganoderma applanatum (Pers.) Pat., Inocutis rheades (Pers.) Bond et Sing. (Красуцкий, 2005). Имаго стафилинов O. maxillosus, L. lunulatus, L. trimaculatus на живых плодовых телах грибов рода Trametes (также дополнительном источнике питания) встречаются реже, а развиваются, главным образом, в грибах порядков Agaricales, Boletales, Tricholomatales, Russulales, Hymenochaetales (Никитский и др., 1996; Nikitsky, Schigel, 2004; Красуцкий, 2005). Что касается лейодид A. axillaris, обнаруженных нами только на стадии имаго, то они развиваются в миксомицетах Fuligo septica (L.) Wiggers., Reticularia lycoperdon Ehrenb., Stemonitis fusca Roth., S. axifera (Bull.) Macbr. (Никитский и др., 1996).
Жуки-грибоеды M. piceus и M. quadripustulatus, будучи специализированными скрытноживущими мицетобионтами, полными мицетофагами-полифагами, на грибах рода Trametes редки и лишь дополнительно питаются на них.
Таким образом, основу комплекса скрытноживущих специализированных мицетобионтов грибов рода Trametes составляют два вида грибовиков, 9 видов трутовиковых жуков и один вид тенелюбов. Поскольку именно эти жуки наиболее активно участвуют в деструкции плодовых тел, дадим более подробную характеристику отдельных видов.
Семейство Erotylidae – грибовики
1. Dacne bipustulata – может развиваться в мертвых плодовых телах T. hirsuta, T. ochracea, T. versicolor, но более характерен для энтомокомплекса Daedaleopsis tricolor (Bull.) Bond. et Sing. (Kп = 0.13), Piptoporus betulinus (Bull.: Fr.) P. Karst. (Kп = 0.40), Laetiporus sulphureus (Bull.: Fr.) Murr., грибов родов Inonotus, Inocutis, Lentinus (Kп = 0.18), Pleurotus (Kп = 0.22), Polyporus (Kп = 0.10) и др. (Никитский и др., 1996; Nikitsky, Schigel, 2004; Красуцкий, 2005, 2006, 2016).
2. Tritoma subbasalis – в своем развитии связан исключительно с грибами семейства Polyporaceae и, кроме грибов рода Trametes (Kп = 0.72), заселяет Lenzites betulinus (Fr.) Fr. (Kп = 0.12) и грибы рода Daedaleopsis (Kп = 0.16).
Семейство Ciidae – трутовиковые жуки
1. Cis boleti – особенно предпочитает грибы рода Trametes (Kп = 0.96), но развивается и в грибах L. betulina (Kп = 0.04) (рис. 4). На других территориях довольно активно заселяет Bjerkandera adusta (Willd.: Fr.) P. Karst., Cerrena unicolor (Bull.: Fr.) Murr., грибы рода Daedaleopsis (Никитский и др., 1996; Nikitsky, Schigel, 2004; Красуцкий, 2005, 2007; Власов, Никитский, 2015).
Рис. 4.
Пищевые преферендумы личинок жуков-мицетобионтов рода Cis Latr.: 1 – С. boleti, 2 – C. comptus, 3 – C. fissicornis, 4 – C. hispidus, 5 – C. jacquemarti, 6 – C. setiger.
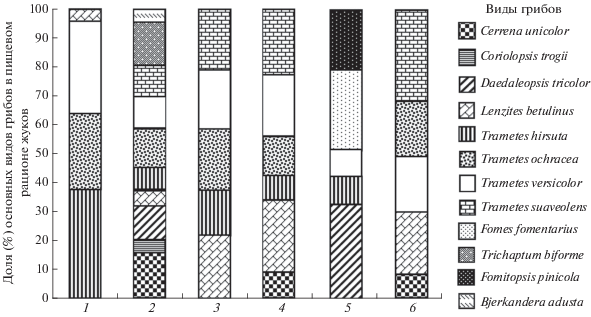
2. C. comptus – полизональный вид с широкой полифагией. Наряду с грибами рода Trametes (Kп = 0.50) заселяет C. unicolor (Kп = 0.18), D. tricolor (Kп = 0.10), T. biforme (Kп = 0.12), реже L. betulinus (Kп = 0.05), Coriolopsis (Trametes) trogii (Berk.) Domanski (Kп = 0.02) и B. adusta (Kп = 0.03) (рис. 4).
