

Зоологический журнал, 2021, T. 100, № 7, стр. 790-801
Причины снижения численности западных подвидов гуменника (Anser fabalis fabalis и Anser fabalis rossicus): о чем говорят данные кольцевания?
И. Н. Панов a, b, *, К. Е. Литвин a, **, Б. С. Эббинге c, С. Б. Розенфельд a, ***
a Институт проблем экологии и эволюции РАН
119071 Москва, Россия
b Зоологический институт РАН
199034 С.-Петербург, Россия
c Вагенингенский университет, Гелдерланд
Вагенинген, Нидерланды
* E-mail: kuksha@yandex.ru
** E-mail: kelitvin@gmail.com
*** E-mail: rozenfeldbro@mail.ru
Поступила в редакцию 05.06.2020
После доработки 11.11.2020
Принята к публикации 16.11.2020
Аннотация
Проведен анализ данных возвратов колец с гуменников (Anser fabalis), помеченных с 1960 по 2013 гг. на зимовках в Нидерландах, где для существенной части птиц был указан подвид (A. f. fabalis или A. f. rossicus). Большая часть находок зарегистрирована на территории бывшего СССР, преимущественно в Российской Федерации. Проверяется гипотеза о том, что подвид A. f. fabalis подвергается большему воздействию пресса охоты, по сравнению с A. f. rossicus. Для этого проводятся сравнение сроков дожития птиц двух подвидов и анализ сезонного распределения находок окольцованных птиц. Обсуждаются в среднем меньшие сроки дожития A. f. fabalis (в сравнении с A. f. rossicus); многолетний сдвиг сроков добычи в сторону весенней охоты; а также возможная смена регионов зимовок гуменника (рост значения восточноевропейских зимовок) как еще одна причина резкого уменьшения численности зимующего A. f. fabalis в Нидерландах.
Гуменник (Anser fabalis) – наиболее широко распространенный евроазиатский охотничий вид гусей. Выделяют четыре подвида гуменника: западный тундровый (A. f. rossicus), восточный тундровый (A. f. serrirostris), лесной (A. f. fabalis) и таежный (A. f. middendorfii). Весной гуменник прилетает раньше, чем другие гуси, использует широкий спектр местообитаний; для гнездования в лесной зоне выбирает труднопроходимые местообитания с густым подростом или высокой травой близ рек. Лесные подвиды мигрируют значительно раньше тундровых и сразу приступают к размножению. Тундровые подвиды после вылупления птенцов переходят на крупные водоемы, где линяют, иногда вместе с неразмножающимися птицами. Крупных линных скоплений лесные подвиды не образуют, отлет на зимовку происходит непосредственно с мест гнездования и линьки без образования предмиграционных концентраций. Неразмножающиеся птицы часто мигрируют на линьку в тундровую зону (Литвин, 2014; Розенфельд и др., 2018; Спицын, 2019; Nilsson et al., 2010).
В единственной на данный момент сводке по миграции гуменника, основанной на данных кольцевания (Лебедева, 1979), нет подразделения материала по подвидам, поскольку подвиды в то время в данных кольцевания не указывались. В общих заключениях к статье по миграции гуменника Кищинский (1979) все же упоминает наличие разделения на подвиды и популяции и делает выводы о существовании двух основных пролетных путей: северного, от Прибалтики к Белому морю и далее на восток, и южного, который проходит от Белоруссии по центральным районам европейской части СССР через Урал на Обь и далее на восток. Полученные данные о подвидовом подразделении гусей, окольцованных в Нидерландах, дают основание для нового анализа результатов кольцевания гуменников.
Лесной гуменник (A. f. fabalis (Latham 1787)) – наиболее редкий, спорадически распространенный подвид, обитающий в таежной зоне Евразии, вплоть до Среднесибирского плоскогорья на восток. Пути миграции лесных гуменников с территории России до сих пор изучены слабо, а сложности проведения учетов затрудняют оценку численности в местах гнездования (Розенфельд и др., 2018). Известно, что гнездящиеся в зарубежной части Фенноскандии птицы зимуют почти исключительно в Дании и на юге Швеции (Nilsson et al., 1999; Nilsson, 2011). Зимовки лесных гуменников из европейской части России сосредоточены в Западной Европе, преимущественно в Нидерландах, на севере Германии и в Польше (Nilsson, 2011; Marjakangas et al., 2015). Лесные гуменники западносибирской субпопуляции, по имеющимся данным (Розенфельд, Стрельников, 2019), зимуют, в том числе, в северо-восточной Германии и северо-западной части Польши, в целом же их области зимовки в настоящее время не очерчены.
По результатам учетов на зимовках, в Западной Европе численность лесного гуменника сократилась на 80% и более, а в целом падение численности гуменника происходит в основном за счет этого подвида (CAFF, 2018; Marjakangas et al., 2015). В Нидерландах с середины 1980-х, по некоторым оценкам, отмечается 50–60 кратное снижение (Hornman et al., 2013). К настоящему времени лесной гуменник внесен в Красную книгу России только на части своего ареала (Приказ Министерства природных ресурсов и экологии Российской Федерации, 2020). Подвид в целом является объектом Международной конвенции по охране мигрирующих видов (Приложение II , http:// docs.cntd.ru/document/1 902 129) и Соглашения по охране афро-евразийских мигрирующих водно-болотных птиц (https://www.unep-aewa.org/). Предполагается, что главной причиной резкого сокращения численности популяций лесного гуменника, зимующих в Западной Европе, является весенняя охота, т.е. прямое истребление репродуктивного ядра птиц.