3. C. fissicornis – кроме грибов рода Trametes (Kп = 0.78) заселяет L. betulinus (Kп = 0.22) (рис. 4), развивается и в других грибах семейства Polyporaceae (Компанцев, 1984; Никитский и др., 1996; Nikitsky, Schigel, 2004; Красуцкий, 2005; Власов, Никитский, 2015).
4. C. hispidus – специализирован на грибах семейства Polyporaceae (рис. 4); развивается, вероятно, в плодовых телах всех видов рода Trametes, а также в L. betulinus (Kп = 0.22) и C. unicolor (Kп = = 0.09).
5. C. jacquemarti – типичный обитатель дереворазрушающих базидиальных грибов с широким кругом пищевых объектов. Помимо грибов рода Trametes (T. hirsuta, T. versicolor) заселяет грибы рода Daedaleopsis, особенно D. tricolor (Kп = 0.32), многолетние базидиомы F. fomentarius (Kп = 0.28), F. pinicola (Kп = 0.21), G. applanatum (Pers.) Pat., (Компанцев, 1984; Никитский и др., 1996; Nikitsky, Schigel, 2004; Красуцкий, 2005, 2006; Власов, Никитский, 2015).
6. C. setiger – заселяет исключительно грибы семейства Polyporaceae, развивается, вероятно, в плодовых телах всех видов рода Trametes, а также в грибах L. betulinus (Kп = 0.21) и C. unicolor (Kп = = 0.08).
7. O. glabriculus – развивается, вероятно, в плодовых телах всех видов рода Trametes и некоторых других представителей семейства Polyporaceae, особенно, L. betulinus (Kп = 0.11), заселяет многолетние базидиомы F. pinicola (Kп = 0.06) (рис. 5), а также (в Западной Сибири) – G. applanatum, однолетние плодовые тела B. adusta и Stereum hirsutum (Willd.: Fr.) Fr. (Красуцкий, 2005), на территории европейской части России – C. trogii (Nikitsky, Schigel, 2004; Власов, Никитский, 2015).
Рис. 5.
Пищевые преферендумы личинок жуков-мицетобионтов родов Sulcacis Dury, Octothemnus Mel., Rhopalodontus Mel.: 1 – S. fronticornis, 2 - S. nitidus, 3 – O. glabriculus, 4 – Rh. strandi, 5 – S. bidentulus (приведен на рисунке в качестве примера вида – строгого монофага).
8. Rh. strandi – единственный представитель трутовиковых жуков, который на Южном Урале и в Западной Сибири нами отмечен в грибах рода Trametes только на имагинальной стадии. Развивается чаще в грибах F. fomentarius (Kп = 0.91), реже – Trichaptum biforme (Fr.) Ryv. (Kп = 0.09) (рис. 5), на территории европейской части России заселяет еще и C. trogii, L. betulinus, L. sulphureus, P. betulinus, G. applanatum, I. rheades (Компанцев, 1984; Никитский и др., 1996; Nikitsky, Schigel, 2004; Цинкевич, 2004; Власов, Никитский, 2015), а в Западной Сибири кроме них – D. confragosa (Bolt. Fr.) Schroet., D. tricolor (Красуцкий, 2005, 2006).
9. S. nitidus – массовый вид, развивающийся в различных древесных грибах. Заселяет все виды рода Trametes, а кроме них – L. betulinus (Kп = 0.11), D. tricolor (Kп = 0.03). Встречается в грибах Pycnoporus cinnabarinus (Jack.: Fr.) P. Karst. (на рис. 5 не отражено из-за малого числа исследованных плодовых тел этого редкого на Урале вида).