Несмотря на многолетние усилия по кольцеванию птиц и немалый зарубежный опыт (особенно опыт США и Канады, см. Maihguy et al., 2002; Klaassen et al., 2005; Arizaga et al., 2014; Ladin et al., 2020) использования данных кольцевания для оценок влияния охоты, в отечественной литературе такие попытки предпринимались нечасто (Паевский, 2008; Харитонов, 2017). В данной статье мы, опираясь на данные стандартного кольцевания, стремимся обсудить выживаемость, выраженную как сроки дожития, лесного гуменника в сравнении с тундровым подвидом; особенности распределения встреч подвидов гуменника в разные сезоны и различия в сроках миграции. Основная цель нашего исследования – оценка влияния пресса охоты на два подвида гусей, зимующих в Европе и гнездящихся на территории России. Найденные различия в выживаемости и сезонном распределении подвидов помогут объяснить снижение численности лесного гуменника и определить возможные меры охраны этого подвида гусей.
При этом в отличие от опыта большинства демографических исследований, на данном этапе мы в целом не используем данные кольцевания, чтобы вскрыть биологические процессы и описать биологически объясняемые параметры, но пытаемся оценить различия воздействия охоты на два подвида. В статье использованы данные, собранные за несколько десятилетий, поэтому мы не можем утверждать, что сделанные выводы в полной мере описывают современную ситуацию, однако это никак не мешает анализировать причины динамики численности птиц. К тому же представленные материалы до сих пор оставались необработанными и никогда не были опубликованы.
МАТЕРИАЛЫ И МЕТОДЫ
Для анализа были выбраны только птицы, окольцованные на зимовках в Нидерландах, где помечено подавляющее большинство тех гуменников, у которых при кольцевании был определен подвид (База данных Центра кольцевания птиц ИПЭЭ РАН; Burgers et al., 1991). Не использовали (относительно немногочисленные) данные по тундровым гуменникам, окольцованным в Германии и России (Ненецкий АО): в этих регионах кольцевание проводили в иные сезоны (миграций и размножения), что может влиять на показатели дожития окольцованных особей.
Использовали все находки (возвраты колец) мертвых или раненых птиц, связанные с деятельностью человека, за исключением возвратов колец с обстоятельствами “найдено только кольцо”, “найдена разложившейся/скелет” (с неизвестной датой гибели). Чаще всего, в случае нарушения тех или иных правил охоты, корреспонденты скрывают факт отстрела, сообщая о находке мертвой птицы или не уточняя обстоятельств. Поэтому большинство встреч, когда птица была “найдена мертвой” и с “неизвестными обстоятельствами находки”, также взяты в исследование (если не было других предпосылок к исключению из выборки, как, например, встречи за пределами области распространения в тот или иной сезон). Не рассматривали также любые находки, дата которых не указана (есть только дата отправления письма) либо точность даты превышает 91 день. Находки с точностью даты “весна (осень, лето) такого-то года” принимали в анализ. В случаях, когда был указан только период находки с точностью 91 день и меньше, за дату принимали начало указанного периода. Поскольку на данном этапе характер материала не позволяет оценить различия вероятности сообщения о находке, мы принимали ее одинаковой для обоих подвидов и в разные сезоны охоты.
Таким образом, из общего числа находок лесных гуменников, окольцованных в Нидерландах (382), в расчет взяты 353 возврата колец. Для тундрового подвида из 395 отобрано 349 возвратов. В целом в выборках для обоих подвидов существенно преобладают встречи с обстоятельствами “застрелена” (табл. 1). Большая часть находок (95.9%) зарегистрирована на территории России, остальные находки – на территории бывших республик СССР и одна – на территории бывшей ГДР.
Таблица 1.
Обстоятельства находки в возвратах колец подвидов гуменника
| Обстоятельства находки | Подвид гуменника | |||
|---|---|---|---|---|
| лесной | тундровый | |||
| n | % | n | % | |
| “Застрелена” | 326 | 92.4 | 321 | 92.0 |
| “Найдена мертвой” | 9 | 2.5 | 8 | 2.3 |
| “Обстоятельства неизвестны” | 10 | 2.8 | 8 | 2.3 |
| Другое* | 8 | 2.3 | 12 | 3.4 |
| Всего | 353 | 100 | 349 | 100 |
Данные по лесному гуменнику имеются для птиц, окольцованных только до 1989 г., включительно, по тундровому гуменнику – до 2013 г. Снижение числа возвратов колец лесных гуменников связано с резким падением численности гусей этого подвида в Нидерландах после 1989 г. (Koffijberg et al., 2011). Очевидно, что у долгоживущего вида использование более поздних данных кольцевания при прочих равных условиях может скорее занижать (но не завышать) общие результаты расчета дожития, поэтому в большинстве случаев мы используем все данные по тундровому подвиду. Чтобы убедиться, что такой подход не искажает картину результатов, провели сравнение дожития тундровых гуменников, окольцованных по 1989 г. и с 1990 г.
При сравнении дожития в разные сезоны “весенними” считали находки с датой по 24.05 включительно; “летними” – отмеченные в период с 25.05 по 25.08 включительно. Группа возвратов колец, датированных начиная с 26.08, обозначена как “осенние”. В последнюю группу вошли также три возврата колец лесного подвида со среднеазиатских зимовок (птицы, сменившие регион зимовки в последующие годы), произошедших 16 и 28 ноября и 28 декабря. Чтобы исключить влияние на показатели дожития более ранних сроков весенней миграции у лесного гуменника, провели сравнение дат встреч двух подвидов весной.
Для оценки многолетних сдвигов в сроках добычи двух подвидов использованы точные даты их обнаружения (добычи).
Для анализа пространственного распределения находок гуменников составлены карты возвратов колец для трех обозначенных выше сезонов. Территориальное положение точек добычи охотничьих видов зависит от распределения охотников не меньше, чем от плотности самих птиц в тот или иной сезон. Поэтому основной задачей картографического анализа было сравнительное описание регионов, где подвиды гуменника встречаются с охотниками, а также сезонной специфики и возможных многолетних сдвигов границ этих регионов. С целью оценить, пользуются ли гуменники в разные годы одними и теми же пролетными путями, для весеннего сезона проанализировали также данные о распределении “прямых” и “непрямых” возвратов колец. К прямым относятся находки, произошедшие в следующую за кольцеванием весну (и можно считать, что данная птица перемещается непосредственно из тех районов зимовки, где ее окольцевали); находки в последующие годы относили к непрямым находкам (из каких районов зимовок возвращается птица – неизвестно).