10. S. fronticornis – более редкий вид рода Sulcacis, наряду с грибами рода Trametes заселяющий другие виды семейства Polyporaceae – L. betulinus (Kп = 0.08), C. trogii (Kп = 0.04), а также C. unicolor из семейства Cerrenaceae (Kп = 0.09) (рис. 5).
Семейство Melandryidae – тенелюбы
1. Orchesia fusiformis – чаще всего развивается в мертвых плодовых телах T. ochracea и T. versicolor, на территории Западной Сибири заселяет наряду с ними также C. unicolor и L. betulinus, а в европейской части России – Inonotus radiatus (Sow.: Fr.) P. Karst., T. hirsuta, T. ochracea, T. pubescens (Никитский и др., 1996; Nikitsky, Schigel, 2004; Красуцкий, 2005).
Группа эврибионтов объединяет в своем составе 17 видов из 9 семейств.
Облигатными мицетофагами являются 12 видов из 7 семейств:
– стафилины C. silphoides развиваются в гнилых плодовых телах (обнаружены также в грибах Lentinus cyathiformis (Schaeff.) Bres., Pleurotus pulmonarius (Fr.) Kumm.), под мертвой корой деревьев, в органических остатках в почве, в навозе, где питаются грибным мицелием;
– гладкотелы C. deplanatum, C. ferrugineum, C. histeroides встречаются, главным образом, под гнилой корой деревьев в мицелиальном слое не только грибов рода Trametes, но и многих других грибов, вызывающих белую гниль;
– скрытники E. fungicola, E. rugosus, L. consimilis, S. pandellei связаны, в основном, с миксомицетами, но могут развиваться в разлагающихся органических остатках (гниющая древесина, сено, подстилка, труха дупел, старые заплесневелые грибы и др.) за счет низших грибов, имаго часто посещают спороносящие базидиомы различных ксилотрофных грибов;
– блестянки E. variegata развиваются на заплесневелых грибах рода Trametes и многих других грибах, в том числе и напочвенных, на гниющих органических субстратах за счет плесневых грибов, а также на забродившем соке лиственных деревьев (Никитский и др., 1996; Никитский, Татаринова, 2003);
– монотомиды Rh. parvulus обитают в мицелиальном слое грибов T. versicolor, T. ochracea, T. hirsuta, T. pubescens, развиваются за счет мицелия этих и некоторых других грибов, например, F. fomentarius, S. hirsutum (Benick, 1952; Никитский и др., 1996; Nikitsky, Schigel, 2004; Цинкевич, 2004; Красуцкий, 2005).
– узкотелки B. crenata обычно развиваются под отмершей корой деревьев за счет некоторых аскомицетов, дейтеромицетов и ксилотрофных базидиомицетов из родов Trametes, Pleurotus, F. fomentarius, F. pinicola, Lentinus strigosus Fr., T. biforme, а также Schizophyllum commune (Fr.) Fr., L. betulinus, P. cinnabarinus, C. unicolor, P. betulinus (Benick, 1952; Никитский и др., 1996; Nikitsky, Schigel, 2004; Красуцкий, 2005), C. trogii, D. tricolor, D. septentrionalis (P. Karst.) Niemela.
– тенелюбы O. fasciata редко встречаются в мицелиальном слое грибов T. ochracea, их личинки развиваются в плодовых телах грибов рода Trichaptum на хвойных, а также Hapalopilus rutilans (Pers.: Fr.) Murill и I. rheades на лиственных (Компанцев, 1984; Никитский и др., 1996; Красуцкий, 2005).
Миксофаги в энтомокомплексе грибов рода Trametes представлены тремя видами – C. haematodes (Cucujidae), Rh. dispar (Rhizophagidae) и D. quadriguttata (Melandryidae). Все они являются обитателями подкорного пространства деревьев с грибами аскомицетами, дейтеромицетами и мицелием многих базидиомицетов.
Для C. haematodes характерны сапроксилофагия и мицетофагия с элементами хищничества и некрофагии. Rh. dispar нередко хищничает в ходах короедов на ели, а на лиственных проявляет себя как мицетофаг, используя в пищу грибы-дейтеромицеты (Никитский и др., 1996). D. quadriguttata развивается в белых гнилях лиственных за счет мицелия грибов C. trogii, Pleurotus calyptratus (Lindbl. ap Fr.) Sacc., возможно, T. ochracea и, таким образом, является мицетофагом и сапроксилофагом.
К факультативным мицетофагам относятся два вида: A. inflata (Staphylinidae) и A. pomonae (Elateridae).
Первый живет и развивается под корой и в гнилой древесине, на вытекающем соке деревьев, на падали, под опавшей листвой, в плазмодиях миксомицетов F. septica (Никитский и др., 1996), в старых плодовых телах некоторых ксилотрофных грибов (Onnia tomentosa (Fr.) Karst.), в том числе и в старых грибах рода Trametes (Benick, 1952; Никитский и др., 1996; Цинкевич, 2004; Красуцкий, 2005).
Второй вид развивается в белых и бурых гнилях хвойных и лиственных деревьев (Никитский и др., 1996), на стадии имаго встречается на генеративных и вегетативных органах различных растений и на плодовых телах некоторых древесных грибов (Красуцкий, 2005).
ЗАКЛЮЧЕНИЕ
Таким образом, основу мицетофильного сообщества грибов рода Trametes составляют 9 видов трутовиковых жуков (Ciidae), 1 вид грибовиков (Erotylidae) и 1 вид тенелюбов (Melandryidae). Самые высокие значения коэффициентов предпочтения этих грибов на Южном Урале и в других регионах (Красуцкий, 2005) у Octotemnus glabriculus, Sulcacis nitidus, Cis hispidus, C. setiger, C. boleti (Ciidae), Tritoma subbasalis (Erotylidae) и Orchesia fusiformis (Melandryidae). Сходная ситуация наблюдается и на территории европейской части России (Компанцев, 1984; Никитский и др., 1996; Nikitsky, Schigel, 2004; Власов, Никитский, 2015). В белых гнилях древесины лиственных пород, в мицелиальном слое грибов рода Trametes формируется другой комплекс видов: Cerylon deplanatum, C. ferrugineum, C. histeroides (Cerylonidae), Rhizophagus dispar, Rh. parvulus (Monotomidae), Cucujus haematodes (Cucujidae), Bitoma crenata (Colydiidae), Dircaea quadriguttata и Orchesia fasciata (Melandryidae). Некоторые эврибионтные жесткокрылые – Cilea silphoides (Staphylinidae), Enicmus fungicola, E. rugosus, Latridius consimilis (Latridiidae), Epuraea variegata (Nitidulidae) – могут использовать траметоидные грибы в качестве дополнительного пищевого ресурса. В целом следует отметить, что общий список жуков, связанных с грибами рода Trametes, существенно пополнился за счет более детального изучения специализированных мицетобионтов-мицетофагов и трофических связей эвритопных видов, заселяющих, помимо древесины, подстилки и почвы, базидиомы этих грибов. Более полно изучен микросукцессионный процесс, характеризующийся сменой комплексов обитателей и посетителей плодовых тел грибов рода Trametes в связи с изменением их состояния и степени разрушения.