Для сравнения рядов дожития и дат применяли тест Манна–Уитни, для сравнения долей – критерий χ2 Пирсона. Использовали статистическую программу Past 1.5. Для оценки различий в распределении находок применяли модификацию теста Мардиа (Mardia, 1967; Batchelet, 1972), когда вычисления проводятся с усреднением совпадающих рангов, а средние координаты совокупности точек определяются в формате градусы-минуты (Robson, 1968). Программа написана James E. Hines и С.П. Харитоновым.
РЕЗУЛЬТАТЫ
Сравнение дожития птиц двух подвидов. В среднем 353 лесных гуменника с момента кольцевания прожили 1265 дней (3.47 года), 349 тундровых – в среднем 1574 дня (4.32 года). Разница в сроках значима: U = 55 570, z = 2.244, p = 0.025. Эта разница также на границе значимой, если выборку тундрового подвида формировать только из птиц, окольцованных до 1989 г.: их средний срок дожития 1615 дней, или 4.42 года (U = 43 130, z = 1.96, p = 0.05). При этом значимых различий в сроке дожития у тундровых гуменников, окольцованных до 1989 г. и начиная с 1990 г., не выявлено. В первой группе (n = 269) птицы в среднем проживали после кольцевания 1615 дней, во второй (n = 80) – 1448 дней (U = 10 650, z = 0.141, p = = 0.888). Таким образом, окольцованные лесные гуменники до момента добычи в среднем проживают значимо меньше тундровых.
95% лесных гуменников были окольцованы в период с 11 декабря по 25 февраля, 95% тундровых гуменников – со 2 декабря по 24 февраля. Средняя дата кольцевания лесных гуменников 13 января, тундровых – 11 января (U = 57 060, z = 1.69, p = 0.091); если рассматривать тундровых гуменников, окольцованных только до 1989 г., – 13 января (U = 4650, z = 0.44, p = 0.660). Соотношение взрослых и молодых птиц (на момент кольцевания) в выборках двух подвидов в высокой степени сходно. У лесного гуменника 39.7% (n = 129) особей определены как молодые, у тундрового – 38.8% (n = 130; χ2 = 0.054, p = 0.816). Имеющиеся у нас данные не позволяют делать вывод о значимых различиях в дожитии между особями, окольцованными молодыми и взрослыми. Молодые лесные гуменники в среднем проживали 1355 дней, а окольцованные взрослыми – 1273 дня (U = 12 140, z = 0.610, p = 0.543). У тундрового гуменника средние сроки дожития возрастных групп – 1538 и 1668 дней соответственно (U = 12 120, z = 1.391, p = 0.164). Такие результаты позволяют считать, что возрастной состав в момент кольцевания и различия в датах кольцевания вряд ли относятся к значимым факторам, влияющим на полученные нами различия в сроках дожития.
Самки лесного гуменника в среднем проживают 1307 дня (3.58 года), самцы – 1233 дня (3.38 года). Самки тундрового подвида в среднем проживают 1501 дня, или 4.11 года, самцы – 1643 дня, или 4.5 года. Статистических различий в дожитии половых групп внутри подвидов нет: p = 0.503 для лесного и p = 0.886 для тундрового гуменника. Различия между подвидами формируются в основном за счет меньшей продолжительности жизни самцов лесного гуменника, которые проживают существенно меньше своих тундровых сородичей: U = 13 580, z = 2.066, p = 0.039. Различия в дожитии самок незначимы; U = 12 930, z = = 1.126, p = 0.26. Таким образом, в целом наиболее уязвимой группой можно признать самцов лесного гуменника.
Добываемые весной лесные гуменники проживают с момента кольцевания в среднем 2.76 года, тундровые 4.39 года (табл. 2). Разница высоко значима: U = 11 890, z = 4.512, p < 0.00001. Средняя дата весенней добычи лесного гуменника – 30 апреля, тундрового - 1 мая, значимой разницы в этом показателе не наблюдается (p = 0.292). Таким образом, более ранние сроки пролета лесного подвида вряд ли оказывают существенное влияние на полученные нами показатели дожития в весенний период. Гуменники, встреченные в летний период, проживают в среднем меньше, чем птицы, которых отстреливают весной (несмотря на то, что с момента кольцевания проходит в среднем больше времени!). Это касается обоих подвидов (табл. 2), но значимы различия только у тундрового гуменника (U = = 3688, z = 2.917, p < 0.01). Для летних и осенних возвратов статистической разницы в дожитии между подвидами нет. Для сравнения сезонных сроков дожития полов внутри подвидов (общая статистика приведена в табл. 2), в большинстве случаев, недостаточно данных. При этом небольшой объем выборки не препятствует выявлению значимых различий в возрасте добытых летом самок и самцов тундрового гуменника (U = 156, z = = 2.136, p = 0.032), у обоих подвидов добытые летом самки проживают в среднем меньше, но у тундрового гуменника в этот сезон разница почти трехкратная (табл. 2).
Таблица 2.
Средняя продолжительность жизни (с момента кольцевания) особей двух подвидов гуменника, добытых в разные сезоны
| Группа | Параметр | Подвид гуменника | |||||
|---|---|---|---|---|---|---|---|
| лесной | тундровый | ||||||
| Сезоны находок | |||||||
| весна | лето | осень | весна | лето | осень | ||
| Самки | n | 69 | 15 | 75 | 116 | 25 | 32 |
| Дней | 903 | 870 | 1782 | 1512 | 598 | 2165 | |
| Лет | 2.47 | 2.38 | 4.88 | 4.14 | 1.64 | 5.93 | |
| Самцы | n | 69 | 23 | 90 | 114 | 18 | 39 |
| Дней | 1135 | 972 | 1375 | 1678 | 1528 | 1591 | |
| Лет | 3.11 | 2.66 | 3.77 | 4.60 | 4.19 | 4.36 | |
| Все | n | 142 | 39 | 172 | 232 | 44 | 73 |
| Дней | 1008 | 930 | 1552 | 1602 | 968 | 1850 | |
| Лет | 2.76 | 2.55 | 4.25 | 4.39 | 2.65 | 5.07 | |
Многолетние сдвиги в сроках добычи гуменников. Тогда как у лесного гуменника дата встречи не зависит от года (рис. 1, p = 0.31), у тундрового подвида средняя дата находки сдвигается на более ранний период (R = 0.21, p < 0.0001). Это происходит за счет того, что в последние три десятилетия (когда возвратов лесного гуменника практически нет) в выборке существенно преобладают птицы, застреленные в весенний период (рис. 1).
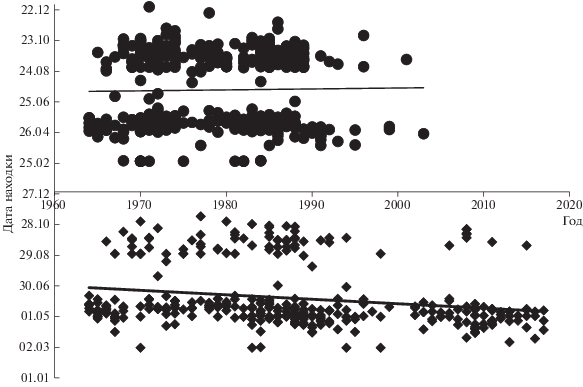
Неоднородность имеющегося для двух подвидов материала, очевидно, главная причина различий в медианном годе находки лесного и тундрового гуменника в разные сезоны. Медианный год весенних находок для лесного гуменника – 1977 г., для тундрового 1988 г. (U = 8885, z = 7.841, p < 0.000); летних возвратов колец – 1978 и 1981, соответственно (U = 613.5, z = 1.267, p = 0.205); осенних – 1979 и 1985 соответственно (U = 4874, z = 2.769, p < 0.01). Отсутствие значимых различий в медианном годе летних находок позволяет нам сравнивать районы летнего распределения подвидов без особых допущений. Не различается также медианная дата отстрела гуменников разных подвидов в период гнездования и линьки: 30 мая для лесного и 31 мая для тундрового (U = = 716.5, z = 0.563, p = 0.573). Таким образом, около половины “летних” птиц добыты в конце мая, другая половина – в течение календарного лета до 24 августа.
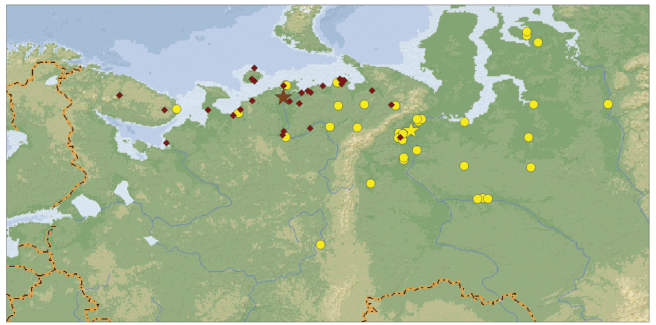
Пространственное распределение находок окольцованных птиц. Распределение летних находок двух подвидов, прямо связанное с различиями в гнездовых частях ареалов, значимо различается (χ2 = 34.019, df = 2, p < 0.0000) и позволяет получить общее представление о районах гнездования и линьки гуменников, зимующих в Нидерландах (рис. 2). Подавляющее большинство встреч тундрового гуменника (средние координаты находок 67°31′ с.ш., 52°36′ в.д.) сосредоточено на территории Ненецкого АО. Отдельные птицы найдены на севере Республики Коми, а также в северных частях Ямало-Ненецкого АО, Мурманской и Архангельской областей. Найденные в пределах северо-таежной подзоны тундровые гуменники, по-видимому, относятся к птицам, еще не достигшим районов размножения. Большая часть находок лесного подвида сосредоточена на территории Ямало-Ненецкого АО и Ханты-Мансийского АО (средние координаты находок 65°54′ с.ш., 65°45′ в.д.). Находки на Гыданском п-ове и на побережьях в пределах тундровой зоны связаны, очевидно, с птицами, уже переместившимися в эти районы на линьку. В европейской части за пределами подзоны северной тайги и зоны тундры имеется лишь один возврат лесного гуменника – с территории Пермского края.
Рис. 2.
Летние возвраты колец лесного гуменника (кружки) и тундрового (ромбы) гуменника. Звездочки – среднее положение находок.
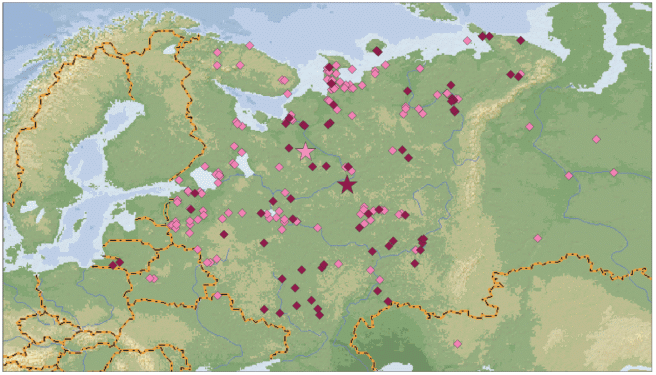
В весенний период имеются существенные различия в положении находок тундровых гуменников, окольцованных по 1989 г. и позднее (χ2 = = 30.86, df = 2, p < 0.0000, рис. 3). В первом случае птиц чаще добывали на территории Северо-Запада и Севера европейской части России (средние координаты находок 62°38′ с.ш., 41°51′ в.д.), тогда как во втором случае больше находок произошло в регионах черноземных областей, Среднего Поволжья и Прикамья (средние координаты находок 60°24′ с.ш., 46°07′ в.д.).