В качестве комментария отметим интересную особенность, позволяющую, на наш взгляд, уточнить систематическое положение трутовика Трога (Polyporaceae). Ранее этот вид входил в состав рода Funalia Pat., затем был помещен в род Coriolopsis Murrill и, позднее, – в Trametes. В работах разных авторов (и микологов, и энтомологов) можно увидеть и разную интерпретацию его родового статуса. Изучение комплекса специализированных мицетобионтов трутовика Трога (рис. 5) показало, что только в плодовых телах этого гриба развивается очень интересный вид трутовиковых жуков – Sulcacis bidentulus (Rosenh.). О строгой монофагии S. bidentulus не раз сообщали Беник (Benick, 1952), Никитский с соавторами (Никитский и др., 1996; Власов, Никитский, 2015), автор настоящей статьи (Красуцкий, 1990, 1994, 1995, 2005). Этот факт позволяет считать более целесообразным помещение трутовика Трога в отдельный род (Coriolopsis или Funalia) и служит примером того, как насекомые могут “помочь” в уточнении систематического положения некоторых “кочующих” грибов.
Список литературы
Аксентьев С.И., Участнова Л.Н., 1986. Проблема симбиоза жуков-ксилофагов и амброзиальных грибов // Научные доклады высшей школы. Биологические науки. № 8 (272). С. 5–22.
Богданов И.И., Чачина С.Б., Коралло Н.П., Дмитриев В.В., 2006. Новый паразитологический индекс для оценки особенностей связей паразита с хозяином и биотопом хозяина // Паразитология. Т. 40. № 2. С. 105– 112.
Бондарцева М.А., 1998. Определитель грибов России. Порядок Афиллофоровые. Вып. 2. Семейства альбатрелловые, апорпиевые, болетопсиевые, бондарцевиевые, ганодермовые, кортициевые (виды с порообразным гименофором), лахнокладиевые (виды с трубчатым гименофором), полипоровые (роды с трубчатым гименофором), пориевые, ригидопоровые, феоловые, фистулиновые). СПб.: Наука. 391 с.
Власов Д.В., Никитский Н.Б., 2014. Фауна жуков-блестянок (Coleoptera, Cucujoidea, Nitidulidae) Ярославской области // Бюллетень Московского общества испытателей природы. Отдел Биологический. Т. 119. Вып. 6. С. 29–35.
Власов Д.В., Никитский Н.Б., 2015. Фауна трутовиковых жуков (Coleoptera, Tenebrionoidea, Ciidae) Ярославской области // Бюллетень Московского общества испытателей природы. Отдел Биологический. Т. 120. Вып. 3. С. 3–11.
Власов Д.В., Никитский Н.Б., 2017. Фауна жуков-челновидок (Coleoptera, Staphylinidae) Ярославской области с указаниями новых и малоизвестных жесткокрылых из некоторых семейств // Бюллетень Московского общества испытателей природы. Отдел Биологический. Т. 122. Вып. 3. С. 8–18.
Власов Д.В., Никитский Н.Б., Салук С.В., 2018. Фауна жуков-скрытников (Coleoptera, Latridiidae) Ярославской области // Бюллетень Московского общества испытателей природы. Отдел Биологический. Т. 123. Вып. 4. С. 34–39.
Ердаков Л.Н., Ефимов В.И., Галактионов Ю.К., Сергеев В.Е., 1978. Количественная оценка верности местообитаниям // Экология. № 3. С. 105–107.
Инголд Ц., 1957. Пути и способы распространения грибов. М.: ИЛ. 183 с.
Компанцев А.В., 1982. Морфо-экологические особенности личинок жуков семейства Erotylidae (Coleoptera) – обитателей плодовых тел высших грибов // Морфо-экологические адаптации насекомых в наземных сообществах. М.: Наука. С. 81–91.
Компанцев А.В., 1984. Комплексы жесткокрылых, связанные с основными дереворазрушающими грибами в лесах Костромской области // Животный мир Южной тайги. М.: Наука. С. 191–196.
Компанцева Т.В., 1987. Экологические особенности ксилофильных и мицетофильных жуков-чернотелок (Coleoptera, Tenebrionidae) // Экология и морфология насекомых – обитателей грибных субстратов. М.: Наука. С. 45–56.