Рис. 3.
Весенние возвраты колец тундрового гуменника с птиц, окольцованных до 1989 г. (малиновые ромбы) и с 1990 г. (вишневые ромбы). Звездочки – среднее положение находок.
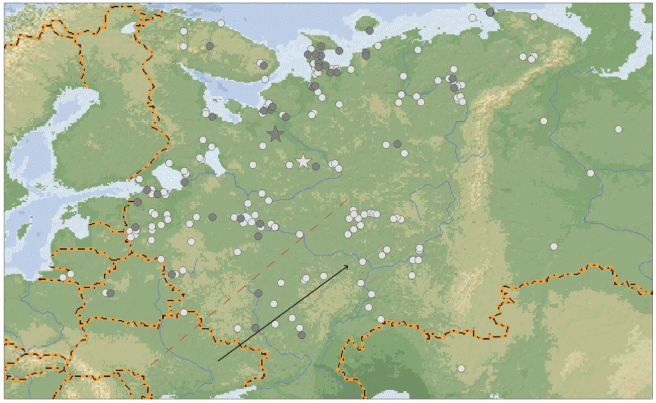
При этом различается также положение прямых и непрямых находок (χ2 = 7.652, df = 2, p < 0.05, рис. 4). В следующую за кольцеванием весну тундровые гуменники были добыты преимущественно в северной и северо-западной части Европейской России (средние координаты находок 61°43′ с.ш., 43°25′ в.д.). С другой стороны, через год и более весенние встречи имели место в среднем чаще в центральных и юго-восточных регионах (средние координаты находок 63°09′ с.ш., 40°51′ в.д.). На территории к юго-востоку от красной пунктирной линии на карте (рис. 4) зарегистрированы 35 из 187 непрямых находок и только 3 из 42 прямых (различия долей на грани достоверности: χ2 = 3.319, df = 2, p = 0.068).
Рис. 4.
Прямые (темные кружки) и непрямые (светлые кружки) весенние возвраты колец тундрового гуменника. Звездочки – среднее положение находок; красная пунктирная линия – граница “юго-восточного пролетного пути”, черная стрелка – его предполагаемое направление.
В отличие от тундрового, у лесного подвида не выявлено различий в положении прямых и непрямых весенних находок (χ2 = 2.622, df = 2, p = = 0.27). Близки друг к другу и средние координаты прямых (62°31′ с.ш., 57°59′ в.д., n = 52) и непрямых встреч (62°08′ с.ш., 56°29′ в.д., n = 94). В целом же весеннее распределение находок двух подвидов ожидаемо значимо различается (p < < 0.0000, рис. 5). В разных регионах европейской части бывшего СССР находки двух подвидов встречаются в сходных пропорциях; тогда как в Западной Сибири резко преобладают находки лесного гуменника (в целом средние координаты 62°16′ с.ш., 57°01′ в.д., n = 146), но также были добыты и отдельные экземпляры тундрового гуменника (в целом средние координаты 62°01′ с.ш., 43°09′ в.д., n = 236). Здесь необходимо отметить, что шесть наиболее южных находок лесного гуменника из европейской части России и две находки с территории Украины относятся к непрямым возвратам колец.
Рис. 5.
Весенние возвраты колец лесного гуменника (кружки) и тундрового гуменника (ромбы). Звездочки – среднее положение находок.
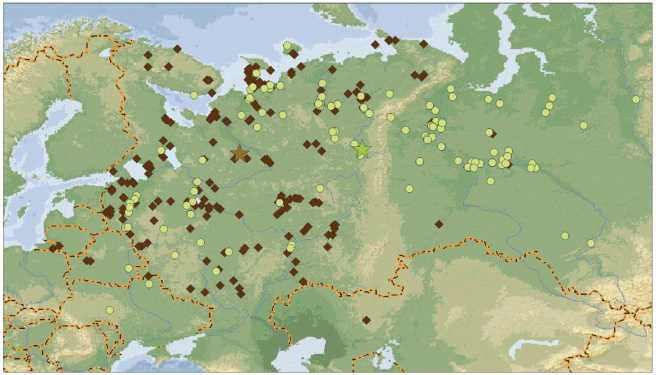
Различия в скорости продвижения весенней миграции двух подвидов ярче всего отражает картина распределения находок в первой декаде мая (рис. 6). В этот период только единичные тундровые гуменники достигают побережий северных морей, а подавляющая часть птиц была добыта в таежной зоне севера европейской части России (средние координаты 61°21′ с.ш., 40°20′ в.д.). В то же время большинство возвратов колец лесных гуменников пришли уже с территории Западной Сибири, из областей гнездования подвида (средние координаты: 62°14′ с.ш., 62°46′ в.д.). Между второй половиной апреля и первой декадой мая имеет место максимальный сдвиг в средних координатах находок лесного гуменника, у тундрового гуменника наибольший сдвиг происходит в мае, между первой и второй декадой месяца (рис. 6). Различия между последующими периодами во всех случаях значимы.
Рис. 6.
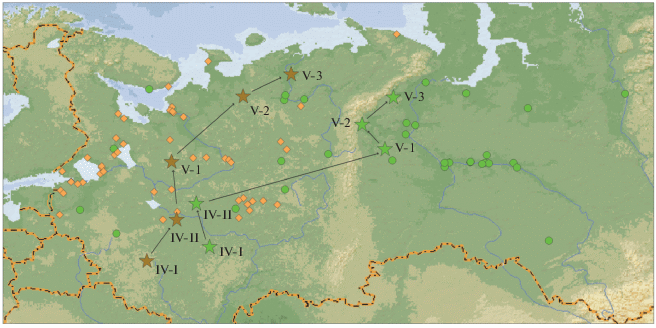
Распределение возвратов колец лесного гуменника (кружки) и тундрового гуменника (ромбы) в первой декаде мая и продвижение средних координат возвратов колец весной по периодам (звездочки). Зеленые звездочки – лесной гуменник: IV-I – первая половина апреля (n = 7), IV-II – вторая половина апреля (n = 19), V-1 – первая декада мая (n = 34), V-2 – вторая декада мая (n = 42), V-3 – третья декада мая (n = 37). Коричневые звездочки – тундровый гуменник: IV-I – первая половина апреля (n = 18), IV-II – вторая половина апреля (n = 45), V-1 – первая декада мая (n = 53), V-2 – вторая декада мая (n = 65), V-3 – третья декада мая (n = 35).
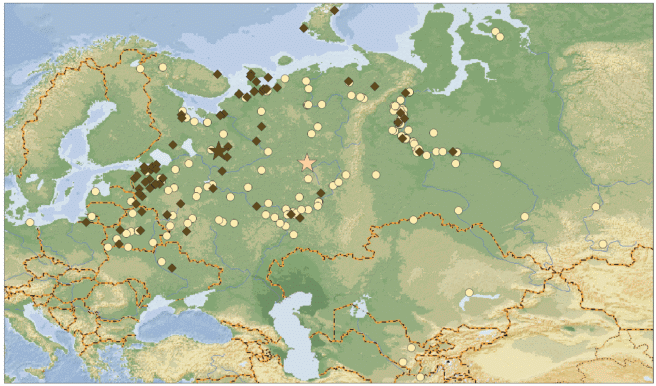
Осенью (рис. 7) большая часть тундровых гуменников продвигается к югу по районам, близким к Беломоро-Балтийскому пролетному пути, а также по верхнему и среднему течению Оби (средние координаты находок 61°42′ с.ш., 39°68′ в.д., n = 73). Большая часть лесных гуменников, в свою очередь, найдена в нижнем и среднем течении Оби, а также в средней полосе европейской части бывшего СССР (средние координаты 60°30′ с.ш., 52°15′ в.д., n = 172). Особенно плотное распределение находок отмечается в Среднем Поволжье и Прикамье. Соответственно различия в осеннем распределении высоко значимы (χ2 = 52.02, df = 2, p < 0.0000). Обращает на себя внимание почти трехкратная разница между подвидами в количестве добытых осенью птиц. Для птиц, окольцованных до 1989 г., имеются сообщения о 65 тундровых и 172 лесных гуменников. От окольцованных после 1989 г. тундровых гуменников пришло всего 8 осенних возвратов.
Рис. 7.
Осенние возвраты колец лесного гуменника (кружки) и тундрового гуменника (ромбы). Звездочки – среднее положение находок.
Местоположение самых восточных находок подвидов (рис. 2, 5, 7) говорит о том, что на нидерландских зимовках могут появляться тундровые гуменники из западносибирских группировок и отдельные лесные гуменники, гнездящиеся к востоку от Енисея. При этом некоторые особи лесного гуменника кардинально меняют регионы зимовок: окольцованные в Нидерландах птицы в последующие годы встречаются в Центральной Азии и на миграционных путях в Восточную Азию (в верхнем течении Оби и Енисея).
ОБСУЖДЕНИЕ
Полученные нами результаты показывают, что добываемые лесные гуменники в целом проживают в среднем меньше, чем тундровые. Значимые различия в сроках дожития получены также при отдельном сравнении самцов обоих подвидов и сравнении особей двух подвидов, добытых в весенний период. Хотя практически во всех остальных случаях сравнения (выборки по возрасту кольцевания, по полу, по сезону добычи) значимых различий не выявлено, средний показатель дожития оказывался неизменно меньше у представителей лесного гуменника. Вероятно, для выявления значимых различий во многих случаях объем выборок оказался недостаточным. Но даже если рассматривать только статистически подтвержденные результаты, общий вывод о большей уязвимости лесного гуменника несомненен.
Можно вполне допускать, что самцы рано гнездящегося лесного гуменника чаще становятся добычей охотников, особенно в весенний период в условиях лесной зоны, когда самцы более заметны, чем сидящие на гнезде самки. И, напротив, результат для летнего сезона – единственный случай, когда среднее дожитие у самок тундрового гуменника оказалось в среднем ниже, чем у лесного (а, в свою очередь, в пределах тундрового подвида добытые самки были в среднем значимо моложе самцов), – можно связывать с большей уязвимостью насиживающих кладку самок в условиях тундры.
Сопоставляя данные о сроках дожития двух подвидов и картину распределения мест их добычи по сезонам, можно сделать следующие обобщения. Меньший срок дожития птиц, добываемых летом, скорее всего, связан с тем, что в этот период для (незаконной) охоты наиболее уязвимы молодые (не размножающиеся птицы), многие из которых раньше отлетают в тундру к местам линьки и становятся более заметными. При этом медианный год добычи тундрового гуменника для летнего сезона наиболее ранний, в сравнении с медианными годами для других сезонов. Это может указывать на то, что в последние десятилетия летняя (незаконная) охота имеет место реже (по крайней мере, в районах распространения этого подвида) и/или в случае добычи в неустановленные сроки охотники реже сообщают о находке.
Учитывая сходство в биологии двух подвидов и единство видовой стратегии, маловероятно, что различия в сроках дожития определяются биологическими факторами, и, соответственно, именно пресс охоты определяет специфику полученных нами результатов. Комбинация особенностей фенологии этапов годового цикла и сезонного распределения птиц, с одной стороны, и традиционных сроков открытия охоты – с другой, создает менее благоприятные условия существования для лесного гуменника. Этот подвид раньше пролетает по районам с наибольшей плотностью охотников весной и начинает гнездиться зачастую задолго до закрытия весенней охоты (Головатин, 2010; наши данные). Это подтверждается и данными от помеченных передатчиками птиц, а также данными авиа- и наземных учетов (Розенфельд и др., 2018; Артемьев и др., 2019; Розенфельд, Стрельников, 2019; Розенфельд и др., 2020; Heldbjerg et al., 2020). На большей части областей гнездования лесного гуменника сроки весенней охоты приходятся на время насиживания кладки. Таким образом, под выстрел попадают в основном представители местных гнездящихся популяций.