Красуцкий Б.В., 1990. Сообщества жесткокрылых, связанные с основными дереворазрушающими грибами Припышминских боров Западной Сибири // Эколого-флористические исследования по споровым растениям Урала. Свердловск: УрО АН СССР. С. 57–67.
Красуцкий Б.В., 1994. Экологическая классификация жесткокрылых – мицетобионтов дереворазрушающих базидиальных грибов // Экология. № 1. С. 71–79.
Красуцкий Б.В., 1995. Жесткокрылые – мицетобионты дереворазрушающих базидиальных грибов в подтаежных лесах Западной Сибири // Энтомологическое обозрение. Т. LXXIV. Вып. 3. С. 542–550.
Красуцкий Б.В., 1996. Мицетофильные жесткокрылые Урала и Зауралья. Т. I. Краткое иллюстрированное руководство к определению по имаго наиболее обычных в энтомокомплексах дереворазрушающих базидиальных грибов видов жесткокрылых. Екатеринбург: Изд-во “Екатеринбург”. 146 с.
Красуцкий Б.В., 1996а. Жесткокрылые – мицетобионты (Coleoptera) основных дереворазрушающих грибов лесостепного Зауралья // Энтомологическое обозрение. Т. LXXV. Вып. 2. С. 274–277.
Красуцкий Б.В., 1997. Жесткокрылые (Сoleoptera) – мицетобионты основных дереворазрушающих грибов южной подзоны Западно-Сибирской тайги // Энтомологическое обозрение. Т. LXXVI. Вып. 2. С. 302–308.
Красуцкий Б.В., 1997а. Жесткокрылые-мицетобионты (Coleoptera) основных дереворазрушающих грибов подзоны средней тайги Западной Сибири // Энтомологическое обозрение. Т. LXXVI. Вып. 4. С. 720–775.
Красуцкий Б.В., 2000. Сообщества жесткокрылых, связанных с основными дереворазрушающими грибами Челябинской области // Труды института биоресурсов и прикладной экологии. Оренбург: Изд-во ОГПУ. Вып. 1. С. 76–89.
Красуцкий Б.В., 2001. Мицетофильные жесткокрылые (Insecta, Coleoptera) Ильменского заповедника. Система “Грибы-насекомые” // Изучение беспозвоночных животных в заповедниках. Проблемы заповедного дела. Вып. 10. С. 126–150.
Красуцкий Б.В., 2005. Мицетофильные жесткокрылые Урала и Зауралья. Т. II: Система “Грибы-насекомые”. Челябинск: ОАО “Челябинский дом печати”. 213 с.
Красуцкий Б.В., 2006. Жесткокрылые (Coleoptera), связанные с березовым трутовиком Piptoporus betulinus (Bull.: Fr.) P. Karst. (Basidiomycetes, Aphyllophorales) в лесах Урала и Зауралья // Энтомологическое обозрение. Т. LXXXV. Вып. 4. С. 758–773.
Красуцкий Б.В., 2007. Жесткокрылые (Coleoptera), связанные с трутовиком Daedaleopsis confragosa (Bolton.: Fr.) J. Schrot (Basidiomycetes, Aphyllophorales) в лесах Урала и Зауралья // Энтомологическое обозрение. Т. LXXXVI. Вып. 2. С. 289–305.
Красуцкий Б.В., 2007а. Жесткокрылые (Coleoptera), связанные с окаймленным трутовиком Fomitopsis pinicola (Sw.: Fr.) Karst. (Basidiomycetes, Aphyllophorales) в лесах Урала и Зауралья // Энтомологическое обозрение. Т. LXXXVI. Вып. 3. С. 532–545.
Красуцкий Б.В., 2010. Жесткокрылые (Coleoptera), связанные с трутовиком Trichaptum biforme (Fr. in Klotzsch) (Basidiomycetes, Aphyllophorales) в лесах Урала и Зауралья // Энтомологическое обозрение. Т. LXXXIX. Вып. 2. С. 367–379.