Пролетные пути лесного гуменника в целом проходят по более населенным территориям. Особенно хорошо это заметно на карте распределения осенних находок окольцованных птиц (рис. 7). Согласно нашим данным и имеющимся публикациям (Лебедева, 1979; Кищинский, 1979), осенью тундровый гуменник продвигается к европейским зимовкам вдоль побережий морей Северного Ледовитого океана (где интенсивность осенней охоты относительно невысока), а затем большая часть птиц использует Беломоро-Балтийский пролетный путь. Большая часть лесных гуменников, в свою очередь, осенью пролетает по центральным областям европейской части России, где чаще становится добычей охотников. Возможно, подвидовые пропорции добытых осенью птиц (почти трехкратное преобладание находок лесного подвида) объективно отражают соотношение охотничьего пресса на два рассматриваемых таксона. В весенний период большая часть лесных гуменников к началу охоты успевает достигнуть малонаселенных областей Западной Сибири, и относительная вероятность их отстрела уменьшается. При этом в весенний период приступившие к размножению лесные гуменники оказываются уязвимы и подвержены отстрелу даже в Западной Сибири, что подтверждается обилием возвратов колец из этих малонаселенных районов. Сдвиг охотничьей активности на весенний период в последние десятилетия, вероятно, имеет общий характер и может усиливать нагрузку на лесной подвид.
Многолетний сдвиг средних координат весенних находок тундрового гуменника (рис. 3) может быть обусловлен как сменой направления миграции, так и изменениями в интенсивности охотничьей нагрузки. Пока у нас недостаточно данных, чтобы предпринимать попытки как-то разделить эти факторы. Однако выявленные различия в распределении прямых и непрямых встреч тундрового подвида (рис. 4) определяются скорее природными, нежели социальными факторами. Эти данные позволяют предполагать, что гуменники год от года могут менять регионы зимовок. В первую после кольцевания весну пролетные пути птиц, окольцованных на западноевропейских зимовках, в среднем проходят значимо северо-западнее, чем в последующие годы. Различия в пролетных путях дают основание предполагать, что через год и более те же тундровые гуменники могут улететь на зимовки в Южную и Восточную Европу, возможно – на Средний и Нижний Дунай. О зимнем распределении тундровых гуменников писал и Кищинский (1979). По его предположению, гуменники, гнездящиеся к востоку от устья Печоры, практически не встречаются на голландских зимовках, а зимуют “в бассейне Дуная, в ЧССР, Австрии, Венгрии и Югославии, где гуменники многочисленны”. Здесь же говорится об увеличении обмена между европейскими зимовками.
Теперь стало известно, что, по данным наблюдений тундровых гуменников, помеченных ошейниками в Ненецком АО на о-ве Вайгач в 1996–1997 г., часть зимних регистраций действительно относится к территории Чехии, Словакии и Венгрии. Есть птицы, которые в разные годы встречались то в западной части Германии, то в Венгрии. Не исключено, что отдельные птицы перемещаются между районами зимовок в течение одной зимы (База данных Центра кольцевания птиц ИПЭЭ РАН). Именно этим можно объяснить наличие трех прямых находок тундрового подвида в юго-восточном регионе (ограничен красным пунктиром на рис. 4). В целом, если выявленные тенденции определяются в основном природными факторами, то можно говорить о том, что в последние 3–4 десятилетия все больше гуменников пользуются юго-восточными пролетными путями, возможно возвращаясь с восточноевропейских зимовок. Смена европейских зимовок может касаться, в первую очередь, тундрового подвида, но может также быть одной из причин исчезновения лесного гуменника с нидерландских зимовок, связанной, возможно, также с климатическими изменениями. Однако пока все это остается в виде предположения, для подтверждения которого необходим сбор дополнительного материала, в том числе, при помощи доступного теперь спутникового слежения. С другой стороны, только новые данные массового кольцевания позволят получить материал для статистической оценки современных параметров выживаемости и влияния пресса охоты на различные виды и подвиды гусей.
ЗАКЛЮЧЕНИЕ
По данным возвратов колец с добытых птиц, лесной гуменник в среднем проживает меньше, чем его тундровый конспецифик, по-видимому, испытывая бóльший пресс охоты. В период весенней охоты лесной подвид гуменника наиболее уязвим в районах Западной Сибири, осенью – в средней полосе европейской части России, начиная от Урала. Все это указывает на необходимость пересмотра сроков весенней охоты и ее ограничения, а также на необходимость принятия дополнительных мер по охране лесного гуменника в ключевых регионах размножения и сезонного распределения. При этом важен региональный подход к включению подвидовых таксонов в Красные книги. Необходимо провести дополнительные исследования для уточнения сведений о пролетных путях и зимнем распределении гусеобразных, в том числе, для проверки гипотезы о смене районов зимовки гуменников, гнездящихся на территории Российской Федерации.
Список литературы
Артемьев А.В., Матанцева М.В., Симонов С.А., 2019. Лесной гуменник на весенней миграционной стоянке гусей и казарок в окрестностях г. Олонца, Республика Карелия, Россия // Казарка. Т. 21. С. 81–90.
Головатин М.Г., 2010. Таежный гуменник // Красная книга ЯНАО. Екатеринбург: Изд-во Баско. С. 212.
Кищинский А.А., 1979. Миграция гуменника по данным кольцевания, полученным в СССР. Общие заключения // Миграции птиц Европы и Северной Азии. Аистообразные – Пластинчатоклювые. М.: Наука. С. 160–163.