Красуцкий Б.В., 2013. Жесткокрылые (Coleoptera, Insecta) в энтомокомплексах ксилотрофных базидиальных грибов порядка Polyporales подтаежных лесов Западной Сибири // Экологический мониторинг и биоразнообразие. № 1(8). С. 50–54.
Красуцкий Б.В., 2013а. Жесткокрылые (Coleoptera, Insecta) в энтомокомплексах ксилотрофных базидиальных грибов порядка Fomitopsidales (Basidiomycetes) подтаежных лесов Западной Сибири // Вестник Ишимского государственного педагогического института. № 6(12). С. 32–38.
Красуцкий Б.В., 2014. Жесткокрылые (Coleoptera, Insecta) в энтомокомплексах ксилотрофных базидиальных грибов порядка Hyphodermatales (Basidiomycetes) подтаежных лесов Западной Сибири // Экологический мониторинг и биоразнообразие. № 2(9). С. 82–86.
Красуцкий Б.В., 2016. Мицетофильные жесткокрылые Южного Урала // Актуальные вопросы современного естествознания Южного Урала (к 170-летию со дня рождения Ю.К. Шелля): Материалы II Всерос. научно-практ. конф. с междунар. участием, 7 декабря 2016 г., Челябинск / под ред. Меркер В.В., Гашек В.А., Попкова П.Н. Челябинск: изд-во Челябинского гос. ун-та. С. 40–56.
Красуцкий Б.В., 2018. Материалы к фауне жуков Челябинского городского бора, связанных с ксилотрофными базидиальными грибами // Фауна Урала и Сибири. Екатеринбург: ИЭРиЖ УрО РАН. № 1. С. 97–103.
Криволуцкая Г.О., 1992. Сем. Erotylidae – грибовики // Определитель насекомых Дальнего Востока СССР. Т. III: Жесткокрылые, или жуки. Ч. 2. СПб.: Наука. С. 285–303.
Никитский Н.Б., 1992. Сем. Tetratomidae – тетратомиды // Определитель насекомых Дальнего Востока СССР. Т. III: Жесткокрылые, или жуки. Ч. 2. СПб.: Наука. С. 426–435.
Никитский Н. Б., 1992а. Сем. Melandryidae – тенелюбы // Определитель насекомых Дальнего Востока СССР. Т. III: Жесткокрылые, или жуки. Ч. 2. СПб.: Наука. С. 435–474.
Никитский Н.Б., Компанцев А.В., 1995. Новые виды жуков-грибовиков (Coleoptera, Erotylidae) с Дальнего Востока России с замечаниями по распространению и биологии других видов // Зоологический журнал. Т. 74. Вып. 6. С.83–92.
Никитский Н.Б., Осипов И.Н., Чемерис М.В., Семенов В.Б., Гусаков А.А., 1996. Жесткокрылые-ксилобионты, мицетобионты и пластинчатоусые Приокско-Террасного биосферного заповедника (с обзором фауны этих групп Московской области. Сборник трудов Зоологического музея МГУ. Т. XXXVI. М.: Изд-во МГУ. 198 с.
Никитский Н.Б., Татаринова А.Ф., 2003. Фауна и экология ксилофильных жуков-блестянок (Coleoptera, Nitidulidae) европейского Северо-Востока России // Бюллетень Московского общества испытателей природы. Отдел Биологический. Т. 108. Вып. 5. С. 28–32.
Определитель насекомых Дальнего Востока СССР., 1992. Т. III. Жесткокрылые, или жуки. Ч. 2. СПб.: Наука. 704 с.
Песенко Ю.А., 1982. Принципы и методы количественного анализа в фаунистических исследованиях. М.: Наука. 287 с.
Степанова-Картавенко Н.Т., 1967. Афиллофоровые грибы Урала. Труды Института экологии растений и животных Уральского филиала АН СССР. Свердловск. Вып. 50. 295 с.