Лебедева М.И., 1979. Миграция гуменника по данным кольцевания, полученным в СССР // Миграции птиц Европы и Северной Азии. Аистообразные – Пластинчатоклювые. М.: Наука. С. 150–160.
Литвин К.Е., 2014. Новые данные о миграциях гусей, гнездящихся в России. Обзор результатов дистанционного прослеживания // Казарка. Т. 17. С. 13–45.
Паевский В.А., 2008. Демографическая структура и популяционная динамика певчих птиц. М.–СПб.: Товарищество научных изданий КМК. 235 с.
Приказ Министерства природных ресурсов и экологии Российской Федерации, 2020. Приказ Министерства природных ресурсов и экологии Российской Федерации от 24.03.2020 № 162 “Об утверждении Перечня объектов животного мира, занесенных в Красную книгу Российской Федерации” (Зарегистрирован 02.04.2020 № 57 940).
Розенфельд С.Б., Замятин Д.О., Ванжелюв Д., Киртаев Г.В., Рогова Н.В., Као Л., Поповкина А.Б., 2018. Лесной гуменник в Ямало-Ненецком автономном округе // Казарка. Т. 20. С. 28–55.
Розенфельд С.Б., Стрельников Е.Г., 2019. Новые сведения о зимовке лесного гуменника Anser fabalis fabalis, гнездящегося в заповеднике “Юганский” // Русский орнитологический журнал. Т. 28. № 1858. С. 5683–5687.
Розенфельд С.Б., Баянов Е.С., Богомякова Н.Г., Стрельников Е.Г., 2020. О значении миграционной остановки лесного гуменника Anser fabalis fabalis в Свердловской области, выявленном с помощью гусыни, помеченной GPRS передатчиком // Русский орнитологический журнал. Т. 29. № 1927. С. 2333–2337.
Спицын В.М., 2019. Встреча лесного гуменника на Южном острове Новой Земли // Казарка. Т. 20. С. 125–126.
Харитонов С.П., 2017. Оценка ежегодной смертности и демографического благополучия популяций птиц по данным кольцевания // Зоологический журнал. Т. 96. № 3. С. 350–359.
Arizaga J., Crespo A., Telletxea I., Ibanez R., Dıez Ph., Tobar J.F., Minondo M., Ibarrola Z., Fuente J.J., Perez J.A., 2014. Solar/Argos PTTs contradict ring-recovery analyses: Woodcocks wintering in Spain are found to breed further east than previously stated // Journal of Ornithology. V. 156. P. 515–523.
Batchelet E., 1972. Recentstatistical methods for orientation data. Animal Orientation and Navigation, NASA SP-262. P. 61–91.
Burgers J., Smit J.J., Voet van der H., 1991. Origins and systematics of 2 types of the Bean Goose Anser fabalis (Latham, 1787) wintering in The Netherlands // Ardea. V. 79. № 2. P. 307–315.
CAFF, 2018. A Global audit of the status and trends of Arctic and Northern Hemisphere goose population (Component 2: population accounts) // Conservation of Arctic Flora and Fauna International Secretariat, Akureyri, Iceland. 173 p.
Heldbjerg H., Fox A.D., Christensen T.K., Clausen P., Kampe-Persson H., Koffijberg K., Kostiushyn V., Liljebäck N., Mitchell C., Nilsson L., Rozenfeld S., Skyllberg U., Alhainen M., 2020. Taiga bean goose population status report 2019-2020 // AEWA European goose management platform (EGMP) data centre. 18 p.
Hornman M., Hustings F., Koffijberg K., Klaassen O., Kleefstra R., van Winden E., Soldaat L., 2013. Watervogels in Nederland in 2011/2012 // Sovon report 2013/66, Sovon Vogelonderzoek, Nijmegen. 34 p.
Klaassen M., Bauer S., Madsen J., Tombre I., 2005. Modelling behavioural and fitness conse-quences of disturbance for geese along their spring flyway // Journal of Applied Ecology. V. 43. P. 90–100.
Koffijberg K., Hustings F., de Jong A., Hornman M., van Winden E., 2011. Recente ontwikkelingen in het voorkomen van Taigarietganzen in Nederland // Limosa. V. 84. P. 117–131.
Ladin Z. S., Costanzo G., Lewis B., Williams C., 2020. Long term survival and harvest of resident Canada Geese in Virginia // The Journal of Wildlife Management. V. 84. P. 666–674.
Maihguy J., Bety J., Gauthier G., Giroux J.F., 2002. Are body condition and reproductive effort of laying Greater Snow Geese affected by the spring hunt? // Condor. V. 104. № 1. P. 156–161.
Mardia K.V., 1967. A nonparametric test for the bivariate two-sample location problem // Journal of the Royal Statistical Society. Ser. B 29. P. 320–342.
Marjakangas A., Alhainen M., Fox A.D., Heinicke T., Madsen J., Nilsson L., Rozenfeld S., 2015. International Single Species Action Plan for the Conservation of the Taiga Bean Goose Anser fabalis fabalis // AEWA Technical Series. V. 56. Bonn, Germany. 88 p.
Nilsson L., 2011. The migrations of Finnish Bean Geese Anser fabalis in 1978–2011 // Ornis Svecica. V. 21. P. 157–166.
Nilsson L., van den Bergh L., Madsen J., 1999. Taiga Bean Goose Anser fabalis fabalis // Goose populations of the Western Palearctic. A review of status and distribution. Wetlands International Publ. V. 48. P. 20–36.
Nilsson L., de Jong A., Heinicke T., Sjöberg K., 2010. Satellite tracking of Bean Geese Anser fabalis fabalis and A. f. rossicus from spring staging areas in Northern Sweden to breeding and moulting areas // Ornis Svecica. V. 20. P. 184–189.
Robson D.S., 1968. The effect of ties on critical values of some two-sample rank tests // Biometrics Unit, Cornell. Univ. BU-258-M. P. 1–4.
Дополнительные материалы отсутствуют.
Инструменты
Зоологический журнал