Цинкевич В.А., 2004. Жесткокрылые (Coleoptera) – обитатели плодовых тел базидиальных грибов (Basidiomycetes) запада лесной зоны Русской равнины (Беларусь) // Бюллетень Московского общества испытателей природы. Серия Биологическая. Т. 109. Вып. 4. С. 17–25.
Яковлев Е.Б., 1986. Особенности консортивных связей насекомых с макромицетами // Микология и фитопатология. Т. 20. Вып. 3. С. 185–191.
Яковлев Е.Б., 1994. Двукрылые Палеарктики, связанные с грибами и миксомицетами. Петрозаводск: Институт леса КНЦ РАН. 128 с.
Alexander K.N. A., 2002. The invertebrates of living and decaying timber in Britain and Ireland – a provisional annotated checklist // English Nature Research Reports. № 467. P. 1–142.
Benick L., 1952. Pilzkafer und Kaferpilz. Okologishe und statistische Untersuchungen // Acta zoologica Fennica. Bd. 70. S. 1–250.
Jablokoff-Khnzorian S.M., 1975. Etude sur les Erotylidae (Coleoptera) Palaearctiques // Acta zoologica Cracoviensia. Tom XX. № 8. P. 201–249.
Kotiranta H., Mukhin V.A., Ushakova N., Dai Y.-C., 2005. Polypore (Aphyllophorales, Basidiomycetes) studies in Russia. 1. South Ural. Annales Botanici Fennici. V. 42. P. 427–451.
Komonen A., 2001. Structure of insect communities inhabiting old-forest specialist bracket fungi // Ecological Entomology. V. 26. P. 63–75.
Komonen A., 2005. Occurrence and abundance of fungus-dwelling beetles (Ciidae) in boreal forests and clearcuts: habitat associacions at two spatial scales // Animal Biodiversity and Conservation. V. 28. P. 137–147.
Komonen A., 2008. Colonization experiment of fungivorous beetles (Ciidae) in lake-island system // Entomologisk Tidskrift. V. 129. № 3. P. 141–145.
Elton C., 1927. Enimal Ecology. London, England: Sidgwick and Jackson. 256 p.
Nikitsky N.B., Schigel D.S., 2004. Beetles in Polypores of the Moscow region: checklist and ecological notes // Entomologica Fennica. № 15. P. 6–22.
Orledge G.M., Reynolds S.E., 2005. Fungivore host-use group from claster analysis: pattern of utilization of fungal fruiting bodies by ciid beetles // Ecological Entomology. V. 30. P. 620–641.
Paine R.T., 1988. Road maps of interactions or grist for theoretical development? // Ecology. V. 69. № 6. P. 1648–1654.
Reibnitz J., 1999. Verbreitungund Lebensträume der Baumschwammfresser Südwestdeutchlands (Coleoptera, Cisidae) // Mitteilungen Entomologisher Verien Stuttgart. V. 34. S. 1–76.
Schigel D.S, 2004. Polyporus and associated beetles of the North Karelian Biosphere Reserve, eastern Finland. Karstenia. V. 44. P. 35–56.
Schigel D.S., 2005. Polypore-inhabiting beetles of four protected forests in South Häme, Central Finland. Sahlbergia. № 10. P. 59–62.
Schigel D.S., 2006. Polyporus of western Finnish Lapland and seasonal dynamics of polypore beetles. Karstenia. V. 46. P. 37–64.
Schigel D.S., 2009. Polypore assemblages in boreal old-growth forests, and notes on associated beetles in Finland // Polypore assemblages in boreal old-growth forests, and associated beetles (Academic dissertation). Helsinki. P. 1–48.
Schigel D.S., 2011. Polypore-beetle associated in Finland // Annales Zoologici Fennici. V. 48. № 6. P. 319–348.
Schigel D.S., 2011a. Fungus-beetle food web pattern in boreal forests // Russian Entomological Journal. V. 20. № 2. P. 141–150.
Дополнительные материалы отсутствуют.
Инструменты
Зоологический журнал